

Abstract
Ever since Virchow introduced the entity myxoma, abundant myxoid extracellular matrix (ECM) has been recognized in various reactive and neoplastic lesions. Nowadays, the term “myxoid” is commonly used in daily pathological practice. But what do today’s pathologists mean by it, and what does the myxoid ECM tell the pathologist? What is known about the exact composition and function of the myxoid ECM 150 years after Virchow? Here, we give an overview of the composition and constituents of the myxoid ECM as known so far and demonstrate the heterogeneity of the myxoid ECM among different tumors. We discuss the possible role of the predominant constituents of the myxoid ECM and attempt to relate them to differences in clinical behavior. Finally, we will speculate on the potential relevance of this knowledge in daily pathological practice.
Keywords: Extracellular matrix, Glycosaminoglycans, Proteoglycan, Collagen, Myxofibrosarcoma, Intramuscular myxoma
Historical perspective
In his 1858 masterpiece Cellularpathologie, Rudolph Virchow introduced the term “myxoma” to describe a soft tissue tumor, histologically resembling the structure of the umbilical cord (Figs. 1 and 2) [1]. This description of myxoma was adopted in the seventh edition of the Medical Lexicon by Robley Dunglison who remarkably added that “[myxoma] was for the first time described in 1838 by Johannes Müller as Collonema” [2]. Müller used the term collonema (κολλα = glue) for “peculiar gelatinous tumours, consisting of a remarkably soft gelatiniform tissue, which trembles on being touched” [3]. Though this description is applicable to most myxoid tumors, it holds also for many nonmyxoid tumors and it is not particularly clear which tumor type Müller had in mind. Today, it has been generally accepted that it was indeed Virchow who introduced myxoma as an entity. The introduction of this new histological concept of tumors containing myxoid (μύξα = “mucus” and ειδος = “resemblance”) areas soon led to the recognition of new entities, such as myxadenoma, myxochondroma, myxofibroma, and myxoneuroma [2]. The term myxosarcoma, introduced in 1802 by Bryant [4], became reserved for malignant tumors and defined as “a mucous transformation of round-celled sarcoma, malignant, and of large volume, usually attacking the omentum and the skin” [2]. Nowadays, myxoid changes/areas are recognized in both benign and malignant neoplasms (primarily classified as mesenchymal or epithelial) as well as non-neoplastic (reactive) lesions (Fig. 3).
Fig. 1.
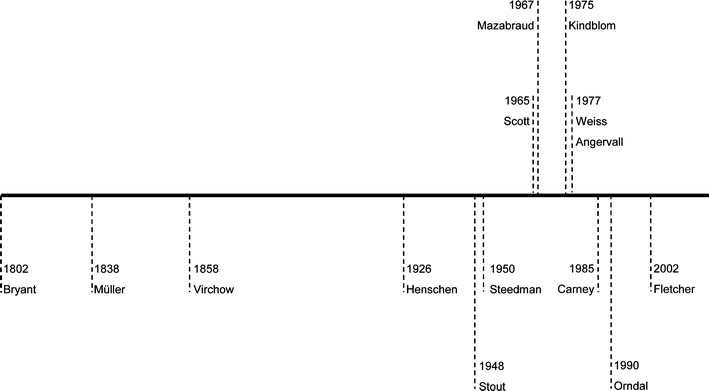
Timetable with key events in studies on myxoid tumors of soft tissue. Though Müller already mentioned tumors with a macroscopically gelatinous appearance in 1838 [3], it was in 1858 when Virchow introduced the term myxoma to describe tumors which morphologically resembled the jelly structure of the umbilical cord [1]. Ever since, the term myxosarcoma, introduced by Bryant in 1802 was reserved for the malignant counterparts [4]. Because of their morphologically overlapping features, both terms were used interchangeably, which was mentioned by Stout in 1948 as unwise, warranting for macroscopical and microscopical criteria for the reliable differential diagnosis between the two entities [5]. The relationship between myxomas and fibrous dysplasia was first described in 1926 by Henschen [90], though it was Mazabraud who proposed it as a syndrome in 1967 [91]. The association of cardiac/cutaneous myxomas, hyperpigmentation of the skin, and endocrine overactivity was only recognized in 1985 by Carney [43]. Progress in the study of the myxoid ECM was made by the invention of the alcian blue staining in 1950 by Steedman [9], and Scott who developed the CEC method to distinguish the different GAGs in 1965 [10]. Based upon this technique, Kindblom showed in 1975 that different bone and soft tissue tumors (including myxoid ones) contained different GAGs [11]. From the late 1980s, it became clear that the ECM is a key player in tumor development and tumor progression, sustained by an exponentially growing number of publications [40]. As myxoid areas were now being recognized as an intrinsic part of a subset of tumors, Weiss and Angervall simultaneously described the myxoid variant of malignant fibrous histiocytoma/myxofibrosarcoma as a distinct entity [92, 93]. Parallel to morphological classification, an increasing number of myxoid tumors showed specific molecular genetics aberrations, such as (activating) mutations and translocations. The concept of malignant progression in myxoid tumors of soft tissue (i.e., myxoid liposarcoma) due to chromosomal instability and subsequent secondary genetic events was described in 1990 by Orndal et al. [94]. Nowadays, classification of myxoid tumors of soft tissues is based upon clinicopathological and molecular/cytogenetic aberrations as published in the 2002 WHO classification [7]
Fig. 2.
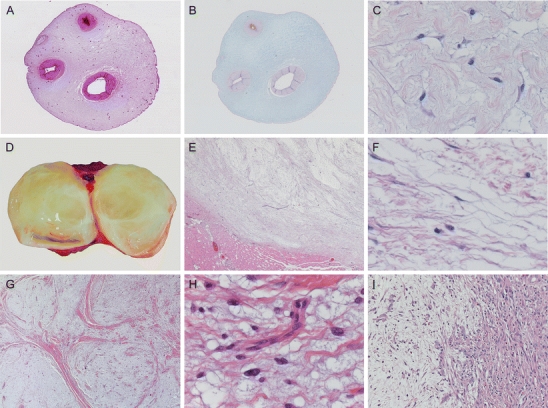
Characteristic macroscopy and histomorphology of the myxoid ECM. Rudolph Virchow introduced the term myxoma for those tumors morphologically resembling Wharton’s jelly of the umbilical cord (a), which contains large amounts of GAGs as detected by alcian blue (b). High-power image of Wharton’s jelly showing abundant myxoid ECM containing fibrillary collagens, interspersed between myofibroblast-like stroma cells (c). Intramuscular myxoma characteristically has a gelatinous appearance on cut surface (d) and is well circumscribed towards its peripheral tissue (e). On higher magnification, it shows the same abundant myxoid ECM as the umbilical cord (c) and no significant atypia of the sparse tumor cells (f). Histological criteria are still a hallmark of diagnosis, showing characteristic lobulated, hypocellular morphology of grade I myxofibrosarcoma at low magnification (g).Curvilinear blood vessels are often seen in grade I myxofibrosarcoma (but are not diagnostic), whereas tumor cells show vesicular, slightly atypical nuclei compared to intramuscular myxoma (h). Another hallmark of myxofibrosarcoma is areas with abrupt transition of grade (i) which was already mentioned by Mentzel et al. [95]
Fig. 3.
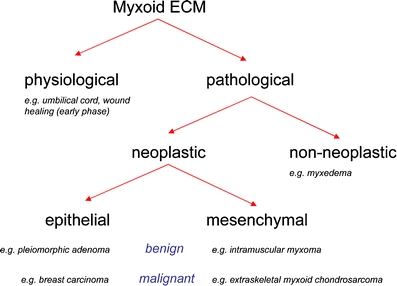
Myxoid ECM is a ubiquitously histological feature in physiological and pathological conditions. Myxoid ECM is a morphological feature in physiological and pathological conditions, such as in myxedema due to increased production of HA. Myxoid areas/changes are also commonly present in tumors (both of epithelial and mesenchymal origin). In epithelial tumors, myxoid changes are often a secondary phenomenon, whereas in mesenchymal tumors, they are more frequently an intrinsic part of the tumor entity. This group of so-called myxoid tumors of soft tissues contains an increasing number of entities (e.g., myxofibrosarcoma, formerly called myxoid variant of malignant fibrous histiocytoma), sometimes sustained by specific distinct molecular/cytogenetic aberrations (e.g., myxoid liposarcoma)
In his first description, Virchow had already recognized the recurrent nature of some myxomatous tumors [1] and it became clear that it was difficult to predict the exact clinical behavior of these different tumors based on their myxoid morphology alone. Subsequently, the terms myxoma and myxosarcoma were used interchangeably till Arthur Stout recognized this as unwise [5] because “myxomas do not metastasize and there is no way to anticipate differences in their growth energy from their histopathology.” Later studies confirmed the distinction between both entities on the basis of macroscopical and microscopical features (necrosis, nuclear atypia, and mitotic figures; Fig. 2) [6]. Today, myxoid tumors of soft tissue are classified according to the World Health Organization (WHO) formulation based on clinicopathological criteria and specific molecular/cytogenetic aberrations (Table 1) [7, 8]. So what is left of the term “myxoid” 150 years after Virchow? What do today’s pathologists mean by it, and what does the myxoid extracellular matrix (ECM) tell the pathologist? What is the exact composition of this myxoid ECM and does it have a function? Here, we give an overview of the composition and constituents of the myxoid ECM as known so far and demonstrate the heterogeneity of the myxoid ECM among different tumors. We discuss the possible role of the predominant constituents of the myxoid ECM and attempt to relate them to differences in clinical behavior. Finally, we will speculate on the potential relevance of this knowledge in daily pathological practice.
Table 1.
Myxoid tumors of soft tissue: overview of clinicopathological and genetic features
| Myxoid tumors of soft tissue | Age | Sex | Predilection site | Molecular/cytogenetic aberrations | References |
|---|---|---|---|---|---|
| Benign | |||||
| Intramuscular myxoma (including its cellular variant) | Adults | F>M | Thigh, shoulder, buttocks, and rarely upper arm | GNAS1 mutations | [34] |
| Myxoid neurothekeoma | Young adults | F>M | Head, neck, and shoulders | Loss of 22q | [8] |
| Myxoid lipoma (myxolipoma) | Any age | F=M | Head and neck | Rearrangement of 13q and/or 16q | [8] |
| Myxoid chondroma | Adults | M>F | Hands and feet | Extra copies of chromosome 5 or 12q13–15 rearrangement | [8] |
| Myxoid neurofibroma | Any age | M=F | All over the body | NF1 mutations | [8] |
| Myxoid dermatofibroma | Young adults | F>M | Lower extremities | Unknown | [42] |
| Cardiac myxoma | Adults | F>M | Atria (predominantly left) | PRKAR1α mutations in Carney complex | [43] |
| Ossifying fibromyxoid tumor | Elderly | M>F | Extremities and trunk | Nonspecific cytogenetic aberrations | [8] |
| Cutaneous myxoid cyst | Any age | F>M | Distal and dorsal portions of fingers (and toes) | Unknown | [8] |
| Cutaneous myxoma (superficial angiomyxoma) | Adults | M>F | Trunk, lower extremities, head, and neck | PRKAR1α mutations in Carney complex | [43] |
| Myxoid nodular fasciitis | Young adults | M=F | Head, neck, and extremities | Nonspecific cytogenetic aberrations | [8, 44] |
| Locally aggressive | |||||
| Odontogenic myxoma | Young adults | F>M | Mandible and maxilla | PRKAR1α mutations in rare cases (not Carney complex) | [45] |
| Myxoinflammatory fibroblastic sarcoma | Adults | M=F | Feet, lower leg | t(1;10)(p22;q24) and amplification of chromosome 3 | [46] |
| Aggressive angiomyxoma | Adults | F>>M | Inguinal region | Rearrangement of 12q13-15 | [8] |
| Malignant | |||||
| Myxofibrosarcoma | Elderly | M=F | Extremities, thigh | Nonspecific cytogenetic aberrations | [47, 48] |
| Extraskeletal myxoid chondrosarcoma | Adults | M>F | Extremities and limb | t(9;22)(q22;q12), t(9;17)(q22;q11) or t(9;15)(q22;q21) | [49] |
| Low-grade fibromyxoid sarcoma | Young adults | M>F | Proximal extremities and trunk | t(7,16) (q33;p11) | [50] |
| Myxoid liposarcoma | Elderly | M>F | Lower extremities and thigh | t(12;16)(q13;p11) or t(12;22)(q13;q12) | [51] |
| Myxoid leiomyosarcoma of soft tissue | Adults | F>>M | Limbs, female genitalia, head, and neck | Nonspecific cytogenetic aberrations | [52] |
| Myxoid malignant peripheral nerve sheath tumor | Adults | F=M | Extremities, trunk | NF1 and TP53 mutations, P16 deletions | [8, 53] |
| Myxoid dermatofibrosarcoma | Adults | M=F | Trunk, groin, and extremities | t(17;22)(q22;q13) | [54] |
Composition of the myxoid extracellular matrix
Glycosaminoglycans and proteoglycans
Substantial progress in the study of the myxoid ECM was made after the introduction of alcian blue staining in 1950 [9]. John Scott was one of the pioneers who used this histochemical stain to distinguish between the different glycosaminoglycans (GAGs) in tissue sections [10]. Kindblom et al. showed that the myxoid ECM of various (non)neoplastic tissues, i.e., Wharton’s jelly and myxoid tumors of soft tissue, contained large amounts of GAGs (Figs. 1 and 2) [11]. GAGs are large macromolecules abundantly present in pericellular and extracellular matrices and consist of unbranched polysaccharide chains of disaccharides which are often sulfated. There are six different types of GAGs: hyaluronic acid (HA), keratin sulfate, chondroitin sulfate, dermatan sulfate, heparan sulfate, and heparin [12]. GAGs form proteoglycans (PGs) once covalently attached to specific core proteins. Core proteins of PGs are synthesized in the endoplasmic reticulum and post-translationally modified as they pass through the Golgi apparatus where hexuronic acid and hexosamine groups are attached. The exception is HA, which is synthesized directly under the cytoplasmic membrane by the hyaluronic acid synthetases 1, 2, and 3 [12]. The most common classification of the different PGs is based upon the properties of the core protein. The three main PG families present in the ECM are lecticans, small leucine-rich proteoglycans (SLRPs), and other ECM PG [13]. Lecticans always contain both a hyaluronan-binding domain and a C-type lectin domain. The lectican family includes: aggrecan, versican, neurocan, and brevican that can be found at different locations (Table 3) [12]. SLRPs can be found extracellularly and intracellularly and at the cell surface. They contain nine to 12 tandem repeats of leucine-rich motifs which involve their collagen-binding domains. The SLRP family includes decorin, biglycan, asporin, ECM protein 2, keratocan, proline/arginine-rich and leucine-rich repeat proteins, osteoadherin, lumican, fibromodulin, opticin, epiphycan, osteoglycin, podocan, chondroadherin, and nyctalopin [14]. ECM PGs do not show significant homology in the content of their core proteins. Perlecan, agrin, and collagen types XV and XVIII belong in this family [14]. Various GAGs and PGs have been identified in the myxoid ECM (Table 2): HA is the most common; none of them are specific for one particular lesion.
Table 3.
GAGs and PGs: their role in physiology and pathologic processes [19]
| Type and presence | Physiology | Pathology | |
|---|---|---|---|
| C4S (CS-A) | Sulfated galactosaminoglycan; cartilage, skin and tendon | Binds Ca2+, Cu2+ and Fe2+ ions; antioxidant (better then C6S and HA) | Decreased in OA, mediate adherence of plasmodium infected red blood cells |
| C6S (CS-C) | Sulfated galactosaminoglycan; cartilage, brain secretory granules | Reduces proinflammatory cytokines, MMPs, NO, and apoptosis | Laryngeal cancer, decreased in OA, increased in early atherosclerotic lesions |
| DS (CS-B) | Sulfated galactosaminoglycan; skin, blood vessels, heart, tendons, lungs | Regulation ECM integrity and cellular signaling; DS selectively activates heparin cofactor II that inactivates thrombin; carcinogenesis, wound repair, and fibrosis; DS binds water, coagulation | Dermatan sulfate accumulates abnormally in several of the mucopolysaccharidosis disorders and in myxomatous degeneration. Involved in cardiovascular disease, infection, fibrosis |
| HA | Lack sulfation and epimerization of glucuronic acid moiety to uronic acid, the only GAG synthesized in the cytoplasm at the plasma membrane and also the only GAG that is synthesized without core protein. Connective, epithelial and neural tissue. abundant in cartilage and bone | Early development, tissue organization, cell proliferation, facilitate migration and condensation of mesenchymal cells, participates in joint cavity formation, binds and immobilizes aggrecan, regulates osteoblast and osteoclast function. HA works as a scaffold for building PGs, suppresses cartilage degeneration and reduce pain perception, associated with cell adhesion and motility, suppresses prostaglandin E2 and IL-1 production, activates SRC, FAK, ERK and PKC whereas interaction with CD44 also regulates ERBB, PI3K, regulates phosphorylation of BAD and hence promotes cell survival, contributes to cell proliferation and migration, bone turnover, involved in tissue repair in skin, binds to receptor CD44 | Used for treatment of osteoarthritis |
| HS | Sulfated glucosaminoglycan; all types of cells, highly abundant in ECM of the skeleton | Coreceptors for morphogens, sequester growth factors and cytokines to regulate cell differentiation and growth, compose ECM scaffolds that make physical separation of the niche from cellular and signaling influence of surrounding environment, involved in skeletal patterning, differentiation, growth and homeostasis, critical for hematopoietic stem cell inch, Ndst1 mutation causes brain/skull defects and lung surfactant problems resulting in perinatal lethality, Ndst2 mutant have defective granule formation in mast cells, stimulates angiogenesis, osteocastogenesis, skeletal patterning, differentiation and homeostasis, coreceptor for morphogens, sequester growth factors and cytokines to regulate cell differentiation and growth, FGF-binding, binds fibronectin | Sequesters chemokines or FGF towards migrating tumor cells, promotes metastasis, multiple osteochondromas (MO)—benign bone cartilaginous tumor caused by mutant in EXT1 or EXT2, accumulated in mucopolysaccharidoses |
| KS | Sulfated glucosaminoglycan; N-glycan KSI or O-glycan KSII. Highly abundant in cornea and cartilage. Also found in epithelial tissue, central nervous system | Maintains proper special organization of the type I collagen fibrils and promotes transparency of cornea, cellular recognition of protein ligands, cell motility | Corneal opacity and corneal dystrophy (KS lacks GlcNAc sulfation), epithelial-derived carcinoma cells, alerted sulfation levels of KS was found in brain of Alzheimer patients |
| Aggrecan | O- and N-linked KSII, CS, DS, KS (HS absent), cartilage | Maintains tissue hydration, contributes to the mechanical properties of tissue, inhibits migration of neural crest cells, null mice show cartilage defects and delay in bone development | Chondrodystrophy, nanomelia, cartilage matrix deficiency (CMD), murine brachymorphism (bm), spondyloepimetaphyseal dysplasia enhanced expression in chondroblastoma, chondroma, chondrosarcoma, osteosarcoma, decreased in squamous cell carcinoma chondrodystrophy, nanomelia, cartilage matrix deficiency, spondyloepimataphyseal dysplasia |
| Biglycan | CS, DS (HS absent), bone, cartilage, skin, connective tissue | Activates cell division, organization of collage fibers, increased in vascular injury, upregulates p27 and downregulates cyclin A and proliferating cell nuclear antigen, maintaining proper number of mature osteoblasts and survival of bone marrow stromal cells, organization of collagen fibers, regulator of cell cycle, binds TGF-beta | Overexpressed in pancreatic cancer and hyperplasic thymus osteoporosis |
| Decorin | CS, DS (HS absent), connective tissue, cornea | Inhibits collagen fiber formation by interaction with col I, col II, and col VI, inhibits cell division, adhesion, increased in vascular injury, downregulates Erbb2 and MAP kinases, upregulates p21 CDK inhibitor leading to inhibition of cell proliferation and specific induction of apoptosis in transformed cells. maintaining proper number of mature osteoblasts and maintaining survival of bone marrow stromal cells, bind nonfibril collagens XII and XIV, regulates cell proliferation, binds TGF-beta, mediates EGF signaling by binding to EGFR | Antiproliferative properties in tumor growth, overexpressed in colorectal carcinoma, colon adenocarcinoma, melanoma, osteosarcoma, basal cell carcinoma, inhibits migration of MG-63 osteosarcoma cells, reduced decorin levels were found in lung adenocarcinoma, squamous carcinoma, breast carcinoma, hepatocellular carcinoma and ovarian tumors, may regulate tumor angiogenesis, overexpression is often associated with shift from DS to CS, osteoporosis |
| Lumican | KS, cornea | Upregulates p27 and downregulates cyclin A and proliferating cell nuclear antigen, regulates collagen fibril organization and circumferential growth, corneal transparency, and epithelial cell migration and tissue repair | Upregulated in pancreatic, colorectal and breast cancers, stroma of salivary pleiotropic adenoma, reduced expression is correlated with progression of breast carcinoma |
| Perlecan | HS, CS, cartilage, limb bud mesenchyme, articular cartilage, bone marrow stroma, all basal membranes, vasculature | Growth factor signaling, collagen fibrillogenesis, structural stability, vasculogenesis, endorepellin, antiangiogenic factor, chondrocyte proliferation and differentiation, collagen I and II fibrillogenesis, vasculogenesis, mediator of Shh signaling, Wnt signaling, TGF-beta signaling in the skeleton, regulates FGF2 signaling | Schwartz–Jampel syndrome, dyssegmental dysplasia Silverman–Handmaker type (DDSH), perlecan-null embryo chondroplasia, prostate tumor metastasis |
| Versican | C6S>C4S>DS (HS, KS absent), connective tissue, aorta, brain; fibroblasts. important for vascular biology | Lipid retention, modification and accumulation, hydration of ECM, cell proliferation, migration, embryo development, binds HA, CD44, and chemokines | Promotes tumor growth and spread, expressed in the stroma of nearly all human cancers (prostate, breast, lung, ovarian cancers and odontogenic tumors, melanoma, brain tumors, pharyngeal squamous cell carcinoma, keratinocyte tumors, atherosclerosis) |
Table 2.
The composition of the myxoid ECM is heterogeneous but not lesion-specific
| Proteoglycans | Collagens (types) | Other ECM components | References | |
|---|---|---|---|---|
| Non-neoplastic myxoid lesions | ||||
| Follicular mucinosis | HA | [55] | ||
| Myxedema | HA | I (III) | [56, 57] | |
| Stenotic arteries | Versican, biglycan, perlecan, HA | I | TBFbeta1 | [58, 59] |
| Pseudoaneurysm | Versican | [60] | ||
| Endocardiosis | HA | [61] | ||
| Valvular degeneration of the heart | I, III | [62] | ||
| Myxoid tumors of soft tissue | ||||
| Intramuscular myxoma | KS, HA, C4S, C6S | I, VI | Albumin, IgGs | [20, 34] |
| Myxofibrosarcoma | KS, HA, C4S, C6S | I, VI, XII, XIV | Thrombospondin Albumin, IgGs | [8, 20, 34] |
| Extraskeletal myxoid chondrosarcoma | C4S, C6S, HA (aggrecan) | I, III, VI (II, IV) | Albumin, IgGs | [20, 63] |
| Odontogenic myxoma | C4S, C6S, DS, KS, HS, aggrecan, versican, biglycan, decorin | [64] | ||
| Low-grade fibromyxoid sarcoma | HA | [65] | ||
| Myxoid neurothekeoma | HA | [66] | ||
| Myxoid lipoma (myxolipoma) | HA | [67] | ||
| Myxoid liposarcoma | HA | FN | [68] | |
| Myxoid chondroma | HA, KS | [69] | ||
| Myxoid neurofibroma | HA, C4S, C6S | [69] | ||
| Chondromyxoid fibroma | Aggrecan | I, III, VI | [70] | |
| Myxoid dermatofibroma | HA | [71] | ||
| Myxoid leiomyosarcoma | HA | [72] | ||
| Cardiac myxoma | C4S, C6S, (HA) | [73] | ||
| Ossifying fibromyxoid tumor | IV (II) | [74, 75] | ||
| Myxoinflammatory fibroblastic sarcoma | A1AT, A1ACT | [76] | ||
| Cutaneous myxoid cyst | HA | [77] | ||
| Aggressive angiomyxoma | HA | [78] | ||
| Cutaneous myxoma (superficial angiomyxoma) | HA | [79] | ||
| Myxoid areas in epithelial tumors | ||||
| Breast carcinoma | Aggrecan, versican, HA | I, II, IV | [80] | |
| Pleiomorphic adenoma | Lumican, perlecan, aggrecan, C4S, C6S, DS, KS, HA | I, III, IV | FGF2, Tenascin, FN, ChM-I | [81–83] |
| Vulvar squamous cell carcinoma | CD44, TGF-beta3 | [84] | ||
| Mixed tumor of skin | IV | Tenascin, FN | [85] | |
| Cholangiocarcinoma | Perlecan | [86] | ||
| Miscellaneous tumors (nonsoft tissue, nonepithelial) | ||||
| Myxopapillary ependymoma | HA, CS, HS | [87] | ||
| Myxoid mesothelioma | HA | [88] | ||
| Myxoid meningioma | HA, CS | [89] | ||
KS keratin sulfate, HA hyaluronic acid, C4S chondroitin sulfate, C6S chondroitin 6 sulfate, DS dermatan sulfate, HS heparan sulfate
Collagens
One of the first papers addressing myxoma and its malignant counterpart (“myxosarcoma”) mentioned the presence of fibrillary collagens as a hallmark for differential diagnosis [6]. Though this criterion did not last long, collagens (κολλα = glue; γηνε = that which produces) are a main component of the myxoid ECM. They are characterized by their regular, triple-stranded helix of so-called alpha-chains forming cord-like strands of 300 nm in length and 1.5 nm in diameter. A separate group of collagens is formed by the fibril-associated collagens with interrupted triple helices (FACIT) and includes collagen types XII and XIV. These collagens have several triple helical domains (collagen type domains [Col]) separated by nontriple helical domains (NC). All collagens contain large amounts of proline and glycine as well as hydroxyproline and hydroxylysine which are formed by post-translational modification. Based on their biochemical differences, more than 30 different types of collagens are recognized [15]. The most common types are I, II, III, and IV, which account for 90% of all collagens in humans. Except for collagen II, which is predominantly present in cartilage, collagen types I, III, and IV as well as VI, XII, and XIV may be found in the myxoid ECM (Table 2).
Other ECM molecules
Other structural molecules identified in the myxoid ECM are fibronectin and tenascin C (Table 2). Fibronectin is a fibril-forming glycoprotein existing in a dimeric or multimeric form. Each monomer contains several binding sites for fibrin, heparin, DNA, and cells. Fibronectin molecules consist of different repeats (types I, II, and III) and three different sites that can be alternatively spliced (EDA, EDB, and V). The dimeric, soluble form is produced by hepatocytes and lacks the alternative EDA and EDB variants. The multimeric form is extensively present in granulation tissue, basement membrane, and on cell surfaces and contains variable proportions of the EDA and EDB domains [16]. Until now, fibronectin has only been found in myxoid liposarcoma but might also be present in the ECM of other myxoid lesions (Table 2).
Tenascin C is a highly conserved glycoprotein of the ECM consisting of 300 kDa monomers, characteristically assembled in 1,800 kDa hexamers [17, 18]. It consists of several functionally independent domains of which the number is dramatically increased by alternative splicing. The N-terminal contains the cysteine-rich assembly domain, followed by EGF-like repeats, eight constant and up to nine alternatively spliced fibronectin type III repeats and a C-terminal fibrinogen-like globular domain [18]. Till today, its presence has only been shown in myxoid areas of epithelial but not (yet) in mesenchymal tumors (Table 2).
Functional role of the different constituents in the myxoid extracellular matrix
Glycosaminoglycans
GAGs have both biophysical and biochemical functions and play important roles in physiologic and neoplastic processes (Table 3) [19]. Due to their high content of sulfate and carboxyl groups, complex patterns of sulfation and uronic acid epimerizations, GAG chains confer upon PGs the diverse capacities to function as ideal physiological barriers, reservoirs for signaling proteins, and binding partners for structural macromolecules [13]. We have shown that the myxoid ECM in soft tissue tumors is heterogeneous in composition and that the relative amount of each GAG is tumor-type- and tumor-grade-dependent [20]. Because of their negative charge, all GAGs, especially HA, are able to trap water molecules. Interestingly, HA is the common denominator in the myxoid ECM (Table 2). This suggests that HA is the major contributor to the edematous appearance of the myxoid ECM. As a result of the biophysical properties of GAGs (their high viscosity and low compressibility), they are ideal for tissue lubrication. On the other hand, their rigidity is responsible for the structural integrity of tissues facilitating diffusion of metabolites and cell migration [21]. The biochemical properties of GAGs are mediated by specific binding to other macromolecules. GAGs can bind to secreted proteases and antiproteases, growth factors, structural ECM proteins, and proteins expressed on (tumor) cells [22]. Chondroitin sulfate modulates cell fate as it appears to prevent apoptosis and is involved in cell proliferation. Since chondroitin sulfate is much more abundant in the ECM of extraskeletal myxoid chondrosarcoma compared to intramuscular myxoma and myxofibrosarcoma, it might, therefore, play a role in the more malignant behavior of this tumor [20]. Large multidomain ECM molecules such as collagen types I, III, V, and XIV and fibronectin contain at least one GAG binding site. This allows them to bind to heparan and chondroitin sulfates on cells or in the ECM, contributing to proper ECM formation.
Proteoglycans
PGs exhibit a wide variety of functions due to their structural diversity (Table 3). As PGs avidly bind proteins, they are involved in all cellular processes concerning cell–matrix, cell–cell, and ligand–receptor interactions. PGs are known to have affinity for a variety of ligands, including growth factors, cell adhesion molecules, ECM components, enzymes, and enzyme inhibitors [22]. Lecticans bind other ECM proteins with its C-type lectin motif, facilitating the formation of networks permissive for cell growth [23]. For example, aggrecan and versican associate tightly with both HA, thereby maintaining tissue hydration. Due to the EGF-like repeats, lecticans are directly involved in growth control. Versican stimulates the proliferation of fibroblasts and is highly expressed by fast-growing cells and present in myxoid areas of both reactive and neoplastic (mesenchymal and epithelial) lesions (Table 2) [24].
Small leucine-rich proteoglycans
SLRPs are structurally and functionally related ECM molecules and abundantly expressed in connective tissues. Decorin and biglycan bind to collagen and influence collagen fibrillogenesis and ECM assembly in various ways: (a) due to its curved shape, decorin is able to bind to collagen types I, II, VI, and XIV linking them together and to fibronectin [25], (b) by decorating collagen fibers, decorin protects them from degradation by collagenases [25, 26], and (c) by its attached GAG chain, decorin is capable of modulating the activity of TGF-beta, which plays a central role in fibrogenesis [13, 25, 27]. Biglycan- and decorin-deficient mice show irregular and defective collagen fibrils, fragile skin, and a phenotype that closely resembles that of patients with Ehlers–Danlos syndrome [28]. In wound healing, there is a spatial temporal regulation of the expression of the different SLRPs (decorin, lumican, fibromodulin, and biglycan) tightly controlling the transformation of the myxoid ECM of granulation tissue towards fibrotic scar formation [29]. Myxoid areas in pleomorphic adenoma of the salivary gland and odontogenic myxoma lack expression of biglycan, lumican, and decorin, whereas these SLRPs are diffusely present in the fibrotic parts of the ECM of these tumors (Table 2). This suggests that the absence of SLRPs might contribute to impaired ECM formation resulting in a mere myxoid morphology.
Collagens
As form follows function [30], different collagens have different properties. In the initial phase of wound healing, collagen type I is quickly produced by fibroblasts and replaced by collagen type III in the later stage of scar formation [29]. The FACIT collagen types XII and XIV do not form collagen fibrils themselves but associate with fibrillary collagens, such as collagen type I, decorin, and GAGs, linking them together [31–33]. Collagen type XIV interacts with dermatan sulfate sequences on the single chondroitin sulfate/dermatan sulfate chain attached to decorin, thereby providing a link between the fibril-forming and the fibril-associated collagens [32]. Collagen type XIV is expressed significantly higher in grade I myxofibrosarcoma than in intramuscular myxoma (including its cellular variant) [34]. On one hand, this might have implications for ECM assembly and tumor development, thereby playing a potential role in the different biology and clinical behavior of these different entities; on the other hand, the effect of cell–ECM interaction in these tumors might work the other way around, as collagen XIV induces differentiation of fibroblastic cells [35]. Higher expression of collagen XIV in myxofibrosarcoma is associated with a more mature morphology of the tumor cells compared to intramuscular myxoma [5].
Other ECM molecules
Fibronectin plays a role in various biological processes, such as wound healing, host defense, regulation of ECM assembly, adhesion, and proliferation [36]. It is also a key molecule in cell–ECM signaling, and aberrant activation of fibronectin–ECM signaling has been described in many tumors [37]. Likewise, tenascin C is an ECM glycoprotein of which the spatial and temporal expression is tightly regulated during fetal development [38]. It has been shown that the tenascin C-encoding gene is upregulated in several pathologic conditions, such as wound healing, inflammation, and malignancies [38]. Elevated tenascin C expression has been found in epithelial and mesenchymal tumors, both in tumor and stromal cells, and is associated with unfavorable disease outcome (Table 2) [18]. Tenascin C attaches to cells via integrins, though the effect of this interaction on cellular signaling and tumor development is largely unknown [39].
Putative relevance for routine pathology practice
In routine general practice, one encounters a group of tumors, mesenchymal as well as epithelial, which may show variable degrees of myxoid appearance of their ECM. Myxoid tumors of soft tissue, the focus of this paper, encompass a heterogeneous group of lesions characterized by a marked abundance of extracellular mucoid/myxoid matrix (Table 1). This group of tumors exhibits a broad range of biological behavior varying from those which are entirely benign via locally aggressive (but nonmetastasizing) behavior to those which are frankly malignant; hence, accurate histopathological diagnosis is essential for appropriate clinical management. Till today, there have been no convincing data which demonstrate that the composition of the myxoid matrix significantly differ between tumors (both of mesenchymal and/or epithelial origin) and reactive lesions. From a biochemical point of view, the term “myxoid” is best considered a wastebasket of many different proteins and other (macro)molecules which might all have a different function (Tables 2 and 3). While determination of the different GAGs in myxoid tumors is interesting from a research point of view, Scott’s critical electrolyte concentration (CEC) method is no longer a routinely used technique by pathologists. Most myxoid tumors of soft tissue tumors contain HA (Table 2), and consequently, its detection is no longer regarded as discriminatory in differential diagnoses. Outside the academic setting (where recourse to sophisticated biochemical and molecular genetics techniques is generally not possible), when confronted with challenging myxoid lesions, the diagnostic process relies primarily on careful and discriminatory histopathological evaluation in order to reach a correct diagnosis. Routine histological criteria (e.g., tumor demarcation, growth and vascular pattern, and nuclear atypia) remain the hallmarks of diagnosis. (Immuno-)histochemistry may be useful as a adjunct in the setting of certain differential diagnostic considerations, but is often not discriminatory and, therefore, not diagnostically useful [8]. Molecular cytogenetics is of increasing importance for diagnosis in difficult cases, as the genetic makeup of an increasing number of myxoid tumors of soft tissue is being elucidated (Table 1). Next to genetic causes (such as driver mutations, translocations, or gene amplifications), environmental factors have been shown to play an important role in tumorigenesis of both epithelial and mesenchymal tumors [40]. Virchow already mentioned the possible relationship between the histological appearance of the myxoid ECM of tumors and their differences in biological behavior [1]. It is very tempting to believe that GAGs and PGs play an important role in the biology of myxoid lesions rather than just being a byproduct of (myo)fibroblastic differentiation. Intramuscular myxoma expresses significantly less decorin and collagen VI and XIV (both protein and mRNA level) than grade I myxofibrosarcoma [34]. Next to decorin, liquid chromatography–mass spectrometry revealed other SLRPs (i.e., lumican, prolargin 4, biglycan) present in tumor lysates of grade I myxofibrosarcoma but not of intramuscular myxoma [34]. This suggest that ECM formation in intramuscular myxoma is impaired compared to grade I myxofibrosarcoma. Increased ECM rigidity activates integrins and subsequently leads to stimulation of the rho/Rock pathway and cell migration and invasion [41]. In this respect, the morphological similarity between the process of wound healing and myxoid changes/areas in (non)neoplastic lesions is rather striking. In the early phase, granulation tissue shows a loose, edematous/myxoid ECM rich in GAGs. In the subsequent phase, there is structural organization of the ECM with deposition of collagens produced by myofibroblasts which locally (de)differentiate from fibroblast/smooth muscle cells or are derived from the bone marrow. The term “myxoid” might not only be synonymous with (aberrant turnover and) the presence of GAGs, but also reflects the absence of proper ECM assembly or incomplete ECM formation. This might well explain that a myxoid morphology of the ECM is not specific at all and can be seen in reactive/non-neoplastic processes as well as in benign and malignant tumors (Fig. 3). As already extensively discussed in this paper, the ECM is dependent on the interplay of various macromolecules (polysaccharides and proteins) which determine its texture and consistency. It is becoming increasingly clear that, within this group of lesions, various tumor types may show differences in the biological composition of their myxoid matrix, as determined by differences in the relative proportions of the constituent macromolecules and the presence or absence of specific stromal constituents. Clearly, this is an important and highly relevant area for further research in order to identify tumor-specific markers and to consequently develop economically viable and user-friendly detection methods to facilitate the diagnostic process and ensure increasingly accurate histopathologic diagnosis.
Acknowledgements
Stefan M. Willems was financially supported by a grant from The Netherlands Organization for Health Research and Development (project no. 920-03-403).
Conflict of interest statement
We declare that we have no conflict of interest.
Open Access
This article is distributed under the terms of the Creative Commons Attribution Noncommercial License which permits any noncommercial use, distribution, and reproduction in any medium, provided the original author(s) and source are credited.
References
- 1.Virchow R. Die cellularpathologie in ihrer Begrundung auf physiologische und pathologische Gewebelehre. Berlin: Hirschwald; 1858. [Google Scholar]
- 2.Dunglison R. Medical lexicon. A dictionary of medical science. Philadelphia: Blanchard and Lea; 1865. [Google Scholar]
- 3.Muller J. Uber den feineren bau und die formen der krankhaften geschwulste. Berlin: Haberling; 1838. [Google Scholar]
- 4.Bryant T. Tumors and tumor formation. New York: Churchill; 1802. [Google Scholar]
- 5.Stout AP. Myxoma, the tumor of primitive mesenchyme. Ann Surg. 1948;127:706–719. doi: 10.1097/00000658-194804000-00013. [DOI] [PubMed] [Google Scholar]
- 6.Sponsel KH, McDonald JR, Ghormley RK. Myxoma and myxosarcoma of the soft tissues of the extremities. J Bone Joint Surg Am. 1952;34(A):820–826. [PubMed] [Google Scholar]
- 7.Fletcher CDM, Unni KK, Mertens F. WHO classification of tumours. Pathology and genetics of tumours of soft tissue and bone. Lyon: IARC; 2002. [Google Scholar]
- 8.Graadt van Roggen JF, Hogendoorn PCW, Fletcher CDM. Myxoid tumours of soft tissue. Histopathology. 1999;35:291–312. doi: 10.1046/j.1365-2559.1999.00835.x. [DOI] [PubMed] [Google Scholar]
- 9.Steedman HF. Alcian Blue 8 GS: a new stain for mucin. Q J Microsc Sci. 1950;3:477–479. [PubMed] [Google Scholar]
- 10.Scott JE, Dorling J. Differential staining of acid glycosaminoglycans (mucopolysaccharides) by alcian blue in salt solutions. Histochemie. 1965;5:221–233. doi: 10.1007/BF00306130. [DOI] [PubMed] [Google Scholar]
- 11.Kindblom LG, Angervall L. Histochemical characterization of mucosubstances in bone and soft tissue-tumors. Cancer. 1975;36:985–994. doi: 10.1002/1097-0142(197509)36:3<985::AID-CNCR2820360322>3.0.CO;2-F. [DOI] [PubMed] [Google Scholar]
- 12.Gandhi NS, Mancera RL. The structure of glycosaminoglycans and their interactions with proteins. Chem Biol Drug Des. 2008;72:455–482. doi: 10.1111/j.1747-0285.2008.00741.x. [DOI] [PubMed] [Google Scholar]
- 13.Kresse H, Schonherr E. Proteoglycans of the extracellular matrix and growth control. J Cell Physiol. 2001;189:266–274. doi: 10.1002/jcp.10030. [DOI] [PubMed] [Google Scholar]
- 14.Schaefer L, Iozzo RV. Biological functions of the small leucine-rich proteoglycans: from genetics to signal transduction. J Biol Chem. 2008;283:21305–21309. doi: 10.1074/jbc.R800020200. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Kadler KE, Hill A, Canty-Laird EG. Collagen fibrillogenesis: fibronectin, integrins, and minor collagens as organizers and nucleators. Curr Opin Cell Biol. 2008;20:495–501. doi: 10.1016/j.ceb.2008.06.008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.White ES, Baralle FE, Muro AF. New insights into form and function of fibronectin splice variants. J Pathol. 2008;216:1–14. doi: 10.1002/path.2388. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Bosman FT, Stamenkovic I. Functional structure and composition of the extracellular matrix. J Pathol. 2003;200:423–428. doi: 10.1002/path.1437. [DOI] [PubMed] [Google Scholar]
- 18.Orend G, Chiquet-Ehrismann R. Tenascin-C induced signaling in cancer. Cancer Lett. 2006;244:143–163. doi: 10.1016/j.canlet.2006.02.017. [DOI] [PubMed] [Google Scholar]
- 19.Lamoureux F, Baud’huin M, Duplomb L, et al. Proteoglycans: key partners in bone cell biology. BioEssays. 2007;29:758–771. doi: 10.1002/bies.20612. [DOI] [PubMed] [Google Scholar]
- 20.Willems SM, Schrage YM, Baelde JJ, et al. Myxoid tumours of soft tissue: the so-called myxoid extracellular matrix is heterogeneous in composition. Histopathology. 2008;52:465–474. doi: 10.1111/j.1365-2559.2008.02967.x. [DOI] [PubMed] [Google Scholar]
- 21.Almond A. Hyaluronan. Cell Mol Life Sci. 2007;64:1591–1596. doi: 10.1007/s00018-007-7032-z. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Rodgers KD, San Antonio JD, Jacenko O. Heparan sulfate proteoglycans: a GAGgle of skeletal-hematopoietic regulators. Dev Dyn. 2008;237:2622–2642. doi: 10.1002/dvdy.21593. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Evanko SP, Angello JC, Wight TN. Formation of hyaluronan- and versican-rich pericellular matrix is required for proliferation and migration of vascular smooth muscle cells. Arterioscler Thromb Vasc Biol. 1999;19:1004–1013. doi: 10.1161/01.atv.19.4.1004. [DOI] [PubMed] [Google Scholar]
- 24.Zimmermann DR, Dours-Zimmermann MT, Schubert M, et al. Versican is expressed in the proliferating zone in the epidermis and in association with the elastic network of the dermis. J Cell Biol. 1994;124:817–825. doi: 10.1083/jcb.124.5.817. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.McEwan PA, Scott PG, Bishop PN, et al. Structural correlations in the family of small leucine-rich repeat proteins and proteoglycans. J Struct Biol. 2006;155:294–305. doi: 10.1016/j.jsb.2006.01.016. [DOI] [PubMed] [Google Scholar]
- 26.Geng Y, McQuillan D, Roughley PJ. SLRP interaction can protect collagen fibrils from cleavage by collagenases. Matrix Biol. 2006;25:484–491. doi: 10.1016/j.matbio.2006.08.259. [DOI] [PubMed] [Google Scholar]
- 27.Ferdous Z, Wei VM, Iozzo R, et al. Decorin-transforming growth factor- interaction regulates matrix organization and mechanical characteristics of three-dimensional collagen matrices. J Biol Chem. 2007;282:35887–35898. doi: 10.1074/jbc.M705180200. [DOI] [PubMed] [Google Scholar]
- 28.Danielson KG, Baribault H, Holmes DF, et al. Targeted disruption of decorin leads to abnormal collagen fibril morphology and skin fragility. J Cell Biol. 1997;136:729–743. doi: 10.1083/jcb.136.3.729. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Martinez DA, Vailas AC, Vanderby R, et al. Temporal extracellular matrix adaptations in ligament during wound healing and hindlimb unloading. Am J Physiol Regul Integr Comp Physiol. 2007;293:R1552–R1560. doi: 10.1152/ajpregu.00423.2007. [DOI] [PubMed] [Google Scholar]
- 30.Sullivan LH. The tall office building artistically considered. Lippincott’s Magazine. 1896;57:403–409. [Google Scholar]
- 31.Ehnis T, Dieterich W, Bauer M, et al. Localization of a binding site for the proteoglycan decorin on collagen XIV (undulin) J Biol Chem. 1997;272:20414–20419. doi: 10.1074/jbc.272.33.20414. [DOI] [PubMed] [Google Scholar]
- 32.Ehnis T, Dieterich W, Bauer M, et al. A chondroitin/dermatan sulfate form of CD44 is a receptor for collagen XIV (undulin) Exp Cell Res. 1996;229:388–397. doi: 10.1006/excr.1996.0384. [DOI] [PubMed] [Google Scholar]
- 33.Canty EG, Kadler KE. Procollagen trafficking, processing and fibrillogenesis. J Cell Sci. 2005;118:1341–1353. doi: 10.1242/jcs.01731. [DOI] [PubMed] [Google Scholar]
- 34.Willems SM, Mohseny AB, Balog C, et al. Cellular/intramuscular myxoma and grade I myxofibrosarcoma are characterized by distinct genetic alterations and specific composition of their extracellular matrix. J Cell Mol Med. 2009;7:1291–1301. doi: 10.1111/j.1582-4934.2009.00747.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Ruehl M, Erben U, Schuppan D, et al. The elongated first fibronectin type III domain of collagen XIV is an inducer of quiescence and differentiation in fibroblasts and preadipocytes. J Biol Chem. 2005;280:38537–38543. doi: 10.1074/jbc.M502210200. [DOI] [PubMed] [Google Scholar]
- 36.Vakonakis I, Campbell ID. Extracellular matrix: from atomic resolution to ultrastructure. Curr Opin Cell Biol. 2007;19:578–583. doi: 10.1016/j.ceb.2007.09.005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Larsen M, Artym VV, Green JA, et al. The matrix reorganized: extracellular matrix remodeling and integrin signaling. Curr Opin Cell Biol. 2006;18:463–471. doi: 10.1016/j.ceb.2006.08.009. [DOI] [PubMed] [Google Scholar]
- 38.Harty M, Neff AW, King MW, et al. Regeneration or scarring: an immunologic perspective. Dev Dyn. 2003;226:268–279. doi: 10.1002/dvdy.10239. [DOI] [PubMed] [Google Scholar]
- 39.Jones FS, Jones PL. The tenascin family of ECM glycoproteins: structure, function, and regulation during embryonic development and tissue remodeling. Dev Dyn. 2000;218:235–259. doi: 10.1002/(SICI)1097-0177(200006)218:2<235::AID-DVDY2>3.0.CO;2-G. [DOI] [PubMed] [Google Scholar]
- 40.De Wever O, Mareel M. Role of tissue stroma in cancer cell invasion. J Pathol. 2003;200:429–447. doi: 10.1002/path.1398. [DOI] [PubMed] [Google Scholar]
- 41.Berrier AL, Yamada KM. Cell–matrix adhesion. J Cell Physiol. 2007;213:565–573. doi: 10.1002/jcp.21237. [DOI] [PubMed] [Google Scholar]
- 42.Fletcher CDM. Benign fibrous histiocytoma of subcutaneous and deep soft tissue: a clinicopathologic analysis of 21 cases. Am J Surg Pathol. 1990;14:801–809. [PubMed] [Google Scholar]
- 43.Carney JA, Gordon H, Carpenter PC, et al. The complex of myxomas, spotty pigmentation, and endocrine overactivity. Medicine (Baltimore) 1985;64:270–283. doi: 10.1097/00005792-198507000-00007. [DOI] [PubMed] [Google Scholar]
- 44.Meng GZ, Zhang HY, Zhang Z, et al. Myofibroblastic sarcoma vs nodular fasciitis: a comparative study of chromosomal imbalances. Am J Clin Pathol. 2009;131:701–709. doi: 10.1309/AJCPV6H2WSYXLKFB. [DOI] [PubMed] [Google Scholar]
- 45.Perdigao PF, Stergiopoulos SG, De Marco L, et al. Molecular and immunohistochemical investigation of protein kinase a regulatory subunit type 1A (PRKAR1A) in odontogenic myxomas. Genes Chromosomes Cancer. 2005;44:204–211. doi: 10.1002/gcc.20232. [DOI] [PubMed] [Google Scholar]
- 46.Hallor KH, Sciot R, Staaf J, et al. Two genetic pathways, t(1;10) and amplification of 3p11–12, in myxoinflammatory fibroblastic sarcoma, haemosiderotic fibrolipomatous tumour, and morphologically similar lesions. J Pathol. 2009;217:716–727. doi: 10.1002/path.2513. [DOI] [PubMed] [Google Scholar]
- 47.Calonje E, Mentzel T, Fletcher CDM. Cellular benign fibrous histiocytoma. Clinicopathologic analysis of 74 cases of a distinctive variant of cutaneous fibrous histiocytoma with frequent recurrence. Am J Surg Pathol. 1994;18:668–676. [PubMed] [Google Scholar]
- 48.Willems SM, Debiec-Rychter M, Szuhai K, et al. Local recurrence of myxofibrosarcoma is associated with increase in tumour grade and cytogenetic aberrations, suggesting a multistep tumour progression model. Mod Pathol. 2006;19:407–416. doi: 10.1038/modpathol.3800550. [DOI] [PubMed] [Google Scholar]
- 49.Enzinger FM, Shiraki M. Extraskeletal myxoid chondrosarcoma. An analysis of 34 cases. Hum Pathol. 1972;3:421–435. doi: 10.1016/S0046-8177(72)80042-X. [DOI] [PubMed] [Google Scholar]
- 50.Evans HL. Low-grade fibromyxoid sarcoma. A report of 12 cases. Am J Surg Pathol. 1993;17:595–600. doi: 10.1097/00000478-199306000-00007. [DOI] [PubMed] [Google Scholar]
- 51.Turc-Carel C, Limon J, Dal CP, et al. Cytogenetic studies of adipose tissue tumors. II. Recurrent reciprocal translocation t(12;16)(q13;p11) in myxoid liposarcomas. Cancer Genet Cytogenet. 1986;23:291–299. doi: 10.1016/0165-4608(86)90011-7. [DOI] [PubMed] [Google Scholar]
- 52.Rubin BP, Fletcher CDM. Myxoid leiomyosarcoma of soft tissue, an underrecognized variant. Am J Surg Pathol. 2000;24:927–936. doi: 10.1097/00000478-200007000-00004. [DOI] [PubMed] [Google Scholar]
- 53.Brems H, Beert E, De Ravel RT, et al. Mechanisms in the pathogenesis of malignant tumours in neurofibromatosis type 1. Lancet Oncol. 2009;10:508–515. doi: 10.1016/S1470-2045(09)70033-6. [DOI] [PubMed] [Google Scholar]
- 54.Pedeutour F, Simon MP, Minoletti F, et al. Translocation, t(17;22)(q22;q13), in dermatofibrosarcoma protuberans: a new tumor-associated chromosome rearrangement. Cytogenet Cell Genet. 1996;72:171–174. doi: 10.1159/000134178. [DOI] [PubMed] [Google Scholar]
- 55.Kaya G, Augsburger E, Chavaz P, et al. CD44 and hyaluronate expression in follicular mucinosis. J Cutan Pathol. 2006;33:227–230. doi: 10.1111/j.0303-6987.2006.00463.x. [DOI] [PubMed] [Google Scholar]
- 56.Alves MF, Filgueira AL, Lorena DE, et al. Type I and type III collagens in cutaneous mucinosis. Am J Dermatopathol. 1998;20:41–47. doi: 10.1097/00000372-199802000-00008. [DOI] [PubMed] [Google Scholar]
- 57.Lengyel J, Vertes B. Hyaluronic acid studies of local myxedema and of the stroma of skin cancer. Dermatologica. 1956;113:219–225. doi: 10.1159/000256238. [DOI] [PubMed] [Google Scholar]
- 58.Chung IM, Gold HK, Schwartz SM, et al. Enhanced extracellular matrix accumulation in restenosis of coronary arteries after stent deployment. J Am Coll Cardiol. 2002;40:2072–2081. doi: 10.1016/S0735-1097(02)02598-6. [DOI] [PubMed] [Google Scholar]
- 59.Wight TN. Versican: a versatile extracellular matrix proteoglycan in cell biology. Curr Opin Cell Biol. 2002;14:617–623. doi: 10.1016/S0955-0674(02)00375-7. [DOI] [PubMed] [Google Scholar]
- 60.Burke AP, Jarvelainen H, Kolodgie FD, et al. Superficial pseudoaneurysms: clinicopathologic aspects and involvement of extracellular matrix proteoglycans. Mod Pathol. 2004;17:482–488. doi: 10.1038/modpathol.3800060. [DOI] [PubMed] [Google Scholar]
- 61.Black A, French AT, Dukes-McEwan J, et al. Ultrastructural morphologic evaluation of the phenotype of valvular interstitial cells in dogs with myxomatous degeneration of the mitral valve. Am J Vet Res. 2005;66:1408–1414. doi: 10.2460/ajvr.2005.66.1408. [DOI] [PubMed] [Google Scholar]
- 62.Nasuti JF, Zhang PJ, Feldman MD, et al. Fibrillin and other matrix proteins in mitral valve prolapse syndrome. Ann Thorac Surg. 2004;77:532–536. doi: 10.1016/S0003-4975(03)01584-4. [DOI] [PubMed] [Google Scholar]
- 63.Aigner T, Oliveira AM, Nascimento AG. Extraskeletal myxoid chondrosarcomas do not show a chondrocytic phenotype. Mod Pathol. 2004;17:214–221. doi: 10.1038/modpathol.3800036. [DOI] [PubMed] [Google Scholar]
- 64.Zhao M, Lu Y, Takata T, Mock D, Nikia H, et al. Immunohistochemical and histochemical characterization of the mucosubstances of odontogenic myxoma: histogenesis and differential diagnosis. Pathol Res Pract. 1999;195:391–397. doi: 10.1016/S0344-0338(99)80012-0. [DOI] [PubMed] [Google Scholar]
- 65.Zamecnik M, Michal M. Low-grade fibromyxoid sarcoma: a report of eight cases with histologic, immunohistochemical, and ultrastructural study. Ann Diagn Pathol. 2000;4:207–217. doi: 10.1053/adpa.2000.8122. [DOI] [PubMed] [Google Scholar]
- 66.Gallager RL, Helwig EB. Neurothekeoma—a benign cutaneous tumor of neural origin. Am J Clin Pathol. 1980;74:759–764. doi: 10.1093/ajcp/74.6.759. [DOI] [PubMed] [Google Scholar]
- 67.Hajdu SI. Pathology of soft tissue Tumors. Philadelphia: Lea and Febiger; 1979. [Google Scholar]
- 68.Fukuda T, Tsuneyoshi M. Adhesion proteins, cellular morphology and fibrous components around the cell/extracellular-matrix interface in myxoid liposarcomas. J Cancer Res Clin Oncol. 2000;126:320–324. doi: 10.1007/s004320050350. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 69.Mackenzie DH. The myxoid tumors of somatic soft tissues. Am J Surg Pathol. 1981;5:443–458. doi: 10.1097/00000478-198107000-00004. [DOI] [PubMed] [Google Scholar]
- 70.Soder S, Inwards C, Muller S, et al. Cell biology and matrix biochemistry of chondromyxoid fibroma. Am J Clin Pathol. 2001;116:271–277. doi: 10.1309/5BKG-E3CH-GGVA-1HDE. [DOI] [PubMed] [Google Scholar]
- 71.Calikoglu E, Chavaz P, Saurat JH, et al. Decreased CD44 expression and stromal hyaluronate accumulation in myxoid dermatofibroma. Dermatology. 2003;207:104–106. doi: 10.1159/000070959. [DOI] [PubMed] [Google Scholar]
- 72.Yasui W, Oda N, Ito H, et al. Myxoid leiomyosarcoma of the stomach: a case report. Jpn J Clin Oncol. 1991;21:447–452. [PubMed] [Google Scholar]
- 73.Lam RM, Hawkins ET, Roszka J. Cardiac myxoma: histochemical and ultrastructural localization of glycosaminoglycans and proteoglycans. Ultrastruct Pathol. 1984;6:69–81. doi: 10.3109/01913128409016666. [DOI] [PubMed] [Google Scholar]
- 74.Folpe AL, Weiss SW. Ossifying fibromyxoid tumor of soft parts: a clinicopathologic study of 70 cases with emphasis on atypical and malignant variants. Am J Surg Pathol. 2003;27:421–431. doi: 10.1097/00000478-200304000-00001. [DOI] [PubMed] [Google Scholar]
- 75.Yang P, Hirose T, Hasegawa T, et al. Ossifying fibromyxoid tumor of soft parts: a morphological and immunohistochemical study. Pathol Int. 1994;44:448–453. doi: 10.1111/j.1440-1827.1994.tb01709.x. [DOI] [PubMed] [Google Scholar]
- 76.Hassanein AM, Atkinson SP, Al-Quran SZ, et al. Acral myxoinflammatory fibroblastic sarcomas: are they all low-grade neoplasms? J Cutan Pathol. 2008;35:186–191. doi: 10.1111/j.1600-0560.2007.00789.x. [DOI] [PubMed] [Google Scholar]
- 77.Johnson WC, Graham JH, Helwig EB. Cutaneous myxoid cyst. A clinicopathological and histochemical study. JAMA. 1965;191:15–20. doi: 10.1001/jama.1965.03080010021004. [DOI] [PubMed] [Google Scholar]
- 78.Begin LR, Clement PB, Kirk ME, et al. Aggressive angiomyxoma of pelvic soft parts: a clinicopathologic study of nine cases. Hum Pathol. 1985;16:621–628. doi: 10.1016/S0046-8177(85)80112-X. [DOI] [PubMed] [Google Scholar]
- 79.Calikoglu E, Augsburger E, Masouye I, et al. Hyaluronate accumulation and decreased CD44 expression in perifollicular solitary cutaneous myxoma. Dermatology. 2002;205:122–126. doi: 10.1159/000063896. [DOI] [PubMed] [Google Scholar]
- 80.Kusafuka K, Muramatsu K, Kasami M, et al. Cartilaginous features in matrix-producing carcinoma of the breast: four cases report with histochemical and immunohistochemical analysis of matrix molecules. Mod Pathol. 2008;21:1282–1292. doi: 10.1038/modpathol.2008.120. [DOI] [PubMed] [Google Scholar]
- 81.Kusafuka K, Ishiwata T, Sugisaki Y, et al. Lumican expression is associated with the formation of mesenchyme-like elements in salivary pleomorphic adenomas. J Pathol. 2004;203:953–960. doi: 10.1002/path.1599. [DOI] [PubMed] [Google Scholar]
- 82.Nara Y, Takeuchi J, Yoshida K, et al. Immunohistochemical characterisation of extracellular matrix components of salivary gland tumours. Br J Cancer. 1991;64:307–314. doi: 10.1038/bjc.1991.297. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 83.Kusafuka K, Hiraki Y, Shukunami C, et al. Cartilage-specific matrix protein chondromodulin-I is associated with chondroid formation in salivary pleomorphic adenomas: immunohistochemical analysis. Am J Pathol. 2001;158:1465–1472. doi: 10.1016/S0002-9440(10)64097-7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 84.Ambros RA, Kallakury BV, Malfetano JH, et al. Cytokine, cell adhesion receptor, and tumor suppressor gene expression in vulvar squamous carcinoma: correlation with prominent fibromyxoid stromal response. Int J Gynecol Pathol. 1996;15:320–325. doi: 10.1097/00004347-199610000-00004. [DOI] [PubMed] [Google Scholar]
- 85.Franchi A, Dini M, Paglierani M, et al. Immunolocalization of extracellular matrix components in mixed tumors of the skin. Am J Dermatopathol. 1995;17:36–41. doi: 10.1097/00000372-199502000-00007. [DOI] [PubMed] [Google Scholar]
- 86.Sabit H, Tsuneyama K, Shimonishi T, et al. Enhanced expression of basement-membrane-type heparan sulfate proteoglycan in tumor fibro-myxoid stroma of intrahepatic cholangiocarcinoma. Pathol Int. 2001;51:248–256. doi: 10.1046/j.1440-1827.2001.01201.x. [DOI] [PubMed] [Google Scholar]
- 87.Giordana MT, Bertolotto A, Mauro A, et al. Glycosaminoglycans in human cerebral tumors. Part II. Histochemical findings and correlations. Acta Neuropathol. 1982;57:299–305. doi: 10.1007/BF00692187. [DOI] [PubMed] [Google Scholar]
- 88.Shia J, Qin J, Erlandson RA, et al. Malignant mesothelioma with a pronounced myxoid stroma: a clinical and pathological evaluation of 19 cases. Virchows Arch. 2005;447:828–834. doi: 10.1007/s00428-005-0035-y. [DOI] [PubMed] [Google Scholar]
- 89.Harrison JD, Rose PE. Myxoid meningioma: histochemistry and electron microscopy. Acta Neuropathol. 1985;68:80–82. doi: 10.1007/BF00688961. [DOI] [PubMed] [Google Scholar]
- 90.Henschen F. Fall von ostitis Fibrosa mit multiplen Tumoren in deer umgebenden Muskulatur. Verh Dtsch Ges Pathol. 1926;21:93–97. [Google Scholar]
- 91.Mazabraud A, Semat P, Roze R. A propos de l’association de fibromyxomes des tissus mous à la dysplasie fibreuse des os. Presse Med. 1967;75:2223–2228. [PubMed] [Google Scholar]
- 92.Weiss SW, Enzinger FM. Myxoid variant of malignant fibrous histiocytoma. Cancer. 1977;39:1672–1685. doi: 10.1002/1097-0142(197704)39:4<1672::AID-CNCR2820390442>3.0.CO;2-C. [DOI] [PubMed] [Google Scholar]
- 93.Angervall L, Kindblom LG, Merck C. Myxofibrosarcoma. A study of 30 cases. Acta Pathol Microbiol Scand A. 1977;85A:127–140. [PubMed] [Google Scholar]
- 94.Orndal C, Mandahl N, Rydholm A, et al. Chromosomal evolution and tumor progression in a myxoid liposarcoma. Acta Orthop Scand. 1990;61:99–105. doi: 10.3109/17453679009006497. [DOI] [PubMed] [Google Scholar]
- 95.Mentzel T, Calonje E, Wadden C, et al. Myxofibrosarcoma. Clinicopathologic analysis of 75 cases with emphasis on the low-grade variant. Am J Surg Pathol. 1996;20:391–405. doi: 10.1097/00000478-199604000-00001. [DOI] [PubMed] [Google Scholar]