

Abstract
Trace amines such as p-tyramine and β-phenylethylamine are found endogenously as well as in the diet. Concomitant ingestion of these foodstuffs with monoamine oxidase inhibitors may result in the hypertensive crisis known as the “beer, wine, and cheese effect” attributed to their sympathomimetic action. Trace amines have been shown to act on one of a novel group of mammalian seven transmembrane spanning G protein-coupled receptors belonging to the rhodopsin superfamily, cloned in 2001. This receptor encoded by the human TAAR1 gene is also present in rat and mouse genomes (Taar1) and has been shown to be activated by endogenous trace amine ligands, including p-tyramine and β-phenylethylamine. A number of drugs, most notably amphetamine and its derivatives, act as agonists at this receptor. This review proposes an official nomenclature designating TAAR1 as the trace amine 1 receptor following the convention of naming receptors after the endogenous agonist, abbreviated to TA1 where necessary. It goes on to discuss briefly the significance of the receptor, agents acting upon it, its distribution, and currently hypothesized physiological and pathophysiological roles. In humans, a further five genes are thought to encode functional receptors (TAAR2, TAAR5, TAAR6, TAAR8, and TAAR9). TAAR3 seems to be a pseudogene in some individuals but not others. TAAR4 is a pseudogene in humans, but occurs with TAAR3 as a functional gene in rodents. Nine further genes are present in rats and mice. The endogenous ligands are not firmly established but some may respond to odorants consistent with their expression in olfactory epithelium.
I. Introduction
Trace amines, such as p-tyramine and β-phenylethylamine (β-PEA1) were discovered more than a century ago [e.g., β-PEA by Nencki in 1876 (reviewed in Grandy, 2007)] and are well known sympathomimetics (Dale and Dixon, 1909; Barger and Dale, 1910) described as “false transmitters.” In mammals, trace amines are synthesized from aromatic amino acids at rates comparable with classic monoamines (for example, tyramine from tyrosine, catalyzed by aromatic l-amino acid decarboxylase) (David et al., 1974; Boulton, 1976; Bowsher and Henry, 1983; Brier et al., 1991). However, they are detectable only at trace levels because they are substrates for monoamine oxidase and have a half-life of approximately 30 s (Durden and Philips, 1980; Paterson et al., 1990). Trace amines are also present in significant amounts in fermented foods (Chaytor et al., 1975; Hannah et al., 1988), and concomitant ingestion of these with monoamine oxidase inhibition may result in the hypertensive crisis known as the “beer, wine, and cheese effect” (Blackwell, 1963; Cooper, 1989; Caston et al., 2002).
In invertebrates, tyramine and octopamine are well characterized neurotransmitters, modulating metabolism and muscle tone (Axelrod and Saavedra, 1977; Roeder, 2005) via their own G protein coupled receptors, which have been cloned (Arakawa et al., 1990; Saudou et al., 1990).
II. Trace Amine Receptors
A. Designation of the Trace Amine 1 Receptor
In 2001, a novel mammalian G-protein-coupled receptor (GPCR) was cloned in a search for further subtypes of the 5HT receptor. It was shown to have high (nanomolar) affinity for trace amines and was therefore named the trace amine 1 (TA1) receptor (Borowsky et al., 2001). Subsequently, a family of genes encoding trace amine receptors was cloned (Borowsky et al., 2001; Lindemann et al., 2005) that showed closest homology to the aminergic receptors. The gene name was at first abbreviated to TA then TAR after the initial pairing. However, it was subsequently shown that not all family members may have high affinity for trace amines, which has led to the adoption of the nomenclature of “trace amine-associated receptors” (TAARs) for the genes encoding the receptors (Lindemann et al., 2005). The Human Genome Organization (HUGO) Gene Nomenclature Committee has approved the gene symbol for the trace amine receptor 1 as TAAR1 (http://www.genenames.org/cgi-bin/hgnc_search.pl) because it also avoids confusion with existing genes and ensures that this family has a unique name that can be searched on databases. This nomenclature links together the other members of the “Associated Receptors” that are homologous but for which the precise pharmacology remains to be determined.
To facilitate comparison between members of this family, current and previous nomenclatures for both the receptor proteins and the family of genes encoding them are given in Table 1. To date, only one receptor, TA1, has been shown to respond to a cognate ligand, and several different groups replicated the results (Borowsky et al., 2001; Bunzow et al., 2001, Liberles and Buck, 2006), leading to the International Union of Pharmacology (IUPHAR)-recommended nomenclature for the receptor protein encoded by the gene TAAR1 as trace amine receptor 1, abbreviated to TA1, first proposed by Borowsky et al. (2001). This follows the agreed convention on naming receptor proteins after the cognate endogenous ligand. According to IUPHAR convention no R is added to the abbreviated name for receptor proteins.
TABLE 1.
Summary of human, mouse, and rat genes encoding trace amine and trace amine-associated receptors
| Receptor |
Human Gene |
Mouse Gene |
Rat Gene |
||||
|---|---|---|---|---|---|---|---|
| Old Name | New Name | Swiss Prot RefSeq | Name | Swiss Prot RefSeq | Name | Swiss Prot RefSeq | |
| IUPHAR Receptor Nomenclature | |||||||
| TA1 | TAR1 | TAAR1 | Q96RJ0 | Taar1 | Q923Y8 | Taar1 | Q923Y9 |
| NP_612200 | NP_444435 | NP_599155 | |||||
| Unofficial Receptor Terminologya | |||||||
| TA2 | Taar4 | Q5QD15 | Taar4 | Q923Y7 | |||
| NP_001008499 | NP_783173 | ||||||
| TA3 | TAR3 | TAAR9 | Q96R19b | Taar9 | Q5QD04 | Taar9 | Q923Y6 |
| NP_778227 | NP_001010831 | NP_783192 | |||||
| TA4 | TAR4 | TAAR6 | Q96R18c | Taar6 | Q5QD13 | Taar6 | Q923Y5 |
| NP_778237 | NP_001010828 | NP_783174 | |||||
| TA5 | GPR102 | TAAR8 | Q969N4 | ||||
| NP_444508 | |||||||
| TA6 | Taar7h | Q923Y4 | |||||
| NP_783177 | |||||||
| TA7 | Taar8b | Q5QD06 | Taar8b | Q923Y3 | |||
| NP_001010837 | NP_783191 | ||||||
| TA8 | Taar7a | Q5QD12 | Taar7a | Q923Y2 | |||
| NP_001010829 | NP_783175 | ||||||
| TA9 | Taar7g | Q923Y1 | |||||
| NP_783178 | |||||||
| TA10 | Taar8c | Q5QD05 | Taar8c | Q923Y0 | |||
| NP_001010840 | NP_783190 | ||||||
| TA11 | Taar8a | Q5QD07 | Taar8a | Q923X9 | |||
| NP_001010830 | NP_783189 | ||||||
| TA12 | Taar7b | Q5QD11 | Taar7b | Q923X8 | |||
| NP_001010827 | NP_783176 | ||||||
| TA13 | Taar7f | Q5QD08 | Taar7f | Pseudogene | |||
| NP_001010839 | |||||||
| TA14 | Taar7e | Q5QD09 | Taar7e | Q923X6 | |||
| NP_001010835 | NP_783180 | ||||||
| TA15 | Taar7d | Q5QD10 | Taar7d | Q923X5 | |||
| NP_001010838 | NP_783181 | ||||||
| GPR57 | Taar3 | Q5QD16 | |||||
| NP_001008429 | |||||||
| GPR58 | GPR58 | TAAR2 | Q9P1P5 | Taar2 | Q5QD17 | Taar2 | Q5QD25 |
| NP_001028252 | NP_001007267 | NP_001008512 | |||||
| or NP_055441 | |||||||
| PNR | PNR | TAAR5 | O14804 | Taar5 | Q5QD14 | Taar5 | Q5QD23 |
| NP_003958 | NP_001009574 | NP_001009650 | |||||
Nomenclature as designated by Borowsky et al. (2001). See Lindemann and Hoener (2005) for further information on TAARs and Foord et al. (2005) and Schöneberg et al. (2007) for further information on pseudogenes
Stop codon present in 10% of humans
Polymorphisms in the human gene have been reported to be associated with schizophrenia and bipolar disorder (Duan et al., 2004; Pae et al., 2008a,b)
It has been shown that the rat TA1 receptor, expressed in HEK293 cells, was also activated by thyronamines (decarboxylated and deiodinated metabolites of the thyroid hormones) (Scanlan et al., 2004; Hart et al., 2006) with a potency similar to that of tyramine (Bunzow et al., 2001). Cardiac effects of iodothyronamines have been reported in rat, but the rank order of potencies and affinities in ligand binding assays were not consistent with activation of TA1 and led the authors to suggest that iodothyronamines might be acting at different trace amine-associated receptors (Frascarelli et al., 2008).
The main focus of this review is the recommendation of nomenclature for TA1 receptors. For excellent, comprehensive reviews on trace amines, TA1, and the related family of receptors, see Grandy (2007), Zucchi et al. (2006), Lewin (2006), Lindemann and Hoener (2005), and Davenport (2003).
B. TAAR1 Gene
TAAR1 is localized in humans, along with genes for other trace amine-associated receptors, to a 109-kilobase region of chromosome 6q23.2 (Borowsky et al., 2001; Bunzow et al., 2001; Lindemann et al., 2005). Taar genes have also been found in the rat (Table 1; Bunzow et al., 2001) and mouse (Table 1; Borowsky et al., 2001). Designation of these genes follows the International Committee on Standardized Genetic Nomenclature for Mice and Rat Genome and Nomenclature Committee recommendation to use lower case italics (e.g., Taar1). This distinguishes the human gene, written in capitals, from those of rodents. The rat and mouse genes are localized to chromosomes 1p12 (Bunzow et al., 2001) and 10A3 (Borowsky et al., 2001; Reese et al., 2007), respectively.
C. Other Family Members
The unofficial receptor names for the remaining receptors are included, for comparison, in Table 1 and await clear identification of their endogenous ligands, together with the official gene names. In humans, a further five genes are predicted to exist encoding trace amine-associated receptors [TAAR2, TAAR5, TAAR6, TAAR8, and TAAR9 (Table 1)], and these are thought to be functional genes. TAAR3 seems to be a pseudogene in some individuals but not others (Gloriam et al., 2005, Vanti et al., 2003). TAAR4 is a pseudogene in humans but occurs, with TAAR3, as a functional gene in rats and mice. Nine further genes are present in rats (Table 1; Bunzow et al., 2001) and mice (Table 1; Borowsky et al., 2001), but not in humans (Foord et al., 2005). The focus of this review is on human and rodent receptors; however, TAAR genes have also been reported in rhesus monkey (Miller et al., 2005) and avian (Mueller et al., 2008) genomes (Foord et al., 2005).
Odorants are detected in the nasal olfactory epithelium by the odorant receptor family, whose ∼1000 members allow the discrimination of many different odorants. In a key article, Liberles and Buck (2006) reported the presence of trace amine-associated receptors that, like odorant receptors, are expressed in unique subsets of neurons dispersed in the mouse olfactory epithelium. Interestingly Taar1 was not detected. They expressed some of the mouse Taar genes in HEK293 cells linked to a fluorescent reporter and found that several responded to various amine ligands: 1) In agreement with previous reports for human and rat TA1, mouse TA1 receptor recognized β-PEA with an EC50 = 0.1 μM (Liberles and Buck, 2006). 2) The mouse Taar3 gene product responded to several primary amines, including isoamylamine (EC50 = 10 μM) and cyclohexylamine but, interestingly, not to the corresponding alcohols, isoamylalcohol and cyclohexanol, indicating slight variations in ligand structure eliminated ligand activity in some cases. 3) Mouse Taar5 gene product (an ortholog also exists in humans; Table 1) responded to tertiary amines trimethylamine (EC50 = 0.3 μM) and N-methylpiperidine, but not to the related compounds methylamine, dimethylamine, and tetramethylammonium chloride. 4) Mouse Taar7f gene product responded to the tertiary amine, N-methylpiperidine (EC50 = 20 μM).
It is noteworthy that three of the ligands identified that activate mouse Taar gene products are natural components of mouse urine, a major source of social cues in rodents. Mouse Taar4 gene product recognizes β-PEA, a compound whose elevation in urine is correlated with increases in stress and stress responses in both rodents and humans. The gene products of mouse Taar3 and Taar5 detected compounds (isoamylamine and trimethylamine, respectively) that are enriched in male versus female mouse urine. Isoamylamine in male urine is reported to act as a pheromone, accelerating puberty onset in female mice (Liberles and Buck, 2006). The authors suggest the Taar family has a chemosensory function that is distinct from odorant receptors, with a role associated with the detection of social cues. In mice, a clear discrepancy between the expression pattern of mRNA encoding Taar1 and the other Taar family members has been confirmed by Regard et al. (2008). They have reported the anatomical distribution of mRNA for GPCRs in mouse tissues and shown highest expression of Taar1 in pancreatic islets of Langerhans and white adipose with no evidence for expression in the olfactory epithelium, in contrast to the other family members that are relatively highly expressed in this tissue (Regard et al., 2008).
Human homologs of Taar3, Taar4, and Taar7 are thought to be pseudogenes but Taar5 does have an apparently functional human ortholog and the results suggest that functional members of the family more generally respond to trace amines. Responses were not reported for the gene products of other human orthologs TAAR2, TAAR6, TAAR8, and TAAR9 (although it was not established that they were successfully expressed in HEK293 cells) and a role in humans for these TAARs is not yet clear.
D. Phylogeny
In addition to Taar genes being present in the rat (Table 1; Bunzow et al., 2001) and mouse (Table 1; Borowsky et al., 2001, Regard et al., 2008), they have also been reported in other species including rhesus monkey (Miller et al., 2005), avian (Mueller et al., 2008), fish, and amphibian genomes (Hashiguchi and Nishida, 2007) (see Foord et al., 2005).
The evolutionary pattern of the TAAR gene family is characterized by lineage-specific phylogenetic clustering (Gloriam et al., 2005; Lindemann et al., 2005; Hashiguchi and Nishida, 2007). These characteristics are very similar to those observed in the olfactory GPCRs and vomeronasal (V1R, V2R) GPCR gene families. Hashiguchi and Nishida (2007) carried out a careful phylogenetic analysis of the trace amine receptors in fish, amphibians, birds, and mammals and concluded (from the species they considered) that there are five types of trace amine receptor genes. The first type included expanded “trace amine-like” GPCR families within fish (fish can respond to catecholamines and their metabolites when these are introduced into their water). However, type I also included the murine Taar receptors demonstrated to have an olfactory role and the human trace amine receptors, TAAR5, TAAR6, and TAAR8. Subfamily II also contained Taar genes that, in the mouse, are expressed in the olfactory epithelium and their products function as receptors for volatile amines (Liberles and Buck, 2006; Regard et al., 2008). Subfamilies III and V had no mammalian members. Subfamily IV contained human and murine TAAR1 and Taar1 genes, respectively. To date, these analyses suggest that the TAAR1 gene alone encodes a trace amine receptor that may serve a nonsensory function. “Trace amine pharmacology” probably extends beyond the putative trace amine receptors. It will require further characterization using pharmacology and physiology to determine whether the trace amine receptors are “sensory” or not (Grandy, 2007).
III. Receptor Structure
The human TA1 receptor is a member of the rhodopsin-type superfamily (i.e., it is a class A GPCR), with 339 amino acids. It has a predicted seven transmembrane spanning domain structure with short N- and C-terminal domains of 23 to 49 and 27 to 33 amino acids, respectively (Lindemann et al., 2005). Rat and mouse TA1 both have 332 amino acids, with sequence identities of 78 and 75% in relation to humans, respectively (see Table 2 and Figure 1).
TABLE 2.
Classification of trace amine 1 receptor
| Receptor | Trace Amine 1, TA1 |
|---|---|
| Previous names | TAR1, TRAR1, BO111, TAAR1 (approved human gene symbol) |
| Structural information | 7TM |
| Human: 339 aa (SwissProt Q96RJ0), chr 6q23.2 (Entrez 134864) | |
| Rat: 332 aa (SwissProt Q923Y9), chr 1p12 (Entrez 113914) | |
| Mouse: 332 aa (SwissProt Q923Y8) chr 10A3 (Entrez 111174) | |
| Functional assays | COS-7/HEK293 cells transfected with TA1 and Gαs (Borowsky et al., 2001; Lindemann et al., 2005; Wolinsky et al., 2007); X. laevis oocytes cotransfected with TA1 and CFTR (Borowsky et al., 2001); CHO cells expressing TA1 and promiscuous Gα16 (Navarro et al., 2006) |
| Endogenous agonists | Tyramine (pEC50 = 6.4–6.7), β-PEA (pEC50 = 6.2–7.0), (Borowsky et al., 2001; Wainscott et al., 2007; Barak et al., 2008) |
| Antagonists | None currently commercially available |
| Radioligand assays | COS-7 cells transiently transfected with human TA1 and rat Gαs (Borowsky et al., 2001) |
| Radioligands | [3H]tyramine (pKD = 7.7) (Borowsky et al., 2001); [125I]-,[2H]-, [3H]3-iodothyronamine (Miyakawa and Scanlan, 2006) |
| Transduction | Couples to Gαs (Borowsky et al., 2001; Lindemann et al., 2005; Wolinsky et al., 2007) and Gα16 (Navarro et al., 2006; Lewin et al., 2008) in vitro |
| Receptor distribution | Studies in humans, RT-PCR showed mRNA encoding TA1 in stomach, amygdala, kidney, lung, small intestine, cerebellum, dorsal root ganglion, hippocampus, hypothalamus, liver, medulla, pancreas, pituitary, reticular formation, prostate, skeletal muscle, spleen (Borowsky et al., 2001), pancreatic islets (Regard et al., 2007), circulating leukocytes (D'Andrea et al., 2003), and intestinal mucosa/endothelium (Kidd et al., 2008); in rats, mRNA found in cardiac ventricles (Chiellini et al., 2007); in mice, in situ hybridization localized mRNA encoding TA1 to discrete CNS areas, including the mitral cell layer of the olfactory bulb, trigeminal nuclei, cerebellar Purkinje cells, spinal cord, amygdala, hippocampus, monoaminergic nuclei (Borowsky et al., 2001), and dopaminergic neurons of the substantia nigra (Xie et al., 2007) |
| Tissue function | Inhibits uptake and induces efflux of monoamines at striatal and thalamic synapses (Xie et al., 2008; Xie and Miller, 2008) |
7TM, seven transmembrane; aa, amino acid(s); chr, chromosome; CFTR, cystic fibrosis transmembrane conductance regulator; CHO, Chinese hamster ovary; RT-PCR, reverse transcriptase polymerase chain reaction; CNS, central nervous system
Fig. 1.

Alignment of amino acid sequences of the rat, mouse, and human trace amine 1 (TA1) receptor. Produced using ClustalW2 (http://www.uniprot.org/?tab=align).
IV. Distribution of Receptor and mRNA Encoding the Receptor
In humans, reverse transcriptase polymerase chain reaction has shown moderate levels (∼100 copies/ng of cDNA) of mRNA encoding TA1 in the stomach; low levels (15–100 copies/ng of cDNA) in the amygdala, kidney, lung, and small intestine; and trace amounts (<15 copies/ng of cDNA) in the cerebellum, hippocampus, hypothalamus, liver, medulla oblongata, pituitary gland, pontine reticular formation, prostate gland, skeletal muscle, and spleen (Borowsky et al., 2001). Further amounts have been detected in pancreatic islets (Regard et al., 2007), circulating leukocytes of healthy subjects (D'Andrea et al., 2003; Nelson et al., 2007), and in normal small intestinal mucosal and endothelial cells (Kidd et al., 2008).
In mouse brain, mRNA encoding the receptor was detected by in situ hybridization in cerebellar Purkinje cells, trigeminal nuclei, olfactory bulb, hypothalamic nuclei, monoaminergic nuclei (such as the dorsal raphé and ventral tegmental area), amygdala, basal ganglia, cortex, and spinal cord (Borowsky et al., 2001). In peripheral tissues, mRNA encoding Taar1 is reported in pancreatic islet and white adipose cells (Regard et al., 2008). Immunohistochemistry has shown TA1 protein expression in the dopaminergic neurons of the substantia nigra (Xie et al., 2007).
In the rat, mRNA encoding TA1 has been found in the cardiac ventricular wall (Chiellini et al., 2007). Bunzow et al. (2001) showed the subcellular localization of the rat TA1 protein in a HEK293 expression system to be intracellular and punctate using immunocytochemistry. For experimental purposes, stable membrane expression of TA1 has been achieved using a human-rat chimera, with a modified coding sequence including an influenza-derived hemagglutinin leader sequence and rat TA1-derived N and C termini and third intracellular loop (Lindemann et al., 2005) or by the generation of an N-glycosylated human TA1 variant (Barak et al., 2008) that shows appropriate activation by β-PEA. The in vivo subcellular localization of TA1 is yet to be confirmed.
V. Radiolabeled Ligands
[3H]Tyramine (pKD = 7.7) has been available for a long time, but few studies have been carried out in expression systems of TA1 (Borowsky et al., 2001). Radioactive thyronamines (e.g., [125I]3-iodothyronamine) have been synthesized (Miyakawa and Scanlan, 2006) but as yet are neither pharmacologically characterized nor commercially available.
VI. Agonists
Endogenous trace amines act as agonists of the TA1 receptor, for example p-tyramine and β-PEA (pEC50 values at human receptor = 6.4–6.7, 6.2–7.0, respectively; Borowsky et al., 2001; Wainscott et al., 2007; Barak et al., 2008). In addition, the thyroid hormone derived thyronamines [e.g., 3-iodothyronamine (T1AM); pEC50 at rat receptor = 7.9; Scanlan et al., 2004; Hart et al., 2006] have affinity for TA1. In rat isolated perfused heart, trace amines and iodothyronamines are negative inotropic agents (Chiellini et al., 2007; Frascarelli et al., 2008), although the authors state that the rank order of agonist potency suggests that this is not a TA1 response but may be mediated by a different trace amine-associated receptor, several of which are expressed in rat heart (Frascarelli et al., 2008).
Of the classic biogenic amines, only dopamine is reported to bind to expressed human TA1 (Ki 422 nM compared with 8 nM for β-PEA and 34 nM for tyramine; Borowsky et al., 2001) and produced functional responses in cAMP assays in expressed human and rodent receptors in the micromolar range (Borowsky et al., 2001; Bunzow et al., 2001). In these studies, noradrenaline, adrenaline, and serotonin had a negligible effect in both binding and functional assays. It is noteworthy that the rhesus monkey TA1 receptor, which shows higher deduced amino acid sequence homology to the human receptor (96%) than do rodent receptors (<79% homology with human TA1) (Miller et al., 2005), responded to tyramine, β-PEA, octopamine, and tryptamine as expected (Miller et al., 2005; Xie and Miller, 2007) but also to comparable concentrations of dopamine, noradrenaline, and serotonin (Xie et al., 2007). The explanation and, indeed, the importance of this discrepancy is as yet unclear.
Exogenous agonists include amphetamine and its derivatives [e.g., S(+)methamphetamine (pEC50 at rat receptor = 6.1; Reese et al., 2007) and 3,4-methylenedioxymethamphetamine (pEC50 at rat receptor = 5.8; Bunzow et al., 2001)]. Both amphetamine and methamphetamine seem to show a species-dependent stereoselectivity, at least in expression systems (Bunzow et al., 2001; Reese et al., 2007) (see Table 3 and Fig. 2).
TABLE 3.
Selected agonists of the TA1 receptor and reported species-specific pEC50 values All values are for receptors in cell expression systems.
Fig. 2.
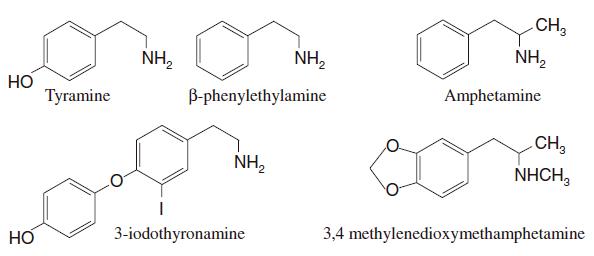
Examples of chemical structures of trace amine 1 (TA1) receptor endogenous and exogenous agonists.
VII. Antagonists
There are no reports yet of fully characterized antagonists of the TA1 receptor, and none are commercially available. Nonselective classical amine receptor antagonists have little TA1 blocking ability (Wainscott et al., 2007). Tan et al. (2008) were able to rationally design and synthesize lead compounds, taking into account the recently described rotamer toggle switch model of GPCR activation (Kobilka and Deupi, 2007; Rasmussen et al., 2007), concluding that a hexyloxy group and the outer or β-phenyl rings are essential for antagonism. It remains to be seen whether such compounds are specific and will become widely available in the near future.
VIII. Receptor Signaling
TA1 stably expressed in HEK293 or COS-7 cells has been shown to couple to Gs, leading to intracellular cAMP accumulation (Borowsky et al., 2001; Lindemann et al., 2005; Wolinsky et al., 2007) and stimulation of the cystic fibrosis transmembrane conductance regulator in Xenopus laevis oocytes (Borowsky et al., 2001). In addition, TA1 has been coupled to the promiscuous Gα16 in Chinese hamster ovary cells, producing an increase in the intracellular Ca2+ concentration as measured by fluorometry (Navarro et al., 2006; Lewin et al., 2008). Inhibitors of both protein kinases A and C have been shown to block TA1-mediated effects in synaptosomes (see section IX; Xie and Miller, 2007). In vivo signal transduction mechanisms are yet to be investigated.
IX. Physiological Role
Levels of trace amines have been measured in human plasma and are in the low nanomolar range (for example, see Zhou et al., 2001; D'Andrea et al., 2003). Tyramine and β-PEA have pKi values for human TA1 of 8.1 and 7.5, respectively (Borowsky et al., 2001) with pEC50 values in the high nanomolar range (6.2–7.0; Table 3). For comparison, the biogenic amines noradrenaline and adrenaline circulate at similar low nanomolar concentrations (Goldstein et al., 2003) but are functional at higher concentrations, with pKi of 6.0 to 6.5 for α1- and β1-adrenergic receptors and pEC50 values again in the high nanomolar range (NC-IUPHAR GPCR database: http://www.iuphar-db.org/GPCR/ReceptorFamiliesForward). Selective TA1 antagonists have not yet been developed but are required to confirm the precise physiology of this receptor system.
The role of the TA1 receptor is most understood in the central nervous system, where it is believed to modulate monoaminergic neurotransmission, thus affecting a number of neural networks and processes. β-PEA inhibits uptake and induces efflux of dopamine and serotonin in striatal synaptosomes and of norepinephrine in thalamic synaptosomes in vitro by interacting with transporters, for example the dopamine transporter (Xie and Miller, 2007, 2008; Xie et al., 2008). The effect is abolished in the TA1 knockout (Lindemann et al., 2008; Xie and Miller, 2008; Xie et al., 2008) and by inhibitors of protein kinases A and C (Xie and Miller, 2007). In addition, TA1 may be immunomodulatory, because mRNA encoding the receptor is up-regulated in circulating leukocytes after administration of the mitogen phytohemagglutinin (Nelson et al., 2007). Trace amines, β-PEA in particular, have long been associated with sustaining mood (Fischer and Heller, 1972; Sabelli and Mosnaim, 1974; Boulton, 1980), although a specific role for TA1 in this has yet to be elucidated. The effect of genetically disrupting synthesis of the endogenous agonists tyramine and β-PEA has not been assessed.
X. Pathophysiological Role
Large increases in plasma trace amine levels can occur in patients or animals on monoamine oxidase inhibitors, and alterations in levels have been reported in some diseases. The TA1 receptor has not been directly linked with any pathophysiological process, although trace amines are known to be associated with the hypertensive “beer, wine, and cheese effect” and are thought to play a role in psychiatric disorders such as schizophrenia (O'Reilly et al., 1991) and depression (Boulton, 1980; Premont et al., 2001) as well as primary headache (D'Andrea et al., 2004). Modulation of trace amine systems may be a potential therapeutic avenue (Branchek and Blackburn, 2003; Berry, 2007), particularly because the receptors are likely to be amenable as drug targets (Davenport, 2003). Linkage analysis has also shown a correlation between schizophrenia and polymorphisms in the chromosomal region encoding the trace amine-associated receptors (Levi et al., 2005; Pae et al., 2008a,b) but not TA1, polymorphisms of which are yet to be reported.
XI. Genetically Modified Animals
Deletion of the Taar1 gene in mice results in viable, fertile animals. They exhibit a phenotype characterized by minor spontaneous hyperactivity, reduced prepulse inhibition, increased sensitization to the psychomotor-stimulatory effects of amphetamine, raised levels of dopamine and norepinephrine in the dorsal striatum, increased striatal D2 receptor expression, and an elevated spontaneous firing rate of dopaminergic neurons in the ventral tegmental area compared with the wild type (Wolinsky et al., 2007; Sotnikova et al., 2008; Xie and Miller, 2008; Xie et al., 2008). This phenotype has been proposed as an animal model of schizophrenia (Wolinsky et al., 2007) and also as hemi-Parkinsonian (Sotnikova et al., 2008). In addition, in TA1-deficient mice, β-PEA was unable to modify the uptake or efflux of classic amine transmitters in striatal or thalamic synaptosomes as had been shown for wild type (Xie and Miller, 2008; Xie et al., 2008). The effect of genetically disrupting synthesis of the endogenous agonists tyramine and β-PEA has not been assessed, although because these compounds are also metabolites, they cannot readily be disrupted-out without identifying enzymes exclusive to their production.
Acknowledgments
This work was supported by British Heart Foundation [Grants PS/02/001, PG/05/127/19872]; and by the Intramural Research Program of the National Institutes of Health National Institute of Mental Health.
This article is available online at http://pharmrev.aspetjournals.org.
doi:10.1124/pr.109.001107.
Footnotes
Abbreviations: β-PEA, β-phenylethylamine; GPCR, G-protein coupled receptor; TA1, trace amine 1; IUPHAR, International Union of Pharmacology; HEK, human embryonic kidney.
References
- Arakawa S, Gocayne JD, McCombie WR, Urquhart DA, Hall LM, Fraser CM, and Venter JC (1990) Cloning, localization, and permanent expression of a Drosophila octopamine receptor. Neuron 4 343-354. [DOI] [PubMed] [Google Scholar]
- Axelrod J and Saavedra JM (1977) Octopamine. Nature 265 501-504. [DOI] [PubMed] [Google Scholar]
- Barak LS, Salahpour A, Zhang X, Masri B, Sotnikova TD, Ramsey AJ, Violin JD, Lefkowitz RJ, Caron MG, and Gainetdinov RR (2008) Pharmacological characterization of membrane-expressed human trace amine associated receptor 1 (TAAR1) by a bioluminescence resonance energy transfer (BRET) cAMP biosensor. Mol Pharmacol 74 585-594. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Barger G and Dale HH (1910) Chemical structure and sympathomimetic action of amines. J Physiol 41 19-59. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Berry MD (2007) The potential of trace amines and their receptors for treating neurological and psychiatric diseases. Rev Recent Clin Trials 2 3-19. [DOI] [PubMed] [Google Scholar]
- Blackwell B (1963) Hypertensive crisis due to monoamine-oxidase inhibitors. Lancet 26 849-850. [DOI] [PubMed] [Google Scholar]
- Borowsky B, Adham N, Jones KA, Raddatz R, Artymyshyn R, Ogozalek KL, Durkin MM, Lakhlani PP, Bonini JA, Pathirana S, et al. (2001) Trace amines: identification of a family of mammalian G protein-coupled receptors. Proc Natl Acad Sci U S A 98 8966-8971. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Boulton AA (1976) Identification, distribution, metabolism, and function of meta and para tyramine, phenylethylamine and tryptamine in brain. Adv Biochem Psychopharmacol 15 57-67. [PubMed] [Google Scholar]
- Boulton AA (1980) Trace amines and mental disorders. Can J Neurol Sci 7 261-263. [DOI] [PubMed] [Google Scholar]
- Bowsher RR and Henry DP (1983) Decarboxylation of p-tyrosine: a potential source of p-tyramine in mammalian tissues. J Neurochem 40 992-1002. [DOI] [PubMed] [Google Scholar]
- Branchek TA and Blackburn TP (2003) Trace amine receptors as targets for novel therapeutics: legend, myth and fact. Curr Opin Pharmacol 3 90-97. [DOI] [PubMed] [Google Scholar]
- Brier ME, Bowsher RR, Mayer PR, and Henry DP (1991) Conversion of p-tyrosine to p-tyramine in the isolated perfused rat kidney: modulation by perfusate concentrations of p-tyrosine. Life Sci 48 901-907. [DOI] [PubMed] [Google Scholar]
- Bunzow JR, Sonders MS, Arttamangkul S, Harrison LM, Zhang G, Quigley DI, Darland T, Suchland KL, Pasumamula S, Kennedy JL, et al. (2001) Amphetamine, 3,4-methylenedioxymethamphetamine, lysergic acid diethylamide, and metabolites of the catecholamine neurotransmitters are agonists of a rat trace amine receptor. Mol Pharmacol 60 1181-1188. [DOI] [PubMed] [Google Scholar]
- Caston JC, Eaton CL, Gheorghiu BP, and Ware LL (2002) Tyramine induced hypertensive episodes and panic attacks in hereditary deficient monoamine oxidase patients: case reports. J S C Med Assoc 98 187-192. [PubMed] [Google Scholar]
- Chaytor JP, Crathorne B, and Saxby MJ (1975) The identification and significance of 2-phenylethylamine in foods. J Sci Food Agric 26 593-598. [DOI] [PubMed] [Google Scholar]
- Chiellini G, Frascarelli S, Ghelardoni S, Carnicelli V, Tobias SC, DeBarber A, Brogioni S, Ronca-Testoni S, Cerbai E, Grandy DK, et al. (2007) Cardiac effects of 3-iodothyronamine: a new aminergic system modulating cardiac function. FASEB J 21 1597-1608. [DOI] [PubMed] [Google Scholar]
- Cooper AJ (1989) Tyramine and irreversible monoamine oxidase inhibitors in clinical practice. Br J Psychiatry Suppl 6 38-45. [PubMed] [Google Scholar]
- Dale HH and Dixon WE (1909) The action of pressor amines produced by putrefaction. J Physiol 39 25-44. [DOI] [PMC free article] [PubMed] [Google Scholar]
- D'Andrea G, Terrazzino S, Fortin D, Farruggio A, Rinaldi L, and Leon A (2003) HPLC electrochemical detection of trace amines in human plasma and platelets and expression of mRNA transcripts of trace amine receptors in circulating leukocytes. Neurosci Lett 346 89-92. [DOI] [PubMed] [Google Scholar]
- D'Andrea G, Terrazzino S, Leon A, Fortin D, Perini F, Granella F, and Bussone G (2004) Elevated levels of circulating trace amines in primary headaches. Neurology 62 1701-1705. [DOI] [PubMed] [Google Scholar]
- Davenport AP (2003) Peptide and trace amine orphan receptors: prospects for new therapeutic targets. Curr Opin Pharmacol 3 127-134. [DOI] [PubMed] [Google Scholar]
- David JC, Dairman W, and Udenfriend S (1974) Decarboxylation to tyramine: a major route of tyrosine metabolism in mammals. Proc Natl Acad Sci U S A 71 1771-1775. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Duan J, Martinez M, Sanders AR, Hou C, Saitou N, Kitano T, Mowry BJ, Crowe RR, Silverman JM, Levinson DF, et al. (2004) Polymorphisms in the trace amine receptor 4 (TRAR4) gene on chromosome 6q23.2 are associated with susceptibility to schizophrenia. Am J Hum Genet 75 624-638. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Durden DA and Philips SR (1980) Kinetic measurements of the turnover rates of phenylethylamine and tryptamine in vivo in the rat brain. J Neurochem 34 1725-1732. [DOI] [PubMed] [Google Scholar]
- Fischer E and Heller B (1972) Phenylethylamine as a neurohumoral agent in brain. Behav Neuropsychiatry 4 8-11. [PubMed] [Google Scholar]
- Foord SM, Bonner TI, Neubig RR, Rosser EM, Pin JP, Davenport AP, Spedding M, and Harmar AJ (2005) International Union of Pharmacology. XLVI. G protein-coupled receptor list. Pharmacol Rev 57 279-288. [DOI] [PubMed] [Google Scholar]
- Frascarelli S, Ghelardoni S, Chiellini G, Vargiu R, Ronca-Testoni S, Scanlan TS, Grandy DK, and Zucchi R (2008) Cardiac effects of trace amines: pharmacological characterization of trace amine-associated receptors. Eur J Pharmacol 587 231-236. [DOI] [PubMed] [Google Scholar]
- Gloriam DE, Bjarnadóttir TK, Schiöth HB, and Fredriksson R (2005) High species variation within the repertoire of trace amine receptors. Ann N Y Acad Sci 1040 323-327. [DOI] [PubMed] [Google Scholar]
- Goldstein DS, Eisenhofer G, and Kopin IJ (2003) Sources and significance of plasma levels of catechols and their metabolites in humans. J Pharmacol Exp Ther 305 800-811. [DOI] [PubMed] [Google Scholar]
- Grandy DK (2007) Trace amine-associated receptor 1: family archetype or iconoclast? Pharmacol Ther 116 355-390. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hannah P, Glover V, and Sandler M (1988) Tyramine in wine and beer. Lancet 1 879. [DOI] [PubMed] [Google Scholar]
- Hart ME, Suchland KL, Miyakawa M, Bunzow JR, Grandy DK, and Scanlan TS (2006) Trace amine-associated receptor agonists: synthesis and evaluation of thyronamines and related analogues. J Med Chem 49 1101-1112. [DOI] [PubMed] [Google Scholar]
- Hashiguchi Y and Nishida M (2007) Evolution of trace amine associated receptor (TAAR) gene family in vertebrates: lineage-specific expansions and degradations of a second class of vertebrate chemosensory receptors expressed in the olfactory epithelium. Mol Biol Evol 24 2099-2107. [DOI] [PubMed] [Google Scholar]
- Kidd M, Modlin IM, Gustafsson BI, Drozdov I, Hauso O, and Pfragner R (2008) The luminal regulation of normal and neoplastic human EC cell serotonin release is mediated by bile salts, amines, tastants and olfactants. Am J Physiol Gastrointest Liver Physiol 295 G260-G272. [DOI] [PubMed] [Google Scholar]
- Kobilka BK and Deupi X (2007) Conformational complexity of G-protein-coupled receptors. Trends Pharmacol Sci 28 397-406. [DOI] [PubMed] [Google Scholar]
- Levi A, Kohn Y, Kanyas K, Amann D, Pae CU, Hamdan A, Segman RH, Avidan N, Karni O, Korner M, et al. (2005) Fine mapping of a schizophrenia susceptibility locus at chromosome 6q23: increased evidence for linkage and reduced linkage interval. Eur J Hum Genet 13 763-771. [DOI] [PubMed] [Google Scholar]
- Lewin AH (2006) Receptors of mammalian trace amines. AAPS J 8 E138-E145. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lewin AH, Navarro HA, and Mascarella SW (2008) Structure-activity correlations for beta-phenylethylamines at human trace amine receptor 1. Bioorg Med Chem 16 7415-7423. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Liberles SD and Buck LB (2006) A second class of chemosensory receptors in the olfactory epithelium. Nature 442 645-650. [DOI] [PubMed] [Google Scholar]
- Lindemann L, Ebeling M, Kratochwil NA, Bunzow JR, Grandy DK, and Hoener MC (2005) Trace amine-associated receptors form structurally and functionally distinct subfamilies of novel G protein-coupled receptors. Genomics 85 372-385. [DOI] [PubMed] [Google Scholar]
- Lindemann L and Hoener MC (2005) A renaissance in trace amines inspired by a novel GPCR family. Trends Pharmacol Sci 26 274-281. [DOI] [PubMed] [Google Scholar]
- Lindemann L, Meyer CA, Jeanneau K, Bradaia A, Ozmen L, Bluethmann H, Bettler B, Wettstein JG, Borroni E, Moreau JL, et al. (2008) Trace amine-associated receptor 1 modulates dopaminergic activity. J Pharmacol Exp Ther 324 948-956. [DOI] [PubMed] [Google Scholar]
- Miller GM, Verrico CD, Jassen A, Konar M, Yang H, Panas H, Bahn M, Johnson R, and Madras BK (2005) Primate trace amine receptor 1 modulation by the dopamine transporter. J Pharmacol Exp Ther 313 983-994. [DOI] [PubMed] [Google Scholar]
- Miyakawa M and Scanlan TS (2006) Synthesis of [125I]-, [2H]-, and [3H]-labeled 3-iodothyronamine (T1AM). Synth Commun 36 891-902. [Google Scholar]
- Mueller JC, Steiger S, Fidler AE, and Kempenaers B (2008) Biogenic Trace Amine-Associated Receptors (TAARS) are encoded in avian genomes: evidence and possible implications. J Hered 99 174-176. [DOI] [PubMed] [Google Scholar]
- Navarro HA, Gilmour BP, and Lewin AH (2006) A rapid functional assay for the human trace amine-associated receptor 1 based on the mobilization of internal calcium. J Biomol Screen 11 688-693. [DOI] [PubMed] [Google Scholar]
- Nelson DA, Tolbert MD, Singh SJ, and Bost KL (2007) Expression of neuronal trace amine-associated receptor (Taar) mRNAs in leukocytes. J Neuroimmunol 192 21-30. [DOI] [PMC free article] [PubMed] [Google Scholar]
- O'Reilly R, Davis BA, Durden DA, Thorpe L, Machnee H, and Boulton AA (1991) Plasma phenylethylamine in schizophrenic patients. Biol Psychiatry 30 145-150. [DOI] [PubMed] [Google Scholar]
- Pae CU, Yu HS, Amann D, Kim JJ, Lee CU, Lee SJ, Jun TY, Lee C, Paik IH, Patkar AA, et al. (2008a) Association of the trace amine associated receptor 6 (TAAR6) gene with schizophrenia and bipolar disorder in a Korean case control sample. J Psychiatr Res 42 35-40. [DOI] [PubMed] [Google Scholar]
- Pae CU, Drago A, Kim JJ, Patkar AA, Jun TY, Lee C, Mandelli L, De Ronchi D, Paik IH, and Serretti A (2008b) TAAR6 variation effect on clinic presentation and outcome in a sample of schizophrenic in-patients: An open label study. Eur Psychiatry 23 390-395. [DOI] [PubMed] [Google Scholar]
- Paterson IA, Juorio AV, and Boulton AA (1990) 2-Phenylethylamine: a modulator of catecholamine transmission in the mammalian central nervous system? J Neurochem 55 1827-1837. [DOI] [PubMed] [Google Scholar]
- Premont RT, Gainetdinov RR, and Caron MG (2001) Following the trace of elusive amines. Proc Natl Acad Sci U S A 98 9474-9475. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rasmussen SG, Choi HJ, Rosenbaum DM, Kobilka TS, Thian FS, Edwards PC, Burghammer M, Ratnala VR, Sanishvili R, Fischetti RF, et al. (2007) Crystal structure of the human beta2 adrenergic G-protein-coupled receptor. Nature 450 383-387. [DOI] [PubMed] [Google Scholar]
- Reese EA, Bunzow JR, Arttamangkul S, Sonders MS, and Grandy DK (2007) Trace amine-associated receptor 1 displays species-dependent stereoselectivity for isomers of methamphetamine, amphetamine, and para-hydroxyamphetamine. J Pharmacol Exp Ther 321 178-186. [DOI] [PubMed] [Google Scholar]
- Regard JB, Kataoka H, Cano DA, Camerer E, Yin L, Zheng YW, Scanlan TS, Hebrok M, and Coughlin SR (2007) Probing cell type-specific functions of Gi in vivo identifies GPCR regulators of insulin secretion. J Clin Invest 117 4034-4043. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Regard JB, Sato IT, and Coughlin SR (2008) Anatomical profiling of G protein-coupled receptor expression. Cell 135 561-571. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Roeder T (2005) Tyramine and octopamine: ruling behavior and metabolism. Annu Rev Entomol 50 447-477. [DOI] [PubMed] [Google Scholar]
- Sabelli HC and Mosnaim AD (1974) Phenylethylamine hypothesis of affective behavior. Am J Psychiatry 131 695-699. [DOI] [PubMed] [Google Scholar]
- Saudou F, Amlaiky N, Plassat JL, Borrelli E, and Hen R (1990) Cloning and characterization of a Drosophila tyramine receptor. EMBO J 9 3611-3617. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Scanlan TS, Suchland KL, Hart ME, Chiellini G, Huang Y, Kruzich PJ, Frascarelli S, Crossley DA, Bunzow JR, Ronca-Testoni S, et al. (2004) 3-Iodothyronamine is an endogenous and rapid-acting derivative of thyroid hormone. Nat Med 10 638-642. [DOI] [PubMed] [Google Scholar]
- Schöneberg T, Hofreiter M, Schulz A, and Römpler H (2007) Learning from the past: evolution of GPCR functions. Trends Pharmacol Sci 28 117-221. [DOI] [PubMed] [Google Scholar]
- Sotnikova TD, Zorina OI, Ghisi V, Caron MG, and Gainetdinov RR (2008) Trace amine associated receptor 1 and movement control. Parkinsonism Relat Disord 14(Suppl 2): S99-S102. [DOI] [PubMed] [Google Scholar]
- Tan ES, Groban ES, Jacobson MP, and Scanlan TS (2008) Toward deciphering the code to aminergic G protein-coupled receptor drug design. Chem Biol 15 343-353. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Vanti WB, Muglia P, Nguyen T, Cheng R, Kennedy JL, George SR, and O'Dowd BF (2003) Discovery of a null mutation in a human trace amine receptor gene. Genomics 82 531-536. [DOI] [PubMed] [Google Scholar]
- Wainscott DB, Little SP, Yin T, Tu Y, Rocco VP, He JX, and Nelson DL (2007) Pharmacologic characterization of the cloned human trace amine-associated receptor1 (TAAR1) and evidence for species differences with the rat TAAR1. J Pharmacol Exp Ther 320 475-485. [DOI] [PubMed] [Google Scholar]
- Wolinsky TD, Swanson CJ, Smith KE, Zhong H, Borowsky B, Seeman P, Branchek T, and Gerald CP (2007) The trace amine 1 receptor knockout mouse: an animal model with relevance to schizophrenia. Genes Brain Behav 6 628-639. [DOI] [PubMed] [Google Scholar]
- Xie Z and Miller GM (2007) Trace amine-associated receptor 1 is a modulator of the dopamine transporter. J Pharmacol Exp Ther 321 128-136. [DOI] [PubMed] [Google Scholar]
- Xie Z and Miller GM (2008) β-phenylethylamine alters monoamine transporter function via trace amine-associated receptor 1: implication for modulatory roles of trace amines in brain. J Pharmacol Exp Ther 325 617-628. [DOI] [PubMed] [Google Scholar]
- Xie Z, Westmoreland SV, Bahn ME, Chen GL, Yang H, Vallender EJ, Yao WD, Madras BK, and Miller GM (2007) Rhesus monkey trace amine-associated receptor 1 signaling: enhancement by monoamine transporters and attenuation by the D2 autoreceptor in vitro. J Pharmacol Exp Ther 321 116-127. [DOI] [PubMed] [Google Scholar]
- Xie Z, Westmoreland SV, and Miller GM (2008) Modulation of monoamine transporters by common biogenic amines via trace amine-associated receptor 1 and monoamine autoreceptors in human embryonic kidney 293 cells and brain synaptosomes. J Pharmacol Exp Ther 325 629-640. [DOI] [PubMed] [Google Scholar]
- Zhou G, Miura Y, Shoji H, Yamada S, and Matsuishi T (2001) Platelet monoamine oxidase B and plasma beta-phenylethylamine in Parkinson's disease. J Neurol Neurosurg Psychiatry 70 229-231. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zucchi R, Chiellini G, Scanlan TS, and Grandy DK (2006) Trace amine-associated receptors and their ligands. Br J Pharmacol 149 967-978. [DOI] [PMC free article] [PubMed] [Google Scholar]