

Summary
Triglyceride storage in adipose tissue comprises the principal energy reserve in mammals. Additionally glucose can be stored as glycogen in the fed state, primarily in liver and skeletal muscle, for mobilization during times of energy deficit. Adipose tissue also contains glycogen stores albeit at very low levels. The physiological role of glycogen metabolism in adipocytes remains unclear. However, both classical literature and more recent work demonstrate that the dynamic regulation of adipose glycogen may serve as an energy sensing modality in the coordination of glucose and lipid metabolism in adipose tissue, especially during the fasted to fed transition.
Keywords: Lipolysis, Glyceroneogenesis, Glycogen, Adipocyte, Pepck, Energy Sensing
1. Introduction
Investigations spanning over a century have addressed the dynamics of adipocyte energy flux and the role of glycogen metabolism in these processes. A recent publication (Birsoy et al. 2008) highlighted the important yet undefined role of glycogen turnover during lipid synthesis in adipose tissue. Their investigation into the altered genetic programming of the adipocyte during the transition from lipid depletion to restoration recapitulated the physiological transient elevation of glycogen levels in adipose first characterized by Tuerkischer and Wertheimer in 1942. These two publications separated by 66 years underscore the need for a fuller understanding of the role of glycogen turnover in the regulation of adipocyte energy metabolism. In concert with the intervening body of literature, data from these two publications suggest a potential role of adipose glycogen metabolism as a sensing modality of adipocyte energy flux. Exploration of the data supporting this novel hypothesis is the central focus of the current review.
2. Adipose Tissue Overview
More than 10–15 kg of body composition in a young, healthy adult is dedicated to adipose tissue. This equates to 135,000 kcal or roughly the energy contained in 200 meals (Large et al. 2004) In contrast, glycogen is the principle energy storage form in liver and muscle and these two tissues are the principle sites for the study of glycogen metabolism. Glycogen is also present in adipocytes albeit at much lower levels and the study of adipose glycogen metabolism has a long yet underappreciated history. Examination of past literature and current studies indicate a potential three pronged role for adipose glycogen that includes it functioning as a metabolic switch under varied nutritional states, providing a carbon source within the adipocyte, and as a sensor of energy flux through the cell. The experimental foundation for these three proposed roles spans the last century.
The prevailing thought prior to the 1940s regarding fat was that lipid storage in adipose tissue was directly related to circulating lipid levels. It was accepted that fat diffused into the adipocyte “as through a sieve” (Tuerkischer and Wertheimer 1942). Thus adipose tissue was regarded merely as a passive and inert storage depot. Although widely accepted at the time several observations were noted at the turn of the 20th century that challenged this perception. In 1903 Rosenfeld noted that geese being gavaged accumulated fat in adipose tissue at a rate that outpaced what could be formed physiologically in liver or through blood transport (Rosenfeld 1903). Thus he supposed that the adipocyte could possibly form fat from carbohydrate via its own internal metabolism. Soon afterward, von Geirke noted the rapid accumulation of a polysaccharide in adipose tissue in the fasted to fed transition period (von Gierke 1906). A multitude of studies conducted in the following years investigating the deposition of this poly-saccharide and lipid within adipose tissue challenged the notion that adipose tissue did not play an active metabolic role (Tuerkischer and Wertheimer 1942). In 1942 a seminal publication by Tuerkischer and Wertheimer confirmed the poly-saccharide as glycogen. The experimental design entailed several days of calorie restriction in rats, followed by unrestricted access to food. Upon this transition a massive, transient deposition of glycogen was observed in adipose tissue comparatively much higher than basal glycogen levels in the fed state. Furthermore the return of lipid stores in the re-fed state occurred only after the initial spike in glycogen. Maximal fat repletion was achieved by 4 days of re-feeding at which point glycogen levels returned to trace amounts. These studies were paramount for two main reasons. First, because glycogen can not be transported across the cell membrane, this result demonstrated that the adipocyte was metabolically active, synthesizing glycogen de novo from glucose. Second, these studies indicated during the fasted to fed transition glycogen and lipid synthesis may be linked. Thus the investigators concluded, “The disappearance of adipose glycogen may thus be said to mark the re-establishment of a fat equilibrium in the organism.” The work of Tuerkischer and Wertheimer laid the foundation by corroborating ‘strong evidence’ for the ‘interdependence’ between the dynamics of fat and glycogen metabolism in adipose tissue that is still actively studied. Subsequent work detailed below demonstrated that glycogen stores are preferentially replenished over lipid synthesis (Moody and Gliemann 1968) further supporting this supposition.
3. Lipid Metabolism in the Adipocyte
To better understand how glycogen metabolism functions in relation to the lipid equilibrium of adipose tissue, a closer examination of adipocyte lipid dynamics in the fed state is necessary. Lipoprotein lipase localized to the plasma membrane of the adipocyte, hydrolyzes dietary lipid (Mead et al. 2002) thereby allowing for entry into the cell through either the action of fatty acid transport proteins (Abumrad et al. 1993; Isola et al. 1995; Schaffer and Lodish 1994) and/or a passive “flip-flop” transition through the plasma membrane. Following cellular uptake, these fatty acids are quickly converted into fatty acyl-CoA by acyl-CoA synthetase. In concert, insulin stimulates glucose uptake via translocation of the GLUT4 glucose transporter to the plasma membrane. The glucose can then be partially metabolized through the initial steps of glycolysis to produce dihydroxyacetone-phosphate (DHAP) which is further reduced forming glycerol-3-phosphate (G3P). Triglyceride is then formed through the esterification of three fatty acyl-CoA molecules to the G3P backbone for storage in the central lipid droplet. Additionally, excess glucose influx may also be utilized for the production of fatty acids de novo and storage as triglyceride. Thus carbohydrate and lipid metabolism are inherently connected in energy metabolism of the adipocyte.
Catecholamines represent the principal signal eliciting energy mobilization from the adipocyte via lipolysis. Hormones including but not limited to glucocorticoids, ACTH, thyroid hormone, and glucagon also play a role. Signaling through beta-adrenergic receptors activate adenylate cyclase resulting in increased cAMP and activation of protein kinase A (PKA) resulting in the phosphorylation and activation of hormone sensitive lipase. A proposed model of sequential lipase action entails the hydrolysis of the initial triacylglycerol ester bond by the lipase desnutrin/ATGL yielding diacylglycerol which is subsequently hydrolyzed by activated hormone sensitive lipase. Monoacylglycerol is produced via hormone sensitive lipase action before it is finally hydrolyzed to fatty acid through monoacylglycerol lipase action (Jaworski et al. 2007). Additionally, PKA phosphorylates perilipin exposing the central lipid drop surface to lipase action. Upon completion of hydrolysis the glycerol and free fatty acids are shuttled into circulation where glycerol is primarily utilized by liver for gluconeogenesis and the free fatty acids are oxidized in skeletal muscle. Catecholamines also trigger glycogenolysis in the adipocyte. As triglyceride is hydrolyzed, already low levels of glycogen become nearly undetectable under these conditions underscoring a potential link between glycogen and lipid metabolism.
4. Glycogen Metabolism in the Adipocyte
Glycogen synthesis, although maintained at much lower levels relative to muscle and liver, is dynamically regulated within the adipocyte. Insulin signaling leads to the activation of glycogen synthase partly through the targeted action of protein phosphatase-1 (PP1). Assembly of the glycogen poly-saccharide is initiated through the self-glucosylation of glycogenin (Roach and Skurat 1997) and activation of glycogen synthase. Glucose that has been taken up by the cell is rapidly modified from glucose-6-phosphate (G6P) to glucose-1-phosphate (G1P) to UDP-glucose that is then incorporated into the growing glycogen particle via α1–4 linkages. Once 11 residues of glucose have been incorporated into the glycogen chain, a α1–6 linkage is utilized by branching enzyme to establish a new branch consisting of at least 7 residues. As the process continues a structure of 12 concentric tiers is achieved (Melendez et al. 1999).
As is the case with lipolysis, elevation of cAMP levels in the adipocyte catalyzes glycogenolysis. PKA phosphorylates and activates phosphorylase kinase which is the sole covalent activator of glycogen phosphorylase. Active phosphorylase promotes glycogenolysis, the net breakdown of glycogen. Specifically, glycogen phosphorylase hydrolyzes the α1–4 linkages allowing for the liberation of G1P. As glycogenolysis proceeds and four glucose residues remain prior to a branch point, debranching enzyme acts via transferase activity to move 3 glucose moieties to a proximal branch of the glycogen particle. This reorganization is followed by the cleavage of the α1–6 bond by debranching enzyme. This structural rearrangement within the glycogen chain allows for the continued hydrolysis and mobilization of the glucose residues out of glycogen and into G1P which can be metabolized for entrance into the glycolytic pathway for production of ATP while fatty acids are mobilized out of the cell. The highly ordered enzymatic regulation of glycogen synthesis and breakdown ensures proper energy flux through glycogen is maintained in relation to the current nutritional state.
5. Metabolic Transitions
It is well understood that elevated blood glucose stimulates insulin secretion. Elevated insulin levels suppress adipocyte fatty acid release thereby allowing muscle to utilize the circulating glucose. Elevated plasma non-esterified fatty acid (NEFA) levels typically correspond to low glucose and insulin levels. This cycle of fuel selectivity in circulation is known as the glucose-fatty acid cycle (Randle et al. 1963). Similar regulatory mechanisms regarding fuel selectivity exist within the adipocyte. The integration of metabolic cues together with detection of carbon flux between glucose and triglycerides is vital during times of metabolic transition ensuring proper fuel utilization in the adipocyte. Examination of these regulatory mechanisms begins to define a potential fundamental purpose of glycogen in adipose tissue.
5.1. The Fed State
In the fed state glucose enters the adipocyte through insulin stimulated GLUT4 translocation and is rapidly phosphorylated to form G6P. As alluded to above, this G6P has several metabolic fates including glycolysis, triglyceride synthesis, the pentose phosphate shunt or glycogen incorporation. Early studies in the 1950s utilizing 14C-glucose demonstrated the interdependence of glucose and lipid metabolism (Hausberger et al. 1954). Utilizing similar techniques, Flatt and Ball were able to calculate the flux of glucose through specific metabolic pathways (Flatt and Ball 1964). They reported the following breakdown of recovered glucose: 37% CO2; 50% triglyceride (88% fatty acid and 12% glycerol); 4.5% glycogen and 6.5% lactate. The percentage of glycogen accounting for total adipocyte energy stores ranks minimal relative to other energy forms but should not negate it from fulfilling a vital integrative role. Again it should be noted that conditions favoring triglyceride formation also favor glycogen synthesis. Studies by Leonards and Landau reported rat adipose in vitro incorporated the greatest amount of carbon-6 labeled 14C-glucose into glycogen in medium containing insulin and low concentrations of glucose (Leonards and Landau 1960). High levels of glucose without insulin were not able to achieve an equivalent rate of incorporation into glycogen. Exceedingly high levels of glucose alone were needed to achieve the equivalent incorporation of 14C-glucose into fatty acids as in media containing the combination of insulin and low glucose. Utilizing similar isotope tracer studies Moody and Gliemann concluded extremely low insulin concentrations preferentially direct glucose flux into the pathway of glycogen metabolism (Moody and Gliemann 1968). These studies demonstrated a preference of the adipocyte to replenish glycogen stores prior to filling lipid stores. How does the adipocyte determine which metabolic pathway will receive the flux of the G6P? Utilization appears dependent upon nutritional status of the cell. Therefore an integrative mechanism linking nutritional status with carbohydrate versus lipid metabolism likely exists. The metabolic transitions that occur during the fasted to fed state suggest that this integration may occur via a glycogen intermediary.
5.2. The Fasted State
As predicted by the glucose-fatty acid model under a fasted state circulating NEFAs are elevated in order to provide fuel substrate to peripheral tissues, sparing glucose for utilization by the brain. The adipocyte is the key source of mobilized NEFAs. Based on the text book model of lipolysis, one would expect the hydrolysis of triglyceride to result in a 3:1 ratio of released free fatty acid to glycerol. However, it had been observed under in vitro conditions lacking the addition of glucose that adipose tissue re-esterified approximately 30% of the free fatty acid liberated through lipolysis (Vaughan 1962). Additionally in humans starved for 60h, approximately 60% of the free fatty acid released through lipolysis was re-esterified back into triacylglyceride (Jensen et al. 2001). This seemingly futile cycling was labeled the triglyceride-fatty acid cycle. Initially this discovery was puzzling. Why would the adipocyte re-esterify free fatty acids back into storage as lipid when the body demanded their mobilization? Also as glycerol kinase activity is negligible in adipocytes and glucose uptake is minimal under fasted conditions, what was the likely source of G3P providing the backbone for the fatty acid re-esterification? These driving questions foreshadowed the discovery that adipocytes express the cytosolic isoform of the key gluconeogenic enzyme phosphoenolpyruvate carboxykinase (PEPCK-C) (Ballard et al. 1967). Its characterization in adipose tissue defined a new pathway termed glyceroneogenesis, in which an abbreviated version of gluconeogenesis yields G3P from TCA intermediaries. PEPCK is up regulated under fasting conditions as a mechanism preventing systemic lipotoxicity that could occur upon excessive efflux of FFA from the adipocyte. Specifically, PEPCK activity catalyzes the conversion of oxalacetate derived from the TCA cycle into phosphoenolpyruvate which is further utilized into G3P production (Figure 1). The characterization of glyceroneogenesis underscores that although the adipocyte is able to rapidly mobilize energy additional mechanisms are active in fine tuning the level of energy released to match utilization by other body tissues (Nye et al. 2008). Integration of these metabolic pathways in the adipocyte is crucial to maintaining global energy balance. Under fasting conditions it is evident that carbon continues to course through the adipocyte allowing for the triglyceride-fatty acid cycle. What remains undetermined is if carbon flux through glycogen can also provide G3P precursors for this cycle (Figure 1).
Figure 1. Free fatty acid re-esterification during fasting.
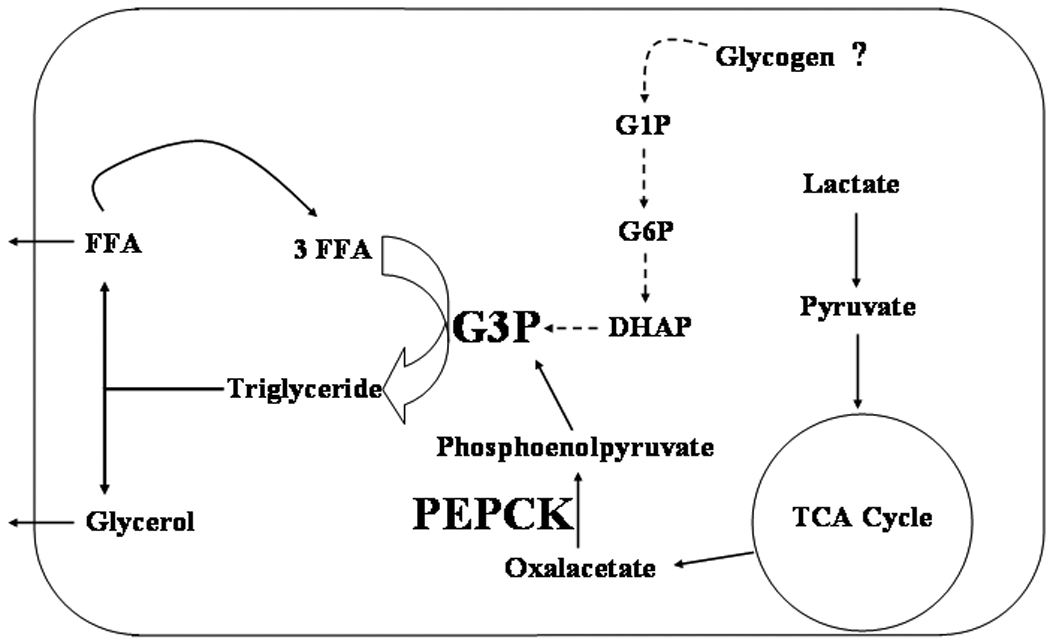
In the absence of glucose, the fatty acid-triglyceride cycle occurs in which hydrolyzed free fatty acids are re-esterified back into lipid storage. Glycerol kinase activity is exceedingly low in adipocytes therefore glycerol released by lipolysis can not be efficiently utilized in the fatty acid-triglyceride cycle. During fasting conditions, PEPCK expression is increased and G3P is produced through glyceroneogenesis. The key step involves the conversion of oxalacetate from the TCA cycle into phosphoenolypyruvate. Alternatively, G1P released from glycogen could be metabolized to G3P, potentially integrating glycogenolysis and lipid metabolism under certain conditions.
5.3. The Re-Fed State
The studies of Tuerkischer and Wertheimer poignantly illustrated the transition of adipose tissue from the fasted to the fed state produces a significant spike in glycogen levels. These experiments underscored that glycogen synthesis precedes lipid synthesis thereby suggesting an integrative role for glycogen under varied nutritional states. Studies through the following decades further investigated this hypothesis. Utilizing adipose from fed versus fasted versus re-fed rats and radioactive substrates, investigators concluded that starvation alters adipocyte glucose metabolism in a manner that does not strongly favor fat synthesis (Rose and Shapiro 1955). The scientists contemplated the plausibility that starvation imposes a “block” on fatty acid synthesis and metabolism favors glycogen precursors yielding the observed glycogen spike upon initial re-feeding. Full transition back into the fed state relieves this “block”, fatty acid synthesis resumes and glycogen dissipates. Conducting similar studies in a transgenic mouse model incapable of sustaining adipocyte glycogen metabolism, such as in a fat-specific glycogen synthase knockout mouse, would be potentially of great interest. Elucidation of this molecular “block” on fatty acid synthesis and how it is integrated with glycogen metabolism is required for further understanding modulation of adipocyte metabolism in the transition from fasted to re-fed states. Collectively, these data with more recent literature to be discussed in the following section strongly suggest a key role for adipose glycogen in times of metabolic transition.
6. Modulation of Glycogen Levels in Adipose Tissue
Insulin regulation of glycogen metabolism is a dynamic process involving coordination of metabolites, enzymes, and phosphatases in time and space. Insulin coordinately regulates glycogen synthase and glycogen phosphorylase through dephosphorylation, resulting in enzymatic activation and inactivation respectively. Insulin appears to regulate both kinase and phosphatase activities mediating its metabolic effects. PP1 is a critical effecter of insulin action as it dephosphorylates both synthase and phosphorylase. Interestingly a family of up to 7 targeting subunits localize the phosphatase with synthase and phosphorylase at the glycogen particles (Cohen 2002, Ceulmans et al. 2002, Munro et al. 2005). To date, protein targeting to glycogen (PTG) is the only reported PP1 glycogen-targeting subunit expressed in adipocytes (Printen et al. 1997).
Multiple studies from various groups have confirmed that modulation of PTG expression in a variety of cell types and tissue in vivo markedly alters glycogen storage (Fong et al. 2000; Gasa et al. 2000; Green et al. 2004; Greenberg et al. 2003; Lerin et al. 2000; O'Doherty et al. 2000; Printen et al. 1997; Yang et al. 2002). PTG gene deletion causes impaired glycogen synthesis and developmental insulin resistance (Crosson et al. 2003). The adenoviral mediated over expression of PTG in the 3T3-L1 adipocyte cell line induced a dramatic dephosphorylation of glycogen synthase and glycogen phosphorylase resulting in an 10-fold increase of cellular glycogen levels (Greenberg et al. 2003). Conversely, the shRNA-mediated knock down of PTG in 3T3-L1 adipocytes produced an approximately 85% reduction in glycogen levels (Greenberg et al. 2006). Interestingly, the loss of cellular glycogen levels resulted in an increase of GLUT1 expression and acute glycogen synthetic rates although these compensatory changes were unable to rescue glycogen levels in the PTG-deficient cells. However these data support the notion that the 3T3-L1 adipocytes ‘sense’ cellular glycogen levels and strive to maintain a physiological set point as also occurs physiologically in skeletal muscle. To expand upon these cell line studies, a transgenic mouse model over expressing PTG under control of the aP2-promoter was generated. It had been hypothesized that the adipocyte was ill suited to sustain significant glycogen storage due to spatial constraints imposed by the large, central lipid droplet. However, in the aP2-PTG transgenic mice adipose glycogen levels were 200- to 400-fold higher than in wild type animals (Jurczak et al. 2007). These elevated glycogen stores were maintained at least one year and could be mobilized following an overnight fast indicating the establishment of a new set point for glycogen storage in the adipose tissue. The sustained elevation of adipose glycogen is significant for it demonstrated that the adipocyte is spatially capable of accommodating far higher glycogen stores than occurs in wild type animals. Conversely, these results suggest that adipose glycogen levels are actively maintained at a low set-point rather than arising through a default mechanism. These increases in glycogen storage occurred without any detectable change in total fat mass, triglyceride storage or acute rates of lipogenesis. It is interesting to speculate that excessive accumulation of glycogen in the adipose tissue of this model has disrupted the lipid equilibrium whereby sustained elevated glycogen is indicative of energy abundance. Although speculative this hypothesis begs to be investigated on the molecular level in the fasted state. Also, the distinct differences between a dynamic process such as the flux of carbon between glucose, glycogen, and lipid and the static read-out of radioactive incorporation should be recognized. To address the question of carbon flux a detailed NMR analysis could be applied to precisely track carbon partitioning between glucose, glycogen, and triglyceride in vivo.
Additional data from this mouse model strongly supports the idea that adipose glycogen is mobilized under varied nutritional states. Transgenic adipose tissue in vivo and in vitro demonstrated robust glycogenolysis throughout a fast and following lipolytic stimulation (Jurczak et al. 2007; Markan et al. 2009). Interestingly, the up-regulation of PEPCK expression in adipose tissue following an overnight fast was largely blunted in the aP2-PTG transgenic mice (Markan et al. 2009). This result potentially indicates that G1P released via glycogenolysis can provide G3P required for free fatty acid re-esterification thus altering the contribution of PEPCK in the process. Although adipose glycogen in wild type animals may not provide a significant fuel source based on limited quantity, these studies suggest the flux of energy through glycogen metabolism is capable of modulating other metabolic pathways in the adipocyte. Additionally, glycogen metabolism may serve as an integrative metabolic regulator under conditions of energy mobilization. Conversely, the glycogen spike in the fasted to fed transition may serve as a carbon reservoir thereby supplying the re-establishment of lipid equilibrium through carbon flux out of glycogen into triglyceride. The molecular mechanisms integrating lipid and glycogen metabolism in the fed and fasted conditions remains an interesting and understudied question.
Physiological conditions do exist where adipose glycogen becomes markedly elevated. Tuerkischer and Wertheimer defined such a scenario in their 1942 publication. Sixty-six years later the phenomenom was again documented (Birsoy et al. 2008). This group’s experimental design utilized leptin administration to mice to decrease food intake in combination with simple calorie restriction in order to deplete adipocytes of detectable lipid. Upon the cessation of leptin administration simultaneously with the first day of re-feeding a massive glycogen spike was observed in adipose tissue. The group conducted an elegant analysis of genes altered during this protocol of re-feeding and found a general up regulation of all genes involved in carbohydrate metabolism, simple sugar flux, and de novo synthesis of fatty acids and triglycerides that was maintained over the next 4 days (Birsoy et al. 2008). However, the enzymatic activities of glycogen synthase and glycogen phosphorylase were not studied under these experimental conditions, so the molecular mechanisms underlying the spike in glycogen remains poorly understood. The results of this study highlight the adipocyte’s ability to shift energy flux dramatically favoring the synthesis of glycogen prior to the re-establishment of lipogenesis. These data further support the supposition that the transient glycogen spike may serve as a metabolic switch for the adipocyte after prolonged energy mobilization, enabling the restoration of triglyceride synthesis and the proper integration of adipocyte metabolism with nutritional and hormonal cues.
7. Potential Roles of Glycogen in Adipose Tissue Metabolism
A delicate equilibrium is maintained within the adipocyte between carbohydrate and lipid metabolism. The fulcrum of this equilibrium lies between energy storage and mobilization. However, the dynamics of adipose tissue extend far beyond these two states and include periods of metabolic transition. Two key points regarding adipose tissue should be recognized: 1) energy is always fluxing through the adipocyte and 2) layers of cellular detection and regulation must exist in order to fine tune this energy flux with the global metabolic requirements of the organism. Processes such as basal lipolysis, entailing the mobilization of energy in a satiated state, or glyceroneogenesis, which plays a role in the fatty acid-triglyceride cycle in the fasted state, illustrate these two key points. It was first postulated more than six decades ago that the dissipation of adipose glycogen signaled the re-setting of the lipogenic program in adipocytes. Collectively these studies have provided a foundation upon which to speculate on three potential physiological roles for glycogen in adipose tissue.
It has been presented that conditions favoring glycogen synthesis also favor lipid synthesis and that a preference for glycogen synthesis prior to lipid formation is most favorable in the transition between fasting and feeding. Due to fluctuating dynamics perhaps we are best served to think of adipose glycogen as a sensory mechanism, equating its flux with total energy flux through the cell. Much as a resting neuron maintains an electrical current around −70mV in order to smoothly transmit action potentials, maintaining low basal levels of glycogen would better enable the adipocyte to detect disequilibrium between carbohydrate and lipid metabolism. When disequilibrium arises and mobilization of energy is demanded, already low glycogen levels dissipate in concert with triglyceride hydrolysis. The observed spike in glycogen in the transition from energy mobilization to storage indicates to the cell conditions are suitable for the restoration of lipid storage. In essence, adipose glycogen could act as a metabolic switch functioning in the metabolic transitions between lipid mobilization and storage.
Based on the preceding argument it is of great interest to elucidate if glycogen itself provides energy to the cell. Essentially, the adipocyte is a fat laden cell consisting of a large central lipid droplet. As highly concentrated energy the central lipid droplet leaves relatively little area for the remaining cellular organelles. Glycogen, with its branching, concentric structure, is a bulky molecule and requires hydration. Therefore a straight-forward explanation is that spatial limitations favor low glycogen levels. Additionally, first observed in the 1960s only a small percentage of glucose taken up by the adipocyte is incorporated into glycogen (4.5%) (Flatt and Ball 1964) and it is known that adipose glucose uptake accounts for less than 10% of total body glucose utilization. Therefore it is unlikely that adipose glycogen serves as a primary fuel storage site in the fed state. However the aP2-PTG transgenic mouse model clearly illustrates that the adipocyte is capable of maintaining markedly elevated glycogen levels. Furthermore, during overnight fasting of the aP2-PTG animals, glycogen stores were mobilized, lactate levels were elevated and PEPCK expression was reduced compared to wild type animals (Markan et al. 2009). These findings indicate that G1P released during glycogenolysis is metabolized and can impact energy metabolism and gene expression in adipose tissue. Furthermore, the spike and subsequent dissipation of glycogen following the calorie restriction of wild type animals illustrates a physiological scenario in which glycogen may be providing substrate for lipid storage. In vitro studies have demonstrated that adipose tissue harvested upon re-feeding demonstrates a respiratory quotient exceeding 1.0 indicating the utilization of glycogen-derived glucose (Mirski 1942). These tantalizing data indicate that glycogen can not be ruled out as a contributing carbon source for lipid synthesis in certain metabolic states.
Delineating the molecular mechanisms that maintain low basal glycogen levels but also allow for the marked increase following a fast will help further define glycogen in the role as an energy sensor in the adipocyte. Logically, it should be postulated that the sequential activation of the key enzymes glycogen synthase and phosphorylase are involved in the transient elevation and reduction of glycogen levels. Since no changes in these enzymes expression levels were detected following withdrawal of leptin administration to mice (Birsoy et al. 2007), it is likely that transient and reversible protein phosphorylation/dephosphorylation is involved, although this supposition needs to be investigated. Such a result could be expected as a series of studies conducted in yeast demonstrated more often than not glycolytic enzymes are regulated via post-translational modification, rather than on the level of gene expression (Daran-Lapujade et al. 2007). Additionally, the role of the mTOR and AMPK signaling pathways as part of a larger metabolic regulatory network have been well studied in muscle and liver. Interplay between mTOR and AMPK pathways has been characterized and are under continued investigation in the adipocyte. Of great interest is the characterization of a glycogen-binding domain within the beta-subunit of AMPK. This subunit has been demonstrated to actively bind glycogen (Polekhina et al. 2003) in proximity to glycogen phosphorylase. Furthermore, it is known that AMPK phosphorylates glycogen synthase in muscle (Towler and Hardie 2007). Whether this glycogen-binding domain plays a functional and significant role in adipocyte glycogen metabolism remains undetermined. A third nutrient sensing molecule that is far less characterized in any tissue is PAS kinase (PASK). PASK homologues are conserved in organisms spanning from yeast to man. In yeast, the PAS domain responds to nutritional cues, thereby relieving auto-inhibition of the kinase domain. PASK then phosphorylates and activates UDP-glucose phosphorylase, the enzyme responsible for converting G1P to UDP-glucose to promote glycogen synthesis. Although the parallel mechanism has not been demonstrated in mammalian cells, the mid-region between the PAS and kinase domains directly interacts with glycogen synthase. Additionally it has been demonstrated that mammalian PASK phosphorylates muscle GS in vitro (Hao and Rutter 2008) although the physiological significance of this interaction remains unclear. Overall, the potential roles of mTOR signaling along with activities AMPK and PASK in adipocyte glycogen metabolism are currently speculative and require further study.
The low basal levels of glycogen metabolism in adipose tissue had been considered to occur through a default mechanism in which glycogen storage was restrained due to biophysical constraints of accommodating bulky glycogen chains in a limited cytoplasmic space. However, the results from the aP2-PTG transgenic mice clearly demonstrated that adipocytes are capable of storing far larger amounts of glycogen for long periods of time than was previously realized. These findings suggest that glycogen levels are actively established and likely defended in adipose tissue, rather than arising through a passive mechanism. Furthermore, the mere elevation of PTG expression induced a dramatic, chronic change in the “set point” of glycogen metabolism in transgenic adipose tissue, indicating that the regulated expression of this protein may be a critical regulator of adipocyte glycogen levels. Additional work is required to determine whether and how PTG expression is controlled under various physiological conditions in adipose tissue in vivo. A potential role for PTG in maintaining low basal levels and mediating the transient glycogen spike in the fasted to fed transition seems logical but lacks experimental proof.
The role of glycogen as an energy sensor is further defined through examination of the relationship between glycogen and the adipokine leptin. Leptin functions as a satiety signal in the hypothalamus indicating sufficient energy storage. Its circulatory levels are directly proportional to adipose tissue levels. Interestingly, in the aP2-PTG transgenic mice a 2-fold increase in circulating leptin levels was observed despite no difference in fat accumulation compared to wild-type animals. Additionally, the transitory elevation of glycogen in Birsoy’s studies was observed under hypoleptinemia, which is a state indicative of insufficient adipocyte energy storage. Leptin secretion in particular due to insulin stimulation is dependent upon nutritional status of the organism (Lee and Fried 2006). It is interesting to speculate how levels of leptin would undulate with varied carbohydrate flux through glycogen in times of metabolic transition. Although the molecular processes regulating leptin are under continued investigation the relationship between glycogen and leptin remains completely unstudied. The characterization of this relationship would further support the role of glycogen as an energy sensor and provide strong evidence further linking adipose glycogen to whole body metabolism.
8. Conclusions
Adipose tissue was first recognized as a metabolically active site due to the characterization of glycogen and not lipid metabolism. Despite this fundamental finding that served as a platform for research on-going to this day, further work is required to define the precise roles of glycogen metabolism in adipose tissue. The dynamics of adipose tissue metabolism involve shifts in energy mobilization and storage dependent on the nutritional status of the organism. A strong interdependence exists within the adipocyte between the metabolism of glucose, glycogen and lipid. The low glycogen levels in the fed state preclude adipose glycogen from serving as a significant site of energy storage. However, more classical and current literature indicate that glycogen metabolism may function as a metabolic switch, helping to re-set the lipogenic program in the adipocyte during the fasted to fed transition. Additionally, glucose flux through glycogen may play a physiological role for the coordinate regulation of lipogenesis and lipolysis under various nutritional states, which in turn could contribute to energy sensing by the adipocyte and the overall regulation of global energy homeostasis.
Footnotes
Publisher's Disclaimer: This is a PDF file of an unedited manuscript that has been accepted for publication. As a service to our customers we are providing this early version of the manuscript. The manuscript will undergo copyediting, typesetting, and review of the resulting proof before it is published in its final citable form. Please note that during the production process errors may be discovered which could affect the content, and all legal disclaimers that apply to the journal pertain.
REFERENCES
- Abumrad NA, el-Maghrabi MR, Amri EZ, Lopez E, Grimaldi PA. Cloning of a rat adipocyte membrane protein implicated in binding or transport of long-chain fatty acids that is induced during preadipocyte differentiation Homology with human CD36. J. Biol. Chem. 1993;268:17665–17668. [PubMed] [Google Scholar]
- Ballard FJ, Hanson RW, Leveille GA. Phosphoenolpyruvate carboxykinase and the synthesis of glyceride-glycerol from pyruvate in adipose tissue. J. Biol. Chem. 1967;242:2746–2750. [PubMed] [Google Scholar]
- Birsoy K, Soukas A, Torrens J, Ceccarini G, Montez J, Maffei M, Cohen P, Fayzikhodjaeva G, Viale A, Socci AD, Friedman JM. Cellular program controlling the recovery of adipose tissue mass: An in vivo imaging approach. Proc. Natl. Acad. Sci. U.S.A. 2008;105:12985–12990. doi: 10.1073/pnas.0805621105. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ceulemans H, Stalmans W, Bollen M. Regulator-driven functional diversification of protein phosphatase-1 in eukaryotic evolution. Bioessays. 2002;24:371–381. doi: 10.1002/bies.10069. [DOI] [PubMed] [Google Scholar]
- Cohen PT. Protein phosphatase 1--targeted in many directions. J. Cell. Sci. 2002;115:241–256. doi: 10.1242/jcs.115.2.241. [DOI] [PubMed] [Google Scholar]
- Crosson SM, Khan A, Printen J, Pessin JE, Saltiel AR. PTG gene deletion causes impaired glycogen synthesis and developmental insulin resistance. J. Clin. Invest. 2003;111:1423–1432. doi: 10.1172/JCI17975. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Daran-Lapujade P, Rossell S, van Gulik WM, Luttik MA, de Groot MJ, Slijper M, Heck AJ, Daran JM, de Winde JH, Westerhoff HV, Pronk JT, Bakker BM. The fluxes through glycolytic enzymes in Saccharomyces cerevisiae are predominantly regulated at posttranscriptional levels. Proc. Natl. Acad. Sci. U.S.A. 2007;104:15753–15758. doi: 10.1073/pnas.0707476104. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Flatt JP, Ball EG. Studies on the Metabolism of Adipose Tissue. An Evaluation of the Major Pathways of Glucose Catabolism as Influenced by Insulin and Epinephrine. J. Biol. Chem. 1964;239:675–685. [PubMed] [Google Scholar]
- Fong NM, Jensen TC, Shah AS, Parekh NN, Saltiel AR, Brady MJ. Identification of binding sites on protein targeting to glycogen for enzymes of glycogen metabolism. J. Biol. Chem. 2000;275:35034–35039. doi: 10.1074/jbc.M005541200. [DOI] [PubMed] [Google Scholar]
- Gasa R, Jensen PB, Berman HK, Brady MJ, DePaoli-Roach AA, Newgard CB. Distinctive regulatory and metabolic properties of glycogen-targeting subunits of protein phosphatase-1 (PTG, GL, GM/RGl) expressed in hepatocytes. J. Biol. Chem. 2000;275:26396–26403. doi: 10.1074/jbc.M002427200. [DOI] [PubMed] [Google Scholar]
- Green AR, Aiston S, Greenberg CC, Freeman S, Poucher SM, Brady MJ, Agius L. The glycogenic action of protein targeting to glycogen in hepatocytes involves multiple mechanisms including phosphorylase inactivation and glycogen synthase translocation. J. Biol. Chem. 2004;279:46474–46482. doi: 10.1074/jbc.M405660200. [DOI] [PubMed] [Google Scholar]
- Greenberg CC, Danos AM, Brady MJ. Central role for protein targeting to glycogen in the maintenance of cellular glycogen stores in 3T3-L1 adipocytes. Mol. Cell Biol. 2006;26:334–342. doi: 10.1128/MCB.26.1.334-342.2006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Greenberg CC, Meredith KN, Yan L, Brady MJ. Protein targeting to glycogen overexpression results in the specific enhancement of glycogen storage in 3T3-L1 adipocytes. J. Biol. Chem. 2003;278:30835–30842. doi: 10.1074/jbc.M303846200. [DOI] [PubMed] [Google Scholar]
- Hao HX, Rutter J. The role of PAS kinase in regulating energy metabolism. IUBMB Life. 2008;60:204–209. doi: 10.1002/iub.32. [DOI] [PubMed] [Google Scholar]
- Hausberger FX, Milstein SW, Rutman RJ. The influence of insulin on glucose utilization in adipose and hepatic tissues in vitro. J. Biol. Chem. 1954;208:431–438. [PubMed] [Google Scholar]
- Isola LM, Zhou SL, Kiang CL, Stump DD, Bradbury MW, Berk PD. 3T3 fibroblasts transfected with a cDNA for mitochondrial aspartate aminotransferase express plasma membrane fatty acid-binding protein and saturable fatty acid uptake. Proc. Natl. Acad. Sci. U.S.A. 1995;92:9866–9870. doi: 10.1073/pnas.92.21.9866. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jaworski K, Sarkadi-Nagy E, Duncan RE, Ahmadian M, Sul HS. Regulation of triglyceride metabolism. IV. Hormonal regulation of lipolysis in adipose tissue. Am. J. Physiol. Gastrointest. Liver Physiol. 2007;293:G1–G4. doi: 10.1152/ajpgi.00554.2006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jensen MD, Ekberg K, Landau BR. Lipid metabolism during fasting. Am. J. Physiol. Endocrinol. Metab. 2001;281:E789–E793. doi: 10.1152/ajpendo.2001.281.4.E789. [DOI] [PubMed] [Google Scholar]
- Jurczak MJ, Danos AM, Rehrmann VR, Allison MB, Greenberg CC, Brady MJ. Transgenic overexpression of protein targeting to glycogen markedly increases adipocytic glycogen storage in mice. Am. J. Physiol. Endocrinol. Metab. 2007;292:E952–E963. doi: 10.1152/ajpendo.00559.2006. [DOI] [PubMed] [Google Scholar]
- Large V, Peroni O, Letexier D, Ray H, Beylot M. Metabolism of lipids in human white adipocyte. Diabetes Metab. 2004;30:294–309. doi: 10.1016/s1262-3636(07)70121-0. [DOI] [PubMed] [Google Scholar]
- Lee MJ, Fried SK. Multilevel regulation of leptin storage, turnover, and secretion by feeding and insulin in rat adipose tissue. J. Lipid Res. 2006;47:1984–1993. doi: 10.1194/jlr.M600065-JLR200. [DOI] [PubMed] [Google Scholar]
- Leonards JR, Landau BR. A study of the equivalence of metabolic patterns in rat adipose tissue: insulin versus glucose concentration. Arch. Biochem. Biophys. 1960;91:194–200. doi: 10.1016/0003-9861(60)90489-6. [DOI] [PubMed] [Google Scholar]
- Lerin C, Montell E, Berman HK, Newgard CB, Gomez-Foix AM. Overexpression of protein targeting to glycogen in cultured human muscle cells stimulates glycogen synthesis independent of glycogen and glucose 6-phosphate levels. J. Biol. Chem. 2000;275:39991–39995. doi: 10.1074/jbc.M006251200. [DOI] [PubMed] [Google Scholar]
- Markan KR, Jurczak MJ, Brady MJ. Keystone Symposia on Type 2 Diabetes and Insulin Resistance. Canada: Banff; 2009. Elevated adipose glycogen stores alter free fatty acid mobilization under a fasted state. [Google Scholar]
- Meade JR, Irvine SA, Ramji D. Lipoprotein lipase structure, function, regulation and role in disease. J. Mol. Med. 2002;80:753–769. doi: 10.1007/s00109-002-0384-9. [DOI] [PubMed] [Google Scholar]
- Melendez R, Melendez-Hevia E, Canela EI. The fractal structure of glycogen: A clever solution to optimize cell metabolism. Biophys. J. 1999;77:1327–1332. doi: 10.1016/S0006-3495(99)76982-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mirski A. Metabolism of adipose tissue in vitro. Biochem. J. 1942;36:232–241. doi: 10.1042/bj0360232. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Moody AJ, Gliemann J. The effect of insulin on the glycogen metabolism of isolated fat cells. Experientia. 1968;24:628–630. doi: 10.1007/BF02153817. [DOI] [PubMed] [Google Scholar]
- Munro S, Ceulemans H, Bollen M, Diplexcito J, Cohen PT. A novel glycogen-targeting subunit of protein phosphatase 1 that is regulated by insulin and shows differential tissue distribution in humans and rodents. FEBS J. 2005;272:1478–1489. doi: 10.1111/j.1742-4658.2005.04585.x. [DOI] [PubMed] [Google Scholar]
- Nye C, Kim J, Kalhan SC, Hanson RW. Reassessing triglyceride synthesis in adipose tissue. Trends Endocrinol. Metab. 2008;19:356–361. doi: 10.1016/j.tem.2008.08.003. [DOI] [PubMed] [Google Scholar]
- O'Doherty RM, Jensen PB, Anderson P, Jones JG, Berman HK, Kearney D, Newgard CB. Activation of direct and indirect pathways of glycogen synthesis by hepatic overexpression of protein targeting to glycogen. J. Clin. Invest. 2000;105:479–488. doi: 10.1172/JCI8673. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Polekhina G, Gupta A, Michell BJ, van Denderen B, Murthy S, Feil SC, Jennings IG, Campbell DJ, Witters LA, Parker MW, Kemp BE, Stapleton D. AMPK beta subunit targets metabolic stress sensing to glycogen. Curr. Biol. 2003;13:867–871. doi: 10.1016/s0960-9822(03)00292-6. [DOI] [PubMed] [Google Scholar]
- Printen JA, Brady MJ, Saltiel AR. PTG, a protein phosphatase 1-binding protein with a role in glycogen metabolism. Science. 1997;275:1475–1478. doi: 10.1126/science.275.5305.1475. [DOI] [PubMed] [Google Scholar]
- Randle PJ, Garland PB, Hales CN, Newsholme EA. The glucose fatty-acid cycle. Its role in insulin sensitivity and the metabolic disturbances of diabetes mellitus. Lancet. 1963;1:785–789. doi: 10.1016/s0140-6736(63)91500-9. [DOI] [PubMed] [Google Scholar]
- Roach PJ, Skurat AV. Self-glucosylating initiator proteins and their role in glycogen biosynthesis. Prog. Nucleic Acid Res. Mol. Biol. 1997;57:289–316. doi: 10.1016/s0079-6603(08)60284-6. [DOI] [PubMed] [Google Scholar]
- Rose G, Shapiro B. The synthesis of fatty acids and glycogen by adipose tissue in vitro. Biochim. Biophys. Acta. 1955;18:504–510. doi: 10.1016/0006-3002(55)90141-4. [DOI] [PubMed] [Google Scholar]
- Rosenfeld G. The formation of fat, parts I and part II. Ergeb. Physiol. 1903 1, 651 and 2, 50. [Google Scholar]
- Schaffer JE, Lodish HF. Expression cloning and characterization of a novel adipocyte long chain fatty acid transport protein. Cell. 1994;79:427–436. doi: 10.1016/0092-8674(94)90252-6. [DOI] [PubMed] [Google Scholar]
- Towler MC, Hardie DG. AMP-activated protein kinase in metabolic control and insulin signaling. Circ. Res. 2007;100:328–341. doi: 10.1161/01.RES.0000256090.42690.05. [DOI] [PubMed] [Google Scholar]
- Tuerkischer E, Wertheimer E. Glycogen and adipose tissue. J. Physiol. 1942;100:385–409. doi: 10.1113/jphysiol.1942.sp003950. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Vaughan M. The production and release of glycerol by adipose tissue incubated in vitro. J. Biol. Chem. 1962;237:3354–3358. [PubMed] [Google Scholar]
- von Gierke E. About the metabolism of adipose tissue. Vert. Deutsch. Ges. Pathol. 1906;10:182. [Google Scholar]
- Yang R, Cao L, Gasa R, Brady MJ, Sherry AD, Newgard CB. Glycogen-targeting subunits and glucokinase differentially affect pathways of glycogen metabolism and their regulation in hepatocytes. J. Biol. Chem. 2002;277:1514–1523. doi: 10.1074/jbc.M107001200. [DOI] [PubMed] [Google Scholar]