

Abstract
Objective
Bexarotene (Targretin®) is a clinically used anti-tumoral agent which exerts its action through binding to and activation of the Retinoid-X-Receptor (RXR). The most frequent side effect of bexarotene administration is an increase in plasma triglycerides, an independent risk factor of cardiovascular disease. The molecular mechanism behind this hypertriglyceridemia remains poorly understood.
Methods and Results
Using wild-type and LXRα/β-deficient mice, we show here that bexarotene induces hypertriglyceridemia and activates hepatic LXR-target genes of lipogenesis in an LXR-dependent manner, hence exerting a permissive effect on RXR/LXR heterodimers. Interestingly, RNA analysis and Chromatin Immunoprecipitation assays performed in the liver reveal that the in vivo permissive effect of bexarotene on the RXR/LXR heterodimer is restricted to lipogenic genes without modulation of genes controlling cholesterol homeostasis.
Conclusion
These findings demonstrate that the hypertriglyceridemic action of bexarotene occurs via the RXR/LXR heterodimer and show that RXR heterodimers can act with a selective permissivity on target genes of specific metabolic pathways in the liver.
Keywords: Animals; Cholesterol; metabolism; DNA-Binding Proteins; chemistry; physiology; Dimerization; Female; Homeostasis; Lipogenesis; Liver; metabolism; Mice; Mice, Inbred C57BL; Orphan Nuclear Receptors; Receptors, Cytoplasmic and Nuclear; chemistry; physiology; Retinoid X Receptors; chemistry; physiology; Tetrahydronaphthalenes; pharmacology; Triglycerides; metabolism
Keywords: Retinoid-X-Receptor, rexinoid, hypertriglyceridemia, Liver-X-Receptor, murine model, ChIP
INTRODUCTION
RXRs (Retinoid-X-Receptors) are ligand-activated transcription factors belonging to the nuclear receptor superfamily. Three subtypes (RXRα, RXRβ and RXRγ) have been identified, which can be activated by natural (9-cis retinoic acid, docosahexaenoic acid) or synthetic ligands, called rexinoids1. RXRs are regulators of a plethora of pathophysiological processes with potential clinical implications. For instance, bexarotene (LGD1069, Targretin®), a rexinoid with potent and selective activity for RXRα, β and γ2, is clinically used in the treatment of cutaneous T cell lymphoma3 and has promising effects in other cancer forms or dermatological disorders like psoriasis4. In addition, synthetic RXR ligands exert beneficial glucose-lowering and insulin-sensitizing effects as well as anti-obesity actions in animal models of insulin-resistance and diabetes5, and bexarotene improves cholesterol homeostasis and inhibits the development of atherosclerosis in a mouse model of mixed dyslipidemia6. However, similar to other rexinoids, bexarotene administration induces an undesirable increase in plasma triglycerides7, 8, an independent risk factor of cardiovascular disease (CVD)9, 10. Until now, the molecular mechanism behind bexarotene-induced hypertriglyceridemia remains unknown.
RXR is a unique nuclear receptor since it can modulate gene transcription by forming either homodimers or heterodimers with several other nuclear receptors, such as the PPARs (Peroxisome Proliferator-Activated Receptors), LXRs (Liver-X-Receptors), FXR (Farnesoid-X-Receptor), PXR (Pregnane-X-Receptor), RARs (Retinoic-Acid-Receptors), VDR (Vitamin D-Receptor) and TR (Thyroid Hormone Receptor). These heterodimers have been further subclassified as permissive or non-permissive heterodimers. This concept states that permissive heterodimers formed by RXR and binding partners are activatable by both RXR-specific and partner specific ligands, whereas non-permissive heterodimers can only be activated by ligands specific for the binding partner. Among the nuclear receptors which require RXR as an obligate partner for their action and form permissive heterodimers are the PPARs, FXRs and LXRs. As PPARs11, FXRs12 and LXRs13 are involved in the regulation of genes implicated in triglyceride metabolism, permissive ligand activation of RXR by rexinoids could elicit responses of the dimerization partner and as such modulate triglyceridemia. However, whereas PPARα and FXR activation leads to decreased plasma triglyceride levels, LXR activation increases triglycerides13, presenting the permissive RXR/LXR heterodimer as a potential mediator of bexarotene-induced hypertriglyceridemia.
LXRs (LXRα and LXRβ) are nuclear receptors that are activated by oxysterols or synthetic ligands such as T0901317, and bind DNA as obligate heterodimers with RXR at specific LXR response elements (LXREs) composed of a direct repeat of 5′-AGGTCA-3′ spaced by four bases. LXR activation regulates triglyceride metabolism and induces hepatic steatosis and hypertriglyceridemia due, in part, to increased hepatic fatty acid synthesis and VLDL secretion13, 14 by upregulation of both sterol regulatory element-binding protein 1c (SREBP1c)13, 15, 16 and carbohydrate response element-binding protein (ChREBP)17 expression. Moreover, LXRs play an important role in the regulation of cholesterol homeostasis18, 19 and their activation leads to the induction of several genes implicated in reverse cholesterol transport and mobilization of cholesterol, such as the ATP binding cassette (ABC) transporters ABCA1, ABCG1, ABCG5, ABCG8 and apolipoprotein E20.
Using wild-type mice and mice deficient for both isoforms of LXR (LXRα/β-deficient mice), we show that the permissive heterodimer RXR/LXR mediates bexarotene-induced hypertriglyceridemia and hepatic lipogenesis in vivo. Surprisingly, RNA levels and Chromatin Immunoprecipitation (ChIP) assays reveal that this LXR-mediated effect of bexarotene in the liver is restricted to genes implicated in triglyceride homeostasis and lipogenesis, without modulation of genes controlling cholesterol homeostasis. These findings demonstrate that RXR heterodimers can act with a selective permissivity on target genes of specific metabolic pathways.
RESULTS
Bexarotene increases plasma and hepatic triglyceride levels in an LXRα/β-dependent manner
To determine whether LXRs are implicated in the bexarotene-induced hypertriglyceridemia, wild type mice and mice lacking both LXRα and LXRβ (LXRα/β-deficient mice) were treated with the RXR agonist (bexarotene, 30 and 100 mpk), the LXR agonist (T0901317, 30 mpk) or vehicle alone for 14 days (figure 1A). In wild type mice, the LXR agonist induced a hypertriglyceridemia, an effect which was not observed in LXRα/β-deficient mice. Interestingly, the RXR agonist bexarotene also induced a robust dose-dependent hypertriglyceridemia, which was strongly blunted in LXRα/β-deficient mice. Thus, the RXR/LXR heterodimer appears to be the principal mediator of the bexarotene-induced hypertriglyceridemia.
Figure 1. RXR or LXR agonist treatment increases plasma and hepatic triglyceride levels in vivo in an LXRα/β-dependent manner.
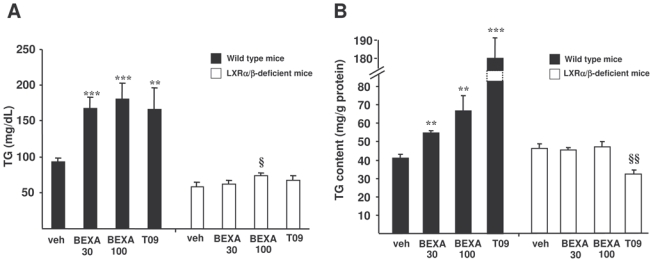
Plasma (A) and hepatic (B) triglyceride concentrations were measured in LXRα/β-deficient mice (□) and wild type mice (■) gavaged with the RXR agonist (bexarotene, 30 and 100 mpk), the LXR agonist (T0901317, 30 mpk) or vehicle alone for 14 days (N=7/group). **p<0.01, ***p<0.001 versus wild type mice treated with vehicle; §p<0.05, §§p<0.01 versus LXRα/β-deficient mice treated with vehicle.
Since LXR is a lipogenic transcription factor in the liver, we analyzed whether bexarotene influences triglyceride metabolism in the liver and whether these effects are mediated by LXR (figure 1B). As previously shown13, the LXR agonist T0901317 increased hepatic triglyceride levels in wild type, but not in LXRα/β-deficient mice. Interestingly, bexarotene induced a dose-dependent increase in hepatic triglyceride concentration in wild type but not in LXRα/β-deficient mice. Thus, the bexarotene-induced hypertriglyceridemia and hepatic triglyceride accumulation are dependent on the RXR/LXR pathway.
Bexarotene induces hepatic lipogenic genes in an LXRα/β-dependent manner
To further explore the role of LXR in the effects of bexarotene on hepatic triglyceride metabolism, the expression of LXR-target genes implicated in lipogenesis (SREBP1c, FAS and SCD1) was measured by quantitative PCR (figure 2). As expected, LXR agonist administration robustly increased the expression of SREBP1c, FAS and SCD1 in wild type mice, an effect which was completely blunted in LXRα/β-deficient mice. Interestingly, the mRNA levels of SREBP1c, FAS, and SCD1 also displayed a strong dose-dependent increase upon bexarotene treatment in wild type, but not in LXRα/β-deficient mice. To determine whether bexarotene could act via other transcription factor pathways and nuclear receptor heterodimers, RNA levels of ChREBP, a glucose-activated lipogenic transcription factor, acylcoA oxidase (ACO, PPARα target gene) and BSEP (FXR target gene) (supplemental figure 1A) as well as Cyp3A11, CD36 and FAE (PXR target genes) (supplemental figure 1B) were measured. Bexarotene treatment increased ChREBP, ACO and BSEP mRNA both in wild type and in LXR-deficient mice (supplemental figure 1A), suggesting that ChREBP gene regulation as well as the permissive heterodimers RXR/PPAR and RXR/FXR do not participate in bexarotene-induced lipogenesis which occurs only in wild type mice. Moreover, we measured the expression of 5′deiodinase type 1 (DIO1), a RXR/TR target gene implicated in the metabolism of thyroid hormones (supplemental figure 1A). Bexarotene treatment decreased DIO1 expression in the liver to a similar extent in wild type and in LXRα/β-deficient mice, suggesting that potential alterations in the thyroid axis do not contribute to bexarotene-induced hypertriglyceridemia. Finally, Cyp3A11 was not modified in bexarotene-treated wild type mice, indicating that PXR does not appear to be implicated in bexarotene-induced lipogenesis (supplemental figure 1B). By contrast to Cyp3A11, CD36 and FAE mRNA levels were strongly increased in T091317-treated wild type but not LXR-deficient mice, suggesting that these genes are activated in a LXR- but not PXR-dependent manner by T091317 (supplemental figure 1B). CD36 expression was induced by bexarotene treatment both in wild type and LXR-deficient mice, suggesting that CD36 is not implicated in bexarotene-induced lipogenesis which is only observed in wild type mice. FAE expression was strongly increased in wild type bexarotene-treated mice, an effect which was strongly reduced in LXR-deficient mice, suggesting that the lipogenic enzyme FAE could be implicated in bexarotene-induced lipogenesis, similar as SREBP-1c, SCD1 and FAS.
Figure 2. RXR or LXR agonist treatment induces hepatic lipogenic gene expression in mice in vivo in an LXRα/β-dependent manner.
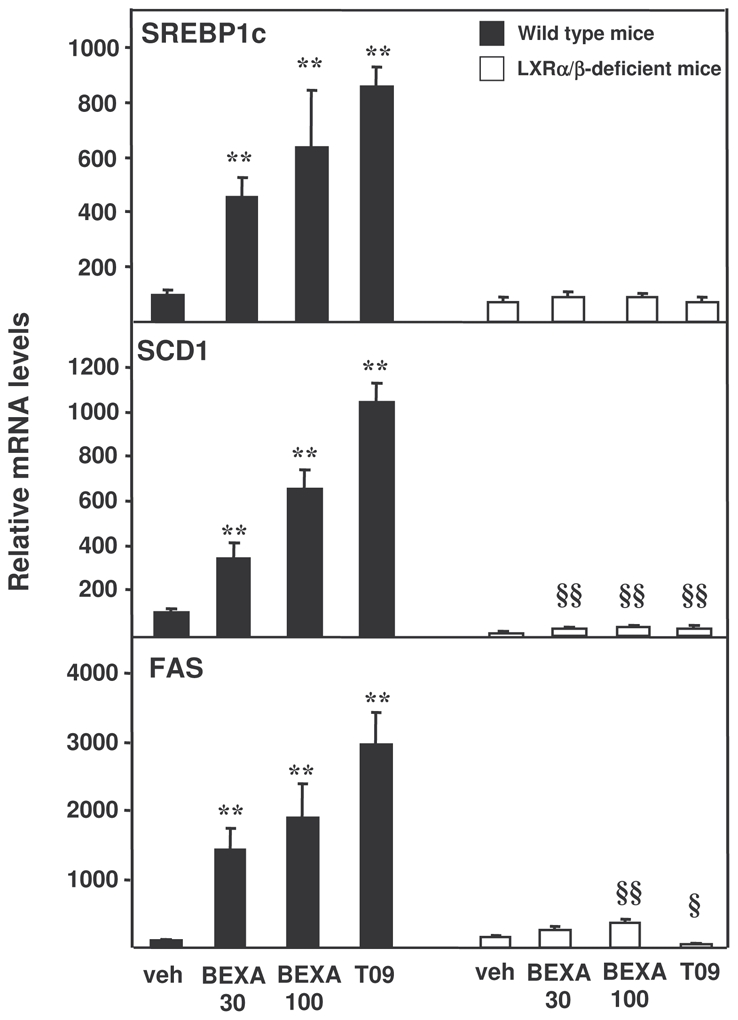
Hepatic mRNA levels of SREBP1c, SCD1 and FAS were measured in LXRα/β-deficient mice (□) and wild type mice (■) gavaged with the RXR agonist (bexarotene, 30 and 100 mpk), the LXR agonist (T0901317, 30 mpk) or vehicle alone for 14 days (N=7/group). Data normalized to 28S mRNA are expressed as a percentage of the wild type mice group treated with vehicle alone, to which an arbitrary value of 100 was attributed. **p<0.01 versus wild type mice treated with vehicle, §p<0.05, §§p<0.01 versus LXRα/β-deficient mice treated with vehicle.
These data show that bexarotene upregulates specific hepatic LXR-target lipogenic genes via the permissive heterodimer RXR/LXR in vivo.
Bexarotene increases RXR binding to the LXREs of hepatic lipogenic genes
To investigate whether the activation of hepatic lipogenic LXR-target genes was associated with an increased ability of RXR complexes to bind LXREs in the promoters of these genes, ChIP assays were performed on livers of mice treated with the highest dose of the RXR agonist, the LXR agonist or vehicle alone (figure 3). Antibodies directed against RXR (figure 3A) and RNA polymerase II (figure 3B) were used, and input as well as immunoprecipitated DNA was quantified by real-time PCR using primers positioned around the LXREs of these genes (figure 3A) or within the gene (figure 3B), respectively. The antibody against RNA polymerase II recognizes both the non-phosphorylated and the phosphorylated extending forms of RNA polymerase II, and occupancy within the gene can be used as a measure of transcriptional activity21. In wild type mice, the RXR ChIP-PCR revealed that both LXR and RXR agonist treatment increased the recruitment of RXR on the LXREs of SREBP1c, SCD1 and FAS genes (figure 3A). RNA polymerase II ChIP-PCR showed that this bexarotene and T0901317-induced RXR recruitment was associated with increased RNA polymerase II occupancy in the genes encoding SREBP1c, FAS and SCD1 in wild type mice (figure 3B). By contrast, bexarotene-induced RXR recruitment to the FAS and SREBP1c LXREs was abolished in the LXRα/β-deficient mice, upon bexarotene and T0901317 treatment, whereas a small, albeit significant, increase was still observed on the SCD1 LXRE (figure 3A). These data suggest that RXR can bind independent of LXR in the vicinity of this LXRE. Moreover, in LXRα/β-deficient mice, neither bexarotene nor T0901317 lead to significant increase of RNA polymerase II occupancy on the SREBP1c and FAS genes. In keeping with the modest LXR independent binding of RXR to the SCD1 gene, a modest bexarotene-induced recruitment of RNA polymerase II was observed after bexarotene treatment. Altogether, both LXR and RXR agonist treatment increased RXR binding to LXREs and increased RNA polymerase II occupancy on hepatic lipogenic genes in wild type mice but not in LXRα/β-deficient mice.
Figure 3. RXR or LXR agonist treatment increases RXR recruitment to the LXREs of lipogenic genes in vivo.
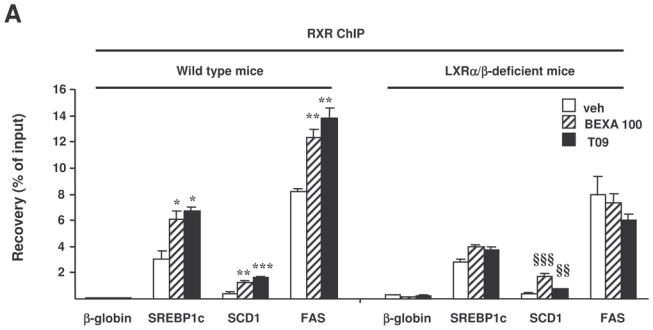
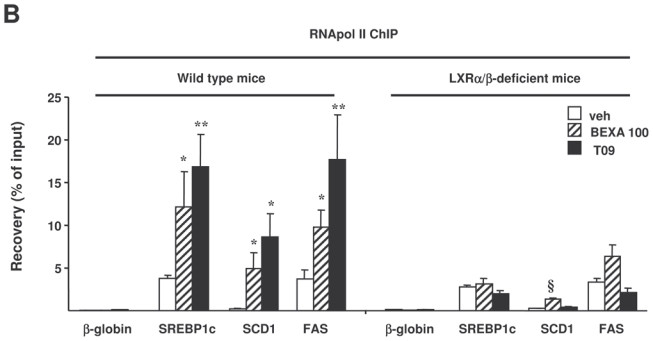
ChIP assays were performed using antibodies against RXR (A) or RNA polymerase II (B) on livers of LXRα/β-deficient mice and wild type mice treated with vehicle alone, bexarotene (100 mpk) or T0901317 (30 mpk). Recovery was determined by real-time PCR using primers positioned at the LXRE of SREBP1c, FAS and SCD1 (A) or primers located in the body of the gene (B). β-globin was used as a control. *p<0.05, **p<0.01, ***p<0.001 versus wild type mice treated with vehicle, §p<0.05, §§p<0.01, §§§p<0.001 versus LXRα/β-deficient mice treated with vehicle.
Plasma cholesterol concentrations are increased after LXR but not RXR agonist treatment
Because LXR is a major regulator of cholesterol homeostasis, plasma cholesterol concentrations were also measured in wild type and LXRα/β-deficient mice treated with the LXR and RXR agonists, respectively (figure 4A). The LXR agonist T0901317 increased plasma total cholesterol by increasing both HDL-C and non-HDL-C in wild type, but not in LXRα/β-deficient mice (figure 4B and 4C). Hepatic cholesterol content was not modified after T0901317 treatment (data not shown). Interestingly, plasma total cholesterol, HDL-C and non-HDL-C did not change upon treatment of wild type and LXRα/β-deficient mice with the RXR agonist bexarotene. Moreover, bexarotene treatment did not influence hepatic cholesterol content (data not shown). Thus, bexarotene-mediated RXR activation of the RXR/LXR heterodimer does not influence plasma cholesterol concentrations, whereas T0901317-induced LXR activation of RXR/LXR increases plasma cholesterol concentrations.
Figure 4. LXR, but not RXR agonist treatment increases plasma cholesterol levels in vivo.
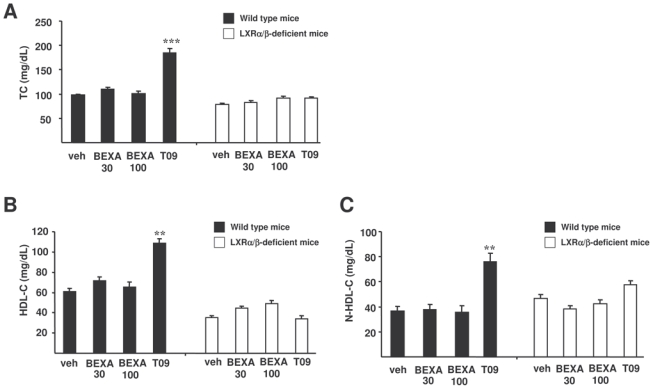
TC (A), HDL-C (B) and non-HDL-C (C) were measured in LXRα/β-deficient mice (□) and wild type mice (■) gavaged, as indicated, with the RXR agonist (bexarotene, 30 and 100 mpk), the LXR agonist (T0901317, 30 mpk) or vehicle alone for 14 days (N=7/group). **p<0.01, ***p<0.001 versus wild type mice treated with vehicle.
Hepatic LXR-target genes implicated in cholesterol homeostasis are induced by LXR but not RXR agonist treatment
LXR influences cholesterol homeostasis in the liver by regulating genes encoding proteins of the ATP Binding Cassette Transporter family22, 23. Therefore, the expressions of these LXR-target genes were measured in the liver. In wild type mice, LXR agonist treatment induced a strong increase in the expression of ABCG1, ABCA1, ABCG5 and ABCG8, an effect which was totally absent in LXRα/β-deficient mice (figure 5). Notably, in contrast to lipogenic LXR target genes, the hepatic mRNA levels of these LXR target genes of cholesterol metabolism only displayed minor or no responsiveness to RXR agonist treatment. Indeed, bexarotene treatment only slightly influenced the mRNA level of these genes and these effects were comparable between wild type and LXRα/β-deficient mice. These data show that, in contrast to hepatic lipogenic genes, only T0901317 induces hepatic LXR-target genes implicated in cholesterol homeostasis via RXR/LXR in vivo, demonstrating that RXR/LXR heterodimers can act with a selective permissivity on target genes of specific pathways in the liver. To analyze whether bexarotene can modulate these LXR-target genes of cholesterol homeostasis in another organ such as the intestine, the expression of ABCG5 and ABCG8 as well as NPC1L1 was measured in the jejunum. In contrast to T091317, the bexarotene-induced decrease of NPC1L1 expression was not LXR-dependent. However, both bexarotene and T091317 increased intestinal ABCG5 and ABCG8 mRNA levels in an LXR-dependent manner (supplemental figure 2). These LXR-dependent increases of ABCG5 and ABCG8 expression by bexarotene in the intestine differ from the absence of LXR-dependent effects of bexarotene on these genes in the liver, identifying tissue-specific mechanisms of RXR/LXR heterodimer permissivity.
Figure 5. LXR, but not RXR agonist treatment upregulates hepatic cholesterol genes in vivo.
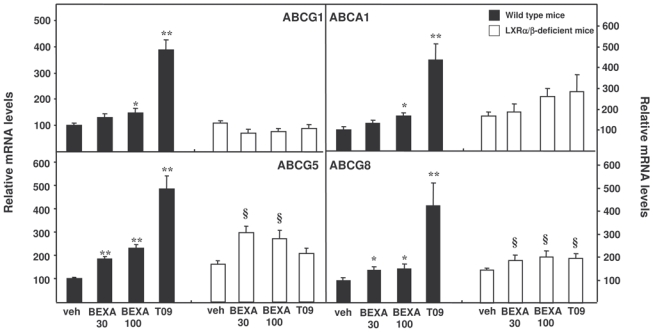
Hepatic mRNA levels of ABCG1, ABCA1, ABCG5 and ABCG8 were measured in LXRα/β-deficient mice (□) and wild type mice (■) gavaged, as indicated, with the RXR agonist (bexarotene, 30 and 100 mpk), the LXR agonist (T0901317, 30 mpk) or vehicle alone for 14 days (N=7/group). Data normalized to 28S gene are expressed as a percentage of the wild type mice group treated with vehicle alone, to which an arbitrary value of 100 was attributed. * p<0.05, **p<0.01 versus wild type mice treated with vehicle, §p<0.05 versus LXRα/β-deficient mice treated with vehicle.
RXR recruitment on the LXREs of hepatic genes implicated in cholesterol homeostasis is induced by LXR but not RXR agonist treatment
To determine how T0901317 and bexarotene affect RXR recruitment to the LXREs of genes implicated in cholesterol homeostasis, RXR ChIP-PCR was performed on livers of mice treated with the highest dose of the RXR agonist, the LXR agonist or vehicle alone (figure 6A). LXR agonist treatment increased RXR occupancy on a putative LXRE sequence adjacent to the ABCG1 gene in wild type mice but not in LXRα/β-deficient mice. Even though ABCA1 has a high basal LXRE occupancy by RXR, RXR recruitment on the reported LXRE in the proximal promoter of ABCA1 also tended to increase in wild type mice but not in LXRα/β-deficient mice. Since the mouse LXREs in ABCG5 and ABCG8 genes have not yet been characterized, we used computational analysis to identify the DR4 elements in the vicinity of these genes and we identified two putative LXREs in the first intron of the ABCG8 gene. Interestingly, LXR agonist treatment increased the recruitment of RXR to these putative LXREs only in wild type mice, indicating that these sequences are indeed functional LXREs. In keeping with ABCG1, ABCA1, ABCG5 and ABCG8 being LXR target genes in the liver, RNA polymerase II ChIP-PCR demonstrated LXR agonist dependent polymerase II occupancy in wild type but not in LXRα/β-deficient livers (figure 6B). Interestingly, in contrast to the LXR agonist, RXR agonist did not induce major changes in RXR binding to the examined LXREs adjacent to genes involved in cholesterol homeostasis neither in wild type nor in LXRα/β-deficient liver. Moreover, recruitment of RNA polymerase II to the ABCG1, ABCA1, ABCG5 and ABCG8 genes was not influenced by bexarotene in wild type and in LXRα/β-deficient mice (figure 6B). These data demonstrate that LXR and RXR agonists differently regulate RXR binding on hepatic genes implicated in cholesterol homeostasis, an effect that may contribute to their distinct effects on plasma and hepatic lipid levels.
Figure 6. LXR, but not RXR agonist treatment increases RXR recruitment to the LXRE of cholesterol genes in vivo.
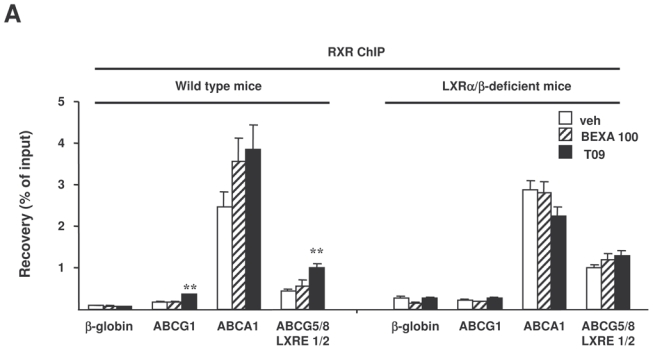
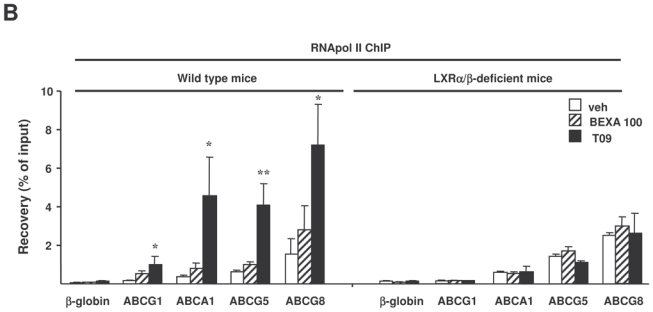
ChIP assays were performed using antibodies against RXR (A) or RNA polymerase II (B) on livers of LXRα/β-deficient mice and wild type mice treated with vehicle alone, bexarotene (100 mpk) or T0901317 (30 mpk). Recovery was determined by real-time PCR using primers positioned at the LXRE of ABCA1, ABCG1 and at two putative LXREs of ABCG5 and ABCG8 (A) or primers located in the body of the gene (B). β-globin was used as a control. *p<0.05, **p<0.01 versus wild type mice treated with vehicle.
DISCUSSION
In the present study, we show that bexarotene, a rexinoid used clinically for the treatment of certain cancers and dermatological disorders, induces hypertriglyceridemia in an LXR-dependent manner in vivo. This observation is of particular importance since rexinoids have been proposed for the treatment of metabolic diseases and insulin-resistance syndromes. However, side effects such as hypertriglyceridemia24, 25 which is now widely recognized to be an independent risk factor of CVD9, have limited their use. Until now, the mechanism underlying this effect was not understood. In a study performed in ZDF rats treated for 14 days with a broad range of bexarotene doses, the authors proposed that the bexarotene-induced dose-dependent hypertriglyceridemia was due to elevated VLDL caused by a primary defect in LPL activity mainly in the muscle26. Another study performed in wild-type and PPARα-deficient mice suggested two separate pathways to explain bexarotene action on plasma triglyceride concentrations, a PPARα-dependent pathway by activation of the RXR/PPARα-heterodimer for a TG-decreasing effect of bexarotene and a PPARα-independent pathway for the TG-raising effect of bexarotene but without identifying the heterodimer implicated27. As a previous study performed in our laboratory in the apolipoprotein E2 knock-in mouse model showed a bexarotene-induced hypertriglyceridemia associated with an increase in the hepatic expression of FAS and SCD1, two direct LXR-target genes implicated in lipogenesis6, we assessed whether hypertriglyceridemia induced by the RXR agonist bexarotene could involve the RXR/LXR pathway. Indeed, the heterodimer RXR/LXR is known to be permissive and the activation of this heterodimer by LXR agonists induces hepatic steatosis and hypertriglyceridemia13, 14. By measuring plasma triglyceride concentrations in wild type and LXRα/β-deficient mice, we demonstrated that bexarotene-induced hypertriglyceridemia occurred in a dose-dependent manner through the RXR/LXR pathway. This effect was associated with an upregulation of hepatic lipogenic genes (SREBP1c, FAS, FAE and SCD1) in a dose- and LXR-dependent manner. The implication of other heterodimers as mediators of the effects of bexarotene in the liver was also analyzed. Bexarotene activation of the permissive heterodimers RXR/PPAR or RXR/FXR was not affected by LXR-deficiency since both ACO (RXR/PPAR target gene) and BSEP (RXR/FXR target gene) expression was strongly increased by bexarotene in wild type and LXRα/β-deficient mice. The potential role of the thyroid hormone axis in the triglyceride response to bexarotene was also dismissed due to the fact that bexarotene decreased DIO1 (RXR/TR target gene) expression to a similar extent in wild type and LXRα/β-deficient mice. Cyp3A11 (RXR/PXR target gene) expression was not modified in the liver of bexarotene-treated wild type mice, suggesting that RXR/PXR heterodimers do not mediate bexarotene-induced lipogenesis in wild type mice.
The fact that the RXR/LXR heterodimer mediates bexarotene-induced hypertriglyceridemia suggests that it could also be implicated in hypertriglyceridemia reported in vivo with many other RXR agonists26, 28. It is, however, difficult to extrapolate to other rexinoids because each rexinoid selectively activates its own pattern of heterodimers29, 30. However, LG10506, a new rexinoid shown to selectively activate RXR/PPAR but not RXR/LXR, RXR/RAR and RXR/FXR heterodimers, has beneficial metabolic effects without inducing hypertriglyceridemia31, reinforcing the hypothesis that, in case of RXR agonist-induced hypertriglyceridemia, LXR seems to be the main partner implicated.
Notably, the results of this study support a new interesting concept concerning the molecular mechanisms of RXR heterodimer action. Indeed, we identify the existence of heterodimer-specific selective permissivity on target genes of specific metabolic pathways in one given tissue, i.e. the liver. The concept of selective permissivity linked to tissue-specific activity has previously been introduced with respect to RXR heterodimers32. The heterodimer RXR/TR, which was shown to be non-permissive, displays a permissive activity depending on the cell type33–35. Conversely, the heterodimer RXR/FXR, shown to be permissive in different assays in vitro, can be antagonized by RXR agonists32. Taken together, these studies have demonstrated tissue-specific differences in permissivity of certain RXR heterodimers. This concept has been extended in our study to the LXR/RXR heterodimer, since bexarotene treatment increased ABCG5 and ABCG8 mRNA levels in an LXR-dependent manner in the jejunum, but not in the liver, suggesting the existence of distinct molecular mechanisms of LXR target gene regulation in the intestine vs the liver. Moreover, we show that, in liver, RXR heterodimers can also act with a selective permissivity on specific metabolic pathways. Indeed, similar to the LXR agonist T0901317, bexarotene increased the recruitment of RXR on the LXREs of specific LXR-target genes implicated in hepatic lipogenesis (SREBP1c, SCD1 and FAS), leading to their upregulation and hepatic accumulation of triglycerides. Thus, the RXR/LXR heterodimer is permissive for lipogenesis in the liver. By contrast, whereas the LXR agonist induces the recruitment of RXR on the LXREs of hepatic genes encoding proteins implicated in cholesterol homeostasis, leading to a strong increase of their expression in an LXR-dependent manner, the RXR agonist bexarotene only slightly increased the expression of these genes and did not influence liver and plasma cholesterol homeostasis, either in wild-type or in LXRα/β-deficient mice. Taken together, these results demonstrate that the heterodimer RXR/LXR is non-permissive for cholesterol homeostasis in the liver. Thus, these data show that, in the same organ (ie the liver in our study), an RXR heterodimer (in this case RXR/LXR) may be permissive for target genes of specific metabolic pathways. These observations reinforce the concept that rexinoids can have molecule-, inter-tissue-and intra-tissue specific effects, which may find application in drug development by synthesizing RXR modulators with appropriate tissue and gene selective profiles for a potential clinical use.
In conclusion, this study shows that the molecular mechanism of bexarotene-induced hypertriglyceridemia is dependent on the RXR/LXR pathway and that in liver the in vivo permissive effect of bexarotene on the RXR/LXR heterodimer is restricted to specific lipogenic LXR-target genes but does not affect LXR-target genes implicated in cholesterol homeostasis. In addition, our study supports the notion of selective permissivity on target genes of specific metabolic pathways for the mode of action of RXR heterodimers.
MATERIALS AND METHODS
Animal experiments
All experiments were performed with the approval of the Pasteur Institute review board, Lille France. Bexarotene was synthesized in the Laboratoire de Chimie Thérapeutique (Faculté des Sciences Pharmaceutiques, Université de Lille 2, France).
Female C57BL6 (13 weeks of age) wild type (n=28) or LXRα/β-deficient (n=28) mice36, 37 obtained from Jan-Ake Gustafsson's laboratory were divided into four groups (n=7/group) and were treated with the RXR agonist bexarotene (suspended in 1% carboxymethylcellulose at the doses of 30 or 100 mg/kg body weight (mpk)), the LXR agonist T0901317 (30 mpk) or vehicle alone by oral gavage once daily for 14 days. At the end of the treatment, blood was collected by retro-orbital venipuncture under isoflurane anesthesia after 4h of fasting (9am–1pm). Plasma was separated by centrifugation (15 min, 4°C, 4 000 rpm) and used within 3 days for biochemical analysis. Animals were weighed and sacrificed by cervical dislocation. Livers were removed immediately, frozen in nitrogen and stored at −80°C for further analysis.
For expanded Material and Methods, please see supplemental information.
Supplementary Material
Acknowledgments
We thank Véronique Touche for excellent technical assistance.
Sources of funding
This work was supported by grants from the Swedish Research Council and European FP6 STREP project X-TRA-NET (018882), the Foundation Coeur et Artères, the Danish Natural Science Research Council, Conseil Régional Nord-Pas de Calais and Fond Européen de Développement.
Footnotes
Disclosure
None
References
- 1.Boehm MF, Zhang L, Zhi L, McClurg MR, Berger E, Wagoner M, Mais DE, Suto CM, Davies JA, Heyman RA, et al. Design and synthesis of potent retinoid X receptor selective ligands that induce apoptosis in leukemia cells. J Med Chem. 1995;38:3146–3155. doi: 10.1021/jm00016a018. [DOI] [PubMed] [Google Scholar]
- 2.Hamann LG. An efficient, stereospecific synthesis of the dimer-selective retinoid X receptor modulator (2E,4E,6Z)-7-[5,6,7,8- tetrahydro-5,5, 8,8-tetramethyl-2-(n-propyloxy)naphthalen-3-yl]-3-methyl octa-2,4, 6-trienoic acid. J Org Chem. 2000;65:3233–3235. doi: 10.1021/jo991936x. [DOI] [PubMed] [Google Scholar]
- 3.Querfeld C, Nagelli LV, Rosen ST, Kuzel TM, Guitart J. Bexarotene in the treatment of cutaneous T-cell lymphoma. Expert Opin Pharmacother. 2006;7:907–915. doi: 10.1517/14656566.7.7.907. [DOI] [PubMed] [Google Scholar]
- 4.Farol LT, Hymes KB. Bexarotene: a clinical review. Expert Rev Anticancer Ther. 2004;4:180–188. doi: 10.1586/14737140.4.2.180. [DOI] [PubMed] [Google Scholar]
- 5.Pinaire JA, Reifel-Miller A. Therapeutic potential of retinoid x receptor modulators for the treatment of the metabolic syndrome. PPAR Res. 2007;2007:94156. doi: 10.1155/2007/94156. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Lalloyer F, Fievet C, Lestavel S, Torpier G, van der Veen J, Touche V, Bultel S, Yous S, Kuipers F, Paumelle R, Fruchart JC, Staels B, Tailleux A. The RXR agonist bexarotene improves cholesterol homeostasis and inhibits atherosclerosis progression in a mouse model of mixed dyslipidemia. Arterioscler Thromb Vasc Biol. 2006;26:2731–2737. doi: 10.1161/01.ATV.0000248101.93488.84. [DOI] [PubMed] [Google Scholar]
- 7.Hurst RE. Bexarotene ligand pharmaceuticals. Curr Opin Investig Drugs. 2000;1:514–523. [PubMed] [Google Scholar]
- 8.Staels B. Regulation of lipid and lipoprotein metabolism by retinoids. J Am Acad Dermatol. 2001;45:S158–167. doi: 10.1067/mjd.2001.113718. [DOI] [PubMed] [Google Scholar]
- 9.Austin MA. Plasma triglyceride as a risk factor for cardiovascular disease. Can J Cardiol. 1998;14 (Suppl B):14B–17B. [PubMed] [Google Scholar]
- 10.McBride PE. Triglycerides and risk for coronary heart disease. Jama. 2007;298:336–338. doi: 10.1001/jama.298.3.336. [DOI] [PubMed] [Google Scholar]
- 11.Lefebvre P, Chinetti G, Fruchart JC, Staels B. Sorting out the roles of PPAR alpha in energy metabolism and vascular homeostasis. J Clin Invest. 2006;116:571–580. doi: 10.1172/JCI27989. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Chiang JY. Bile acid regulation of gene expression: roles of nuclear hormone receptors. Endocr Rev. 2002;23:443–463. doi: 10.1210/er.2000-0035. [DOI] [PubMed] [Google Scholar]
- 13.Schultz JR, Tu H, Luk A, Repa JJ, Medina JC, Li L, Schwendner S, Wang S, Thoolen M, Mangelsdorf DJ, Lustig KD, Shan B. Role of LXRs in control of lipogenesis. Genes Dev. 2000;14:2831–2838. doi: 10.1101/gad.850400. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Grefhorst A, Elzinga BM, Voshol PJ, Plosch T, Kok T, Bloks VW, van der Sluijs FH, Havekes LM, Romijn JA, Verkade HJ, Kuipers F. Stimulation of lipogenesis by pharmacological activation of the liver X receptor leads to production of large, triglyceride-rich very low density lipoprotein particles. J Biol Chem. 2002;277:34182–34190. doi: 10.1074/jbc.M204887200. [DOI] [PubMed] [Google Scholar]
- 15.Yoshikawa T, Shimano H, Amemiya-Kudo M, Yahagi N, Hasty AH, Matsuzaka T, Okazaki H, Tamura Y, Iizuka Y, Ohashi K, Osuga J, Harada K, Gotoda T, Kimura S, Ishibashi S, Yamada N. Identification of liver X receptor-retinoid X receptor as an activator of the sterol regulatory element-binding protein 1 c gene promoter. Mol Cell Biol. 2001;21:2991–3000. doi: 10.1128/MCB.21.9.2991-3000.2001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Repa JJ, Liang G, Ou J, Bashmakov Y, Lobaccaro JM, Shimomura I, Shan B, Brown MS, Goldstein JL, Mangelsdorf DJ. Regulation of mouse sterol regulatory element-binding protein-1c gene (SREBP-1c) by oxysterol receptors, LXRalpha and LXRbeta. Genes Dev. 2000;14:2819–2830. doi: 10.1101/gad.844900. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Cha JY, Repa JJ. The liver X receptor (LXR) and hepatic lipogenesis. The carbohydrate-response element-binding protein is a target gene of LXR. J Biol Chem. 2007;282:743–751. doi: 10.1074/jbc.M605023200. [DOI] [PubMed] [Google Scholar]
- 18.Janowski BA, Willy PJ, Devi TR, Falck JR, Mangelsdorf DJ. An oxysterol signalling pathway mediated by the nuclear receptor LXR alpha. Nature. 1996;383:728–731. doi: 10.1038/383728a0. [DOI] [PubMed] [Google Scholar]
- 19.Lehmann JM, Kliewer SA, Moore LB, Smith-Oliver TA, Oliver BB, Su JL, Sundseth SS, Winegar DA, Blanchard DE, Spencer TA, Willson TM. Activation of the nuclear receptor LXR by oxysterols defines a new hormone response pathway. J Biol Chem. 1997;272:3137–3140. doi: 10.1074/jbc.272.6.3137. [DOI] [PubMed] [Google Scholar]
- 20.Repa JJ, Berge KE, Pomajzl C, Richardson JA, Hobbs H, Mangelsdorf DJ. Regulation of ATP-binding cassette sterol transporters ABCG5 and ABCG8 by the liver X receptors alpha and beta. J Biol Chem. 2002;277:18793–18800. doi: 10.1074/jbc.M109927200. [DOI] [PubMed] [Google Scholar]
- 21.Nielsen R, Pedersen TA, Hagenbeek D, Moulos P, Siersbaek R, Megens E, Denissov S, Borgesen M, Francoijs KJ, Mandrup S, Stunnenberg HG. Genome-wide profiling of PPARgamma:RXR and RNA polymerase II occupancy reveals temporal activation of distinct metabolic pathways and changes in RXR dimer composition during adipogenesis. Genes Dev. 2008;22:2953–2967. doi: 10.1101/gad.501108. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Wang Y, Kurdi-Haidar B, Oram JF. LXR-mediated activation of macrophage stearoyl-CoA desaturase generates unsaturated fatty acids that destabilize ABCA1. J Lipid Res. 2004;45:972–980. doi: 10.1194/jlr.M400011-JLR200. [DOI] [PubMed] [Google Scholar]
- 23.Yu L, York J, von Bergmann K, Lutjohann D, Cohen JC, Hobbs HH. Stimulation of cholesterol excretion by the liver X receptor agonist requires ATP-binding cassette transporters G5 and G8. J Biol Chem. 2003;278:15565–15570. doi: 10.1074/jbc.M301311200. [DOI] [PubMed] [Google Scholar]
- 24.Miller VA, Benedetti FM, Rigas JR, Verret AL, Pfister DG, Straus D, Kris MG, Crisp M, Heyman R, Loewen GR, Truglia JA, Warrell RP., Jr Initial clinical trial of a selective retinoid X receptor ligand, LGD1069. J Clin Oncol. 1997;15:790–795. doi: 10.1200/JCO.1997.15.2.790. [DOI] [PubMed] [Google Scholar]
- 25.Rizvi NA, Marshall JL, Dahut W, Ness E, Truglia JA, Loewen G, Gill GM, Ulm EH, Geiser R, Jaunakais D, Hawkins MJ. A Phase I study of LGD1069 in adults with advanced cancer. Clin Cancer Res. 1999;5:1658–1664. [PubMed] [Google Scholar]
- 26.Davies PJ, Berry SA, Shipley GL, Eckel RH, Hennuyer N, Crombie DL, Ogilvie KM, Peinado-Onsurbe J, Fievet C, Leibowitz MD, Heyman RA, Auwerx J. Metabolic effects of rexinoids: tissue-specific regulation of lipoprotein lipase activity. Mol Pharmacol. 2001;59:170–176. doi: 10.1124/mol.59.2.170. [DOI] [PubMed] [Google Scholar]
- 27.Ouamrane L, Larrieu G, Gauthier B, Pineau T. RXR activators molecular signalling: involvement of a PPAR alpha-dependent pathway in the liver and kidney, evidence for an alternative pathway in the heart. Br J Pharmacol. 2003;138:845–854. doi: 10.1038/sj.bjp.0705113. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Claudel T, Leibowitz MD, Fievet C, Tailleux A, Wagner B, Repa JJ, Torpier G, Lobaccaro JM, Paterniti JR, Mangelsdorf DJ, Heyman RA, Auwerx J. Reduction of atherosclerosis in apolipoprotein E knockout mice by activation of the retinoid X receptor. Proc Natl Acad Sci U S A. 2001;98:2610–2615. doi: 10.1073/pnas.041609298. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Lala DS, Mukherjee R, Schulman IG, Koch SS, Dardashti LJ, Nadzan AM, Croston GE, Evans RM, Heyman RA. Activation of specific RXR heterodimers by an antagonist of RXR homodimers. Nature. 1996;383:450–453. doi: 10.1038/383450a0. [DOI] [PubMed] [Google Scholar]
- 30.Cesario RM, Klausing K, Razzaghi H, Crombie D, Rungta D, Heyman RA, Lala DS. The rexinoid LG100754 is a novel RXR:PPARgamma agonist and decreases glucose levels in vivo. Mol Endocrinol. 2001;15:1360–1369. doi: 10.1210/mend.15.8.0677. [DOI] [PubMed] [Google Scholar]
- 31.Leibowitz MD, Ardecky RJ, Boehm MF, Broderick CL, Carfagna MA, Crombie DL, D’Arrigo J, Etgen GJ, Faul MM, Grese TA, Havel H, Hein NI, Heyman RA, Jolley D, Klausing K, Liu S, Mais DE, Mapes CM, Marschke KB, Michellys PY, Montrose-Rafizadeh C, Ogilvie KM, Pascual B, Rungta D, Tyhonas JS, Urcan MS, Wardlow M, Yumibe N, Reifel-Miller A. Biological characterization of a heterodimer-selective retinoid X receptor modulator: potential benefits for the treatment of type 2 diabetes. Endocrinology. 2006;147:1044–1053. doi: 10.1210/en.2005-0690. [DOI] [PubMed] [Google Scholar]
- 32.Kassam A, Miao B, Young PR, Mukherjee R. Retinoid X receptor (RXR) agonist-induced antagonism of farnesoid X receptor (FXR) activity due to absence of coactivator recruitment and decreased DNA binding. J Biol Chem. 2003;278:10028–10032. doi: 10.1074/jbc.M208312200. [DOI] [PubMed] [Google Scholar]
- 33.Li D, Li T, Wang F, Tian H, Samuels HH. Functional evidence for retinoid X receptor (RXR) as a nonsilent partner in the thyroid hormone receptor/RXR heterodimer. Mol Cell Biol. 2002;22:5782–5792. doi: 10.1128/MCB.22.16.5782-5792.2002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Li D, Yamada T, Wang F, Vulin AI, Samuels HH. Novel roles of retinoid X receptor (RXR) and RXR ligand in dynamically modulating the activity of the thyroid hormone receptor/RXR heterodimer. J Biol Chem. 2004;279:7427–7437. doi: 10.1074/jbc.M311596200. [DOI] [PubMed] [Google Scholar]
- 35.Castillo AI, Sanchez-Martinez R, Moreno JL, Martinez-Iglesias OA, Palacios D, Aranda A. A permissive retinoid X receptor/thyroid hormone receptor heterodimer allows stimulation of prolactin gene transcription by thyroid hormone and 9-cis-retinoic acid. Mol Cell Biol. 2004;24:502–513. doi: 10.1128/MCB.24.2.502-513.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Alberti S, Schuster G, Parini P, Feltkamp D, Diczfalusy U, Rudling M, Angelin B, Bjorkhem I, Pettersson S, Gustafsson JA. Hepatic cholesterol metabolism and resistance to dietary cholesterol in LXRbeta-deficient mice. J Clin Invest. 2001;107:565–573. doi: 10.1172/JCI9794. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Schuster GU, Parini P, Wang L, Alberti S, Steffensen KR, Hansson GK, Angelin B, Gustafsson JA. Accumulation of foam cells in liver X receptor-deficient mice. Circulation. 2002;106:1147–1153. doi: 10.1161/01.cir.0000026802.79202.96. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.