

Abstract
Cannabinoids are a group of compounds that mediate their effects through cannabinoid receptors. The discovery of Δ9-tetrahydrocannabinol (THC) as the major psychoactive principle in marijuana, as well as the identification of cannabinoid receptors and their endogenous ligands, has led to a significant growth in research aimed at understanding the physiological functions of cannabinoids. Cannabinoid receptors include CB1, which is predominantly expressed in the brain, and CB2, which is primarily found on the cells of the immune system. The fact that both CB1 and CB2 receptors have been found on immune cells suggests that cannabinoids play an important role in the regulation of the immune system. Recent studies demonstrated that administration of THC into mice triggered marked apoptosis in T cells and dendritic cells, resulting in immunosuppression. In addition, several studies showed that cannabinoids downregulate cytokine and chemokine production and, in some models, upregulate T-regulatory cells (Tregs) as a mechanism to suppress inflammatory responses. The endocannabinoid system is also involved in immunoregulation. For example, administration of endocannabinoids or use of inhibitors of enzymes that break down the endocannabinoids, led to immunosuppression and recovery from immune-mediated injury to organs such as the liver. Manipulation of endocannabinoids and/or use of exogenous cannabinoids in vivo can constitute a potent treatment modality against inflammatory disorders. This review will focus on the potential use of cannabinoids as a new class of anti-inflammatory agents against a number of inflammatory and autoimmune diseases that are primarily triggered by activated T cells or other cellular immune components.
Cannabis, commonly known as marijuana, is a product of the Cannabis sativa plant and the active compounds from this plant are collectively referred to as cannabinoids. For several centuries, marijuana has been used as an alternative medicine in many cultures and, recently, its beneficial effects have been shown in: the treatment of nausea and vomiting associated with cancer chemotherapy; anorexia and cachexia seen in HIV/AIDS patients; and in neuropathic pain and spasticity in multiple sclerosis [1–4]. Cannabinoid pharmacology has made important advances in recent years after the discovery of the cannabinoid receptors (CB1 and CB2). Cannabinoid receptors and their endogenous ligands have provided an excellent platform for the investigation of the therapeutic effects of cannabinoids. It is well known that CB1 and CB2 are heterotrimeric Gi/o-protein-coupled receptors and that they are both expressed in the periphery and the CNS. However, CB1 expression is predominant in the CNS, especially on presynaptic nerves, and CB2 is primarily expressed on immune cells [5,6].
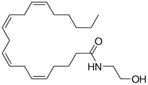
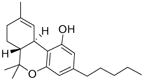
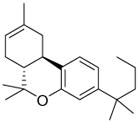
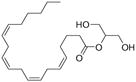
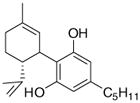
Arachidonic acid metabolites have been shown to exhibit properties similar to compounds found in Cannabis sativa. These metabolites are hence referred to as endocannabinoids. These ubiquitous endogenous cannabinoids act as natural ligands for the cannabinoid receptors expressed in mammalian tissue, thus constituting an important lipid-signaling system termed the endocannabinoid system. The endocannabinoid system is an important biological regulatory system that has been shown to be highly conserved from lower invertebrates to higher mammals [7]. Other than the lipid transmitters that serve as ligands for the cannabinoid receptors, the endocannabinoid family also comprises the enzymes for biosynthesis and degradation of the ligands. The endocannabinoids include N-arachidonoylethanolamine, anandamide (AEA), 2-arachidonoyl glycerol (2-AG), N-arachydonoyldopamine, noladin ether and virodhamine. AEA was discovered by Devane et al. and is an amide formed from arachidonic acid and ethanolamine [8]. AEA binds to brain CB1 with high affinity and mimics the behavioral actions of exogenous cannabinoid Δ9-tetrahydrocannabinol (THC) when injected into rodents. 2-AG was discovered independently 3 years later by Mechoulam et al. [9] and Sugiura et al. [10]. It was found to exist in much higher concentration in serum and brain than AEA. 2-AG has similar affinities for both CB1 and CB2 receptors, as does AEA, but it exhibits higher efficacy. Endocannabinoids are derivatives of arachidonic acid conjugated with either ethanolamine or glycerol. They are synthesized on demand from phospholipid precursors residing in the cell membrane in response to a rise in intracellular calcium levels. Inside cells, endocannabinoids are catalytically hydrolyzed by the aminohydrolase fatty acid amide hydrolase (FAAH), which degrades AEA into arachidonic acid and ethanolamine [11]. 2-AG is hydrolyzed into AEA and glycerol by either FAAH or by monoacyl glycerol lipase (MAGL). Fatty acid-binding proteins (FABPs) have been reported to play an important role as intracellular carriers in the transport of AEA from the plasma membrane to FAAH for their subsequent inactivation [12]. Studies to date indicate that the main pharmacological function of the endocannabinoid system is in neuromodulation: controlling motor functions, cognition, emotional responses, homeostasis and motivation. However, in the periphery, this system is an important modulator of the ANS, immune system and microcirculation [13]. Some well-known natural and synthetic cannabinoids and endocannabinoids are depicted in Table 1.
Table 1.
Selected cannabinoid molecules.
| Endocannabinoids | Natural cannabinoids | Synthetic cannabinoids |
|---|---|---|
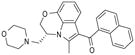
| Arachidonylethanolamide | Δ9-tetrahydrobannbinol | JWH-133 |
 |
 |
 |
| 2-arachidonoylglycerol | Cannabidiol | WIN55,212-2 |
 |
 |
 |
Cannabinoids are potent anti-inflammatory agents and they exert their effects through induction of apoptosis, inhibition of cell proliferation, suppression of cytokine production and induction of T-regulatory cells (Tregs). In this review, we provide an in-depth description of all four different mechanisms and we further discuss the immunosuppressive properties of cannabinoids in the context of inflammatory and autoimmune disease states, triggered by cellular rather than humoral components of the immune system.
Apoptotic effects of cannabinoids on immune cell populations
One major mechanism of immunosupression by cannabinoids is the induction of cell death or apoptosis in immune cell populations. Under normal conditions, apoptosis is required in order to maintain homeostasis and it involves morphological changes (i.e., cell shrinkage, nuclear fragmentation and membrane blebbing) as well as molecular changes (i.e., induction of caspases and cytochrome c leakage) [14]. The extrinsic pathway of apoptosis is initiated with the ligation of death receptors (i.e., CD95) on the cell surface, leading to activation of major caspases, such as caspase 3, 8 and 10. The intrinsic pathway of apoptosis is initiated via mitochondria and caspase 9; cytochrome c and caspase 3 are the major players in the induction of cell death [14,15].
Δ9-THC and its apoptotic effects on immune cell populations have been studied extensively: in 1998, Zhu et al. demonstrated that in vitro THC induced apoptosis in murine macrophages and T cells. This study also showed that the process was mediated via activation of Bcl-2 and caspases [16]. It was difficult to demonstrate the apoptotic effects of THC on lymphocytes, in vivo, and our laboratory speculated that this might be due to rapid clearance of dead cells by phagocytic cells. Therefore, we exposed C57BL/6 mice to 10 mg/kg bodyweight THC and, after several time points, (4, 6, 24 and 72 h), obtained lymphocytes from the thymus and spleen of these animals. The cells were incubated for 12–24 h ex vivo and, since the phagocytosis was excluded in the cultures, we detected significant levels of THC-induced apoptosis in T cells, B cells and macrophages [17]. We have also demonstrated that THC induced higher levels of apoptosis in naive lymphocytes, when compared with mitogen-activated lymphocytes, because activated cells downregulated the levels of CB2 on their cell surface [17]. Several studies also reported THC-induced apoptosis in antigen-presenting cells. In bone marrow-derived dendritic cells (DCs), THC induced apoptosis via ligation of both CB1 and CB2 and activation of caspases such as caspase 2, 8 and 9. In vivo, THC administration decreased the number of splenic DCs, as well as MHCII expression by DCs [18,19]. Furthermore, THC increased Bcl-2 and caspase 1 activity in naive and lipopolysaccharide (LPS)-activated macrophages isolated from the peritoneal cavity of mice [16].
Other natural and synthetic cannabinoid compounds (CBD, AEA, ajulemic acid [AjA] and JWH-015), whose structures are depicted in Table 1, have also been shown to induce apoptosis in murine and human T lymphocytes. Cannabidiol, the nonpsychoactive ingredient in cannabis, induced apoptosis in CD4+ and CD8+ T cells at 4–8-μM concentrations by increasing reactive oxygen species (ROS) production as well as caspase 3 and 8 activity [20].
Ajulemic acid, a side-chain synthetic analog of Δ(8)-THC-11-oic acid, has been shown to induce apoptosis in human peripheral blood T lymphocytes via the intrinsic pathway at concentrations of 1, 3 and 10 μM [21]. In addition, the use of synthetic CB2 agonist JWH-015 treatment in vitro led to cell death via both the death-receptor pathway and the intrinsic pathway. When JWH-015 was administered in vivo, the antigen-specific response to Staphylococcal enterotoxin A was inhibited significantly [22].
It is important to note that, unlike in immune cells, cannabinoids can protect from apoptosis in nontransformed cells of the CNS, which can play a protective role in autoimmune conditions such as multiple sclerosis. Cannabinoids protect against apoptosis of oligodendrocytes via CB1 and CB2 receptors, by signaling through the PI3K/AKT pathway. In vivo and in vitro exposure to arachidonyl-2-ethylamide (ACEA) and WIN55,212-12 protected the cells, while pretreatment with CB1 receptor antagonist SR141716A and CB2 receptor antagonist SR144528 blocked the action of these cannabinoids [23]. In a different study by Jackson et al., 3D mouse brain aggregate cell cultures were compared between wild-type mice and CB1 receptor knockout mice. IFN-γ treatment led to decrease in the neurofilament-H expression in knockout cultures but not in wild-type cultures. In addition, caspase 3 activation was higher in knockout cultures, indicating a protective role of CB1 in neuronal cells [24].
Cannabinoid action on cytokines
Cytokines are the signaling proteins synthesized and secreted by immune cells upon stimulation. They are the modulating factors that balance initiation and resolution of inflammation. One of the possible mechanisms of immune control by cannabinoids during inflammation is the dys-regulation of cytokine production by immune cells and disruption of the well-regulated immune response [25]. Furthermore, cannabinoids may affect immune responses and host resistance by perturbing the balance between the cytokines produced by T-helper subsets, Th1 and Th2. In vitro studies were performed to compare the effect of THC and cannabinol on cytokine production by human T, B, CD8+, NK and eosinophilic cell lines. However, the results were variable, depending on the cell line and the concentration used [26]. Both pro-inflammatory and anti-inflammatory effects of THC were demonstrated in this study, proposing that different cell populations have varied thresholds of response to cannabinoids. Generally, TNF-α, GM-CSF and IFN-γ levels decreased with drug treatment. Interestingly, while the anti-inflammatory cytokine IL-10 decreased following THC treatment, there was an increase in the proinflammatory cytokine IL-8. In other studies, cannabinoid CP55,940 at nanomolar concentrations was shown to have a stimulatory effect on several cytokines in the human promyelocytic cell line HL-60 [27]. At the molecular level, THC has also been shown to inhibit LPS-stimulated mRNA expression of IL-1α, IL-1β, IL-6 and TNF-α in cultured rat microglial cells; however, the effect was independent of the cannabinoid receptors [28]. In a different study, mice were challenged with Corynebacterium parvum, in vivo, following the administration of the synthetic cannabinoids WIN55,212-2 and HU210. The animals were then challenged with LPS. The results showed decreased levels of TNF-α and IL-12 but increased levels of IL-10 in the serum [29]. This effect was shown to be CB1 receptor dependent.
During chronic inflammation, IL-6 suppression can decrease tissue injury [30]. AjA has been reported to prevent joint-tissue injury in animal models of adjuvant arthritis [31]. Recent studies showed that addition of AjA to human monocyte-derived macrophages in vitro reduced the secretion of IL-6 from activated cells, suggesting that AjA may have a value for treatment of joint inflammation in patients with systemic lupus erythematosus (SLE), rheumatoid arthritis (RA) and osteoarthritis [32]. It has been observed that the CB2 agonist HU-308 attenuated the hepatic ischemia/reperfusion injury by decreasing the levels of TNF-α, MIP-1α and MIP-2 in the serum and in liver homogenates [33]. Recent in vitro studies have also shown the potent anti-inflammatory effect of synthetic cannabinoids (CP55,940 and WIN55,212-2). Both CP55,940 and WIN55,212-2 downregulated IL-6 and IL-8 cytokine production from IL-1β-stimulated rheumatoid fibroblast-like synoviocytes (FLS), via a non-CB1/CB2-mediated mechanism [34].
Endocannabinoids have also been reported to affect the cytokine biology of various cell systems. Antiproliferative effects of endocannabinoids on cancer cell lines are well established and are discussed in the later section of the review. However, AEA has also been reported to increase cytokine-induced proliferation. Mouse bone marrow cells, when cultured in the presence of IL-3 and AEA, were observed to produce more hematopoietic colonies than with IL-3 alone [35]. Significant suppression of IL-2 expression by 2-AG and the nonhydrolyzable 2-AG ether was observed in leukocytes via activation of peroxisome proliferator-activated receptor-γ (PPAR-γ) [36]. Furthermore, in undifferentiated and macrophage-like differentiated HL-60 cells, 2-AG induced CB2-dependent acceleration in the production of IL-8 [37]. In Theiler’s virus immune-mediated demyelinating disease, inactivation of endocannabinoids through the use of two selective inhibitors of their transport; (R)-N-oleoyl-(1′-hydroxybenzyl)-2′-ethanolamine] (OMDM2) and [(S)-N-oleoyl-(1′-hydroxybenzyl)-2′-ethanolamine (OMDM1) led to decreased production of the proinflammatory cytokines IL-1β and IL-12 [38]. On a contrary note, cytokines have also been shown to affect the endocannabinoid system. IL-12 and IFN-γ have been shown to reduce FAAH activity and protein expression of FAAH, whereas IL-4 or IL-10 stimulated FAAH activity [39]. Table 2 provides a summary of the effect of cannabinoids on cytokines and chemokines in various cell models [26,28,29,32–34,37,40,41].
Table 2.
Effect of cannabinoids on cytokine and chemokine production.
| Cannabinod | Receptor | Cell/tissue/serum | Effect | Ref. |
|---|---|---|---|---|
| THC | ND | Macrophage cell line (RAW264.7) | Decreases TNF-α | [40] |
| THC | ND | Peritoneal macrophages | Increases IL-1α and IL-1β | [41] |
| THC | ND | Human cell lines | Decreases TNF-α, GM-CSF and IFN-γ, IL-10 Increases IL-8 |
[26] |
| THC | CB1 and CB2 independent | Rat microglial cells | Decreases TNF-α, IL-1α, IL-1β and IL-6 | [28] |
| In vivo WIN55,212-2 and HU-210 | CB1 dependent | Serum | Decreases TNF-α, IL-12 Increases IL-10 |
[29] |
| Ajulemic acid | ND | Human synovial monocyte-derived macrophage | Decreases IL-6 and IL-1β | [32] |
| HU-308 | CB2 dependent | Serum and liver homogenates | Decreases TNF-α, MIP-1α and MIP-2 | [33] |
| CP55,940 WIN55,212–2 | CB1 and CB2 independent | Rheumatoid fibroblast-like synoviocytes | IL-6 and IL-8 | [34] |
| 2-AG | CB2 dependent | Promyelocytic leukemia cell line (HL-60) | Increases IL-8, CXCL8 and CCL2 | [37] |
AG: Arachidonoylglycerol; CB: Cannabinoid receptor; CCL: CC-chemokine ligand; CXCL8: CXC-chemokine ligand 8; ND: Not determined; THC: Tetrahydrocannabinol.
Cannabinoids & multiple sclerosis
Multiple sclerosis (MS) is an autoimmune disorder that is mediated by myelin-specific self-reactive T cells, macrophages/microglial cells and astrocytes [3,42]. The action of these cells leads to the demyelination of nerve fibers and axons in the CNS of humans and results in many signs and symptoms, such as muscle spasms, tremor, ataxia, weakness or paralysis, constipation and loss of bladder control [42]. There is both anecdotal and clinical evidence to show the effectiveness of cannabinoids in the treatment of MS. In 1994, a survey of 112 MS patients (57 men and 55 women) from the USA and UK was conducted; all of the patients were self-medicating with a form of cannabis. The results of the survey showed that cannabis use improved symptoms such as spasticity, pain, tremor and depression in more than 90% of patients. In eight different clinical studies, MS patients have also reported the benefits of THC (administered via ingestion, inhalation, injection or rectal suppository), cannabis (administered via ingestion or inhalation) and the cannabinoid receptor agonist Nabilone™, (administered via ingestion) in treating spasticity, pain, tremor and ataxia [43]. Use of cannabinoids also improved objective test results such as hand-writing tests and bladder control tests [43,44]. In general, cannabinoids are useful in treating MS because they have neuroprotective as well as immunosuppressive properties [44,45]. In this section, we will focus on the latter and discuss the action of endogenous, natural and synthetic cannabinoids on immune cells within the CNS during MS.
The destruction of the blood–brain barrier in MS is initiated by myelin-specific self-reactive T cells. Infiltration of these cells into the spinal cord and CNS, and their subsequent activation, leads to the elimination of the myelin sheath around the nerves and axons [46,47]. The myelin- specific T cells are usually CD4+, IL-2R+ or MHCII-restricted Th1 cells and they secrete proinflammatory cytokines such as IFN-γ and TNF-α [47]. More recently, Th17 cells have been shown to be involved in the pathogenesis of MS [48,49]. One mechanism of immunosuppression by cannabinoids is the induction of apoptosis and Sanchez et al. demonstrated that WIN55,212-2 blocks a passive form of experimental authoimmune encephalomyelitis (EAE) by inducing apoptosis in encephalitogenic cells through partial activation of the CB2 receptor [50]. A CB1-mediated suppressive pathway has also been shown in myelin-specific T cells [24]. This study demonstrated that ex vivo WIN55,212-2 inhibited T-cell recall response to myelin oligodendrocyte glycoprotein (MOG) peptide, as well as decreasing IL-2, IFN-γ and TNF-α production by MOG-activated T cells. Other synthetic cannabinoids, such as JWH-015 and ACEA, also decreased the number of CD4+ infiltrates in the spinal cord of Theiler’s murine encephalomyelitis virus (TMEV)-infected mice [51]. Mestre et al. showed that decreased infiltration of CD4+ T cells upon WIN55,212-2 treatment in EAE mice is due to decreased intercellular and vascular cell adhesion molecules (ICAM-1 and VCAM-1) expression by endothelial cells. Another novel finding of this study demonstrated that WIN55,212-2 exerted its effects by acting through nuclear receptor PPAR-γ [52].
Microglial cells are the macrophages of the CNS and, during MS, they mediate tissue injury in two main ways: antigen presentation and cytokine/chemokine secretion [51,52]. In the initial stages of inflammation, after activation, microglial cells present antigens to myelin-specific T cells, which results in the activation and proliferation of Th1 lineage cells. Arevalo-Martin et al. demonstrated that cannabinoid agonists WIN55,212-2, ACEA or JWH-015 inhibited the activation of microglial cells by TMEV [51]. The investigators confirmed this finding by studying the morphology of the cells (reactive vs resting) as well as by immunohistochemistry. They showed that, after TMEV activation, MHCII molecules co-localized with Mac-1 in the spinal cord sections; however, after 1-day treatment with various cannabinoid agonists, MHCII expression almost disappeared. During this initial stage, co-stimulatory molecule expression, such as that of CD40, also increased and resulted in TNF-α production via the MAPK and JAK/STAT pathways. Ehrhart and colleagues demonstrated that selective stimulation of the CB2 receptor with JWH-015 on murine microglial cells decreased CD40 expression upon IFN-γ activation. This inhibition in CD40 levels translated into decreased JAK/STAT phosphorylation, and decreased TNF-α and nitric oxide production [53].
In the later stages of disease, microglial cells secrete IL-12, IL-13 and IL-23, nitric oxide and glutamate and contribute to myelin sheath destruction. IL-12 drives the proliferation of Th1 cells while IL-23 is important in the maintenance of Th17 cells. A recent study by Correa et al. showed that the endogenous cannabinoid AEA inhibited the expression of IL-12 as well as IL-23 in LPS/IFN-γ-activated human and murine microglia. This inhibition of cytokine production occurred via activation of CB2 and signaling through ERK1/2 and JNK pathways [54]. Palazuelos et al. also showed that the CB2 receptor is involved in myeloid progenitor trafficking, which is necessary for microglia replenishment and activation during MS. Their studies demonstrated that CB2−/− mice had exacerbated EAE symptoms and CD34+ myeloid progenitor cells had greatly infiltrated into the spinal cords of these animals. As an explanation for the mechanism, they showed that, in the bone marrow, CB2 receptor manipulation with HU-308 increased the expression of chemokines and their receptors (CCL2, CCL3, CCL5, CCR1 and CCR2), which are important in trafficking of progenitor cells into the neuroinflamed tissue [55].
Cannabinoids also exert their immunosuppressive effects on astrocytes. Astrocytes make up 60–70% of brain cells in the CNS and play important roles in neuronal growth, neuronal signaling, glucose metabolism and glutamate removal [54]. During disease progression, astrocytes are activated to secrete cytokines, chemokines and nitric oxide, thereby contributing to the overall inflammatory response. Because astrocytes express both CB1 and CB2 receptors, several studies investigated the inhibitory role of cannabinoids on this cell population in the context of MS. One study investigated the effects of AEA on TMEV-activated primary murine astrocytes. This study showed that AEA stimulated astrocytes and triggered the production of IL-6 in a CB1-mediated pathway [56]. The precise role of IL-6 in the CNS is still unclear; however, it has been reported that IL-6 secretion potentiates neuronal growth factor production. In addition, IL-6 has been shown to inhibit TNF-α production by IFN-γ/IL-1β-stimulated glial cells [57]. In a different study, Molina-Holgado and coworkers showed that AEA and the synthetic CB1 agonist CP-55940 inhibited nitric oxide production by LPS-stimulated astrocytes isolated from 1-day-old mice in a CB1-dependent manner [23]. In 2005, Sheng et al. demonstrated that human fetal astrocytes express both CB1 and CB2 receptors and that treatment of IL-1β-stimulated astrocytes with WIN55,212-2 decreased inflammatory products including nitric oxide, TNF-α, CXCL10, CCL2 and CCL5 (Figure 1) [54].
Figure 1. Cannabinoids and multiple sclerosis.

The three main cell types that are involved in demyelination of the nerve fibers and axons in the CNS include activated T-cells, microglia and astrocytes. In activated T-cells, treatment with WIN 55,212-12, AEA and JWH-015 has been shown to inhibit cytokine production, infiltration of cells into the spinal cord and in vitro recall response to myelin oligodendrocyte glycoprotein by T-cells. Cannabinoids also inhibit the antigen presenting abilities of microglia by downregulating MHCII expression, costimulatory molecule CD40 expression, as well as cytokine secretion. Astrocytes, the major cell population in the brain, are also affected, as cannabinoid binding to the receptors leads to inhibition of inflammatory molecules, such as nitric oxide, cytokines and chemokines. In addition, anandamide binding leads to secretion of neural growth factor secretion and protection of the neurons in the CNS.
ACEA: Arachidonyl-2-ethylamide; NGF: Neuronal growth factor.
Cannabinoids & colitis
During inflammation, several different cellular pathways are activated in the intestinal tract, leading to a pathological state [58]. Functional CB1 receptor has been shown to be expressed in the human ileum and colon and the number of CB1-expressing cells was found to be significantly increased after inflammation [59,60]. A protective role for these CB1 receptors during inflammation has been shown in a study analyzing the role of the endogenous cannabinoid system in the development of experimental colitis in mice, induced by intrarectal 2,4-dinitrobenzene sulfonic acid (DNBS) treatment or oral dextran sodium sulfate (DSS) administration [59]. The DSS model, originally reported by Okayasu et al., has been used to investigate the role of leukocytes in the development of colitis [61]. Oral administration of 5% DSS in drinking water can induce acute colitis due to chemical injury in the colon. Furthermore, long-term DSS administration produces colorectal carcinoma, which is similar to the dysplasia–carcinoma sequence seen in the course of cancer development in human ulcerative colitis [62]. On the other hand, intestinal inflammation induced by the intrarectal administration of DNBS has many of the characteristic features of Crohn’s disease in humans, involving induction of an IL-12-driven inflammation with a massive Th1-mediated response [63]. The involvement of the endogenous cannabinoid system in the modulation of the acute phase of DNBS-induced colitis was further supported by the increased levels of transcripts coding for CB1 in wild-type mice after induction of inflammation. It was observed that genetic ablation of CB1 receptors rendered mice more sensitive to inflammatory insults. Furthermore, similar to results observed in CB1-deficient mice, pharmacological blockade of CB1 with the specific antagonist SR141716A led to a worsening of colitis [59]. The protective role of the endogenous cannabinoid system was observed 24 h after DNBS treatment and became more evident on days 2 and 3. However, increased spontaneous spiking activity of smooth muscle cell membrane in DNBS-treated colons from CB1−/− mice was already visible 8 h after DNBS treatment, indicating that inflammation-induced irritation of smooth muscle occurred at an earlier stage than in wild-type mice. This gives further support to the notion that the endogenous cannabinoid system is protective against inflammatory changes. These data indicated that the activation of CB1 and the endogenous cannabinoid system is an early and important physiological step in self-protection of the colon against inflammation.
Pharmacological stimulation of cannabinoid receptors with the potent agonist HU210 also induced a reduction of experimental colitis. It has been reported that cannabinoid receptor stimulation could have a beneficial effect on experimental colitis [64]. Intraperitoneal application of ACEA, a CB1-selective agonist, and JWH-133, a CB2-selective agonist, inhibited oil of mustard (OM)-induced colitis and subsequent symptoms such as induced distal colon weight gain, colon shrinkage, inflammatory damage, diarrhea and histological damage. This study demonstrated a role for CB2 activation in experimental colitis. The fact that both CB1 and CB2 agonists are active in colitis models lends additional support to the theory that signaling through cannabinoid receptors may mediate protective mechanisms in colitis.
In the small intestine, the involvement of CB1 receptors in the control of intestinal motility during croton oil-induced inflammation was recently demonstrated. Izzo et al. showed that pharmacological administration of cannabinoids is able to delay gastrointestinal transit in croton oil-treated mice [65]. It was further suggested that increased levels of CB1 receptor expression in inflamed jejuna may contribute to this protective effect. CB1 receptors were shown to modulate gastrointestinal motility during croton oil-induced inflammation in mice.
Fatty acid amide hydrolase is the major enzyme involved in the degradation of several bioactive fatty amides, in particular anandamide [11], and its genetic deletion in mice leads to a strongly decreased ability to degrade this endocannabinoid and an increase of anandamide levels in several tissues [66]. FAAH-deficient mice showed significant protection against DNBS treatment. However, because anandamide is believed to act not only through cannabinoid receptors but also through other targets, including the peripheral vanilloid receptor TRPV1 [67], the decreased inflammation in FAAH−/− mice could also be due to the activation of targets other than cannabinoid receptors.
In conclusion, cannabinoids have been shown to regulate the tissue response to excessive inflammation in the colon, mediated by both dampening smooth-muscular irritation caused by inflammation and suppressing proinflammatory cytokines, thus controlling the cellular pathways leading to inflammatory responses. These results strongly suggest that modulation of the physiological activity of the cannabinoid system during colonic inflammation might be a promising therapeutic tool for the treatment of several diseases characterized by inflammation of the GI tract.
Cannabinoid system & liver injury
During the past few years, awareness of the cannabinoid system in the pathophysiology of liver disease has gained momentum. Both CB1 and CB2 receptors have been shown to be upregulated in the early stages of liver injury [68–72]. Although embryonic liver has been shown to express CB2 receptor mRNA, adult liver hepatocytes and endothelial cells displayed only a faint physiological level of expression of CB1 receptors and were shown to produce low levels of endocannabinoids. CB1 receptors have been found to be upregulated in the vascular endothelium and in myofibroblasts located in fibrotic bands of cirrhotic livers in human and rodents [72]. CB2 receptors are also expressed in myofibroblasts, inflammatory cells and biliary epithelial cells [69]. There has been growing evidence in recent years to suggest that endocannabinoids may regulate the pathophysiology of liver diseases, including both acute forms of hepatic injury, liver fibrosis and cirrhosis. The endocannabinoids are found in low levels in normal liver, which may be due to high levels of expression of FAAH, which is responsible for the breakdown of AEA [11]. The levels of AEA have been shown to increase in the liver and serum during acute hepatitis and fatty liver disease [70]. In fatty liver, the increase in AEA results from decreased ability of FAAH to degrade AEA. Together, the above studies suggest that endocannabinoids and their receptors may play a critical role in regulating liver fibrogenesis; therefore, targeting the cannabinoid receptors may serve as a novel tool to prevent and treat liver injury.
While the mechanisms of inflammatory liver injury are unclear, they are accompanied by infiltration of activated polymorphonuclear leukocytes, activation of Kupffer cells, production of proinflammatory cytokines and generation of ROS. Many recent studies indicated strongly the increased upregulation of the endocannabinoid system during liver diseases involving hepatocyte injury, inflammation, fibrogenesis, hepatic encephalopathy, cirrhotic cardiomyopathy and portal hypertension [73]. The role of hepatic expression of anandamide and 2-AG is apparent in hepatic ischemia-reperfusion (I/R) injury, in which their levels are significantly increased, correlating with the extent of liver damage. Moreover, pretreatment of mice with JWH-133, a CB2 receptor agonist, was shown to decrease the degree of liver tissue injury and inflammatory cell infiltration and decrease serum levels of cytokines, chemokines and adhesion molecules [74]. Furthermore, CB2−/− receptor mice were shown to develop greater inflammation and I/R-induced liver damage than their wild-type counterparts. The data also highlights the protective role of CB2 receptor activation in the inflammatory response associated with chronic liver diseases such as viral hepatitis and alcoholic or nonalcoholic fatty liver diseases.
Viral hepatitis, alcohol abuse and nonalcoholic fatty liver are some of the conditions that can induce chronic liver injury and inflammation, leading to activation of fibrogenesis as a wound-healing mechanism. However, persistence of fibrogenic stimuli can enhance deposition of the extracellular matrix by hepatic myofibroblasts, thus disrupting normal liver architecture and, ultimately, leading to cirrhosis and liver failure. CB1 and CB2 receptors are shown to be markedly upregulated in cirrhotic human liver samples, demonstrating the impact of endocannabinoids in liver fibrogenesis. In addition, increases in circulating levels of anadamide and hepatic 2-AG have also been reported in cirrhosis and liver fibrosis, respectively [73]. CB2−/− mice exposed to CCl4 showed enhanced liver fibrosis when compared with wild-type mice, thereby suggesting a protective role for CB2 receptor activation in liver fibrosis. By contrast, activation of CB1 receptors was found to promote profibrotic response [72]. The pharmacological inactivation of CB1 with rimonabant® (SR141716) results in the reduction of obesity and hepatic steatosis in rodents [75], demonstrating that CB1 and CB2 receptors exert opposite effects on liver fibrosis and further suggesting that endocannabinoid system regulates both pro- and anti-fibrogenic responses in the liver. Further effects of the endocannabinoids have also been shown to be receptor independent. AEA and 2-AG have been shown to induce necrosis and apoptosis, respectively, in activated hepatic stellate cells, through increased generation of ROS [76].
The abuse of cannabis has been shown to promote liver fibrosis in patients with chronic hepatitis C, indicating that cannabinoids may exacerbate liver fibrogenesis and that CB1 receptor antagonists may play a role as anti-fibrosing molecules [71]. However, an alternative explanation could be that marijuana can trigger immunosuppression. For example, CB2 activation in immune cells can trigger apoptosis and this, in turn, can have an immunosuppressive effect in patients with hepatitis C. As such patients require immunocompetent cells to keep hepatitis under control, chronic marijuana abuse may promote fibrogenesis through the activation of CB2 and consequent suppression of antiviral immunity [77].
Endocannabinoids may also regulate liver cirrhosis by acting as mediators of vascular and cardiac functions. Endocannabinoids can trigger vasorelaxation, while an upregulated CB1-mediated cannabinoid tone causes enhanced mesenteric vasodialation leading to portal hypertension [73,75]. A recent in vivo study by Batkai et al. in rats with CCl4-induced cirrhosis, indicated that increased local production of AEA mediated the inhibition of β-adrenergic responsiveness. Further improvement in contractile function of isolated papillary muscles was observed following treatment with AM251, a CB1 receptor antagonist, suggesting therapeutic potential against cirrhotic cardiomyopathy [75].
There are limited, but reliable, data on the neuroprotective role of the endocannabinoid system in hepatic encephalopathy. It has been demonstrated in a murine model that, during fulminant hepatic failure, levels of 2-AG in the brain are elevated, potentially as a response to liver damage. The administration of the CB2 endogenous ligand 2-AG, an antagonist of CB1 receptor SR141716A or an agonist of CB2 receptor HU308, resulted in a marked improvement in neurological score. Thus, influencing the endocannabinoid system with exogenous cannabinoid derivates specific for the CB1 or CB2 receptor may have a beneficial therapeutic effect on neurological dysfunction in liver diseases [78]. Recently, we noted that both exogenous and endogenous cannabinoids protected mice from concanavalin-A (ConA)-induced acute hepatitis, a model that mimics viral or autoimmune hepatitis, in which T cells play a critical role in triggering liver injury. We found that administration of a single dose of THC or anandamide could ameliorate Con-A-induced hepatitis. We found that this effect was mediated through multiple pathways, including suppression of pro-inflammatory cytokines, induction of apoptosis in activated T cells and induction of forkhead helix transcription factor p3(Foxp3)+ Treg cells [79]. This overwhelming evidence shows that the cannabinoid system must play a major role in the pathophysiology of various liver diseases and its therapeutic potential should be exploited for the treatment of chronic liver injuries (Figure 2).
Figure 2.

Endocannabinoids, CB1 antagonists and CB2 agonists as potential drugs for the treatment of liver injury.
Cannabinoids & RA
Rheumatoid arthritis is a chronic inflammatory disease that affects approximately 1% of the human population and is manifested in joint destruction, deformity and loss of function associated with joint stiffness, pain, swelling and tenderness [80]. The major immune cell populations involved in joint injury are macrophages, T cells, fibroblast-like synoviocytes and DCs. Major cytokines are TNF-α and IL-1 [80,81]. Cannabinoids and their anti-inflammatory properties have been studied in animal models of RA and on human cells from RA patients and these studies demonstrate the anti-arthritic properties of these natural plant compounds [32, 82–84]. Interestingly, most of the studies on RA and cannabinoids focus on the use of nonpsychoactive cannabinoids.
CBD is the major nonpsychoactive component of the cannabis plant and its protective effect has been shown in murine collagen-induced arthritis [85]. In this study, the authors showed that daily oral (5 mg/kg) or intra-peritoneal (25 mg/kg) administration of CBD inhibited disease progression. Furthermore, the study demonstrated that CBD-treated mice had less proliferation in ex vivo-activated draining lymph node cells, decreased levels of IFN-γ secreted by the activated lymph node cells and diminished TNF-α production by knee synovial cells. Sumariwalla et al. used another synthetic nonpsychoactive cannabinoid, HU-320, and demonstrated that this compound improved already established arthritis in mice [86]. Lymph node cells from HU-320-treated mice showed decreased proliferative responses when the cells from 7-day post-inflammation mice were incubated with collagen II.
Furthermore, AjA has been shown to inhibit IL-1β production by human monocytes isolated from RA patients [82]. In this study, the investigators isolated peripheral blood monocytes (PBMs) as well as synovial fluid monocytes (SFMs) from normal and RA patients. The isolated cells were pretreated with 0–30 μM AjA and were then stimulated with LPS. The results showed that AjA treatment decreased IL-1β production in both PBMs and SFMs, but did not affect TNF-α production. In a different study, Parker et al. demonstrated that AjA reduced IL-6 secretion by activated human monocyte-derived-macrophages [32].
AjA also exerts its immunomodulatory effects by inducing apoptosis in mature osteoclast-like cells and, therefore, protecting the host from osteoclastogenesis. George et al. demonstrated that treatment of osteoclast-like cells with 15 and 30 μM of AjA led to induction of caspase 3 and 8 in these cells, resulting in apoptosis and protection from osteoclastogenesis [83].
Cannabinoids & cancers with inflammatory components
The role of inflammation in evolution of certain types of cancer has been strongly suggested, linking the inflammatory response to 15–20% of all deaths from cancer worldwide [84]. The hallmarks of cancer-related inflammation include the presence of inflammatory cells in tumor tissue, and the regulation of tumor growth, metastasis and angiogenesis by inflammatory mediators (e.g., chemokines, cytokines and prostaglandins). The connection between inflammation and cancer is now generally accepted and nonsteroidal anti-inflammatory drugs have been shown to reduce varied cancer risk. Use of these drugs reduces colon cancer risk by 40–50% and is indicated to be preventative for lung, esophagus and stomach cancer [87]. Hence, inflammation can be considered as a therapeutic opportunity in certain types of cancer. Recent applications of cannabinoids have been extended as antitumor agents [1,88], which relies on their ability to inhibit tumor angiogenesis [89] or induce direct apoptosis or cell cycle arrest in neoplastic cells [89–92]. A focus on the antiproliferative effects of these compounds in various tumors, such as breast and prostate cancers, pheochromocytoma and malignant gliomas, has been proposed [1,92–94]. Our laboratory reported that, in vitro, THC and other cannabinoids could induce apoptosis in transformed murine and human T cells [95], including primary acute lymphoblastic human leukemia cells. Furthermore, the treatment of mice bearing a T-cell leukemia with THC could cure approximately 25% of the mice [95]. We further showed that treatment with THC caused interruption of the MAPK/ERK kinase/ERK signaling module that was required for apoptotic lethality [96,97]. The role of endocannabinoids as potential endogenous tumor growth inhibitors has been suggested in a study where it was observed that levels of both AEA and 2-AG were higher in precancerous polyps than in fully developed carcinomas in the colon [98]. Recent in vivo studies proposed that selective targeting of CB2 receptors resulted in colorectal tumor growth inhibition via apoptosis, which was mediated through the stimulation of ceramide [98]. In a xenograft model of thyroid cancer, substances that blocked endocannabinoid degradation also increased the levels of AEA and 2-AG in the tissue and reduced tumor growth [99]. Various attempts have been made to inactivate cannabinoid-degrading enzymes, thereby increasing the local concentration of endocannabinoids at the tumor cell surface. This leads to anti-tumor effects of CB receptor signaling in various cancer types, such as thyroid, brain and prostate cancer [99–103]. Although the majority of the effects of cannabinoids are CB receptor mediated, AEA has been shown to induce its effects on cancerous cells by interacting with TRPV1 receptor [104,105] or cholesterol-rich lipid rafts [106]. Furthermore, it has been reported that signaling pathways are differentially regulated by cannabinoids in normal cells versus cancer cells. In malignancies, such as thyroid cancer, lymphoma, melanoma, pancreas and breast cancer, the levels of cannabinoid receptors are often higher in the tumor compared with normal cells of the same origin, resulting in increased sensitivity to cannabinoids in the malignancies [89,107–110]. Moreover, many animal studies have reported antiproliferative and pro-apoptotic effects of cannabinoids on tumor cells but not on normal tissue [89,91]. Thus, the role of the cannabinoid system in cancer indicates that this system is involved in regulating many of the functions that are essential in cancer development.
Cannabinoids & other inflammatory diseases
Allergic asthma is a complex inflammatory disorder characterized by airway hyper-responsiveness, elevated serum IgE, recruitment of eosinophils into the lung and mucus hypersecretion by goblet cells [111]. Murine models of allergic airway disease, employing ovalbumin (OVA) as an aeroallergen, indicated that CD4+ Th2 cells (IL-4, IL-5 and IL-13) played a pivotal role in the pathophysiology of the allergic airway response [112]. Intraperitoneal administration of THC or cannabinol (CBN) in OVA-sensitized and challenged A/J mice led to attenuation of serum IgE, IL-2, IL-4, IL-5 and IL-13 mRNA expression and decreased allergen-induced mucus production, indicating that cannabinoid-based compounds may represent a novel class of therapeutic agents for the treatment of allergic airway diseases [113]. While most studies have shown that cannabinoids, such as THC, facilitate a Th1 to Th2 cytokine switch, as discussed previously, it is surprising that cannabinoids can also suppress allergic asthma triggered primarily by Th2 cytokines. It is possible that THC may affect other cells such as DCs and B cells directly in this model. Previous findings indicated that aerosolized THC was capable of causing significant bronchodilatation with minimal systemic side effects, but had a local irritating effect on the airways [114]. Further bronchodilator effects of cannabinoids administered orally or by aerosol to asthmatic patients have also been reported [115,116]. Similarly, endogenous cannabinoids have been shown to regulate airway responsiveness. In rodent lungs, a Ca2+-activated mechanism for the biosynthesis of anandamide was observed and CB1 receptors were found predominantly on axon terminals of airway nerves, indicating that endocannabinoids may regulate bronchial smooth muscle tone [117]. It was reported that activation of CB1 receptors by locally released anandamide may participate in the control of bronchial contractility. Blocking of AEA-induced CB1 activity can enhance capsaicin-induced bronchospasm. However, the authors further suggested that the effects of AEA may depend on the state of the bronchial muscle. During capsaicin-evoked bronchospasm, AEA may reduce the muscle contraction, whereas AEA may cause bronchoconstriction in the absence of vagus nerve-constricting tone [117].
Cannabidiol has been shown to be effective in protecting endothelial function and integrity in human coronary artery endothelial cells (HCAECs). The study demonstrated that CBD reversed the harmful effects of high glucose on HCAECs by inhibiting [118]:
Reactive oxygen species production by mitochondria
NF-κB activation
Transendothelial migration of monocytes
Monocyte–endothelial adhesion in HCAECs
In a different experiment, HCAECs were stimulated with TNF-α in order to mimic the inflammatory processes during atherosclerosis, and the effect of different CB2 receptor agonists on the activated cells was studied. It was demonstrated that activation of cells with TNF-α led to increased expression of CB2 and activation of Ras, p38, MAPK, JNK and AKT pathways. In addition, proliferation and migration was markedly increased in activated cell populations. The use of CB2 agonists JWH-133 and HU-308 inhibited all activated pathways in a dose-dependent manner, establishing a novel use for these cannabinoid compounds [119].
Experimental autoimmune uveoretinitis (EAU), is a CD4+ T-cell-mediated autoimmune disease, which can be induced in rodents by challenge with retinal antigens or their peptides [120]. In a recent study, it was observed that the CB2-selective agonist JWH-133 had a high in vivo immunosuppressive effect in EAU model. EAU was strongly inhibited when the CB2 was engaged and the effects of CB2 engagement appeared to be mediated predominantly through downregulation of T-cell function with a less-marked effect on antigen presentation [121]. An impaired T-cell-proliferative response in leukocytes from JWH-133-treated mice was also accompanied by marked reductions in cytokine production. A more recent study showed that JWH-133 (10 nM–5 μM) suppressed IL-12p40 and enhanced IL-10 production in mouse macrophages induced by LPS [122]. As IL-12p40 is critical for the development of EAU and IL-10 could suppress EAU [123,124], the authors suggested that this may also be one of the possible mechanisms responsible for the effect of JWH-133 on EAU.
Insulin-dependent Type 1 diabetes mellitus (T1DM) is an autoimmune disease resulting in destruction of insulin-producing pancreatic β cells, a process that is assumed to be mediated mainly by CD4 Th1 and CD8 T lymphocytes [125]. In rodents, T1D is induced by administration of multiple low doses of streptozotocin (MLDSTZ). This model is used for studying autoimmune processes associated with pancreatic β-cell pathogenesis. A study performed by Li et al. indicated that Δ9-THC could exert a transient attenuation of MLDSTZ-induced autoimmune diabetes. Δ9-THC treated (150 mg/kg) CD-1 mice exhibited reduced hyperglycemia and a significant decrease in the loss of pancreatic insulin. MLDSTZ-induced insulitis was also significantly attenuated by decreases in CD3+ inflammatory cells in the pancreatic islets and in mRNA expression for IL-12, IFN-γ and TNF-α. It was suggested that in this model, the autoimmune component was most effectively modulated by Δ9-THC treatment [126]. Similarly, CBD treatment has been shown to significantly inhibit and delay destructive insulitis and inflammatory Th1-associated cytokine production in nonobese diabetes-prone (NOD) female mice. CBD-treated mice exhibited significant reduction of plasma levels of the proinflammatory cytokines IFN-γ and TNF-α, whereas production of the Th2-associated cytokines IL-4 and IL-10 was increased when compared with untreated control mice, thus shifting the immune response from Th1 to Th2 dominance [127]. A recent study indicated that treatment of 11–14-week-old female NOD mice, either in a latent diabetes stage (after 14 weeks) or with initial symptoms of diabetes (appearing up to 14 weeks) with CBD for 4 weeks, could lead to sustained inhibition of insulitis [128]. CBD treatment inhibited specific destruction of the islets and reduced the infiltrates by mononuclear cells into the islets, thus preventing diabetes. Furthermore, cannabinoids have also been demonstrated to possess additional beneficial effects in animal models of diabetes. It has been reported that rats treated with CBD for periods of 1–4 weeks experienced significant protection from diabetic retinopathy [129]. Cannabinoids have also been shown to alleviate neuropathic pain associated with the disease. Mice injected with a cannabis receptor agonist experienced a reduction in diabetic-related tactile allodynia compared with nontreated controls [130]. Thus, cannabinoids can be considered useful for controlling T1D due to their anti-inflammatory properties.
Future perspective
It is becoming increasingly clear that cannabinoid receptors and their endogenous ligands play a crucial role in the regulation of the immune system. Exogenous cannabinoids have been shown to suppress T-cell-mediated immune responses by primarily inducing apoptosis and suppressing inflammatory cytokines and chemokines. Such observations indicate that targeting cannabinoid receptor–ligand interactions may constitute a novel window of opportunity to treat inflammatory and autoimmune disorders. As CB2 receptors are primarily expressed on immune cells, targeting CB2 may result in selective immunomodulation without overt toxicity. The future challenges for the use of cannabinoids as anti-inflammatory drugs include synthesis of cannabinoid receptor agonists that are nonpsychoactive with anti-inflammatory activity and then identifying their mode of action. Although current studies suggest that cannabinoids are useful therapeutic agents in the treatment of various inflammatory disorders, further evaluation of the mechanisms that account for their anti-inflammatory properties is necessary. Such studies may involve the use of cannabinoid receptor-knockout mice and use of receptor-specific compounds. While most studies have focused on the effect of cannabinoids on cytokines, apoptosis and Th1 cell functions in the past, additional investigations on their effect on Th17 cells, DCs, NK cells, B cells and Fox-P3+ regulatory T cells is critical as such cells play an important role in the regulation and mediation of inflammatory or autoimmune disease response. Whether endocannabinoids and cannabinoid receptors play a critical role during normal inflammatory response also requires further consideration. Moreover, cannabinoid receptor signaling and effect of cannabinoids on adhesion molecules, co-stimulatory molecules and chemokines require further study in order to increase our understanding of cannabinoids and their intricate effects on immune system disorders. Overall, cannabinoids have exhibited significant potential to be used as novel anti-inflammatory agents and specific targeting of CB2 receptors holds the promise of mediating immunosuppressive effects without exerting psychotropic side effects.
Executive summary
Cannabinoids, the active components of Cannabis sativa, and endogenous cannabinoids mediate their effects through activation of specific cannabinoid receptors known as cannabinoid receptor 1 and 2 (CB1 and CB2).
The cannabinoid system has been shown both in vivo and in vitro to be involved in regulating the immune system through its immunomodulatory properties.
Cannabinoids suppress inflammatory response and subsequently attenuate disease symptoms. This property of cannabinoids is mediated through multiple pathways such as induction of apoptosis in activated immune cells, suppression of cytokines and chemokines at inflammatory sites and upregulation of FoxP3+ regulatory T cells.
Cannabinoids have been tested in several experimental models of autoimmune disorders such as multiple sclerosis, rheumatoid arthritis, colitis and hepatitis and have been shown to protect the host from the pathogenesis through induction of multiple anti-inflammatory pathways.
Cannabinoids may also be beneficial in certain types of cancers that are triggered by chronic inflammation. In such instances, cannabinoids can either directly inhibit tumor growth or suppress inflammation and tumor angiogenesis.
Acknowledgments
This work was supported in part by NIH grants R01AI053703, R01ES09098, R01AI058300, R01DA016545, R01HL058641 and P01AT003961. The authors have no other relevant affiliations or financial involvement with any organization or entity with a financial interest in or financial conflict with the subject matter or materials discussed in the manuscript. This includes employment, consultancies, honoraria, stock ownership or options, expert testimony, grants or patents received or pending, or royalties.
- Cannabinoids
Active components extracted from Cannabis sativa that act through cannabinoid receptors
- Cannabinoid receptors
G-protein-coupled receptors that mediate the action of cannabinoids
- Cytokines
Signaling proteins synthesized and secreted by immune cells upon activation
- Autoimmune diseases
Disorders in which the immune system starts recognizing an individual’s own cells as foreign and mounts a reaction, leading to tissue injury
- Inflammation
A defense mechanism that results from the action of activated immune cells in response to foreign antigens. Sometimes, response to self antigens can trigger severe tissue injury
Footnotes
For reprint orders, please contact reprints@futurescience.com
Financial & competing interests disclosure
No writing assistance was utilized in the production of this manuscript.
Bibliography
- 1.Guzman M. Cannabinoids: potential anticancer agents. Nat Rev Cancer. 2003;3:745–755. doi: 10.1038/nrc1188. [DOI] [PubMed] [Google Scholar]
- 2.Inui A. Cancer anorexia-cachexia syndrome: current issues in research and management. CA Cancer J Clin. 2002;52:72–91. doi: 10.3322/canjclin.52.2.72. [DOI] [PubMed] [Google Scholar]
- 3.Pollmann W, Feneberg W. Current management of pain associated with multiple sclerosis. CNS Drugs. 2008;22:291–324. doi: 10.2165/00023210-200822040-00003. [DOI] [PubMed] [Google Scholar]
- 4.Tramer MR, Carroll D, Campbell FA, Reynolds DJ, Moore RA, McQuay HJ. Cannabinoids for control of chemotherapy induced nausea and vomiting: quantitative systematic review. BMJ. 2001;323:16–21. doi: 10.1136/bmj.323.7303.16. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Croxford JL, Yamamura T. Cannabinoids and the immune system: potential for the treatment of inflammatory diseases? J Neuroimmunol. 2005;166:3–18. doi: 10.1016/j.jneuroim.2005.04.023. [DOI] [PubMed] [Google Scholar]
- 6.Mackie K. Cannabinoid receptors as therapeutic targets. Annu Rev Pharmacol Toxicol. 2006;46:101–122. doi: 10.1146/annurev.pharmtox.46.120604.141254. [DOI] [PubMed] [Google Scholar]
- 7.Stefano GB, Liu Y, Goligorsky MS. Cannabinoid receptors are coupled to nitric oxide release in invertebrate immunocytes, microglia, and human monocytes. J Biol Chem. 1996;271:19238–19242. doi: 10.1074/jbc.271.32.19238. [DOI] [PubMed] [Google Scholar]
- 8.Devane WA, Hanus L, Breuer A, et al. Isolation and structure of a brain constituent that binds to the cannabinoid receptor. Science. 1992;258:1946–1949. doi: 10.1126/science.1470919. [DOI] [PubMed] [Google Scholar]
- 9.Mechoulam R, Ben-Shabat S, Hanus L, et al. Identification of an endogenous 2-monoglyceride, present in canine gut, that binds to cannabinoid receptors. Biochem Pharmacol. 1995;50:83–90. doi: 10.1016/0006-2952(95)00109-d. [DOI] [PubMed] [Google Scholar]
- 10.Sugiura T, Kondo S, Sukagawa A, et al. 2-arachidonoylglycerol: a possible endogenous cannabinoid receptor ligand in brain. Biochem Biophys Res Commun. 1995;215:89–97. doi: 10.1006/bbrc.1995.2437. [DOI] [PubMed] [Google Scholar]
- 11.Cravatt BF, Giang DK, Mayfield SP, Boger DL, Lerner RA, Gilula NB. Molecular characterization of an enzyme that degrades neuromodulatory fatty-acid amides. Nature. 1996;384:83–87. doi: 10.1038/384083a0. [DOI] [PubMed] [Google Scholar]
- 12.Kaczocha M, Glaser ST, Deutsch DG. Identification of intracellular carriers for the endocannabinoid anandamide. Proc Natl Acad Sci USA. 2009;106:6375–6380. doi: 10.1073/pnas.0901515106. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Rodriguez de Fonseca F, Del Arco I, Bermudez-Silva FJ, Bilbao A, Cippitelli A, Navarro M. The endocannabinoid system: physiology and pharmacology. Alcohol Alcoholism. 2005;40:2–14. doi: 10.1093/alcalc/agh110. [DOI] [PubMed] [Google Scholar]
- 14.Hengartner MO. The biochemistry of apoptosis. Nature. 2000;407:770–776. doi: 10.1038/35037710. [DOI] [PubMed] [Google Scholar]
- 15.Igney FH, Krammer PH. Death and anti-death: tumour resistance to apoptosis. Nat Rev Cancer. 2002;2:277–288. doi: 10.1038/nrc776. [DOI] [PubMed] [Google Scholar]
- 16.Zhu W, Friedman H, Klein TW. Δ9-tetrahydrocannabinol induces apoptosis in macrophages and lymphocytes: involvement of Bcl-2 and caspase-1. J Pharmacol Exp Ther. 1998;286:1103–1109. [PubMed] [Google Scholar]
- 17.McKallip RJ, Lombard C, Martin BR, Nagarkatti M, Nagarkatti PS. Δ(9)-tetrahydrocannabinol-induced apoptosis in the thymus and spleen as a mechanism of immunosuppression in vitro and in vivo. J Pharmacol Exp Ther. 2002;302:451–465. doi: 10.1124/jpet.102.033506. [DOI] [PubMed] [Google Scholar]
- 18.Do Y, McKallip RJ, Nagarkatti M, Nagarkatti PS. Activation through cannabinoid receptors 1 and 2 on dendritic cells triggers NF-κB-dependent apoptosis: novel role for endogenous and exogenous cannabinoids in immunoregulation. J Immunol. 2004;173:2373–2382. doi: 10.4049/jimmunol.173.4.2373. [DOI] [PubMed] [Google Scholar]
- 19.Lu T, Newton C, Perkins I, Friedman H, Klein TW. Cannabinoid treatment suppresses the T-helper cell-polarizing function of mouse dendritic cells stimulated with Legionella pneumophila infection. J Pharmacol Exp Ther. 2006;319:269–276. doi: 10.1124/jpet.106.108381. [DOI] [PubMed] [Google Scholar]
- 20.Lee CY, Wey SP, Liao MH, et al. A comparative study on cannabidiol-induced apoptosis in murine thymocytes and EL-4 thymoma cells. Int Immunopharmacol. 2008;8:732–740. doi: 10.1016/j.intimp.2008.01.018. [DOI] [PubMed] [Google Scholar]
- 21.Bidinger B, Torres R, Rossetti RG, et al. Ajulemic acid, a nonpsychoactive cannabinoid acid, induces apoptosis in human T lymphocytes. Clin Immunol. 2003;108:95–102. doi: 10.1016/s1521-6616(03)00064-0. [DOI] [PubMed] [Google Scholar]
- 22.Lombard C, Nagarkatti M, Nagarkatti P. CB2 cannabinoid receptor agonist, JWH-015, triggers apoptosis in immune cells: potential role for CB2-selective ligands as immunosuppressive agents. Clin Immunol. 2007;122:259–270. doi: 10.1016/j.clim.2006.11.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Molina-Holgado F, Molina-Holgado E, Guaza C, Rothwell NJ. Role of CB1 and CB2 receptors in the inhibitory effects of cannabinoids on lipopolysaccharide-induced nitric oxide release in astrocyte cultures. J Neurosci Res. 2002;67:829–836. doi: 10.1002/jnr.10165. [DOI] [PubMed] [Google Scholar]
- 24.Croxford JL, Pryce G, Jackson SJ, et al. Cannabinoid-mediated neuroprotection, not immunosuppression, may be more relevant to multiple sclerosis. J Neuroimmunol. 2008;193:120–129. doi: 10.1016/j.jneuroim.2007.10.024. [DOI] [PubMed] [Google Scholar]
- 25.Klein TW, Newton CA, Nakachi N, Friedman H. Δ9-tetrahydrocannabinol treatment suppresses immunity and early IFN-γ, IL-12, and IL-12 receptor β 2 responses to Legionella pneumophila infection. J Immunol. 2000;164:6461–6466. doi: 10.4049/jimmunol.164.12.6461. [DOI] [PubMed] [Google Scholar]
- 26.Srivastava MD, Srivastava BI, Brouhard B. Δ9 tetrahydrocannabinol and cannabidiol alter cytokine production by human immune cells. Immunopharmacology. 1998;40:179–185. doi: 10.1016/s0162-3109(98)00041-1. [DOI] [PubMed] [Google Scholar]
- 27.Derocq JM, Jbilo O, Bouaboula M, Segui M, Clere C, Casellas P. Genomic and functional changes induced by the activation of the peripheral cannabinoid receptor CB2 in the promyelocytic cells HL-60. Possible involvement of the CB2 receptor in cell differentiation. J Biol Chem. 2000;275:15621–15628. doi: 10.1074/jbc.275.21.15621. [DOI] [PubMed] [Google Scholar]
- 28.Puffenbarger RA, Boothe AC, Cabral GA. Cannabinoids inhibit LPS-inducible cytokine mRNA expression in rat microglial cells. Glia. 2000;29:58–69. [PubMed] [Google Scholar]
- 29.Smith SR, Terminelli C, Denhardt G. Effects of cannabinoid receptor agonist and antagonist ligands on production of inflammatory cytokines and anti-inflammatory interleukin-10 in endotoxemic mice. J Pharmacol Exp Ther. 2000;293:136–150. [PubMed] [Google Scholar]
- 30.Smolen JS, Maini RN. Interleukin-6: a new therapeutic target. Arthritis Res Ther. 2006;8(Suppl. 2):S5. doi: 10.1186/ar1969. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Zurier RB, Rossetti RG, Lane JH, Goldberg JM, Hunter SA, Burstein SH. Dimethylheptyl-THC-11 oic acid: a nonpsychoactive antiinflammatory agent with a cannabinoid template structure. Arthritis Rheum. 1998;41:163–170. doi: 10.1002/1529-0131(199801)41:1<163::AID-ART20>3.0.CO;2-9. [DOI] [PubMed] [Google Scholar]
- 32.Parker J, Atez F, Rossetti RG, Skulas A, Patel R, Zurier RB. Suppression of human macrophage interleukin-6 by a nonpsychoactive cannabinoid acid. Rheumatol Int. 2008;28:631–635. doi: 10.1007/s00296-007-0489-0. [DOI] [PubMed] [Google Scholar]
- 33.Rajesh M, Pan H, Mukhopadhyay P, et al. Cannabinoid-2 receptor agonist HU-308 protects against hepatic ischemia/reperfusion injury by attenuating oxidative stress, inflammatory response, and apoptosis. J Leukoc Biol. 2007;82:1382–1389. doi: 10.1189/jlb.0307180. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Selvi E, Lorenzini S, Garcia-Gonzalez E, et al. Inhibitory effect of synthetic cannabinoids on cytokine production in rheumatoid fibroblast-like synoviocytes. Clin Exp Rheumatol. 2008;26:574–581. [PubMed] [Google Scholar]
- 35.Valk P, Verbakel S, Vankan Y, et al. Anandamide, a natural ligand for the peripheral cannabinoid receptor is a novel synergistic growth factor for hematopoietic cells. Blood. 1997;90:1448–1457. [PubMed] [Google Scholar]
- 36.Rockwell CE, Snider NT, Thompson JT, Vanden Heuvel JP, Kaminski NE. Interleukin-2 suppression by 2-arachidonyl glycerol is mediated through peroxisome proliferator-activated receptor γ independently of cannabinoid receptors 1 and 2. Mol Pharmacol. 2006;70:101–111. doi: 10.1124/mol.105.019117. [DOI] [PubMed] [Google Scholar]
- 37.Kishimoto S, Kobayashi Y, Oka S, Gokoh M, Waku K, Sugiura T. 2-arachidonoylglycerol, an endogenous cannabinoid receptor ligand, induces accelerated production of chemokines in HL-60 cells. J Biochem. 2004;135:517–524. doi: 10.1093/jb/mvh063. [DOI] [PubMed] [Google Scholar]
- 38.Mestre L, Correa F, Arevalo-Martin A, et al. Pharmacological modulation of the endocannabinoid system in a viral model of multiple sclerosis. J Neurochem. 2005;92:1327–1339. doi: 10.1111/j.1471-4159.2004.02979.x. [DOI] [PubMed] [Google Scholar]
- 39.Maccarrone M, Valensise H, Bari M, Lazzarin N, Romanini C, Finazzi-Agro A. Progesterone up-regulates anandamide hydrolase in human lymphocytes: role of cytokines and implications for fertility. J Immunol. 2001;166:7183–7189. doi: 10.4049/jimmunol.166.12.7183. [DOI] [PubMed] [Google Scholar]
- 40.Fischer-Stenger K, Dove Pettit DA, Cabral GA. Δ9-tetrahydrocannabinol inhibition of tumor necrosis factor-α: suppression of post-translational events. J Pharmacol Exp Ther. 1993;267:1558–1565. [PubMed] [Google Scholar]
- 41.Zhu W, Newton C, Dka Y, Friedman H, Klein TW. Δ9-tetrahydrocannabinol enhances the secretion of interleukin 1 from endotoxin-stimulated macrophages. J Pharmacol Exp Ther. 1994;270:1334–1339. [PubMed] [Google Scholar]
- 42.Weiner HL. The challenge of multiple sclerosis: how do we cure a chronic heterogeneous disease? Ann Neurol. 2009;65:239–248. doi: 10.1002/ana.21640. [DOI] [PubMed] [Google Scholar]
- 43.Pertwee RG. Cannabinoids and multiple sclerosis. Pharmacol Ther. 2002;95:165–174. doi: 10.1016/s0163-7258(02)00255-3. [DOI] [PubMed] [Google Scholar]
- 44.Arevalo-Martin A, Garcia-Ovejero D, Gomez O, et al. CB2 cannabinoid receptors as an emerging target for demyelinating diseases: from neuroimmune interactions to cell replacement strategies. Br J Pharmacol. 2008;153:216–225. doi: 10.1038/sj.bjp.0707466. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Centonze D, Bari M, Rossi S, et al. The endocannabinoid system is dysregulated in multiple sclerosis and in experimental autoimmune encephalomyelitis. Brain. 2007;130:2543–2553. doi: 10.1093/brain/awm160. [DOI] [PubMed] [Google Scholar]
- 46.Ghosh S, Preet A, Groopman JE, Ganju RK. Cannabinoid receptor CB2 modulates the CXCL12/CXCR4-mediated chemotaxis of T lymphocytes. Mol Immunol. 2006;43:2169–2179. doi: 10.1016/j.molimm.2006.01.005. [DOI] [PubMed] [Google Scholar]
- 47.Iglesias A, Bauer J, Litzenburger T, Schubart A, Linington C. T- and B-cell responses to myelin oligodendrocyte glycoprotein in experimental autoimmune encephalomyelitis and multiple sclerosis. Glia. 2001;36:220–234. doi: 10.1002/glia.1111. [DOI] [PubMed] [Google Scholar]
- 48.Aranami T, Yamamura T. Th17 Cells and autoimmune encephalomyelitis (EAE/MS) Allergol Int. 2008;57:115–120. doi: 10.2332/allergolint.R-07-159. [DOI] [PubMed] [Google Scholar]
- 49.Frisullo G, Nociti V, Iorio R, et al. IL17 and IFNγ production by peripheral blood mononuclear cells from clinically isolated syndrome to secondary progressive multiple sclerosis. Cytokine. 2008;44:22–25. doi: 10.1016/j.cyto.2008.08.007. [DOI] [PubMed] [Google Scholar]
- 50.Sanchez AJ, Gonzalez-Perez P, Galve-Roperh I, Garcia-Merino A. R-(+)-[2,3-dihydro-5-methyl-3-(4-morpholinylmethyl)-pyrrolo-[1,2,3-de]-1,4 -benzoxazin-6-yl]-1-naphtalenylmethanone (WIN-2) ameliorates experimental autoimmune encephalomyelitis and induces encephalitogenic T cell apoptosis: partial involvement of the CB(2) receptor. Biochem Pharmacol. 2006;72:1697–1706. doi: 10.1016/j.bcp.2006.08.018. [DOI] [PubMed] [Google Scholar]
- 51.Arevalo-Martin A, Vela JM, Molina-Holgado E, Borrell J, Guaza C. Therapeutic action of cannabinoids in a murine model of multiple sclerosis. J Neurosci. 2003;23:2511–2516. doi: 10.1523/JNEUROSCI.23-07-02511.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Mestre L, Docagne F, Correa F, et al. A cannabinoid agonist interferes with the progression of a chronic model of multiple sclerosis by downregulating adhesion molecules. Mol Cell Neurosci. 2009;40:258–266. doi: 10.1016/j.mcn.2008.10.015. [DOI] [PubMed] [Google Scholar]
- 53.Ehrhart J, Obregon D, Mori T, et al. Stimulation of cannabinoid receptor 2 (CB2) suppresses microglial activation. J Neuroinflammation. 2005;2:29. doi: 10.1186/1742-2094-2-29. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Correa F, Docagne F, Mestre L, et al. A role for CB2 receptors in anandamide signalling pathways involved in the regulation of IL-12 and IL-23 in microglial cells. Biochem Pharmacol. 2009;77:86–100. doi: 10.1016/j.bcp.2008.09.014. [DOI] [PubMed] [Google Scholar]
- 55.Palazuelos J, Davoust N, Julien B, et al. The CB(2) cannabinoid receptor controls myeloid progenitor trafficking: involvement in the pathogenesis of an animal model of multiple sclerosis. J Biol Chem. 2008;283:13320–13329. doi: 10.1074/jbc.M707960200. [DOI] [PubMed] [Google Scholar]
- 56.Sheng WS, Hu S, Min X, Cabral GA, Lokensgard JR, Peterson PK. Synthetic cannabinoid WIN55,212–212 inhibits generation of inflammatory mediators by IL-1β-stimulated human astrocytes. Glia. 2005;49:211–219. doi: 10.1002/glia.20108. [DOI] [PubMed] [Google Scholar]
- 57.Molina-Holgado F, Molina-Holgado E, Guaza C. The endogenous cannabinoid anandamide potentiates interleukin-6 production by astrocytes infected with Theiler’s murine encephalomyelitis virus by a receptor-mediated pathway. FEBS Lett. 1998;433:139–142. doi: 10.1016/s0014-5793(98)00851-5. [DOI] [PubMed] [Google Scholar]
- 58.Wood JD, Alpers DH, Andrews PL. Fundamentals of neurogastroenterology. Gut. 1999;45(Suppl. 2):II6–II16. doi: 10.1136/gut.45.2008.ii6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Massa F, Marsicano G, Hermann H, et al. The endogenous cannabinoid system protects against colonic inflammation. J Clin Invest. 2004;113:1202–1209. doi: 10.1172/JCI19465. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.Pertwee RG. Cannabinoids and the gastrointestinal tract. Gut. 2001;48:859–867. doi: 10.1136/gut.48.6.859. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61.Okayasu I, Hatakeyama S, Yamada M, Ohkusa T, Inagaki Y, Nakaya R. A novel method in the induction of reliable experimental acute and chronic ulcerative colitis in mice. Gastroenterology. 1990;98:694–702. doi: 10.1016/0016-5085(90)90290-h. [DOI] [PubMed] [Google Scholar]
- 62.Yamada Y, Marshall S, Specian RD, Grisham MB. A comparative analysis of two models of colitis in rats. Gastroenterology. 1992;102:1524–1534. doi: 10.1016/0016-5085(92)91710-l. [DOI] [PubMed] [Google Scholar]
- 63.Neurath MF, Fuss I, Kelsall BL, Stuber E, Strober W. Antibodies to interleukin 12 abrogate established experimental colitis in mice. J Exp Med. 1995;182:1281–1290. doi: 10.1084/jem.182.5.1281. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64.Kimball ES, Schneider CR, Wallace NH, Hornby PJ. Agonists of cannabinoid receptor 1 and 2 inhibit experimental colitis induced by oil of mustard and by dextran sulfate sodium. Am J Physiol Gastrointest Liver Physiol. 2006;291:G364–G371. doi: 10.1152/ajpgi.00407.2005. [DOI] [PubMed] [Google Scholar]
- 65.Izzo AA, Fezza F, Capasso R, et al. Cannabinoid CB1-receptor mediated regulation of gastrointestinal motility in mice in a model of intestinal inflammation. Br J Pharmacol. 2001;134:563–570. doi: 10.1038/sj.bjp.0704293. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 66.Cravatt BF, Demarest K, Patricelli MP, et al. Supersensitivity to anandamide and enhanced endogenous cannabinoid signaling in mice lacking fatty acid amide hydrolase. Proc Natl Acad Sci USA. 2001;98:9371–9376. doi: 10.1073/pnas.161191698. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 67.Di Marzo V, De Petrocellis L, Fezza F, Ligresti A, Bisogno T. Anandamide receptors. Prostaglandins Leukot Essent Fatty Acids. 2002;66:377–391. doi: 10.1054/plef.2001.0349. [DOI] [PubMed] [Google Scholar]
- 68.Biecker E, Sagesser H, Reichen J. Vasodilator mRNA levels are increased in the livers of portal hypertensive NO-synthase 3-deficient mice. Eur J Clin Invest. 2004;34:283–289. doi: 10.1111/j.1365-2362.2004.01331.x. [DOI] [PubMed] [Google Scholar]
- 69.Julien B, Grenard P, Teixeira-Clerc F, et al. Antifibrogenic role of the cannabinoid receptor CB2 in the liver. Gastroenterology. 2005;128:742–755. doi: 10.1053/j.gastro.2004.12.050. [DOI] [PubMed] [Google Scholar]
- 70.Osei-Hyiaman D, DePetrillo M, Pacher P, et al. Endocannabinoid activation at hepatic CB1 receptors stimulates fatty acid synthesis and contributes to diet-induced obesity. J Clin Invest. 2005;115:1298–1305. doi: 10.1172/JCI23057. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 71.Schwabe RF, Siegmund SV. Potential role of CB2 receptors in cannabis smokers with chronic hepatitis C. Hepatology. 2005;42:975–977. doi: 10.1002/hep.20895. [DOI] [PubMed] [Google Scholar]
- 72.Teixeira-Clerc F, Julien B, Grenard P, et al. CB1 cannabinoid receptor antagonism: a new strategy for the treatment of liver fibrosis. Nat Med. 2006;12:671–676. doi: 10.1038/nm1421. [DOI] [PubMed] [Google Scholar]
- 73.Batkai S, Jarai Z, Wagner JA, et al. Endocannabinoids acting at vascular CB1 receptors mediate the vasodilated state in advanced liver cirrhosis. Nat Med. 2001;7:827–832. doi: 10.1038/89953. [DOI] [PubMed] [Google Scholar]
- 74.Batkai S, Osei-Hyiaman D, Pan H, et al. Cannabinoid-2 receptor mediates protection against hepatic ischemia/reperfusion injury. FASEB J. 2007;21:1788–1800. doi: 10.1096/fj.06-7451com. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 75.Gary-Bobo M, Elachouri G, Gallas JF, et al. Rimonabant reduces obesity-associated hepatic steatosis and features of metabolic syndrome in obese Zucker fa/fa rats. Hepatology. 2007;46:122–129. doi: 10.1002/hep.21641. [DOI] [PubMed] [Google Scholar]
- 76.Siegmund SV, Qian T, de Minicis S, et al. The endocannabinoid 2-arachidonoyl glycerol induces death of hepatic stellate cells via mitochondrial reactive oxygen species. FASEB J. 2007;21:2798–2806. doi: 10.1096/fj.06-7717com. [DOI] [PubMed] [Google Scholar]
- 77.Hezode C, Roudot-Thoraval F, Nguyen S, et al. Daily cannabis smoking as a risk factor for progression of fibrosis in chronic hepatitis C. Hepatology. 2005;42:63–71. doi: 10.1002/hep.20733. [DOI] [PubMed] [Google Scholar]
- 78.Avraham Y, Israeli E, Gabbay E, et al. Endocannabinoids affect neurological and cognitive function in thioacetamide-induced hepatic encephalopathy in mice. Neurobiol Dis. 2006;21:237–245. doi: 10.1016/j.nbd.2005.07.008. [DOI] [PubMed] [Google Scholar]
- 79.Hegde VL, Hegde S, Cravatt BF, Hofseth LJ, Nagarkatti M, Nagarkatti PS. Attenuation of experimental autoimmune hepatitis by exogenous and endogenous cannabinoids: involvement of regulatory T cells. Mol Pharmacol. 2008;74:20–33. doi: 10.1124/mol.108.047035. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 80.Hurlimann D, Enseleit F, Ruschitzka F. Rheumatoid arthritis, inflammation, and atherosclerosis. Herz. 2004;29:760–768. doi: 10.1007/s00059-004-2636-7. [DOI] [PubMed] [Google Scholar]
- 81.Pratt AG, Isaacs JD, Mattey DL. Current concepts in the pathogenesis of early rheumatoid arthritis. Best Pract Res Clin Rheumatol. 2009;23:37–48. doi: 10.1016/j.berh.2008.08.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 82.Zurier RB, Rossetti RG, Burstein SH, Bidinger B. Suppression of human monocyte interleukin-1β production by ajulemic acid, a nonpsychoactive cannabinoid. Biochem Pharmacol. 2003;65:649–655. doi: 10.1016/s0006-2952(02)01604-0. [DOI] [PubMed] [Google Scholar]
- 83.George KL, Saltman LH, Stein GS, Lian JB, Zurier RB. Ajulemic acid, a nonpsychoactive cannabinoid acid, suppresses osteoclastogenesis in mononuclear precursor cells and induces apoptosis in mature osteoclast-like cells. J Cell Physiol. 2008;214:714–720. doi: 10.1002/jcp.21263. [DOI] [PubMed] [Google Scholar]
- 84.Balkwill F, Mantovani A. Inflammation and cancer: back to Virchow? Lancet. 2001;357:539–545. doi: 10.1016/S0140-6736(00)04046-0. [DOI] [PubMed] [Google Scholar]
- 85.Malfait AM, Gallily R, Sumariwalla PF, et al. The nonpsychoactive cannabis constituent cannabidiol is an oral anti-arthritic therapeutic in murine collagen-induced arthritis. Proc Natl Acad Sci USA. 2000;97:9561–9566. doi: 10.1073/pnas.160105897. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 86.Sumariwalla PF, Gallily R, Tchilibon S, Fride E, Mechoulam R, Feldmann M. A novel synthetic, nonpsychoactive cannabinoid acid (HU-320) with antiinflammatory properties in murine collagen-induced arthritis. Arthritis Rheum. 2004;50:985–998. doi: 10.1002/art.20050. [DOI] [PubMed] [Google Scholar]
- 87.Mamytbekova A, Rezabek K, Kacerovska H, Grimova J, Svobodova J. Antimetastatic effect of flurbiprofen and other platelet aggregation inhibitors. Neoplasma. 1986;33:417–421. [PubMed] [Google Scholar]
- 88.Sarfaraz S, Adhami VM, Syed DN, Afaq F, Mukhtar H. Cannabinoids for cancer treatment: progress and promise. Cancer Res. 2008;68:339–342. doi: 10.1158/0008-5472.CAN-07-2785. [DOI] [PubMed] [Google Scholar]
- 89.Carracedo A, Gironella M, Lorente M, et al. Cannabinoids induce apoptosis of pancreatic tumor cells via endoplasmic reticulum stress-related genes. Cancer Res. 2006;66:6748–6755. doi: 10.1158/0008-5472.CAN-06-0169. [DOI] [PubMed] [Google Scholar]
- 90.Casanova ML, Blazquez C, Martinez-Palacio J, et al. Inhibition of skin tumor growth and angiogenesis in vivo by activation of cannabinoid receptors. J Clin Invest. 2003;111:43–50. doi: 10.1172/JCI16116. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 91.Galve-Roperh I, Sanchez C, Cortes ML, et al. Anti-tumoral action of cannabinoids: involvement of sustained ceramide accumulation and extracellular signal-regulated kinase activation. Nat Med. 2000;6:313–319. doi: 10.1038/73171. [DOI] [PubMed] [Google Scholar]
- 92.Sanchez C, de Ceballos ML, Gomez del Pulgar T, et al. Inhibition of glioma growth in vivo by selective activation of the CB(2) cannabinoid receptor. Cancer Res. 2001;61:5784–5789. [PubMed] [Google Scholar]
- 93.Sarfaraz S, Afaq F, Adhami VM, Mukhtar H. Cannabinoid receptor as a novel target for the treatment of prostate cancer. Cancer Res. 2005;65:1635–1641. doi: 10.1158/0008-5472.CAN-04-3410. [DOI] [PubMed] [Google Scholar]
- 94.Velasco G, Galve-Roperh I, Sanchez C, Blazquez C, Guzman M. Hypothesis: cannabinoid therapy for the treatment of gliomas? Neuropharmacology. 2004;47:315–323. doi: 10.1016/j.neuropharm.2004.04.016. [DOI] [PubMed] [Google Scholar]
- 95.Lombard C, Nagarkatti M, Nagarkatti PS. Targeting cannabinoid receptors to treat leukemia: role of cross-talk between extrinsic and intrinsic pathways in Δ9-tetrahydrocannabinol (THC)-induced apoptosis of Jurkat cells. Leuk Res. 2005;29:915–922. doi: 10.1016/j.leukres.2005.01.014. [DOI] [PubMed] [Google Scholar]
- 96.Jia W, Hegde VL, Singh NP, et al. Δ9-tetrahydrocannabinol-induced apoptosis in Jurkat leukemia T cells is regulated by translocation of Bad to mitochondria. Mol Cancer Res. 2006;4:549–562. doi: 10.1158/1541-7786.MCR-05-0193. [DOI] [PubMed] [Google Scholar]
- 97.McKallip RJ, Lombard C, Fisher M, et al. Targeting CB2 cannabinoid receptors as a novel therapy to treat malignant lymphoblastic disease. Blood. 2002;100:627–634. doi: 10.1182/blood-2002-01-0098. [DOI] [PubMed] [Google Scholar]
- 98.Ligresti A, Bisogno T, Matias I, et al. Possible endocannabinoid control of colorectal cancer growth. Gastroenterology. 2003;125:677–687. doi: 10.1016/s0016-5085(03)00881-3. [DOI] [PubMed] [Google Scholar]
- 99.Cianchi F, Papucci L, Schiavone N, et al. Cannabinoid receptor activation induces apoptosis through tumor necrosis factor α-mediated ceramide de novo synthesis in colon cancer cells. Clin Cancer Res. 2008;14:7691–7700. doi: 10.1158/1078-0432.CCR-08-0799. [DOI] [PubMed] [Google Scholar]
- 100.Bifulco M, Laezza C, Valenti M, et al. A new strategy to block tumor growth by inhibiting endocannabinoid inactivation. FASEB J. 2004;18:1606–1608. doi: 10.1096/fj.04-1754fje. [DOI] [PubMed] [Google Scholar]
- 101.De Lago E, Gustafsson SB, Fernandez-Ruiz J, et al. Acyl-based anandamide uptake inhibitors cause rapid toxicity to C6 glioma cells at pharmacologically relevant concentrations. J Neurochem. 2006;99:677–688. doi: 10.1111/j.1471-4159.2006.04104.x. [DOI] [PubMed] [Google Scholar]
- 102.Endsley MP, Aggarwal N, Isbell MA, et al. Diverse roles of 2-arachidonoylglycerol in invasion of prostate carcinoma cells: location, hydrolysis and 12-lipoxygenase metabolism. Int J Cancer. 2007;121:984–991. doi: 10.1002/ijc.22761. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 103.Nithipatikom K, Endsley MP, Isbell MA, Wheelock CE, Hammock BD, Campbell WB. A new class of inhibitors of 2-arachidonoylglycerol hydrolysis and invasion of prostate cancer cells. Biochem Biophys Res Commun. 2005;332:1028–1033. doi: 10.1016/j.bbrc.2005.05.049. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 104.Contassot E, Tenan M, Schnuriger V, Pelte MF, Dietrich PY. Arachidonyl ethanolamide induces apoptosis of uterine cervix cancer cells via aberrantly expressed vanilloid receptor-1. Gynecol Oncol. 2004;93:182–188. doi: 10.1016/j.ygyno.2003.12.040. [DOI] [PubMed] [Google Scholar]
- 105.Contassot E, Wilmotte R, Tenan M, et al. Arachidonylethanolamide induces apoptosis of human glioma cells through vanilloid receptor-1. J Neuropathol Exp Neurol. 2004;63:956–963. doi: 10.1093/jnen/63.9.956. [DOI] [PubMed] [Google Scholar]
- 106.DeMorrow S, Glaser S, Francis H, et al. Opposing actions of endocannabinoids on cholangiocarcinoma growth: recruitment of Fas and Fas ligand to lipid rafts. J Biol Chem. 2007;282:13098–13113. doi: 10.1074/jbc.M608238200. [DOI] [PubMed] [Google Scholar]
- 107.Bifulco M, Laezza C, Portella G, et al. Control by the endogenous cannabinoid system of ras oncogene-dependent tumor growth. FASEB J. 2001;15:2745–2747. doi: 10.1096/fj.01-0320fje. [DOI] [PubMed] [Google Scholar]
- 108.Caffarel MM, Sarrio D, Palacios J, Guzman M, Sanchez C. Δ9-tetrahydrocannabinol inhibits cell cycle progression in human breast cancer cells through Cdc2 regulation. Cancer Res. 2006;66:6615–6621. doi: 10.1158/0008-5472.CAN-05-4566. [DOI] [PubMed] [Google Scholar]
- 109.Ek S, Hogerkorp CM, Dictor M, Ehinger M, Borrebaeck CA. Mantle cell lymphomas express a distinct genetic signature affecting lymphocyte trafficking and growth regulation as compared with subpopulations of normal human B cells. Cancer Res. 2002;62:4398–4405. [PubMed] [Google Scholar]
- 110.Islam TC, Asplund AC, Lindvall JM, et al. High level of cannabinoid receptor 1, absence of regulator of G protein signalling 13 and differential expression of cyclin D1 in mantle cell lymphoma. Leukemia. 2003;17:1880–1890. doi: 10.1038/sj.leu.2403057. [DOI] [PubMed] [Google Scholar]
- 111.Wills-Karp M. Immunologic basis of antigen-induced airway hyperresponsiveness. Annu Rev Immunol. 1999;17:255–281. doi: 10.1146/annurev.immunol.17.1.255. [DOI] [PubMed] [Google Scholar]
- 112.Anderson GP, Coyle AJ. TH2 and ‘TH2-like’ cells in allergy and asthma: pharmacological perspectives. Trends Pharmacol Sci. 1994;15:324–332. doi: 10.1016/0165-6147(94)90027-2. [DOI] [PubMed] [Google Scholar]
- 113.Jan TR, Farraj AK, Harkema JR, Kaminski NE. Attenuation of the ovalbumin-induced allergic airway response by cannabinoid treatment in A/J mice. Toxicol Appl Pharmacol. 2003;188:24–35. doi: 10.1016/s0041-008x(03)00010-3. [DOI] [PubMed] [Google Scholar]
- 114.Tashkin DP, Reiss S, Shapiro BJ, Calvarese B, Olsen JL, Lodge JW. Bronchial effects of aerosolized Δ9-tetrahydrocannabinol in healthy and asthmatic subjects. Am Rev Respir Dis. 1977;115:57–65. doi: 10.1164/arrd.1977.115.1.57. [DOI] [PubMed] [Google Scholar]
- 115.Abboud RT, Sanders HD. Effect of oral administration of Δ-tetrahydrocannabinol on airway mechanics in normal and asthmatic subjects. Chest. 1976;70:480–485. doi: 10.1378/chest.70.4.480. [DOI] [PubMed] [Google Scholar]
- 116.Hartley JP, Nogrady SG, Seaton A. Bronchodilator effect of Δ1-tetrahydrocannabinol. Br J Clin Pharmacol. 1978;5:523–525. doi: 10.1111/j.1365-2125.1978.tb01667.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 117.Calignano A, Katona I, Desarnaud F, et al. Bidirectional control of airway responsiveness by endogenous cannabinoids. Nature. 2000;408:96–101. doi: 10.1038/35040576. [DOI] [PubMed] [Google Scholar]
- 118.Rajesh M, Mukhopadhyay P, Batkai S, et al. Cannabidiol attenuates high glucose-induced endothelial cell inflammatory response and barrier disruption. Am J Physiol Heart Circ Physiol. 2007;293:H610–H619. doi: 10.1152/ajpheart.00236.2007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 119.Rajesh M, Mukhopadhyay P, Hasko G, Huffman JW, Mackie K, Pacher P. CB2 cannabinoid receptor agonists attenuate TNF-α-induced human vascular smooth muscle cell proliferation and migration. Br J Pharmacol. 2008;153:347–357. doi: 10.1038/sj.bjp.0707569. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 120.de Kozak Y, Sakai J, Thillaye B, Faure JP. S antigen-induced experimental autoimmune uveo-retinitis in rats. Curr Eye Res. 1981;1:327–337. doi: 10.3109/02713688108998359. [DOI] [PubMed] [Google Scholar]
- 121.Xu H, Cheng CL, Chen M, et al. Anti-inflammatory property of the cannabinoid receptor-2-selective agonist JWH-133 in a rodent model of autoimmune uveoretinitis. J Leukoc Biol. 2007;82:532–541. doi: 10.1189/jlb.0307159. [DOI] [PubMed] [Google Scholar]
- 122.Correa F, Mestre L, Docagne F, Guaza C. Activation of cannabinoid CB2 receptor negatively regulates IL-12p40 production in murine macrophages: role of IL-10 and ERK1/2 kinase signaling. Br J Pharmacol. 2005;145:441–448. doi: 10.1038/sj.bjp.0706215. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 123.Broderick CA, Smith AJ, Balaggan KS, et al. Local administration of an adeno-associated viral vector expressing IL-10 reduces monocyte infiltration and subsequent photoreceptor damage during experimental autoimmune uveitis. Mol Ther. 2005;12:369–373. doi: 10.1016/j.ymthe.2005.03.018. [DOI] [PubMed] [Google Scholar]
- 124.De Kozak Y, Thillaye-Goldenberg B, Naud MC, Da Costa AV, Auriault C, Verwaerde C. Inhibition of experimental autoimmune uveoretinitis by systemic and subconjunctival adenovirus-mediated transfer of the viral IL-10 gene. Clin Exp Immunol. 2002;130:212–223. doi: 10.1046/j.1365-2249.2002.01969.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 125.Mandrup-Poulsen T. β cell death and protection. Ann NY Acad Sci. 2003;1005:32–42. doi: 10.1196/annals.1288.005. [DOI] [PubMed] [Google Scholar]
- 126.Li X, Kaminski NE, Fischer LJ. Examination of the immunosuppressive effect of Δ9-tetrahydrocannabinol in streptozotocin-induced autoimmune diabetes. Int Immunopharmacol. 2001;1:699–712. doi: 10.1016/s1567-5769(01)00003-0. [DOI] [PubMed] [Google Scholar]
- 127.Weiss L, Zeira M, Reich S, et al. Cannabidiol lowers incidence of diabetes in non-obese diabetic mice. Autoimmunity. 2006;39:143–151. doi: 10.1080/08916930500356674. [DOI] [PubMed] [Google Scholar]
- 128.Weiss L, Zeira M, Reich S, et al. Cannabidiol arrests onset of autoimmune diabetes in NOD mice. Neuropharmacology. 2008;54:244–249. doi: 10.1016/j.neuropharm.2007.06.029. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 129.El-Remessy AB, Al-Shabrawey M, Khalifa Y, Tsai NT, Caldwell RB, Liou GI. Neuroprotective and blood–retinal barrier-preserving effects of cannabidiol in experimental diabetes. Am J Pathol. 2006;168:235–244. doi: 10.2353/ajpath.2006.050500. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 130.Dogrul A, Gul H, Yildiz O, Bilgin F, Guzeldemir ME. Cannabinoids blocks tactile allodynia in diabetic mice without attenuation of its antinociceptive effect. Neurosci Lett. 2004;368:82–86. doi: 10.1016/j.neulet.2004.06.060. [DOI] [PubMed] [Google Scholar]