

Abstract
Immune-mediated liver diseases including autoimmune and viral hepatitis are a major health problem worldwide. Natural cannabinoids such as Δ9-tetrahydrocannabinol (THC) effectively modulate immune cell function, and they have shown therapeutic potential in treating inflammatory diseases. We investigated the effects of THC in a murine model of concanavalin A (ConA)-induced hepatitis. Intraperitoneal administration of THC after ConA challenge inhibited hepatitis as shown by significant decrease in liver enzymes and reduced liver tissue injury. Furthermore, THC treatment resulted in significant suppression of crucial inflammatory cytokines in ConA-induced hepatitis. It is noteworthy that THC treatment in ConA-injected mice led to significant increase in absolute number of Forkhead helix transcription factor p3+ T regulatory cells in liver. We were surprised to find that select cannabinoid receptor (CB1 or CB2) agonists were not able to block hepatitis either independently or in combination. However, CB1/CB2 mixed agonists were able to efficiently attenuate hepatitis similar to THC. The modulatory effect of THC in ConA-induced hepatitis was reversed by both CB1 and CB2 antagonists. We also observed that endogenous cannabinoid anandamide was able to reduce hepatitis by suppressing cytokine levels. In addition, deficiency or inhibition of endocannabinoid hydrolyzing enzyme fatty acid amide hydrolase (FAAH), which leads to increased levels of endogenous cannabinoids, resulted in decreased liver injury upon ConA challenge. Our data demonstrate that targeting cannabinoid receptors using exogenous or endogenous cannabinoids and use of FAAH inhibitors may constitute novel therapeutic modalities to treat immune-mediated liver inflammation.
Liver diseases are a serious human health problem worldwide. The pathogenesis of immune-mediated liver diseases is very complex, and it involves several inflammatory cells and cytokines. Growing evidence suggests that major human liver diseases such as autoimmune and viral hepatitis are caused, in general, by activated T-cell-mediated immune responses (Chisari and Ferrari, 1995; Kita et al., 2001). Autoimmune hepatitis (AIH) is a severe form of liver disease characterized by progressive destruction of the hepatic parenchyma, cellular infiltration, hypergammaglobulinemia, and autoantibodies. The prevalence of AIH is estimated to range between 50 and 200 cases per million population, and it accounts for 5.9% of all liver transplantations in the United States. AIH affects women more than men (3.6:1), and all ages and ethnic groups are susceptible to AIH (Czaja and Freese, 2002).
T-cell-mediated hepatitis can be induced in mice by intravenous injection of the mitogenic plant lectin concanavalin A (ConA), which leads to polyclonal activation of T cells, resulting in clinical and histological symptoms of acute hepatitis, including elevation of transaminase activities within 8 to 24 h (Tiegs et al., 1992). It is a well established mouse model for AIH. Activated CD4+ T cells, natural killer T (NKT) cells, and Kupffer cells are the most prominent effector cells in this model. Together with macrophages and eosinophils, they induce hepatocyte killing by contact or indirectly by producing large amounts of inflammatory cytokines (Tiegs et al., 1992; Gantner et al., 1995; Schümann et al., 2000; Takeda et al., 2000). Among several proinflammatory cytokines involved, TNF-α and IFN-γ play a critical role in direct induction of liver cytotoxicity, because anti-TNF-α and anti-IFN-γ monoclonal antibodies (mAbs) protect mice from ConA-induced liver injury (Gantner et al., 1995; Küsters et al., 1996).
δ-9-Tetrahydrocannabinol (THC), the major psychoactive component of marijuana (Cannabis sativa), has wide-ranging pharmacological properties (Mackie, 2006). The cannabinoid compounds possess significant immunosuppressive and anti-inflammatory properties (Croxford and Yamamura, 2005). THC and cannabinoid receptor agonists have shown promise in several models of inflammation (Pertwee, 2002; Quartilho et al., 2003; Steffens et al., 2005). Cannabinoid receptors CB1 and CB2 are heterotrimeric G protein-coupled receptors, which are activated by THC, as well as by endogenous cannabinoids. Arachidonoylethanolamide (anandamide or AEA) has been identified as the major endogenous ligand for cannabinoid receptors (Devane et al., 1992). Recent studies have demonstrated the involvement of endocannabinoid system in chronic liver diseases (Julien et al., 2005; Teixeira-Clerc et al., 2006; Siegmund and Schwabe, 2008). It has been shown that endocannabinoid signaling through CB1 and CB2 receptors may produce pro- and antifibrogenic responses, respectively, in the liver during fibrosis (Julien et al., 2005; Teixeira-Clerc et al., 2006).
The aim of the present study was to determine the immunomodulatory effect of THC in the murine model of ConA-induced hepatitis. We demonstrate that a single injection of THC significantly ameliorates ConA-induced T-cell-mediated liver injury by up-regulating Forkhead helix transcription factor p3 (Foxp3)+ regulatory T cells and down-regulating inflammatory cytokines. Using select cannabinoid receptor agonists and antagonists, we demonstrate that THC mediates immune modulation in this model by signaling through both CB1 and CB2 receptors. We also demonstrate that anandamide, an endocannabinoid can effectively attenuate the disease.
Materials and Methods
Reagents
Concanavalin A, JWH-133, and methylarachidonyl fluorophosphate (MAFP) were purchased from Sigma-Aldrich (St. Louis, MO). Arachidonyl-2′-chloroethylamide (ACEA), AM251, AM630, CP55,940 and WIN55212 were from Tocris Cookson Inc. (Ellisville, MO). THC, AEA, and SR144528 were provided by National Institute on Drug Abuse, National Institutes of Health (Bethesda, MD). URB532 was from Calbiochem (San Diego, CA). The mAbs fluorescein isothiocyanate-conjugated anti-CD3, anti-CD25, phycoerythrin-conjugated anti-CD4, anti-CD19, anti-Mac3, anti-CD11a, anti-CD11b, anti-Gr-1, anti-F4/80, and phycoerythrin-Cy7 conjugated anti-NK1.1 were purchased from BD Biosciences (San Jose, CA). Anti-mouse CD25 mAb was a product of Cedarlane Laboratories (Hornby, ON, Canada).
Mice
Female C57BL/6 mice (8–12 weeks old) were purchased from National Cancer Institute, National Institutes of Health (Frederick, MD). Fatty acid amide hydrolase (FAAH) knockout mice have been described previously (Cravatt et al., 2001). Animals were housed in specific pathogen-free conditions, and all experiments were approved by Institutional Animal Care and Use Committee.
Induction of ConA-Induced Hepatitis and Drug Administration
Concanavalin A was dissolved in pyrogen-free PBS at a concentration of 2.5 mg/ml, and then it was injected intravenously at a dose of 12.5 mg/kg body weight (in 0.1 ml of PBS) to induce hepatitis as described previously (Chen et al., 2001). THC was dissolved in ethanol and diluted in PBS, and then it was administered i.p. 5 or 30 min after ConA injection. Anandamide dissolved in ethanol and JWH-133 dissolved in DMSO were diluted in PBS and injected i.p. 5 min after injecting ConA. Cannabinoid receptor antagonists AM251, AM630, or SR144528 were dissolved in DMSO containing one drop of Tween 80 (Sigma-Aldrich), and then they were further diluted in PBS and injected i.p. 1 h before injecting ConA and THC. Control groups received vehicle ethanol, DMSO or DMSO + Tween 80 vehicle similarly diluted in PBS.
Analysis of Transaminase Activities
Blood from individual mice was obtained at various times after ConA injection. The activities of liver enzymes aspartate transaminase (AST) and alanine transaminase (ALT) in the serum were determined by spectrophotometric method using commercially available assay kits (Pointe Sci. Inc., Pointe-Claire, QC, Canada), as described previously (Chen et al., 2001).
Liver Histology and TUNEL Staining
Mice were euthanized 24 h after ConA injection. Livers were fixed in 10% buffered paraformaldehyde, pH 7.4, and embedded in paraffin. Sections (5 μm) on slides were deparaffinized in xylene, rehydrated in decreasing concentrations of ethanol, and stained with hematoxylin and eosin (H&E). Slides were examined by light microscopy for infiltrating leukocytes and tissue injury. TUNEL staining on the sections was done by dead end colorimetric method using In Situ Cell Death Detection kit (Promega. Madison, WI).
Cytokine and Chemokine Measurement
Sera from individual mice were collected 6, 12, and 24 h after ConA injection and stored below −20°C. IL-2, TNF-α, IFN-γ, IL-1α, IL-1β, IL-3, IL-4, IL-5, IL-6, IL-10, IL-12p40, IL-12p70, IL-17, GM-CSF, G-CSF, KC, MIP-1α, RANTES, and eotaxin concentrations were determined by Bio-Plex chemiluminescence assay (Bio-Rad, Hercules, CA).
Cell Preparation and Flow Cytometric Analysis
Hepatic mononuclear cells (MNCs) were isolated using Histopaque density gradient as described previously (Chen et al., 2001). In brief, livers were flushed with ice-cold PBS, crushed using a tissue homogenizer, and passed through sterile mesh (70 μm). Cell suspension washed once with PBS was layered over 15 ml of Histopaque 1083 (Sigma-Aldrich) and centrifuged at 2000 rpm for 15 min at room temperature. Cells at the interface were transferred to a fresh tube and washed twice with PBS. Contaminating red blood cells were lysed using RBC-lysis solution (Sigma-Alrich). For fluorescence-activated cell sorting analysis, cells were blocked using mouse Fc-block (anti-CD16/CD32) and stained for various cell surface markers using fluorescently labeled mAbs (10 μg/ml, in PBS containing 2% fetal bovine serum). After washing, stained cells were analyzed in a flow cytometer (FC500; Beckman Coulter, Fullerton, CA). Only live cells were counted by setting gates on forward and side scatters to exclude debris and dead cells. Isotype antibody-treated cells served as staining controls. Data obtained were analyzed in Cytomics CXP software (Beckman Coulter).
Preparation of ConA Supernatant
Murine splenocytes (5 × 106/well) were seeded onto a 24-well plate and cultured with complete RPMI-1640 medium containing 4 μg/ml ConA. After 72 h, the culture supernatant was harvested, filtered, aliquoted, and stored at −20°C until use.
Detection of Apoptosis by TUNEL Assay
Cells were fixed at room temperature with 4% paraformaldehyde for 30 min, and then they were stained for apoptosis using the TUNEL assay (Roche Diagnostics, Indianapolis, IN) as described previously (Kamath et al., 1997). In brief, the cells were permeabilized on ice for 2 min with 100 μl of 0.1% Triton X-100 in 0.1% sodium citrate, washed, and incubated for 1 h at 37°C with 25 μl of TUNEL reaction mixture. The cells were analyzed in a flow cytometer.
Cell Depletion
CD25+ T cells were depleted using anti-CD25 (IL-2Rα) mAb (clone PC61, rat IgG1; Cedarlane Laboratories) as described previously (Haeryfar et al., 2005). To deplete CD25+ cells, one dose (500 μg) of the ascites was injected intraperitoneally, 12 h before ConA administration. This procedure resulted in >96% decrease in CD25+ cells as determined by flow cytometry using fluorescein isothiocyanate-conjugated anti-CD25 mAb (BD Biosciences).
Reverse Transcription-PCR for Foxp3
This was performed by standard protocol. In brief, total RNA was prepared using the total RNeasy kit (QIAGEN, Valencia, CA), and cDNA was prepared using random hexamer primers (Invitrogen). Foxp3 message was determined by PCR using gene-specific primers (forward, 5′-GGG GAA GCC ATG GCA ATA GTT-3′ and reverse, 5′-TGA AGT AGG CGA ACA TGC GAG TAA-3′). Amplified products were visualized by electrophoresis using 1.5% agarose gels containing ethidium bromide.
Statistical Analysis
Data are expressed as mean ± S.E.M. Student’s t test was used to compare two data sets, and p<0.05 was considered statistically significant.
Results
THC Ameliorates ConA-Induced Liver Injury
THC has been shown to possess significant immunomodulatory and anti-inflammatory properties. To see whether THC treatment prevents ConA-induced liver injury, various doses of THC were administered intraperitoneally into ConA-injected mice. Mice injected with ConA + vehicle developed acute hepatitis as indicated by elevated serum AST and ALT levels at 16 h compared with vehicle alone (Fig. 1A). THC administration resulted in a THC dose-dependent decrease in ConA-induced AST and ALT levels. Although 20 mg/kg THC had a significant effect, the dose of 50 mg/kg was most effective. We used this dose of THC and determined AST and ALT levels at various times after ConA challenge (Fig. 1B). As can be seen, liver enzyme levels peaked at approximately 12 h, and then they started to subside by 24 h in ConA-injected mice. Mice treated with 50 mg/kg b.wt. THC, immediately after ConA challenge, showed significant decrease in AST levels at 6, 12, and 24 h and ALT levels at 12 and 24 h after treatment. Even when administered 30 min after ConA injection, THC significantly reduced the serum AST levels (data not shown). It is important to note that mice injected with THC alone did not develop hepatitis, and they showed AST and ALT levels in the serum similar to vehicle-injected group.
Fig. 1.
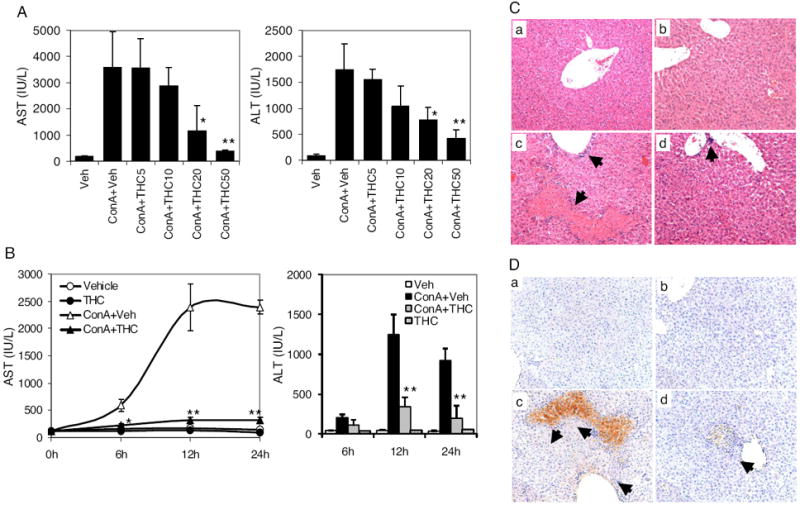
THC ameliorates ConA-induced hepatitis in mice. A, C57BL/6 mice (six/group) were administered with vehicle, ConA + vehicle or ConA + 5, 10, 20, or 50 mg/kg b.wt. THC. Serum was collected 16 h after ConA injection. AST and ALT were determined by spectrophotometric method. B, mice were injected with vehicle, ConA + vehicle or ConA + 50 mg/kg b.wt. THC or 50 mg/kg THC alone. Sera were collected after various times. AST and ALT levels were determined by spectrophotometric method. C, 48 h after ConA treatment, livers were subjected to histological examination. Representative pictures from H&E-stained sections are shown (10× objective): vehicle (a), THC (50 mg/kg) (b), ConA + vehicle (c), and ConA + THC (50 mg/kg) (d). Damaged areas surrounded by infiltrating leukocytes are indicated by arrows. D, sections were stained for apoptosis by TUNEL (brown) and counterstained with hematoxylene (blue). *, p < 0.05; **, p < 0.01 compared with ConA + vehicle.
Liver histological examination was also used to determine the protective effect of THC in ConA-induced hepatitis. Light microscopy showed normal tissue histology for vehicle-injected mice, but dramatic leukocyte infiltration and massive tissue damage in ConA-injected mice. ConA-injected mice administered with THC showed dramatic decrease in hepatic tissue injury (Fig. 1C). THC alone did not cause any liver damage. TUNEL staining showed massive hepatocyte apoptosis in mice challenged with ConA compared with control group, which was markedly reduced upon THC treatment (Fig. 1D).
THC Protects against ConA-Induced Hepatitis by Suppressing Cytokines and Chemokines
ConA-induced hepatitis is associated with the production of various cytokines. To determine whether THC was acting by modulating cytokine and chemokine levels, we measured various cytokine and chemokines at different times (6, 12, and 24 h). As shown in Fig. 2, ConA challenge lead to elevation of most of the cytokines and chemokines tested compared with vehicle-injected control group, including IL-2, TNF-α, IFN-γ, IL-1α, IL-1β, IL-5, IL-6, IL-10, IL-12, IL-17, GM-CSF, G-CSF, KC, MIP-1α, and RANTES. Administration of THC into ConA-injected mice resulted in significant suppression of most of the inflammatory cytokines and chemokines, except IL-2, IL-4, and IL-5.
Fig. 2.
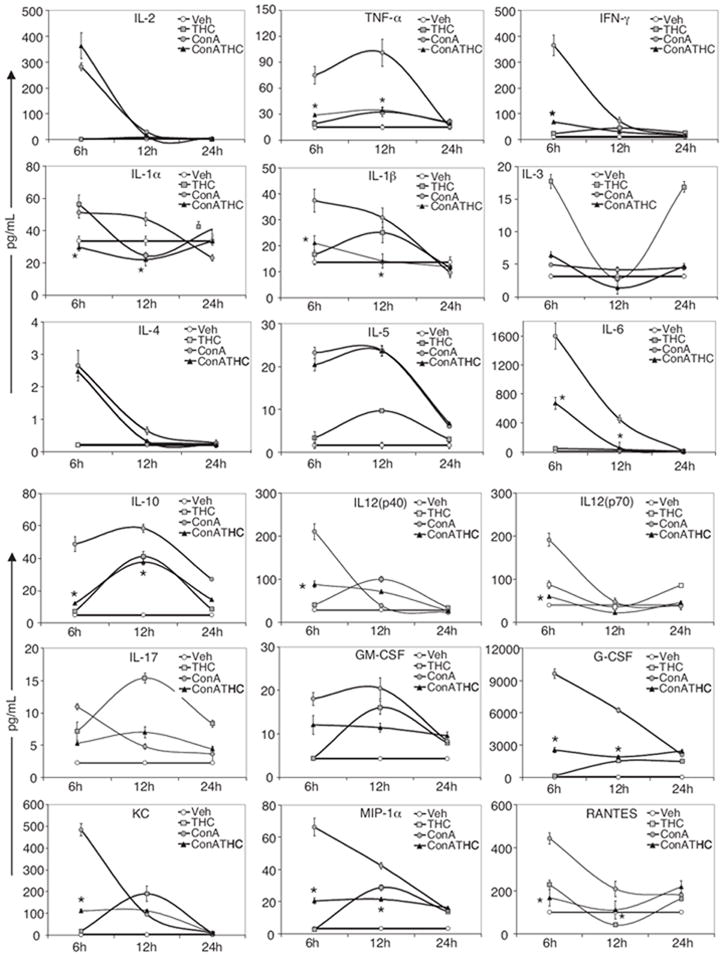
Analysis of cytokines and chemokines in ConA-induced hepatitis. Mice (four/group) were injected with vehicle, THC, ConA + vehicle, or ConA + THC (50 mg/kg). Serum cytokines at 6, 12, and 24 h after ConA injection were analyzed by chemiluminescence method using Bio-Plex system. Significant decrease in the levels of cytokines in ConA + THC group compared with ConA + vehicle group at 6 and 12 h are indicated (*, p < 0.05).
Analysis of T-Cell Subpopulations and T-Cell Apoptosis in Liver
Previous studies from our laboratory have shown that THC can trigger apoptosis in T cells (McKallip et al., 2005). In addition, T cells and NKT cells play critical roles in ConA-induced hepatitis. Thus, we investigated whether THC-induced suppression of hepatic injury results from apoptosis of T or NKT cells in liver. To this end, we isolated intrahepatic mononuclear cells 12 h after ConA challenge and stained them for CD3 and NK1.1 markers. ConA administration resulted in significant increase in percentage and absolute number of T cells in liver compared with vehicle (Fig. 3, A and B). ConA + vehicle-injected mice showed a decrease in the percentage of hepatic NKT cells compared with vehicle alone group (Fig. 3A). These data are consistent with previous reports that ConA administration in vivo leads to early activation and elimination of NKT cells in liver (Takeda et al., 2000). Although there was not much difference in percentages (Fig. 3A), interestingly, we noted that ConA + THC-treated groups showed a significant increase in the absolute numbers of hepatic T and NK cell numbers compared with ConA + vehicle-treated groups (Fig. 3B). We have shown previously that THC triggers apoptosis in T cells. We therefore investigated the levels of apoptosis in hepatic T cells harvested 12 h after ConA challenge in vivo, and we found that significantly higher percentage of apoptotic hepatic T cells were detected in ConA + THC group (58%) compared with ConA + vehicle-injected mice (34.5%) (Fig. 3C). That ConA + THC-treated mice showed increased levels of apoptosis in T cells while demonstrating an increase in their numbers in the liver suggested that additional mechanisms, such as increased migration of cells into the liver, may play a role in ConA + THC-treated groups or that certain subsets of T cells may be more susceptible to THC-induced apoptosis than others.
Fig. 3.
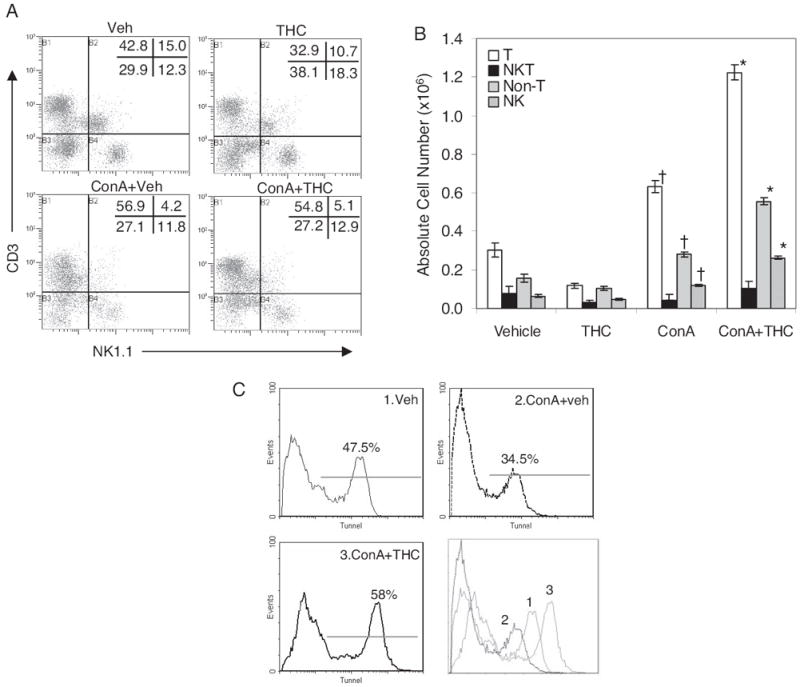
Enumeration of lymphocyte subpopulations in the liver of ConA injected mice. Mice (four/group) were injected with vehicle, THC, ConA + vehicle, or ConA + THC (50 mg/kg). After 12 h, hepatic MNCs were harvested, and cells were stained for various markers and analyzed by fluorescence-activated cell sorting. A, representative double-stain flow profiles showing percentages of each cell population. B, absolute cell numbers of various lymphocyte subpopulations were calculated by taking into consideration total number of liver MNCs isolated per mouse and percentage of cells as shown in A (†, p < 0.05 compared with vehicle; *, p < 0.05 compared with ConA). C, detection of apoptosis in liver MNCs using TUNEL assay. MNCs isolated 12 h after ConA injection were analyzed by TUNEL for apoptotic cells by flow cytometry. Histograms represent TUNEL staining on CD3+ cells. The last panel shows an overlay of three histograms.
Similar to immune cells, hepatocytes are also known to express cannabinoid receptors (Biswas et al., 2003; Osei-Hyiaman et al., 2005). To investigate whether the effect of THC on hepatocytes played a role in ConA-induced hepatitis, we cultured normal mouse hepatocyte cells (BNL.CL.2) with various concentrations of THC for 24 h, and then we determined the percentage of viable cells by trypan blue exclusion. We also measured cells for apoptosis by TUNEL staining and flow cytometry. Mouse splenocytes served as controls. Splenocytes were susceptible to THC-induced apoptosis as indicated by dose-dependent decrease in viable cell number and increase in the percentage of apoptotic cells (Fig. 4, B and C). However, BNL.CL.2 cells were unresponsive to similar concentrations of THC (Fig. 4, A and C) indicating that unlike lymphocytes, murine hepatocytes may be resistant to THC-induced apoptosis. These studies also ruled out the possible direct toxic effects of THC on hepatocytes. Next, we tested whether THC could protect hepatocytes against apoptosis induced by culture supernatant from ConA-activated splenocytes (CS) in vitro. CS induced significant apoptosis in hepatocytes compared with medium control (Fig. 4D). However, the percentage of cells undergoing apoptosis in response to CS was decreased in the presence of THC in a dose-dependent manner (Fig. 4D). These data suggested that THC may afford protection to the hepatocytes from apoptosis induced by activated immune cell-derived factors.
Fig. 4.
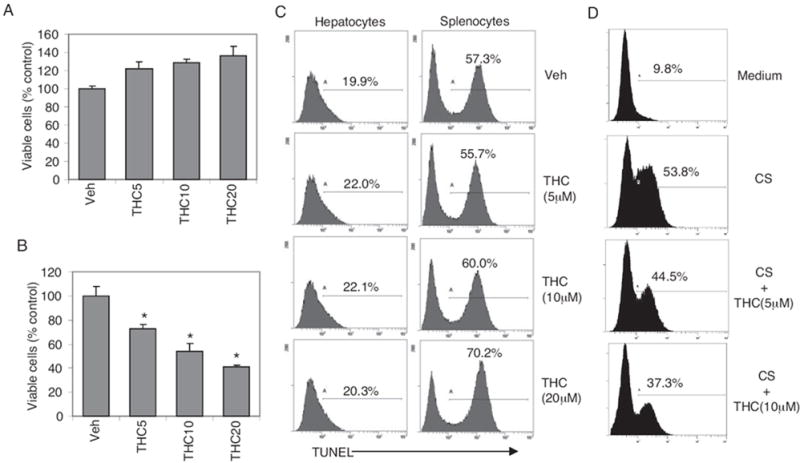
Murine hepatocyte cells are resistant to THC-induced apoptosis. Normal murine hepatocyte cells (BNL.CL.2) (A) and splenocytes (B) were cultured with various doses of THC in complete medium containing 5% fetal bovine serum for 24 h, and percentage of viable cells was determined by trypan blue exclusion (*, p < 0.05). C, cells were stained by TUNEL for apoptosis and analyzed by flow cytometry. Numbers on histograms indicate percentage of apoptotic cells. D, THC protects hepatocyte cells against apoptosis. BNL.CL.2 cells were preincubated with various doses of THC for 1 h. ConA supernatant [CS; 20% (v/v)] was added, and cells were further cultured for 24 h. Apoptosis was analyzed by TUNEL staining and flow cytometry. Percentage of TUNEL-positive cells is indicated on each histogram.
THC Treatment Increases CD4+CD25+ Cells in Liver in ConA-Induced Hepatitis
Although the number of T and NKT cells was significantly increased in the liver upon THC administration in ConA-injected mice, they seemed to be functionally suppressed based on the cytokine data. One possibility was that THC treatment in ConA-injected mice triggered increased levels of CD4+CD25+ Tregs in the liver that were responsible for decreased hepatitis. To this end, we investigated the levels of these cells in liver. ConA + vehicle injection resulted in significant increase in the percentage of CD4+CD25+ cell population in the liver over vehicle-injected controls. In addition, ConA + THC-treated mice did not show any significant increase in the percentage of CD4+CD25+ cells. However, when we enumerated the total number of CD4+CD25+ cells, we noted that there was a significant increase in ConA + THC-treated groups compared with ConA + vehicle-treated mice (Fig. 5, A and B).
Fig. 5.
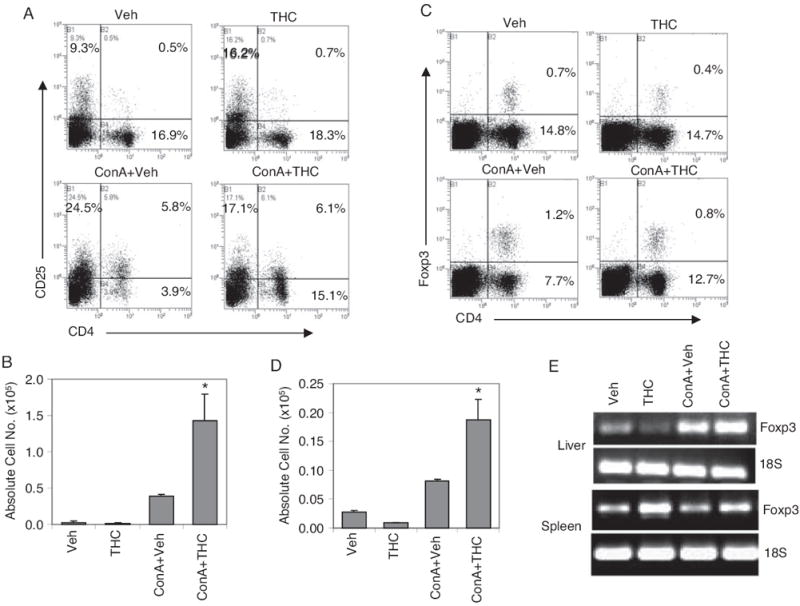
Analysis of regulatory T cells in liver. Hepatitis was induced in mice using ConA as described in Fig. 1. Twelve hours after ConA injection, liver MNCs were analyzed for CD4+CD25+ and CD4+Foxp3+ T cells by double staining. Representative flow profiles with percentage of cell population for each treatment group are shown (A and C) along with absolute cell numbers (B and D). *, p < 0.05 compared with ConA + vehicle. E, 4 h after ConA injection, spleens and livers were collected from each group. cDNA was synthesized from total mRNA, and PCR was carried out using gene-specific primers for Foxp3 and 18S genes.
THC Treatment Increases Foxp3+ Regulatory T Cells in Liver in ConA-Induced Hepatitis
In mice injected with ConA, the CD4+CD25+ population includes both effector and regulatory T-cell populations. foxp3 is the intracellular marker unique to regulatory T cells (Fontenot et al., 2003). Therefore, to further confirm the role of regulatory T cells in THC-induced immune suppression in ConA-induced hepatitis, we looked at CD4+Foxp3+ cells in liver. ConA + THC-injected mice showed a slight decrease in the percentage of CD4+Foxp3+ regulatory T cells compared with ConA + vehicle-injected mice (Fig. 5C). However, they showed a significant increase in the absolute number of CD4+Foxp3+ regulatory T cells (Fig. 5D) as a result of an increase in the absolute number of T cells seen after ConA + THC treatment (Fig. 3B). It should be noted that ConA-injected mice showed a significant increase regulatory T cells over vehicle controls. This is in accordance with recent reports that T regulatory cells play a crucial role in natural tolerance in ConA-induced hepatitis (Erhardt et al., 2007). Nonetheless, we observed that THC treatment further significantly enhanced the number of T regulatory cells in this model.
To further corroborate the role of Tregs, we also analyzed Foxp3 mRNA expression by reverse transcription-PCR in splenocytes and liver cells obtained 4 h after ConA challenge in vivo (Fig. 5E). ConA + THC-treated mice showed significantly higher levels of Foxp3 mRNA in the liver and spleen, compared with ConA + vehicle-treated groups. Mice treated with THC alone showed slight decrease in Foxp3 mRNA in the liver and an increase in spleen.
CD25+ Cells Are Required for THC-Mediated Suppression of ConA-Induced Hepatitis
To further understand the crucial nature of regulatory T cells in ConA-induced hepatitis and THC-mediated suppression, we used anti-mouse CD25 mAbs. We noted that these antibodies depleted CD25+ cells very effectively (Fig. 6B). Mice pretreated with anti-CD25 and injected with ConA showed significantly higher serum AST levels compared with mice pretreated with control IgG (Fig. 6A). These data suggested that in ConA-induced hepatitis, regulatory T cells do play a critical role in trying to reduce the severity of hepatitis. In these studies, mice receiving anti-CD25 alone showed normal AST levels. In the ConA + THC group, preinjection with anti-CD25 significantly reversed the modulatory effect of THC as opposed to control IgG pretreatment, indicating that CD25+ regulatory cells are essential for THC-mediated suppression of hepatitis.
Fig. 6.
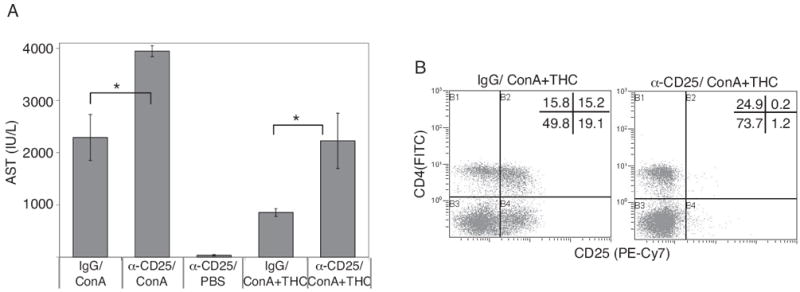
Role of CD4+CD25+ T cells in the pathogenesis of ConA-induced hepatitis and suppression by THC. BL/6 mice (six/group) were injected with isotype control antibody or anti-CD25 mAbs. After 12 h, animals received ConA + vehicle or ConA + THC (50 mg/kg). Plasma AST levels were determined 12 h after ConA injection (A). Control mice received isotype IgG alone. *, p < 0.05 between groups as indicated. B, representative flow profiles showing depletion of CD25+ cells in ConA + THC group.
Modulatory Effect of THC in ConA-Induced Hepatitis Is Mediated through Cannabinoid Receptors
THC is known to act through cannabinoid receptors CB1 and CB2. To delineate the role of cannabinoid receptors involved in the modulation of ConA-induced hepatitis by THC, we used CB1-and CB2-selective agonists. It is noteworthy that CB1 (ACEA) and CB2 (JWH-133)-selective agonists independently or in combination did not decrease AST levels in ConA-induced hepatic injury as shown by serum AST levels (Fig. 7, A and B). In fact, ConA + ACEA, at higher concentrations (20 mg/kg), caused an increase in AST levels compared with ConA + vehicle groups. It should be noted that these cannabinoid receptor agonists by themselves were not hepatotoxic.
Fig. 7.
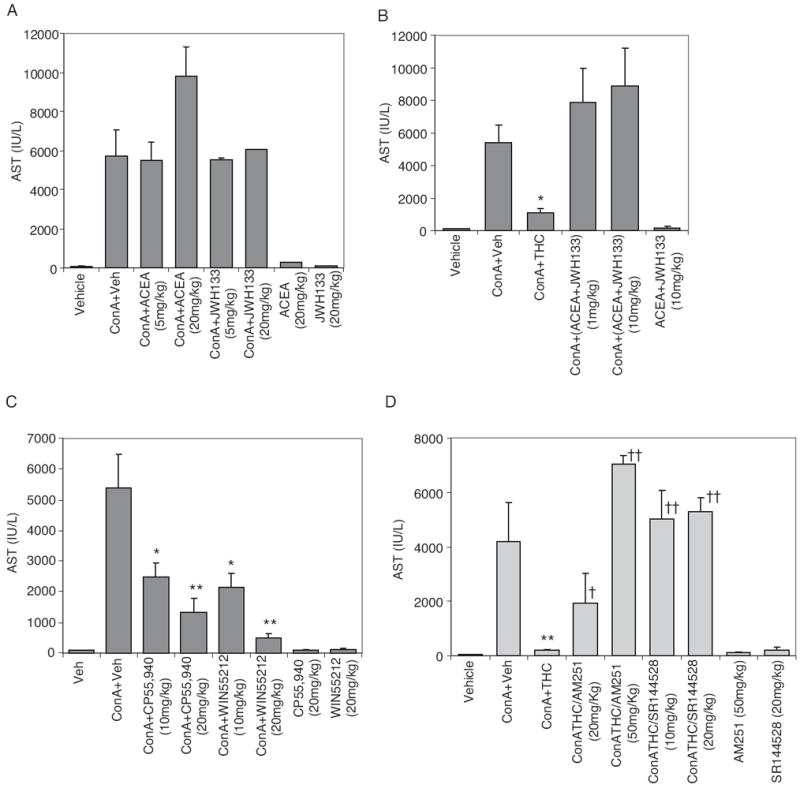
Effect of CB1 or CB2 or mixed agonists on ConA-induced AST levels. A, mice (four/group) were injected with vehicle or ConA. ConA-injected mice were treated with vehicle, CB1 (ACEA), or CB2 (JWH-133) agonists (A) or both ACEA and JWH-133 (B) or CB1/CB2 mixed agonists CP55,940 and WIN55212 (C), with doses as indicated. Control groups received agonists alone as indicated. D, mice received various doses of select CB1 (AM251) or CB2 (SR144528) antagonist 1 h before injecting ConA and THC (50 mg/kg). Serum AST levels in all cases were determined 12 h after ConA injection. (*, p < 0.05; **, p < 0.01 compared with ConA + vehicle; †, p < 0.05; ††, p < 0.01 compared with ConA + THC).
We next tested CB1/CB2 mixed agonists (CP55,940 and WIN55212), which bind to both CB1 and CB2 receptors, similar to THC. We found that these compounds were very effective in suppressing ConA-induced hepatitis in a dose-dependent manner (Fig. 7C). Next, we addressed whether THC acts through CB1 or CB2 receptors. To this end, we pretreated mice with CB1 (AM251) or CB2 antagonists (SR144528) before injecting ConA + THC, and we determined whether this would reverse the THC-mediated suppression of hepatitis (Fig. 7D). It was noted that blocking either one of the receptors could reverse the THC-mediated decrease in AST levels in ConA-treated mice. In addition, control groups receiving antagonists alone showed normal AST levels, indicating that these compounds were not toxic to the liver at the indicated doses. These data demonstrate that immunomodulatory function of THC in ConA-induced hepatitis is mediated by signaling through both CB1 and CB2 receptors, and blocking either of these receptors was sufficient to reverse the effects of THC.
Endocannabinoid Anandamide Attenuates ConA-Induced Hepatitis
Anandamide an endocannabinoid acts as a partial agonist toward CB1 and CB2 receptors (Devane et al., 1992). To see whether anandamide can attenuate hepatitis, we administered exogenous anandamide into mice injected with ConA. ConA + anandamide treatment resulted in decreased AST and ALT levels (Fig. 8A) and less liver damage (Fig. 8B) compared with ConA + vehicle-injected mice, suggesting that anandamide can also effectively block ConA-induced hepatitis similar to THC. We also analyzed cytokine and chemokine levels 12 h after anandamide treatment in ConA-injected mice, and we observed that there was a significant decrease in inflammatory cytokines TNF-α, IL-1β, IL-6, IL-9, IL-12, and IL-17, and in chemokines KC, monocyte chemoattractant protein-1, and eotaxin (Fig. 8C). It is noteworthy that anandamide treatment in ConA-injected mice, similar to THC treatment, failed to decrease IL-2 levels compared with ConA + vehicle-injected mice. We also tested whether the suppressive effect of anandamide is mediated by cannabinoid receptors using CB1- and CB2-specific antagonists. Blocking of CB1 or CB2 receptors by pretreating mice with antagonists AM251 or AM630, respectively, resulted in abrogation of anandamide-mediated suppression of AST levels induced by ConA (Fig. 8D), indicating that anandamide is acting in a CB1- and CB2-dependent manner.
Fig. 8.
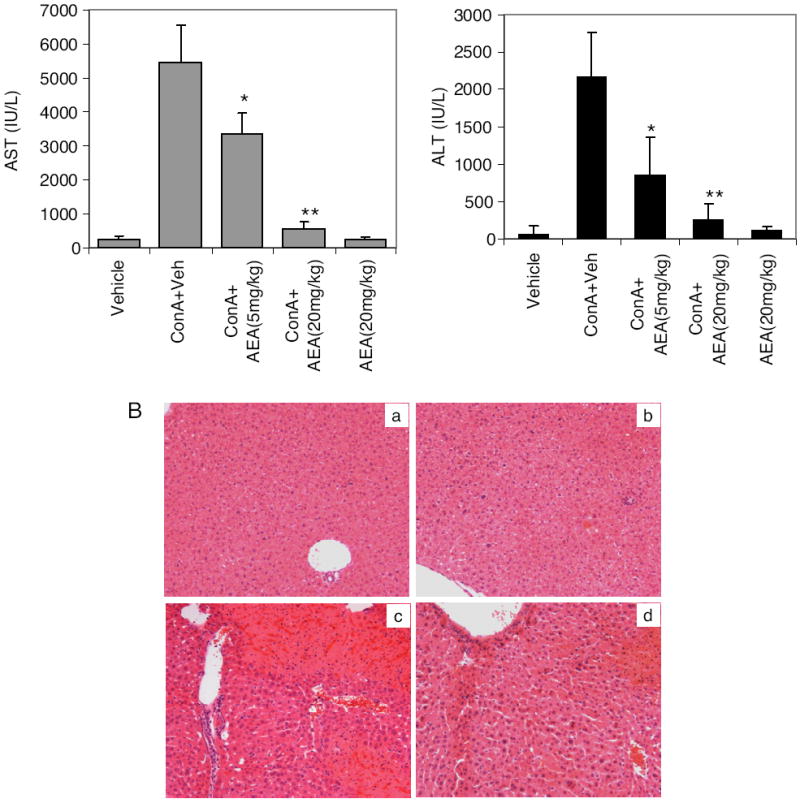
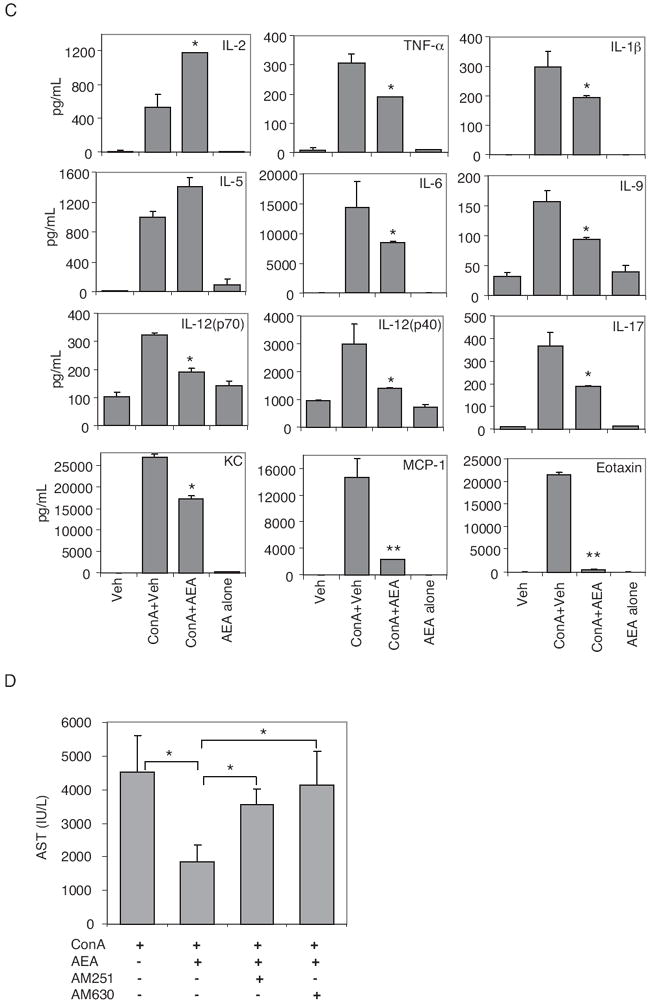
Effect of endocannabinoid AEA on ConA-induced hepatitis. Mice (six/group) were injected with vehicle, AEA alone, ConA + vehicle, or ConA + AEA. A, serum AST and ALT levels determined 12 h after ConA challenge. B, liver histology, H&E-stained sections 48 h after ConA challenge: vehicle (a) AEA (20 mg/kg) (b), ConA + vehicle (c), and ConA + AEA (20 mg/kg) (d). C, serum cytokines and chemokines were determined 12 h after ConA challenge by Bio-Plex method. *, p < 0.05; **, p < 0.01 compared with ConA + vehicle. D, mice received 5 mg/kg b.wt. CB1 (AM251) or CB2 (AM630) antagonists 1 h before injecting ConA and AEA (10 mg/kg). Serum AST levels were determined 12 h after ConA injection. *, p < 0.05 as denoted.
Absence or Inhibition of FAAH Makes Mice Resistant to ConA-Induced Hepatitis
Endogenous cannabinoids such as anandamide are rapidly hydrolyzed by FAAH (Cravatt et al., 1996). Thus, inhibition of FAAH is known to raise anandamide levels (Chang et al., 2006). In addition, FAAH-KO mice are known to have increased levels of endocannabinoids, including anandamide (Cravatt et al., 2001). We therefore investigated whether FAAH-KO mice are more resistant to ConA-induced hepatitis. To test this, we injected wild-type and FAAH-deficient mice with ConA, and we measured AST levels in the serum after 12 h and we also analyzed liver tissue injury by histology after 24 h. As shown in Fig. 9A, FAAH-deficient mice showed significantly reduced AST levels compared with wild-type mice challenged with ConA. Moreover, FAAH-deficient mice showed less severe liver tissue damage and less leukocyte infiltration upon ConA challenge (Fig. 9B). We also tested the effect of pharmacological inhibition of FAAH using inhibitors MAFP or URB532. URB532 has been shown to be a very potent and selective inhibitor of FAAH (Kathuria et al., 2003). Administration of MAFP or URB532 before ConA challenge in wild-type mice resulted in significant decrease in AST levels (Fig. 9C). These data suggest that deficiency or pharmacological inhibition of FAAH imparts mice with some level of resistance to the development of ConA-induced hepatitis.
Fig. 9.
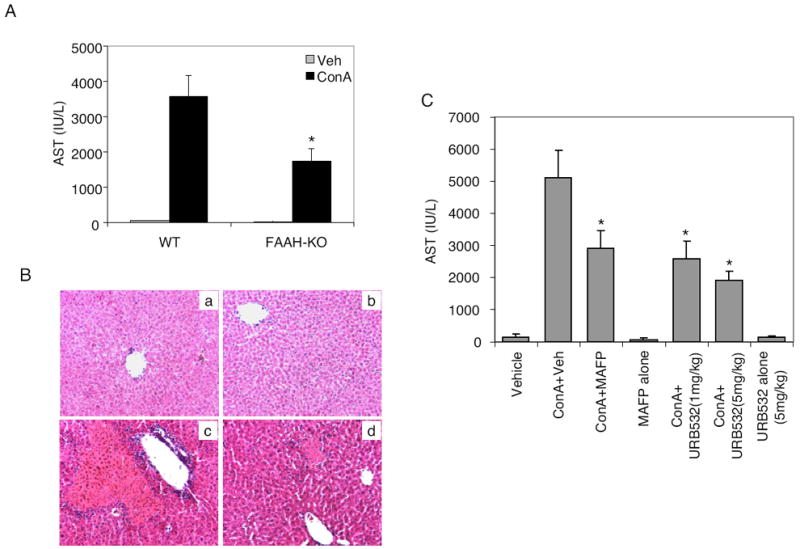
Effect of deficiency or inhibition of FAAH on ConA-mediated hepatitis. Wild-type (BL/6) and FAAH-KO mice (four/group) were injected with vehicle or ConA. A, serum AST levels were analyzed spectrophotometrically 12 h after ConA challenge. *, p < 0.05 compared with wild-type (WT) mice injected with ConA. B, 24 h after ConA injection, livers were harvested and H&E-stained sections were analyzed for histological changes: WT-vehicle (a), FAAH-KO-vehicle (b), WT-ConA (c), and FAAH-KO-ConA (d). C, Wild-type mice were injected with ConA and treated with vehicle or FAAH inhibitors MAFP (10 mg/kg b.wt.) and URB532. AST levels were determined 12 h after ConA injection. Controls injected with vehicle or inhibitors alone are also shown. *, p < 0.05 compared with ConA + vehicle.
Discussion
In vivo administration of THC alleviated ConA-induced hepatitis as indicated by decreased transaminase levels and markedly attenuated inflammatory lesions in liver. THC at 20 and 50 mg/kg b.wt. could significantly suppress AST/ALT levels induced by ConA. A chronic study by the National Toxicology Program (Chan et al., 1996) concluded that a daily dose of up to 150 mg/kg THC for 13 weeks in mice did not produce significant adverse effects. Based on body surface area normalization (Center for Drug Evaluation and Research, 2002), 20- and 50-mg/kg THC doses convert to 60 and 150 mg/m2, respectively. THC may cause central side effects at these doses in humans. However, this is within the range of synthetic THC recommended for clinical use. For example, for dronabinol (Marinol) used as an antiemetic during chemotherapy, 15 mg/m2 is recommended for up to six doses per day, which equates to 90 mg/m2/day. Unlike the prolonged use of dronabinol during chemotherapy, we noted that in our study, a single dose of THC was effective, suggesting that psychotropic effects may not pose a problem if a similar regimen is used in humans to treat autoimmune hepatitis.
TNF-α, IFN-γ and IL-2 play crucial roles in ConA-induced hepatitis (Gantner et al., 1995; Tagawa et al., 1997). We noted that THC could suppress TNF-α and IFN-γ in ConA-injected mice to almost basal levels. IFN-γ-inducing cytokine IL-12 was also significantly decreased after THC treatment. A recent study showed that IL-6 makes CD25+CD4+ effector T cells resistant to the suppression by Tregs (Pasare and Medzhitov, 2003). Decreased IL-6 levels observed in ConA + THC-injected mice may be contributing to suppression of effector T-cell function by Tregs. We also observed that chemokines GM-CSF, G-CSF, KC, MIP-1α, and RANTES that were elevated after ConA challenge, were significantly suppressed upon THC treatment. This is especially important because chemotaxis and activation of macrophages and eosinophils contribute to ConA-induced hepatitis (Schümann et al., 2000; Louis et al., 2002; Bonder et al., 2004).
Previous studies showed that activated T and NKT cells are increased in liver after ConA challenge. However, these cells rapidly undergo apoptosis, which may be crucial for mice to recover from ConA-induced hepatitis (Russell, 1995). We have noted previously that THC induces apoptosis in thymic and splenic T cells (McKallip et al., 2002). In the current study, the frequency of hepatic T lymphocytes undergoing apoptosis was increased in ConA-injected mice upon THC treatment. Nonetheless, absolute numbers of hepatic T cells in THC-treated hepatitis-induced mice were significantly higher. This may result from induction of Tregs as shown, as well as possible increased migration of T cells into the liver. It is interesting to note that IL-2 was not decreased upon THC treatment, and it may be contributing to proliferation of T cells. However, T cells were functionally suppressed by THC as indicated by decreased serum cytokine levels and liver injury. This could be due to direct suppressive effect of THC on activated T cells as well as increase in CD4+Foxp3+ Tregs. IL-2 is an essential growth and survival factor for Tregs, and it is required for their function (Furtado et al., 2002). Sufficient levels of IL-2 observed in ConA-injected mice treated with THC or anandamide could contribute to increased Treg function, resulting in significant suppression of inflammatory cytokines. The crucial role of Tregs was further confirmed by THC not being able to prevent hepatitis in CD25+ Treg-depleted mice. It is important to note that CD25+ cell-depleted mice showed increased AST levels after ConA injection compared with naive mice. One possible explanation for this would be that depletion of Tregs resulted in uncontrolled hepatotoxicity upon polyclonal activation of T cells and other inflammatory cells by ConA. This supports the recent observation that Tregs mediate tolerance in ConA-induced hepatitis model (Erhardt et al., 2007). Previous studies have shown that THC suppresses cytokine secretion by its direct action on lymphocytes (Blanchard et al., 1986; Klein et al., 2000). In the current model, THC may be acting directly on T cells and also by enhancing Treg function in the presence of ConA to effect cytokine suppression and disease attenuation.
There is growing interest in recent years to target cannabinoid receptors for treating liver diseases (Mallat and Lotersztajn, 2008; Pacher and Gao, 2008). In the current study, CB1 or CB2 activation alone had no anti-inflammatory effect on hepatitis. However, cannabinoids that bind to both CB1 and CB2 receptors (THC, CP55,940, WIN55212, and anandamide) effectively attenuated hepatitis. That CB1/CB2 mixed agonists could suppress the disease but not the coadministered CB1 and CB2 agonists indicates that both the cannabinoid receptors need to be activated simultaneously to produce the observed effect and that the different pharmacokinetics of the two coadministered agonists may not allow this to happen. Signaling through both the receptors is important because blocking either CB1 or CB2 could reverse the effect of THC. Activation through CB1 per se worsened the effect of ConA, which is in agreement with previous studies showing CB1 may contribute toward liver inflammatory disorders (Teixeira-Clerc et al., 2006). However, activation of both CB1 and CB2 not only prevented the worsening effect of CB1 stimulation alone but also resulted in a strong counteraction of inflammation. The need for simultaneous activation of CB1 and CB2 to see the beneficial effect is consistent with some previous observations (Van Sickle et al., 2005). However, these results are contradictory to previous understanding that anti-inflammatory properties of cannabinoids are mainly mediated by CB2. The finding that hepatocytes (Biswas et al., 2003; Osei-Hyiaman et al., 2005) and dendritic cells (Matias et al., 2002; Do et al., 2004) express CB1 and CB2 receptors may explain the contribution of both receptors toward protection observed in this study.
The endocannabinoid system plays a protective role in various inflammatory diseases, and it is considered an attractive therapeutic target (Pacher et al., 2006). Activation of endocannabinoid system has been shown to attenuate cutaneous hypersensitivity (Karsak et al., 2007). Anandamide could attenuate hepatitis similar to THC in the current study. Anandamide levels are elevated in the absence of FAAH activity (Cravatt et al., 1996). Increased endogenous anandamide levels may be causing decreased hepatic injury upon ConA challenge in FAAH-KO mice and wild-type mice administered with FAAH inhibitors. Anandamide and 2-arachidonoylglycerol induce cell death in hepatic stellate cells (Siegmund et al., 2006, 2007), and increase in hepatic stellate cell apoptosis is associated with the resolution of liver fibrosis (Iredale et al., 1998; Oakley et al., 2005). However, FAAH−/− mice displayed increased hepatocellular injury upon bile duct ligation (Siegmund et al., 2006). In contrast, we observed that in ConA-induced immune-mediated hepatitis FAAH−/− mice were at least partially resistant compared with wild types. Studies on the role of endocannabinoids in liver diseases have so far mainly focused on hepatic fibrosis, a result of chronic liver disease. CB1−/− mice showed decreased fibrogenesis (Teixeira-Clerc et al., 2006), whereas CB2−/− mice displayed increased fibrogenesis after CCL4 treatment, a chemical-induced liver injury (Julien et al., 2005). This was attributed to opposing effects of CB1 and CB2 in cell death and proliferation of myofibroblasts (Julien et al., 2005). However, other cell types and mechanisms may also play crucial roles because CB1 and CB2 are highly expressed in other hepatic cell types, and CB1 is expressed on the vascular endothelium and in hepatocytes in the injured liver (Siegmund and Schwabe, 2008). The induction of endocannabinoids and their receptors constitute complementary mechanisms that render the liver responsive toward endocannabinoids in hepatic fibrosis (Siegmund and Schwabe, 2008). In addition, endocannabinoids exert numerous and sometimes opposite effects on target cells. Strong up-regulation of CB1 occurs only in some types of liver injury, whereas CB2 is highly induced in most liver diseases (Siegmund and Schwabe, 2008). The regulation of endocannabinoids and their receptors on immune and hepatic cell types during ConA-induced immune-mediated acute liver injury is not known. In contrast to the studies on liver fibrosis, we find that THC and anandamide act through both CB1 and CB2 receptors to produce the observed effect in ConA-induced hepatitis. The levels of endocannabinoids and related receptors on various cell types and hence their effect, as well as other mechanisms of protection that may be involved in different types of liver injury, may be distinct and are yet to be clearly elucidated. This may explain varying response of anandamide and FAAH−/− mice in different liver disease models. Modulation of immune response by cannabinoids seems to play a crucial role in immune-mediated liver injury.
Taken together, our data suggest that exogenous cannabinoids such as THC upon binding to CB1 and CB2 receptors on immune cells, induce apoptosis in effector T cells, up-regulate Treg function, and suppress inflammatory cytokines there by preventing ConA-induced activated T-cell-mediated liver injury. The observation that the anandamide treatment ameliorates ConA-induced hepatitis, together with FAAH deficiency or inhibition leading to increased resistance to the disease, strongly suggests that the endocannabinoid system serves to attenuate the inflammatory response in ConA-induced acute hepatitis. These findings raise the promising potential of developing novel pharmacological treatments for T-cell-mediated liver diseases.
Acknowledgments
This study was funded by National Institutes of Health grants R01-DA016545, R01-ES09098, R01-AI053703, R01-AI058300, R01-HL058641, and P01-AT003961 (to P.S.N. and M.N.).
ABBREVIATIONS
- AIH
autoimmune hepatitis
- NKT
natural killer T
- TNF
tumor necrosis factor
- IFN
interferon
- mAb
monoclonal antibody
- THC
δ-9-tetrahydrocannabinol
- CB
cannabinoid
- AEA
arachidonoylethanolamide (anandamide)
- ConA
concanavalin A
- JWH-133
1,1-dimethylbutyl-1-deoxy-Δ9-tetrahydrocannabinol
- foxp3
Forkhead helix transcription factor p3
- MAFP
methylarachidonyl fluorophosphate
- ACEA
arachidonyl-2′-chloroethylamide
- CP55,940
(1R,3R,4R)-3-[2-hydroxy-4-(1,1-dimethylheptyl)phenyl]-4-(3-hydroxypropyl)cyclohexan-1-ol
- WIN55212
(R)-(+)-[2,3-dihydro-5-methyl-3-(4-morpholinylmethyl) pyrrolo-[1,2,3-d,e]-1,4-benzoxazin-6-yl]-1-naphthalenyl-methanone
- SR144528
N-[(1S)-endo-1,3,3-trimethyl bicyclo heptan-2-yl]-5-(4-chloro-3-methylphenyl)-1-(4-methylbenzyl)-pyrazole-3-carboxamide)
- URB532
4-benzyloxyphenyl-n-butylcarbamate
- FAAH
fatty acid amide hydrolase
- PBS
phosphate-buffered saline
- DMSO
dimethyl sulfoxide
- AST
aspartate transaminase
- ALT
alanine transaminase
- TUNEL
terminal deoxynucleotidyl transferase dUTP nick-end labeling
- H&E
hematoxylin and eosin
- IL
interleukin
- GM-CSF
granulocyte macrophage–colony-stimulating factor
- G-CSF
granulocyte–colony-stimulating factor
- KC
CXC-chemokine
- MIP
macrophage inflammatory protein
- RANTES
regulated on activation normal T cell expressed and secreted
- MNC
mononuclear cell
- PKC
protein kinase C
- CS
ConA-activated splenocyte
- Treg
regulatory T cell
- KO
knockout
- WT
wild type
- AM251
N-(piperidin-1-yl)-1-(2,4-dichlorophenyl)-5-(4-iodophenyl)-4-methyl-1H-pyrazole-3-carboxamide
- AM630
iodopravadoline
References
- Biswas KK, Sarker KP, Abeyama K, Kawahara K, Iino S, Otsubo Y, Saigo K, Izumi H, Hashiguchi T, Yamakuchi M, et al. Membrane cholesterol but not putative receptors mediates anandamide-induced hepatocyte apoptosis. Hepatology. 2003;38:1167–1177. doi: 10.1053/jhep.2003.50459. [DOI] [PubMed] [Google Scholar]
- Blanchard DK, Newton C, Klein TW, Stewart WE, 2nd, Friedman H. In vitro and in vivo suppressive effects of delta-9-tetrahydrocannabinol on interferon production by murine spleen cells. Int J Immunopharmacol. 1986;8:819–824. doi: 10.1016/0192-0561(86)90020-2. [DOI] [PubMed] [Google Scholar]
- Bonder CS, Ajuebor MN, Zbytnuik LD, Kubes P, Swain MG. Essential role for neutrophil recruitment to the liver in concanavalin A-induced hepatitis. J Immunol. 2004;172:45–53. doi: 10.4049/jimmunol.172.1.45. [DOI] [PubMed] [Google Scholar]
- Center for Drug Evaluation and Research. Estimating the safe starting dose in clinical trials for therapeutics in adult healthy volunteers. Food and Drug Administration; Rockville, MD: 2002. Available at: http://www.fda.gov/cber/gdlns/dose.htm. [Google Scholar]
- Chan PC, Sills RC, Braun AG, Haseman JK, Bucher JR. Toxicity and carcinogenicity of delta 9-tetrahydrocannabinol in Fischer rats and B6C3F1 mice. Fundam Appl Toxicol. 1996;30:109–117. doi: 10.1006/faat.1996.0048. [DOI] [PubMed] [Google Scholar]
- Chang L, Luo L, Palmer JA, Sutton S, Wilson SJ, Barbier AJ, Breitenbucher JG, Chaplan SR, Webb M. Inhibition of fatty acid amide hydrolase produces analgesia by multiple mechanisms. Br J Pharmacol. 2006;148:102–113. doi: 10.1038/sj.bjp.0706699. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chen D, McKallip RJ, Zeytun A, Do Y, Lombard C, Robertson JL, Mak TW, Nagarkatti PS, Nagarkatti M. CD44-deficient mice exhibit enhanced hepatitis after concanavalin A injection: evidence for involvement of CD44 in activation-induced cell death. J Immunol. 2001;166:5889–5897. doi: 10.4049/jimmunol.166.10.5889. [DOI] [PubMed] [Google Scholar]
- Chisari FV, Ferrari C. Hepatitis B virus immunopathogenesis. Annu Rev Immunol. 1995;13:29–60. doi: 10.1146/annurev.iy.13.040195.000333. [DOI] [PubMed] [Google Scholar]
- Cravatt BF, Demarest K, Patricelli MP, Bracey MH, Giang DK, Martin BR, Lichtman AH. Supersensitivity to anandamide and enhanced endogenous cannabinoid signaling in mice lacking fatty acid amide hydrolase. Proc Natl Acad Sci U S A. 2001;98:9371–9376. doi: 10.1073/pnas.161191698. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cravatt BF, Giang DK, Mayfield SP, Boger DL, Lerner RA, Gilula NB. Molecular characterization of an enzyme that degrades neuromodulatory fatty acid amides. Nature. 1996;384:83–87. doi: 10.1038/384083a0. [DOI] [PubMed] [Google Scholar]
- Croxford JL, Yamamura T. Cannabinoids and the immune system: potential for the treatment of inflammatory diseases? J Neuroimmunol. 2005;166:3–18. doi: 10.1016/j.jneuroim.2005.04.023. [DOI] [PubMed] [Google Scholar]
- Czaja AJ, Freese DK. Diagnosis and treatment of autoimmune hepatitis. Hepatology. 2002;36:479–497. doi: 10.1053/jhep.2002.34944. [DOI] [PubMed] [Google Scholar]
- Devane WA, Hanus L, Breuer A, Pertwee RG, Stevenson LA, Griffin G, Gibson D, Mandelbaum A, Etinger A, Mechoulam R. Isolation and structure of a brain constituent that binds to the cannabinoid receptor. Science. 1992;258:1946–1949. doi: 10.1126/science.1470919. [DOI] [PubMed] [Google Scholar]
- Do Y, McKallip RJ, Nagarkatti M, Nagarkatti PS. Activation through cannabinoid receptors 1 and 2 on dendritic cells triggers NF-kappaB-dependent apoptosis: novel role for endogenous and exogenous cannabinoids in immunoregulation. J Immunol. 2004;173:2373–2382. doi: 10.4049/jimmunol.173.4.2373. [DOI] [PubMed] [Google Scholar]
- Erhardt A, Biburger M, Papadopoulos T, Tiegs G. IL-10, regulatory T cells, and Kupffer cells mediate tolerance in concanavalin A-induced liver injury in mice. Hepatology. 2007;45:475–485. doi: 10.1002/hep.21498. [DOI] [PubMed] [Google Scholar]
- Fontenot JD, Gavin MA, Rudensky AY. Foxp3 programs the development and function of CD4+CD25+ regulatory T cells. Nat Immunol. 2003;4:330–336. doi: 10.1038/ni904. [DOI] [PubMed] [Google Scholar]
- Furtado GC, Curotto de Lafaille MA, Kutchukhidze N, Lafaille JJ. Interleukin 2 signaling is required for CD4(+) regulatory T cell function. J Exp Med. 2002;196:851–857. doi: 10.1084/jem.20020190. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gantner F, Leist M, Lohse AW, Germann PG, Tiegs G. Concanavalin A-induced T-cell-mediated hepatic injury in mice: the role of tumor necrosis factor. Hepatology. 1995;21:190–198. doi: 10.1016/0270-9139(95)90428-x. [DOI] [PubMed] [Google Scholar]
- Haeryfar SM, DiPaolo RJ, Tscharke DC, Bennink JR, Yewdell JW. Regulatory T cells suppress CD8+ T cell responses induced by direct priming and cross-priming and moderate immunodominance disparities. J Immunol. 2005;174:3344–3351. doi: 10.4049/jimmunol.174.6.3344. [DOI] [PubMed] [Google Scholar]
- Iredale JP, Benyon RC, Pickering J, McCullen M, Northrop M, Pawley S, Hovell C, Arthur MJ. Mechanisms of spontaneous resolution of rat liver fibrosis. Hepatic stellate cell apoptosis and reduced hepatic expression of metalloproteinase inhibitors. J Clin Invest. 1998;102:538–549. doi: 10.1172/JCI1018. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Julien B, Grenard P, Teixeira-Clerc F, Van Nhieu JT, Li L, Karsak M, Zimmer A, Mallat A, Lotersztajn S. Antifibrogenic role of the cannabinoid receptor CB2 in the liver. Gastroenterology. 2005;128:742–755. doi: 10.1053/j.gastro.2004.12.050. [DOI] [PubMed] [Google Scholar]
- Kamath AB, Xu H, Nagarkatti PS, Nagarkatti M. Evidence for the induction of apoptosis in thymocytes by 2,3,7,8-tetrachlorodibenzo-p-dioxin in vivo. Toxicol Appl Pharmacol. 1997;142:367–377. doi: 10.1006/taap.1996.8049. [DOI] [PubMed] [Google Scholar]
- Karsak M, Gaffal E, Date R, Wang-Eckhardt L, Rehnelt J, Petrosino S, Starowicz K, Steuder R, Schlicker E, Cravatt B, et al. Attenuation of allergic contact dermatitis through the endocannabinoid system. Science. 2007;316:1494–1497. doi: 10.1126/science.1142265. [DOI] [PubMed] [Google Scholar]
- Kathuria S, Gaetani S, Fegley D, Valino F, Duranti A, Tontini A, Mor M, Tarzia G, La Rana G, Calignano A, et al. Modulation of anxiety through blockade of anandamide hydrolysis. Nat Med. 2003;9:76–81. doi: 10.1038/nm803. [DOI] [PubMed] [Google Scholar]
- Kita H, Mackay IR, Van De Water J, Gershwin ME. The lymphoid liver: considerations on pathways to autoimmune injury. Gastroenterology. 2001;120:1485–1501. doi: 10.1053/gast.2001.22441. [DOI] [PubMed] [Google Scholar]
- Klein TW, Newton CA, Nakachi N, Friedman H. Delta 9-tetrahydrocannabinol treatment suppresses immunity and early IFN-gamma, IL-12, and IL-12 receptor beta 2 responses to Legionella pneumophila infection. J Immunol. 2000;164:6461–6466. doi: 10.4049/jimmunol.164.12.6461. [DOI] [PubMed] [Google Scholar]
- Küsters S, Gantner F, Kunstle G, Tiegs G. Interferon gamma plays a critical role in T cell-dependent liver injury in mice initiated by concanavalin A. Gastroenterology. 1996;111:462–471. doi: 10.1053/gast.1996.v111.pm8690213. [DOI] [PubMed] [Google Scholar]
- Louis H, Le Moine A, Flamand V, Nagy N, Quertinmont E, Paulart F, Abramowicz D, Le Moine O, Goldman M, Deviere J. Critical role of interleukin 5 and eosinophils in concanavalin A-induced hepatitis in mice. Gastroenterology. 2002;122:2001–2010. doi: 10.1053/gast.2002.33620. [DOI] [PubMed] [Google Scholar]
- Mackie K. Cannabinoid receptors as therapeutic targets. Annu Rev Pharmacol Toxicol. 2006;46:101–122. doi: 10.1146/annurev.pharmtox.46.120604.141254. [DOI] [PubMed] [Google Scholar]
- Mallat A, Lotersztajn S. Endocannabinoids and liver disease. I. Endocannabinoids and their receptors in the liver. Am J Physiol Gastrointest Liver Physiol. 2008;294:G9–G12. doi: 10.1152/ajpgi.00467.2007. [DOI] [PubMed] [Google Scholar]
- Matias I, Pochard P, Orlando P, Salzet M, Pestel J, Di Marzo V. Presence and regulation of the endocannabinoid system in human dendritic cells. Eur J Biochem. 2002;269:3771–3778. doi: 10.1046/j.1432-1033.2002.03078.x. [DOI] [PubMed] [Google Scholar]
- McKallip RJ, Lombard C, Martin BR, Nagarkatti M, Nagarkatti PS. Delta(9)-tetrahydrocannabinol-induced apoptosis in the thymus and spleen as a mechanism of immunosuppression in vitro and in vivo. J Pharmacol Exp Ther. 2002;302:451–465. doi: 10.1124/jpet.102.033506. [DOI] [PubMed] [Google Scholar]
- McKallip RJ, Nagarkatti M, Nagarkatti PS. Delta-9-tetrahydrocannabinol enhances breast cancer growth and metastasis by suppression of the antitumor immune response. J Immunol. 2005;174:3281–3289. doi: 10.4049/jimmunol.174.6.3281. [DOI] [PubMed] [Google Scholar]
- Oakley F, Meso M, Iredale JP, Green K, Marek CJ, Zhou X, May MJ, Millward-Sadler H, Wright MC, Mann DA. Inhibition of inhibitor of kappaB kinases stimulates hepatic stellate cell apoptosis and accelerated recovery from rat liver fibrosis. Gastroenterology. 2005;128:108–120. doi: 10.1053/j.gastro.2004.10.003. [DOI] [PubMed] [Google Scholar]
- Osei-Hyiaman D, DePetrillo M, Pacher P, Liu J, Radaeva S, Batkai S, Harvey-White J, Mackie K, Offertaler L, Wang L, et al. Endocannabinoid activation at hepatic CB1 receptors stimulates fatty acid synthesis and contributes to diet-induced obesity. J Clin Invest. 2005;115:1298–1305. doi: 10.1172/JCI23057. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pacher P, Batkai S, Kunos G. The endocannabinoid system as an emerging target of pharmacotherapy. Pharmacol Rev. 2006;58:389–462. doi: 10.1124/pr.58.3.2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pacher P, Gao B. Endocannabinoid effects on immune cells: implications for inflammatory liver diseases. Am J Physiol Gastrointest Liver Physiol. 2008;294:G850–G854. doi: 10.1152/ajpgi.00523.2007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pasare C, Medzhitov R. Toll pathway-dependent blockade of CD4+CD25+ T cell-mediated suppression by dendritic cells. Science. 2003;299:1033–1036. doi: 10.1126/science.1078231. [DOI] [PubMed] [Google Scholar]
- Pertwee RG. Cannabinoids and multiple sclerosis. Pharmacol Ther. 2002;95:165–174. doi: 10.1016/s0163-7258(02)00255-3. [DOI] [PubMed] [Google Scholar]
- Quartilho A, Mata HP, Ibrahim MM, Vanderah TW, Porreca F, Makriyannis A, Malan TP., Jr Inhibition of inflammatory hyperalgesia by activation of peripheral CB2 cannabinoid receptors. Anesthesiology. 2003;99:955–960. doi: 10.1097/00000542-200310000-00031. [DOI] [PubMed] [Google Scholar]
- Russell JH. Activation-induced death of mature T cells in the regulation of immune responses. Curr Opin Immunol. 1995;7:382–388. doi: 10.1016/0952-7915(95)80114-6. [DOI] [PubMed] [Google Scholar]
- Schümann J, Wolf D, Pahl A, Brune K, Papadopoulos T, van Rooijen N, Tiegs G. Importance of Kupffer cells for T-cell-dependent liver injury in mice. Am J Pathol. 2000;157:1671–1683. doi: 10.1016/S0002-9440(10)64804-3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Siegmund SV, Qian T, de Minicis S, Harvey-White J, Kunos G, Vinod KY, Hungund B, Schwabe RF. The endocannabinoid 2-arachidonoyl glycerol induces death of hepatic stellate cells via mitochondrial reactive oxygen species. FASEB J. 2007;21:2798–2806. doi: 10.1096/fj.06-7717com. [DOI] [PubMed] [Google Scholar]
- Siegmund SV, Schwabe RF. Endocannabinoids and Liver Disease. II. Endocannabinoids in the pathogenesis and treatment of liver fibrosis. Am J Physiol Gastrointest Liver Physiol. 2008;294:G357–G362. doi: 10.1152/ajpgi.00456.2007. [DOI] [PubMed] [Google Scholar]
- Siegmund SV, Seki E, Osawa Y, Uchinami H, Cravatt BF, Schwabe RF. Fatty acid amide hydrolase determines anandamide-induced cell death in the liver. J Biol Chem. 2006;281:10431–10438. doi: 10.1074/jbc.M509706200. [DOI] [PubMed] [Google Scholar]
- Steffens S, Veillard NR, Arnaud C, Pelli G, Burger F, Staub C, Karsak M, Zimmer A, Frossard JL, Mach F. Low dose oral cannabinoid therapy reduces progression of atherosclerosis in mice. Nature. 2005;434:782–786. doi: 10.1038/nature03389. [DOI] [PubMed] [Google Scholar]
- Tagawa Y, Sekikawa K, Iwakura Y. Suppression of concanavalin A-induced hepatitis in IFN-gamma(-/-) mice, but not in TNF-alpha(-/-) mice: role for IFN-gamma in activating apoptosis of hepatocytes. J Immunol. 1997;159:1418–1428. [PubMed] [Google Scholar]
- Takeda K, Hayakawa Y, Van Kaer L, Matsuda H, Yagita H, Okumura K. Critical contribution of liver natural killer T cells to a murine model of hepatitis. Proc Natl Acad Sci U S A. 2000;97:5498–5503. doi: 10.1073/pnas.040566697. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Teixeira-Clerc F, Julien B, Grenard P, Tran Van Nhieu J, Deveaux V, Li L, Serriere-Lanneau V, Ledent C, Mallat A, Lotersztajn S. CB1 cannabinoid receptor antagonism: a new strategy for the treatment of liver fibrosis. Nat Med. 2006;12:671–676. doi: 10.1038/nm1421. [DOI] [PubMed] [Google Scholar]
- Tiegs G, Hentschel J, Wendel A. A T cell-dependent experimental liver injury in mice inducible by concanavalin A. J Clin Invest. 1992;90:196–203. doi: 10.1172/JCI115836. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Van Sickle MD, Duncan M, Kingsley PJ, Mouihate A, Urbani P, Mackie K, Stella N, Makriyannis A, Piomelli D, Davison JS, et al. Identification and functional characterization of brainstem cannabinoid CB2 receptors. Science. 2005;310:329–332. doi: 10.1126/science.1115740. [DOI] [PubMed] [Google Scholar]