

Abstract
Curcumin, a plant polyphenol, is a widely studied chemopreventive agent with demonstrated antitumor activities in preclinical studies and low toxicity profiles in multiple clinical trials against human malignancies. We previously demonstrated that curcumin radiosensitizes cervical tumor cells without increasing the cytotoxic effects of radiation on normal human fibroblasts. Here we report that an inhibitory activity of curcumin on the anti-oxidant enzyme Thioredoxin Reductase-1 (TxnRd1) is required for curcumin-mediated radiosensitization of squamous carcinoma cells. Stable knockdown of TxnRd1 in both HeLa and FaDu cells nearly abolished curcumin-mediated radiosensitization. TxnRd1 knockdown cells demonstrated decreased radiation-induced reactive oxygen species and sustained ERK1/2 activation, which we previously demonstrated was required for curcumin-mediated radiosensitization. Conversely, overexpressing catalytically active TxnRd1 in HEK293 cells, with low basal levels of TxnRd1, increased their sensitivity to curcumin alone and to the combination of curcumin and ionizing radiation. These results demonstrate the critical role of TxnRd1 in curcumin-mediated radiosensitization and suggest that TxnRd1 levels in tumors could have clinical value as a predictor of response to curcumin and radiotherapy.
Keywords: Thioredoxin reductase-1(TxnRd1), curcumin, radiosensitization
Introduction
The success of radiotherapy depends on the ability to target malignant cells with radiation while simultaneously protecting surrounding normal tissues from its cytotoxic effects. One pharmacological approach for increasing the clinical response to radiotherapy is the use of radiation response modifiers that either preferentially sensitize tumor cells or protect normal tissues from radiation-induced damage. Curcumin, a naturally occurring plant polyphenol, has an extensive history of use in diet and traditional medicine. Several preclinical studies carried out over the last 20 years have demonstrated direct anti-tumorigenic activity in mouse tumor models (1). Curcumin's anti-tumor properties occur through pleiotropic mechanisms including inhibition of proliferation, induction of apoptosis, inhibition of angiogenesis, immune modulation and suppression of inflammation (Reviewed in 2). Curcumin has a low toxicity profile and has been well tolerated fin multiple phase I and II clinical trials for malignant and non- malignant diseases (2).
Several groups, including our own, have shown that curcumin selectively radiosensitizes tumor cells (3–5) without enhancing the effects of radiation on non-transformed cells or normal tissue (6, 7). We previously demonstrated that curcumin sensitized cervical tumor cells but not normal fibroblasts to ionizing radiation (IR). Mechanistically, we found that pretreatment of tumor cells with curcumin and IR resulted in elevated levels of reactive oxygen species (ROS) leading to sustained activation ERK1/2-MAPK and that both were required for curcumin-mediated radiosensitization (3). However, the molecular mechanism accounting for the differential effects of curcumin in fully transformed and normal cells remained unclear.
Thioredoxin Reductase 1 (TxnRd1) is one of several identified molecular targets of curcumin (16). The thioredoxin reductases are essential mammalian selenocysteine containing flavoenzymes that act in homodimeric form to catalyze NADPH-dependent reduction of thioredoxin and small molecular weight oxidants including ROS (8, 9). TxnRd1 and TxnRd2 (in cytosolic/nuclear and mitochondrial form respectively) are two ubiquitously expressed isoforms of this enzyme family. They play a key role in maintaining redox-regulated cellular functions including transcription, DNA damage recognition and repair, proliferation and apoptosis (8). In response to oxidative stress, TxnRds sustain signaling pathways that regulate transcription of genes to protect the cell from oxidative damage (10, 11, 12). Cytosolic TxnRd1 expression is often upregulated in human cancers where it is associated with aggressive tumor growth and poor prognosis (13). TxnRd1 has been shown to confer protection against the lethal effects of IR in tumor cells (10). Moreover, agents that selectively target TxnRd1 have shown promising results as anticancer drugs in preclinical and clinical studies (14, 15) when used alone or when combined with IR (11). Fang et al, recently (16) demonstrated that Curcumin covalently binds to the selenocysteine residue of TxnRd1 (16), thereby inhibiting its catalytic activity.
Based on these findings and our previous studies (3) we hypothesized that curcumin-mediated inhibition of TxnRd1 activity could enhance radiation sensitivity by decreasing the capacity of TxnRd1 to scavenge IR-induced ROS and/or decrease the activity of pro-survival signaling cascades activated by IR. We report that knockdown of TxnRd1 in squamous carcinoma cell lines significantly decreases curcumin-mediated radiosensitization. Conversely, overexpression in cells with low basal levels of TxnRd1 increased their sensitivity to curcumin alone and to the combination of curcumin and IR. These results demonstrate that TxnRd1 is required for curcumin-mediated radiosensitization.
Methods & Materials
Cell culture
HeLa, SiHa, FaDu (ATCC, Manassas, VA), SCC-1 (a gift from Dr. Bert O’Malley Jr., University of Pennsylvania, PA), HEK293-pIRES and HEK293-TxnRd1 (a gift from Dr. Giannis Spyrou, Academy of Athens, Greece) cell lines were grown in Dulbecco's modified Eagle's medium supplemented with penicillin/streptomycin, 10% fetal bovine serum. MSK-Leuk1 (a gift from Dr. Peter Sacks and Memorial Sloane Kettering Hospital) and Keratinocytes (Invitrogen, Carlsbad, CA) were maintained in Defined Keratinocyte Serum Free Media with growth factor supplements.
Reagents
All chemical reagents were obtained from Sigma-Aldrich, St.Louis, MO. Curcumin and tetrahydrocurcumin (THC) were dissolved in DMSO (10 mM stock solution). Puromycin and G418 were dissolved in water (1 mg/ml and 200 mg/ml stock solutions respectively). 2', 7'-dichlorofluorescein diacetate (DCF-DA) was dissolved in DMSO (20 mM stock solution).
Generation of stable TxnRd1 knockdown cells
A set of non-targeting shRNA (shNT) and four shRNA plasmids targeting different coding regions of the TxnRd1 gene (shTR1) were purchased (SA Biosciences, KH02104P). HeLa and FaDu cells were transfected using Lipofectamine 2000 reagent (Invitrogen). At 48 h post-transfection, equal numbers of cells were plated from mock and plasmid transfected plates. 24 h post-plating, cells were treated with 2 µg/ml puromycin for 48 h and clones were allowed to grow. Selected clones were maintained in 0.5 µg/ml puromycin.
Clonogenic survival assays
(Low density). HeLa cells were plated in 60mm dishes at low densities and allowed to attach overnight. Cells were irradiated in the presence of curcumin or DMSO, using a 137Cs source (dose rate of 1.3 Gy/min). Colony formation, fixation and counting were performed as described (3). (High density). FaDu cells were plated in 60mm dishes at high densities and allowed to attach overnight. Cells were pretreated for 8 h with curcumin and then irradiated using a 137Cs source (dose rate of 1.3 Gy/min). 1 h after irradiation cells were trypsinized and subcultured at low densities. Colony formation, fixation and counting were performed as described (3). For both types of the assay, survival curves were fitted using a second order polynomial function. The average normalized surviving fraction from 3 independent experiments were reported (±S.E.). DER values were calculated at 0.37 survival fraction. Survival curves were fitted using a second order polynomial function. The average normalized surviving fraction from 3 independent experiments was reported (±S.E.).
Cell growth assay (MTT assay)
MTT assays were performed using the cell proliferation kit (Roche Applied Sciences, Germany). Cells were plated in 24-well plates and treated with different curcumin doses for 8 h. Cell viability was assessed according to the manufacturer’s protocol when the untreated controls reached confluence. 200 µl aliquots from each well were transferred into a 96-well plate and absorbance measured at 570 nm with an automated plate reader (Thermo Scientific, Sunnyvale, CA).
Analysis of ROS levels
HeLa clones (shNT and shTR1) were treated with DMSO or 10 µM curcumin for 8 h. Cells were incubated with 20 µM DCF-DA during the last 30 min of treatment at 37°C. The probe was washed off with PBS and the cells were either mock irradiated or exposed to a 4Gy dose of IR. 30 min later, the cells were trypsinized, resuspended in PBS and analyzed for DCF fluorescence by flow cytometry (FACS-Calibur, CellQuest Pro; BD Biosciences).
Immunoblotting
Whole cell lysates were obtained as described previously (22). For detection of TxnRd1, TxnRd2, pERK1/2, ERK1/2 and β-actin, 20 – 50 µg of total protein was resolved on 10% SDS-PAGE gels, transferred to polyvinylidene difluoride (PVDF) membrane, blocked for 30 min in TBS-Tween solution containing 5% milk and incubated for 1–2 h with the following antibodies in TBS-Tween solution containing 1% milk: anti-TxnRd1 antibody (Abcam, MA), anti-TxnRd2 antibody (Abcam, MA), anti-β-actin (Sigma), anti-pERK1/2 and anti-ERK1/2 (Cell signaling, Boston, MA). All membranes were incubated with horseradish peroxidase-conjugated anti-mouse or anti-rabbit secondary antibodies (Santa Cruz Biotechnology, Santa Cruz, CA), and immunoreactive bands were detected using ECL-Plus chemiluminescence (GE Healthcare, UK).
Real time RT-PCR
RNA was isolated from cells following TRI Reagent protocol (Invitrogen). Reverse transcription was performed using AMV Reverse Transcriptase (Promega). Real time PCR was done on Applied Biosystems 7300 Real-Time PCR System using Power SYBR® Green PCR Master Mix. The primer sequences for detecting human TxnRd1 cDNA were: ccactggtgaaagaccacgtt (forward), aggagaaaagatcatcactgctgat (reverse). Primer sequences for 18S rRNA were aaacggctaccacatccaag (forward), caattacagggcctcgaaag (reverse)
TxnRd activity assay
Cells were plated to 90% confluence, allowed to attach overnight and treated with 10 µM curcumin or THC for 8 h. Following treatment, media was aspirated and replaced with 0.5 ml of media with Na-R-Lipoate (Na-RALA, GeroNova Research Inc) at a final concentration of 1 mM. After a 2 hour incubation, an aliquout was removed from the media overlying the cell monolayer and spun at 1000 g for 5 min. to get rid of cellular debris. 50 µl of cleared media was then added to 100 µl of 5, 5'-Dithio-bis (2-nitrobenzoic acid) (DTNB) solution in triplicates wells of a 96 well plate. The plates were incubated for 10 min in the dark. Absorbance was measured at 450nm using a plate reader (Thermo Scientific, Sunnyvale, CA). TxnRd activity was defined by the concentration of Sodium-Lipoate (disulfide) reduced to dehydrolipoate (dithiol) per mg protein.
Statistical Analysis
For all the clonogenic survival assays, mean and standard error (SEM) of survival fraction (SF) from 3 independent experiments at 4 and 6 Gy for each treatment group were calculated. ANOVA was used to determine whether there were significant differences among 3 or more independent experimental conditions, specific pairwise comparisons were evaluted by planned contrasts. Student’s t test was employed to test for differences between 2 independent experimental conditions. Statistical significance was set at 0.05. All tests were two-sided. Statistical analyses were performed with SPSS Version 17 software (SPSS, Inc., Chicago, IL).
Results
Tumor cells express higher levels of TxnRd1 compared to normal or minimally transformed cells
TxnRd1 expression is shown to be elevated in several primary human malignancies and in a number of human cancer cell lines (13, 17). A subset of aggressive head and neck cancers also exhibit elevated expression of TxnRd1 mRNA and protein when compared to normal mucosa (13). We analyzed the expression of TxnRd1 protein levels and activity in various squamous carcinoma cell lines (FaDu, SCC-1 and HeLa) and compared it to that in primary keratinocytes and in MSK-Leuk1, a pre-malignant cell line derived from a human oral leukoplakia. Based on immunoblot analysis, TxnRd1 expression was elevated by >50 fold in FaDu and HeLa cells, and by >20 fold in SCC-1 compared to either MSK-Leuk1 or keratinocytes (Fig 1A). Total TxnRd activity was determined using an assay based on work by Arner et al (18) and Biaglow et al (19). These investigators demonstrated that mammalian TxnRds reduce lipoate to dehydrolipoate more efficiently than lipoate dehydrogenase (18) and that the reduction of lipoate in intact tumor cells was largely dependent on thioredoxin reductase activity (19). As shown in Fig 1B, basal TxnRd activity correlated with levels of TxnRd1 protein; TxnRd activity was ~20 fold higher in FaDu and HeLa cells and ~7 fold higher in SCC-1 compared to that in Keratinocytes or MSK-Leuk1 cells.
Figure 1. TxnRd1 protein and activity levels in cells with different transformation status correlate with response to curcumin.
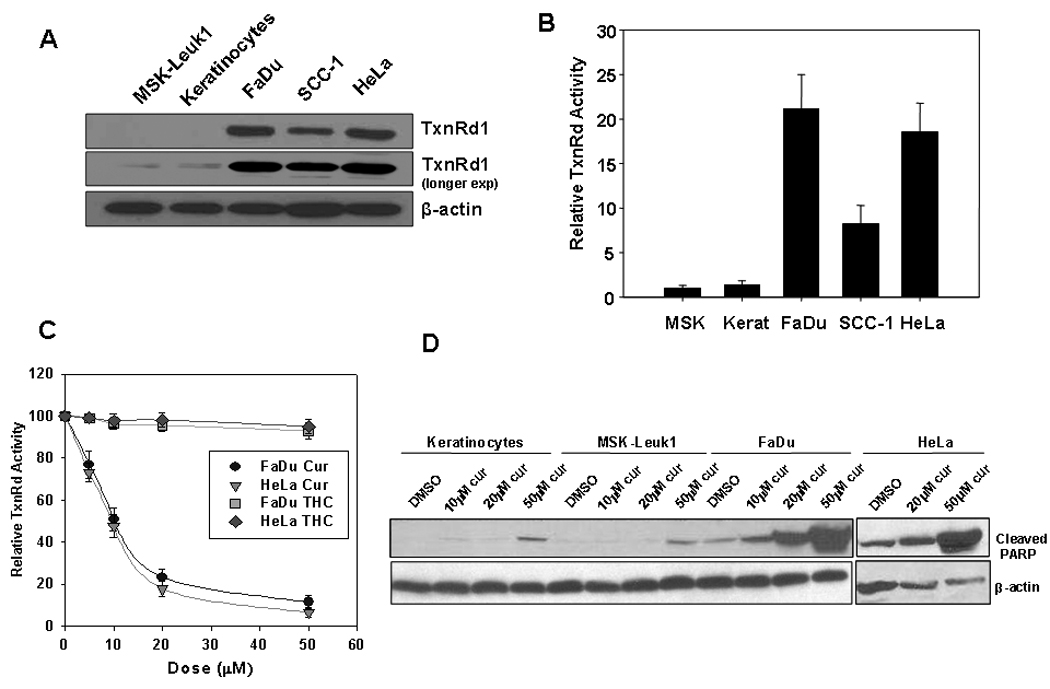
MSK-Leuk1, human keratinocytes and three squamous carcinoma cell lines: HeLa (cervical), FaDu and SCC-1 (Head & Neck) were analyzed for (A) TxnRd1 protein levels by immunoblot analysis. β-actin was used as a loading control. (B) Basal levels of whole cell TxnRd activity. Activity was measured as nmoles dihydrolipoate formed per mg protein and was normalized to that in MSK-Leuk1 cells. Results represent average of 3 independent experiments (±S.E.) (C) The effect of curcumin on TxnRd activity. FaDu and HeLa cells were treated with 0, 5, 10, 20 and 50 µM curcumin or THC for 8 h and assayed for TxnRd activity. Activity was measured as nmoles dihydrolipoate formed per mg protein and normalized to own cells untreated control. (D) Keratinocytes, MSK-Leuk1, FaDu and HeLa cells were treated with DMSO, 10, 20 or 50 µM curcumin for 8 h. Whole-cell lysates were analyzed for curcumin-induced apoptosis by immunoblot assay using antibodies against cleaved PARP and β-actin.
Curcumin inhibits TxnRd activity and enhances apoptosis in a manner dependent on transformation status
We subsequently examined the ability of curcumin to inhibit TxnRd activity in HeLa and FaDu cells. Curcumin treatment resulted in a dose-dependent decrease in TxnRd activity with an IC50 of approximately 10 µM in both cell lines (Fig 1C). These results are in agreement with those of Fang et. al., where TxnRd1 activity was measured by assaying the reduction of insulin in curumin-treated and untreated HeLa cell lysates (16). In that study, the Michael acceptor function of curcumin was proposed to mediate the inhibition of TxnRd activity. To test this possibility more stringently, we used tetra-hydro curcumin (THC), a curcumin metabolite lacking the α,β-unsaturated carbon double bond that is presumably required for curcumin to covalently inhibit TxnRd1. As predicted, THC had no significant effect on TxnRd activity at doses up to 50 µM (Fig. 1C).
Curcumin has been shown to selectively induce apoptosis in transformed cells (21). Therefore, we determined the effects of curcumin on levels of cleaved PARP, a marker of apoptosis, in keratinocytes (normal untransformed cells), MSK-Leuk1 (minimally transformed), FaDu and HeLa cells (fully transformed) at 24 h post-curcumin treatment. Levels of cleaved PARP did not appreciably increase in keratinocytes or MSK-Leuk1 cells treated with curcumin doses up to 20 µM. However, there was a modest increase in cleaved PARP observed with 50 µM curcumin. In contrast, curcumin induced a robust increase in cleaved PARP in both FaDu and HeLa cells. This increase exhibited a dose-dependence which was evident at doses as low as 10 µM in FaDu (Fig 1D). In accordance with reports that THC retains the antioxidant effects but not the pro-apoptotic activity of the parental drug (20), we did not observe an increase in cleaved PARP in any of the cells lines treated with THC doses up to 50 µM (data not shown). These results demonstrate that curcumin enhances apoptosis preferentially in fully-transformed squamous carcinoma cells and that this activity parallels its inhibitory activity on TxnRd1.
Efficient knockdown of TxnRd1 in squamous carcinoma cell lines
To test whether TxnRd1 is required for curcumin-mediated radiosensitization, we generated stable clones from both the HeLa and FaDu cell lines expressing an shRNA targeting a unique sequence in TxnRd1 gene (shTR1) or a non-targeting shRNA (shNT). For HeLa cells, two stable clones (shTR1–3.2, shTR1–3.3) were selected using shRNA-3 while another two stable clones (shTR1–1.2, shTR1–1.5) were selected using shRNA-1. For FaDu cells, one stable clone (shTR1–1.1) was selected using shRNA-1 and one stable clone using (shTR1–2.1) was selected using shRNA-2. All six knockdown clones exhibited >70% reduction in TxnRd1 mRN A l e vels compared to corresponding shNT clones transfected with the shNT plasmid (Fig 2A). Consistent with mRNA levels, the HeLa-shTR1 clones demonstrated over a 90% decrease (Fig 2B, left panel) and FaDu-shTR1 clones demonstrated an 80% decrease (Fig 2B, right panel) in TxnRd1 protein levels as determined by immunoblot analysis. Moreover, there was a 50% reduction of TxnRd activity observed in HeLa-shTR1 clones (Fig 2C, left panel) and 40% reduction in FaDu-shTR1 clones (Fig 2C, right panel) compared to the respective shNT clones. The residual activity most likely reflects activity due to the mitochondrial TxnRd2 isoform, whose levels are not substantially affected by these shRNA sequences (Fig 2B).
Figure 2. Stable knockdown of TxnRd1 in HeLa and FaDu squamous carcinoma cell lines.
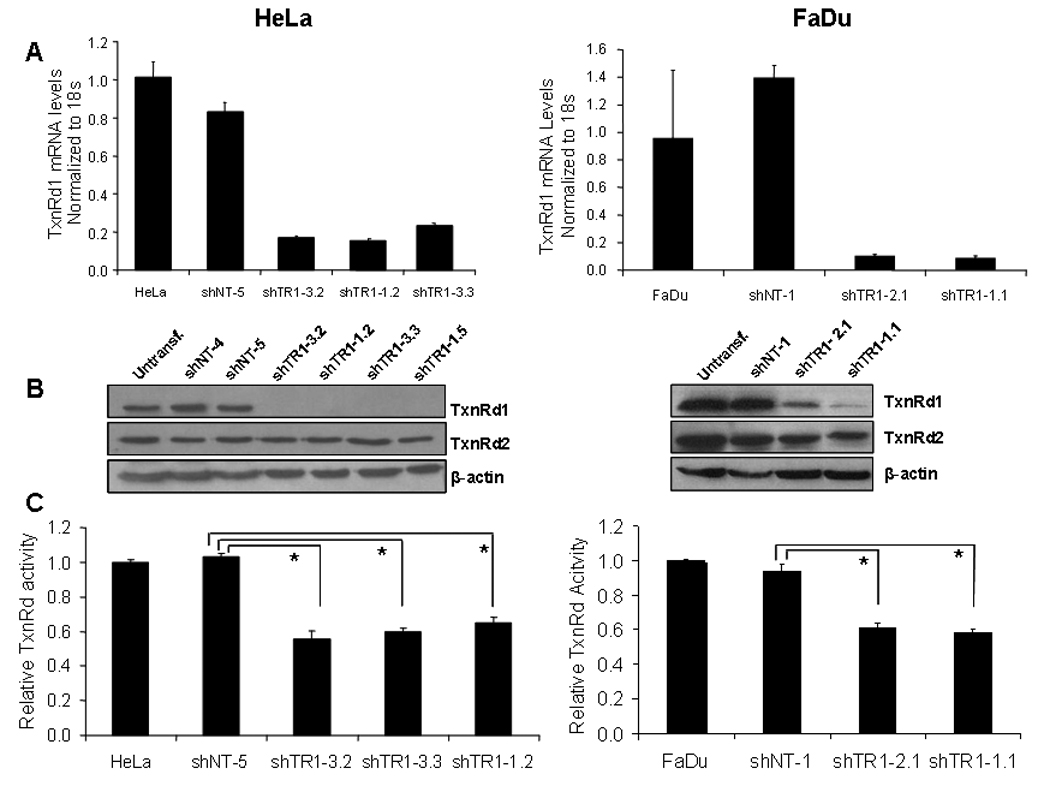
Stable clones were generated from HeLa (left panel) and FaDu (right panel) cells by expressing a non-targeting shRNA (shNT) or an shRNA targeting human TxnRd1 (shTR1). Untransfected cells, shNT and shTR1 clones were analyzed for (A) TxnRd1 mRNA levels by real time RT-PCR analysis. TxnRd1 mRNA levels were normalized to 18s rRNA control and normalized to mRNA levels in untransfected HeLa or FaDu cells. PCR reactions were done in triplicate (±S.E.; *p<0.05). (B) TxnRd1 protein levels assayed by immunoblot analysis. β-actin was used as a loading control. (C) TxnRd enzymatic activity. Results represent the average of 3 independent experiments (±S.E; *p<0.05).
Stable TxnRd1 knockdown in squamous carcinoma cells inhibits curcumin-mediated radiosensitization
To determine whether TxnRd1 is involved in the enhancement of the cytotoxic response of curcumin in combination with IR, we performed clonogenic survival assays on shNT and shTR1 clones from HeLa, SiHa (another cervical, squamous carcinoma cell line) and FaDu cells. As predicted from our previous report (3), HeLa, SiHa and FaDu cells stably expressing control shRNA (shNT clones) were significantly sensitized to IR by pretreatment with curcumin. The dose enhancement ratios (DERs) at a surviving fraction of 0.37 ranged from 1.32–1.92 (Fig 3A–D and Table 1). Knocking down TxnRd1 in HeLa and FaDu cells resulted in enhanced sensitivity to IR (ANOVA, p<0.05), in agreement with previous reports (11) at 6 Gy (Fig 3 A–D). Notably, curcumin failed to increase the radiosensitivity in the HeLa- and FaDu shTR1 clones (Figs 3A–D). Interestingly, in one of the HeLa knockdown clones (shTR1–3.2), curcumin induced a statistically significant radioprotective effect (Fig. 3A). Taken together, these results indicate that TxnRd1 is required for curcumin-mediated radiosensitization of squamous carcinoma cell lines.
Figure 3. Squamous carcinoma cells with TxnRd1 knockdown are less sensitive to curcumin-mediated radiosensitization.
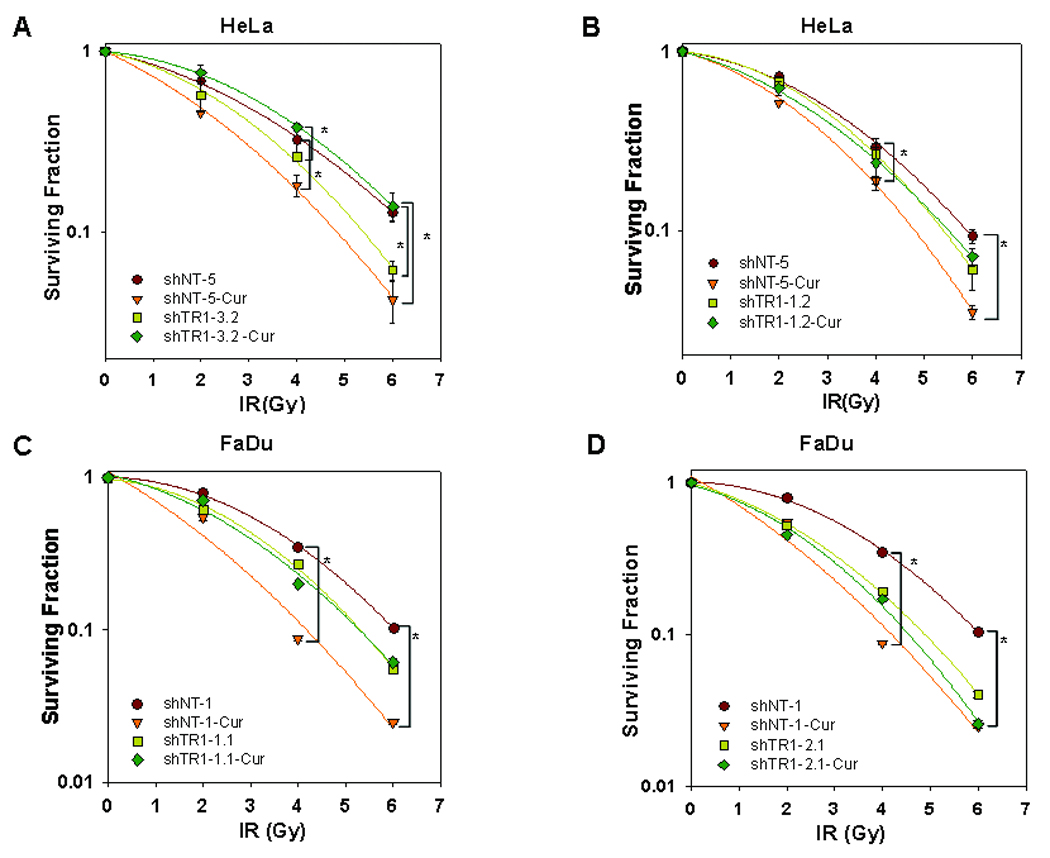
HeLa (A, B) and FaDu (C, D)-shNT control and shTR1 clones were treated with DMSO or 10 µM curcumin for 8 h followed by 0, 2, 4 or 6 Gy doses of IR. Survival was assessed by clonogenic assays. Results represent the averages of 3 independent experiments (± S.E) and data were analyzed by pairwise comparisons. Treatment groups with statistically significant differences are indicated by brackets (* = p<0.05). Differences in survival in response to IR between untreated (no curcumin) shNT and shTR1 cells were analyzed by ANOVA and were found to be statistically significant at 6 Gy (p<0.05) (see results text for details).
Table 1. Summary of Dose Enhancement Ratios (DERs) in TxnRd1 knockdown and overexpressing HEK293 cells.
DERs (expressed as the ratio of radiation dose required to produce 0.37% survival with no curcumin present to the dose required to produce the same survival) in presence of 10µM curcumin). N represents the number of times the experiment was replicated. shNT: the control, non-targeting vector; shTR1: TxnRd1 knockdown clones; pIRES: empty expression vector; TxnRd1: TxnRd1 expressing vector.
| Cell line | Plasmid (Clone) | (N) | DER (at 0.37 surv. fraction) |
|---|---|---|---|
| HeLa | shNT-5 | 3 | 1.32 ± 0.06 |
| shTR1-3.2 | 3 | 0.78 ± 0.18 | |
| shTR1-3.3 | 2 | 0.98 | |
| shTR1-1.2 | 3 | 1.04 ± 0.07 | |
| shTR1-1.5 | 2 | 0.95 | |
| SiHa | shNT-2 | 2 | 1.92 |
| shTR1-1.4 | 2 | 1.50 | |
| FaDu | shNT-1 | 3 | 1.75 ± 0.28 |
| shTR1-1.1 | 3 | 1.10 ± 0.27 | |
| shTR1-2.1 | 3 | 1.04 ± 0.14 | |
| HEK293 | pIRES | 3 | 1.06 ± 0.08 |
| TxnRd1 | 3 | 1.52 ± 0.19 |
TxnRd1 knockdown leads to reduced ROS generation and ERK1/2 activation following curcumin and IR treatments
In our previous study (3), we demonstrated that pretreatment of tumor cells with curcumin led to enhanced production of IR-induced ROS resulting in sustained ERK1/2-MAP kinase activation. To test the effect of TxnRd1 knockdown on ROS generation following combined treatment with curcumin and IR, we used DCF-DA, a probe whose fluorescence is directly proportional to intracellular ROS levels (23). Treatment of HeLa shNT (control) and shTR1–3.2 (TxnRd1 knockdown) clones with curcumin did not significantly increase ROS levels (Fig 4A). A single dose of 4Gy induced an increase in ROS levels that was significantly higher in the shTR1 clones, which is consistent with the role of TxnRd1 as a ROS scavenger. Combined curcumin and IR treatment caused a substantial increase in ROS in the shNT clone but not in the shTR1–3.2 clone. Quantitation of 3 independent experiments demonstrated a similar trend (Fig. 4B). Similar results in terms of curcumin-mediated ROS changes were observed in a second HeLa clone, shTR1–1.2 (Sup Fig. 1). Interestingly, clone shTR1–3.2 which exhibited a larger increase in ROS induction upon IR treatment compared to shTR1–1.2, also showed a larger effect in terms of radiosensitization upon knockdown of TxnRd1. These results indicate that higher levels of TxnRd1 result in increased levels of ROS in response to the combined treatment of curcumin and IR.
Figure 4. Effect of TxnRd1 knockdown on ROS production and ERK1/2 activation.
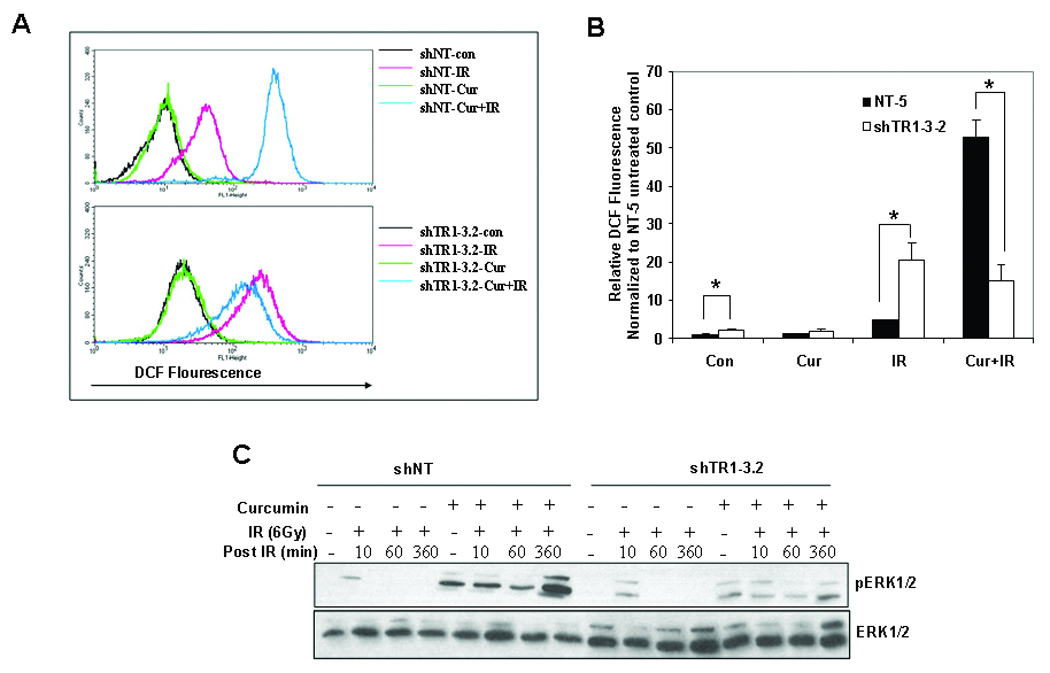
(A) Increase in ROS in HeLa-shNT or shTR1–3.2 clones was analyzed by flow cytometry using DCF-DA. At least 30,000 events were collected for each treatment condition. Results are representative of three independent experiments. (B) Results from (A) were quantified, represented as percentage increase in fluorescence normalized to shNT unirradiated control (±S.E.; *p<0.05). (C) ERK phosphorylation in response to curcumin treatment. shNT and shTR1-3.2 cells were serum-starved for 24 h. The cells were pretreated with DMSO or 10 µM curcumin for 8 h followed by mock irradiation or irradiation with 6 Gy. Whole cell lysates were analyzed by immunoblot using antibodies against p-ERK1/2 and total-ERK1/2. The results are representative of 3 independent experiments.
Our previous study (3) indicated that increased ROS production following treatment with curcumin and IR led to sustained ERK1/2 activation. To determine the effects of TxnRd1 knockdown on ERK1/2 activation, we performed immunoblot analysis on irradiated HeLa-shNT and HeLa-shTR1 cells that were either pretreated with curcumin or DMSO. As shown in Fig 4C, irradiated cells showed a transient increase in phosphoERK1/2 levels that decrease to baseline within an hour after irradiation. Pretreatment with curcumin prolonged IR-induced ERK1/2 activation in the HeLa-shNT cells for up to 6 h after IR. This sustained ERK1/2 activation was not observed in the HeLa-shTR1 cells after curcumin treatment. These results demonstrate that the increased production of ROS and the sustained ERK1/2 activation (which we previously demonstrated is required for curcumin-induced radiosensitization) are dependent on the levels of TxnRd1 protein.
Overexpression of TxnRd1 increases radiosensitivity to curcumin
To determine if overexpression of TxnRd1 is sufficient to cause curcumin-mediated radiosensitization, we attempted to overexpress TxnRd1 in keratinocytes and the minimally transformed MSK-Leuk1 cells. However, the intricate mammalian selenoprotein synthesis machinery imposes a major obstacle in constructing stable selenoprotein-expressing mammalian cell lines. Specifically, previous studies demonstrated that TxnRd1 expression is tightly regulated by multiple AU-rich sequences in the 3’-UTR (24, 25). Indeed, our attempts to overexpress TxnRd1 in either keratinocytes or MSK-Leuk1 cells were unsuccessful.
To overcome this problem, we searched for cell lines with low basal TxnRd1 levels which could allow for TxnRd1 overexpression. One such line reported in the literature is the HEK293 cell line (24). We obtained the HEK-293 cells stably expressing the full-length TxnRd1 gene (HEK293-TxnRd1) and also HEK-293 cells expressing the empty pIRES vector (HEK293-pIRES). Immunoblot analysis confirmed the overexpression of TxnRd1 protein in HEK293–TxnRd1 cells (Fig. 5A). Moreover, TxnRd1 activity was 6-fold higher in HEK293-TxnRd1 cells compared to HEK293-pIRES cells, indicating that the expressed protein was enzymatically active (Fig 5B). Also as shown in Fig 5C, HEK293-TxnRd1 cells exhibited significantly reduced survival in response to curcumin compared to the HEK293-pIRES cells, while THC did not result in significant cytotoxicity in either of the HEK293 cell lines confirming the role of TxnRd1 in determining curcumin mediated cytotoxicity.
Figure 5. TxnRd1 overexpression enhances curcumin-induced toxicity and radiosensitivity in HEK293 cells.
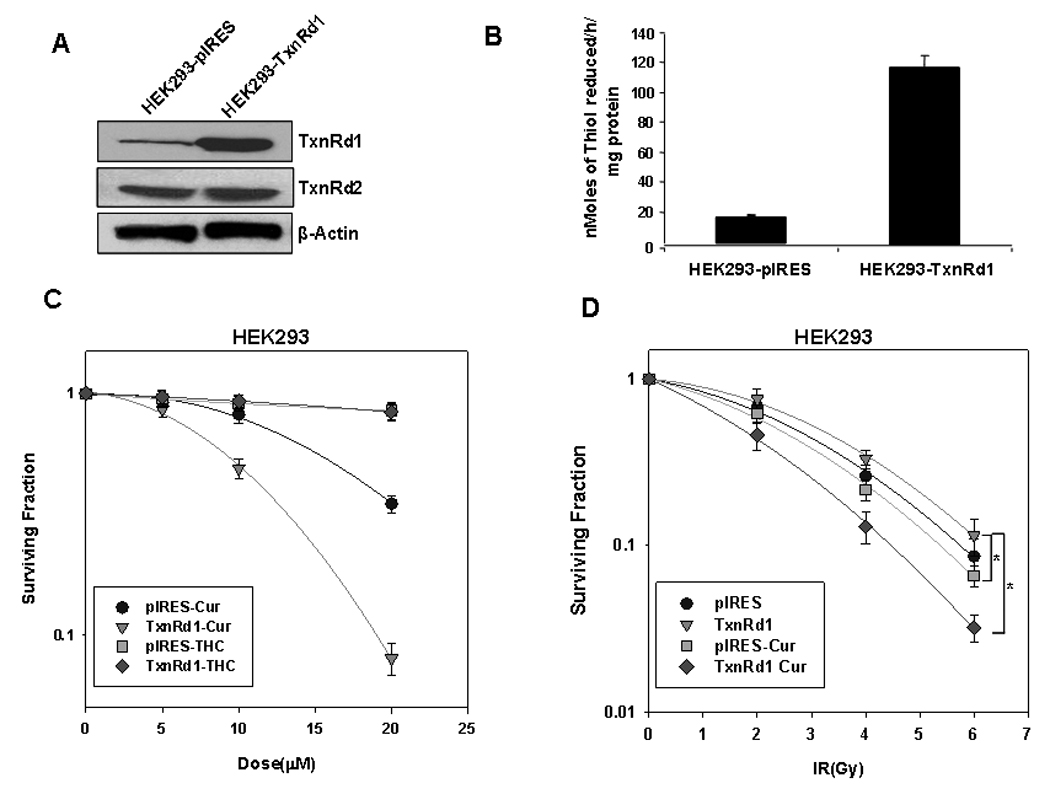
(A) Immunoblot analysis of HEK293 cells stably expressing an empty pIRES-neo vector (HEK293-pIRES) or the pIRES vector containing the full length TxnRd1 gene (HEK293-TxnRd1). β-actin was used as a loading control. (B) Basal TxnRd activity in HEK293-pIRES and HEK293-TxnRd1 cells. Results represent the average of 3 independent experiments (±S.E). (C) Cytotoxic effect of curcumin and THC on HEK293-pIRES and HEK293-TxnRd1 cells. Cells were treated with increasing concentrations of curcumin or THC for 8 h. When the untreated cells were confluent, MTT assays were performed. Results represent the averages of 4 independent experiments (±S.E). (D) Effect of curcumin on radiosensitivity of HEK293-pIRES and HEK293-TxnRd1 cells. Cells were treated with DMSO or 10 µM curcumin for 8 h followed by 0, 2, 4 or 6 Gy doses of IR. Long-term survival was assessed by high-density clonogenic survival assay. Results represent the averages of 3 independent experiments (± S.E) and data were analyzed by pairwise comparisons. Treatment groups with statistically significant differences are indicated by brackets (* = p<0.05).
Using a dose of 10 µM curcumin, we performed clonogenic survival assays on irradiated HEK293-pIRES and HEK293-TxnRd1 cells. As predicted from the work of Smart et al. (11), HEK293-TxnRd1 cells were more radioresistant than HEK 293-pIRES cells. Pretreating HEK293-pIRES cells with curcumin did not induce significant radiosensitization. In contrast, the HEK293-TxnRd1 cells were significantly radiosensitized by 10 µM curcumin, with a DER of 1.52 at 0.37 survival fraction (Fig 5D). Taken together, these results indicate that TxnRd1 confers increased radioresistance and that TxnRd1 overexpressing cells exhibit enhanced sensitivity to combined treatment with radiation and curcumin, supporting the role for TxnRd1 as a crucial target mediating curcumin-induced radiosensitization.
Discussion
Several reports have identified curcumin as a potent protector of normal tissue against radiation-induced damage. Administration of curcumin significantly reduced various normal tissue toxicities in rodent models treated with whole-body radiation (6, 26, 27). Intriguingly, curcumin has also been shown to radiosensitize various tumor cell lines in vitro (3–5) and induce a pronounced tumor growth delay following irradiation in mouse tumor models (28, 29). These findings indicate that curcumin has the ability to preferentially radiosensitize tumor but not normal tissue, a remarkable property for a radiation response modifier that could translate into substantial clinical benefit. However, the mechanism behind this selective property of curcumin has remained elusive and potential targets including NF-κB, Akt etc (4, 30, 31) have not been rigorously tested as causative factors in this effect.
Growing evidence suggests that some cancer cells produce higher basal levels of ROS than normal cells (32). Under this persistent intrinsic oxidative stress, cancer cells develop an enhanced, endogenous antioxidant capacity which makes them more resistant to exogenous oxidants (33, 34). The upregulation of the antioxidant enzyme TxnRd1 is observed in multiple primary human malignancies and its loss has been associated with a reversal of tumor phenotype and a decrease in tumorigenicity (35). These observations support the speculation that some malignant cells could be sensitized to oxidants including IR by inhibition of this key antioxidant protein (11, 36).
It has been hypothesized that TxnRd1 may be necessary to counteract IR-induced changes in intracellular protein thiol oxidation/reduction status and to directly scavenge cytotoxic free radicals formed during exposure to IR. This hypothesis was tested by Smart et al (11) using cell lines overexpressing wild-type or dominant negative (dn) form of TxnRd1 (cysteine mutant). This study demonstrated that HeLa cells overexpressing the wild-type but not dnTxnRd1 were more resistant to the lethal effects of IR, suggesting that TxnRd1 is a clinically relevant target for novel radiosensitizing agents. Moreover, in a separate study, Motexafin gadolinium, a potent inhibitor of TxnRd, has been shown to enhance the tumor cell response to IR and currently is in phase I clinical trials for patients with brain metastases from lung and breast cancers (14). Our results showing a significant increase in the radiosensitivity of HeLa and FaDu cells with knockdown TxnRd1 levels are in agreement with these data and further support a role of TxnRd1 as a major determinant of intrinsic tumor cell radiosensitivity.
Our attempts to overexpress TxnRd1 in non-transformed primary keratinocytes that normally express low levels of the protein were not successful, even when using a plasmid that had been used to express TxnRd1 in HEK293 cells (24). However, this was not unexpected since previous reports demonstrated major obstacles exist to overexpressing TxnRd1 in mammalian cells (24, 25). TxnRd1 expression is regulated by sequences in the 3’-UTR including AU-rich elements (ARE) that exert stringent regulatory control over translation of TxnRd1 mRNA. ARE-binding proteins and microRNAs that selectively recognize AU-rich sequences can affect stability of TxnRd1 mRNA levels, and protein expression (40). One or more of these factors may contribute to the differential expression of TxnRd1 in tumor and normal cells. This is an area of ongoing investigation in our laboratory.
Based on the findings presented here, we propose that the preferential radiosensitization of tumor cells by curcumin is, in part, due to the elevated expression of the TxnRd1. The involvement of TxnRd1 in curcumin-mediated radiosensitization appears to be two fold: First, curcumin covalently binds to TxnRd1 irreversibly inhibiting its ability to reduce thioredoxin, which in turn is required for many of the pleiotropic anti-oxidant effects associated with TxnRd1 (16). Second, modification of the selenocysteine residue of TxnRd1 converts the protein from essential antioxidant enzyme to a protein with NADPH oxidase activity, thereby elevating oxidative stress. Both effects would enhance the oxidative burden on the cell, leading to greater clonogenic cell death (a finding supported by our previous work demonstrating ablation of curcumin-mediated radiosensitization by the antioxidant NAC (3)). Normal cells which express very low levels of TxnRd1 are not radiosensitized by curcumin. While this phenomenon could be explained by low levels of TxnRd1, we have not excluded other possible mechanisms of radioprotection. For example, at least in vivo, it has been postulated that the radioprotective effects of curcumin could be due its inhibitory effects on the expression of inflammatory and fibrogenic cytokines (6).
Our findings further suggest that TxnRd1 may mediate radiosensitization by additional natural and chemical compounds. Curcumin reacts as a potent Michael acceptor due to the presence of α, β-unsaturated ketone moieties coupled with electron withdrawing phenolic hydroxyl groups. Curcumin-induced inhibition of TxnRd1 is dependent on the Michael acceptor function by which curcumin binds covalently to the highly nucleophilic selenocysteine residue in the C-terminus of TxnRd1 (16, 41). THC, which lacks the α,β-unsaturated ketone moieties, does not inhibit TxnRd1 activity and does not radiosensitize tumor cells. These observations suggest that compounds with Michael acceptor functionality could represent a novel family of radiosensitizers that are selective for transformed cells. Moreover, direct inhibition of TxnRd1 activity could be a useful means for screening natural and synthetic compounds for Michael acceptor functionality. Notably, caffeic acid phenethyl ester and oleocanthal acid, two additional natural compounds with Michael acceptor functions, are also inhibitors of TxnRd1 as well as potent tumor radiosensitizers (Tuttle et al., unpublished observations).
In conclusion, our results support a critical role of TxnRd1 in curcumin-mediated radiosensitization. Analysis of TxnRd1 levels in the tumors of patients enrolled in curcumin clinical trials could provide a valuable tool in identifying patients that would respond favorably to combined use of curcumin and radiotherapy.
Supplementary Material
Acknowledgements
We thank Dr. Peter Sacks (Memorial Sloane Kettering Hospital) for his gift of MSK-Leuk1 cells, Dr. Giannis Spyrou for his gift of HEK293-pIRES, HEK293-TxnRd1 cells and pIRES TxnRd1 plasmid and Dr. Bert W. O'Malley Jr. for his gift of SCC-1 cells.
Grant Support: This study was supported by grants CA104922 (to C.K.) and CA137398 (to SWT) by the National Cancer Institute.
Abbreviations
- TxnRd
Thioredoxin reductase
- ROS
reactive oxygen species
- ERK
extracellular signal-regulated kinase
- MAPK
mitogen-activated protein kinase
- DMSO
dimethyl sulfoxide
- Gy
gray
- MTT
3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium
- PBS
phosphate-buffered saline
- DCF-DA
2',7'-dichlorofluorescein diacetate
- IR
ionizing radiation
- PARP
poly(ADP-ribose) polymerase.
References
- 1.Howells LM, Moiseeva EP, Neal CP, et al. Predicting the physiological relevance of in vitro cancer preventive activities of phytochemicals. Acta Pharmacol Sin. 2007;28:1274–1304. doi: 10.1111/j.1745-7254.2007.00690.x. [DOI] [PubMed] [Google Scholar]
- 2.Goel A, Kunnumakkara AB, Aggarwal BB. Curcumin as “Curecumin”: From kitchen to clinic. Biochem Pharmacol. 2008;75(4):787–809. doi: 10.1016/j.bcp.2007.08.016. [DOI] [PubMed] [Google Scholar]
- 3.Javvadi P, Segan AT, Tuttle SW, Koumenis C. The chemopreventive agent curcumin is a potent radiosensitizer of human cervical tumor cells via increased reactive oxygen species production and overactivation of the mitogen-activated protein kinase pathway. Mol Pharmacol. 2008;73(5):1491–1501. doi: 10.1124/mol.107.043554. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Chendil D, Ranga RS, Meigooni D, Sathishkumar S, Ahmed MM. Curcumin confers radiosensitizing effect in prostate cancer cell line PC-3. Oncogene. 2004;23:1599–1607. doi: 10.1038/sj.onc.1207284. [DOI] [PubMed] [Google Scholar]
- 5.Khafif A, Hurst R, Kyker K, Fliss DM, Gil Z, Medina JE. Curcumin: A new radio-sensitizer of squamous cell carcinoma cells. Otolaryngol Head Neck Surg. 2005;132(2):317–321. doi: 10.1016/j.otohns.2004.09.006. [DOI] [PubMed] [Google Scholar]
- 6.Okunieff P, Xu J, Hu D, et al. Curcumin protects against radiation-induced acute and chronic cutaneous toxicity in mice and decreases mRNA expression of inflammatory and fibrogenic cytokines. Int J of Radiat Oncol Biol Phys. 2006;65(3):890–898. doi: 10.1016/j.ijrobp.2006.03.025. [DOI] [PubMed] [Google Scholar]
- 7.Choudhary D, Chandra D, Kale RK. Modulation of radioresponse of glyoxalase system by curcumin. J Ethnopharmacol. 1999;64(1):1–7. doi: 10.1016/s0378-8741(98)00064-6. [DOI] [PubMed] [Google Scholar]
- 8.Mustacich DPG. Thioredoxin reductase. Biochem J. 2000;346(1):1–8. [PMC free article] [PubMed] [Google Scholar]
- 9.Becker K, Gromer S, Schirmer RH, Müller S. Thioredoxin reductase as a pathophysiological factor and drug target. European Journal of Biochemistry. 2000;267(20):6118–6125. doi: 10.1046/j.1432-1327.2000.01703.x. [DOI] [PubMed] [Google Scholar]
- 10.Karimpour S, Lou J, Lin LL, et al. Thioredoxin reductase regulates AP-1 activity as well as thioredoxin nuclear localization via active cysteines in response to ionizing radiation. Oncogene. 2002;21(41):6317–6327. doi: 10.1038/sj.onc.1205749. [DOI] [PubMed] [Google Scholar]
- 11.Smart DK, Ortiz KL, Mattson D, et al. Thioredoxin reductase as a potential molecular target for anticancer agents that induce oxidative stress. Cancer Res. 2004;64(18):6716–6724. doi: 10.1158/0008-5472.CAN-03-3990. [DOI] [PubMed] [Google Scholar]
- 12.Choi JH, Kim TN, Kim S, et al. Overexpression of mitochondrial thioredoxin reductase and peroxiredoxin III in hepatocellular carcinomas. Anticancer Res. 2002;22(6A):3331–3335. [PubMed] [Google Scholar]
- 13.Lincoln DT, Ali Emadi EM, Tonissen KF, Clarke FM. The thioredoxin-thioredoxin reductase system: Over-expression in human cancer. Anticancer Res. 2003;23(3B):2425–2433. [PubMed] [Google Scholar]
- 14.Carde P, Timmerman R, Mehta MP, et al. Multicenter phase Ib/II trial of the radiation enhancer motexafin gadolinium in patients with brain metastases. J Clin Oncol. 2001;19(7):2074–2083. doi: 10.1200/JCO.2001.19.7.2074. [DOI] [PubMed] [Google Scholar]
- 15.Urig SBK. On the potential of thioredoxin reductase inhibitors for cancer therapy. Seminars in cancer biology. 2006;16(6):452–465. doi: 10.1016/j.semcancer.2006.09.004. [DOI] [PubMed] [Google Scholar]
- 16.Fang J, Lu J, Holmgren A. Thioredoxin reductase is irreversibly modified by curcumin: A novel molecular mechanism for its anticancer activity. J Biol Chem. 2005;280(26):25284–25290. doi: 10.1074/jbc.M414645200. [DOI] [PubMed] [Google Scholar]
- 17.Tamura T, Stadtman TC. A new selenoprotein from human lung adenocarcinoma cells: Purification, properties, and thioredoxin reductase activity. Proc Natl Acad Sci U S A. 1996;93(3):1006–1011. doi: 10.1073/pnas.93.3.1006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Arnér ESJ, Nordberg J, Holmgren A. Efficient reduction of lipoamide and lipoic acid by mammalian thioredoxin reductase. Biochem Biophys Res Commun. 1996;225(1):268–274. doi: 10.1006/bbrc.1996.1165. [DOI] [PubMed] [Google Scholar]
- 19.Biaglow JE, Donahue J, Tuttle S, Held K, Chrestensen C, Mieyal J. A method for measuring disulfide reduction by cultured mammalian cells: Relative contributions of glutathione-dependent and glutathione-independent mechanisms. Anal Biochem. 2000;281(1):77–86. doi: 10.1006/abio.2000.4533. [DOI] [PubMed] [Google Scholar]
- 20.Somparn P, Phisalaphong C, Nakornchai, Unchern S, Morales NP. Comparative antioxidant activities of curcumin and its demethoxy and hydrogenated derivatives. Biol Pharm Bull. 2007;30(1):74–78. doi: 10.1248/bpb.30.74. [DOI] [PubMed] [Google Scholar]
- 21.Syng-ai C, Kumari AL, Khar A. Effect of curcumin on normal and tumor cells: Role of glutathione and bcl-2. Mol Cancer Ther. 2004;3(9):1101–1108. [PubMed] [Google Scholar]
- 22.Koumenis C, Naczk iC, Koritzinsky M, et al. Regulation of protein synthesis by hypoxia via activation of the endoplasmic reticulum kinase PERK and phosphorylation of the translation initiation factor eIF2a. Mol Cell Biol. 2002;22:7405–7416. doi: 10.1128/MCB.22.21.7405-7416.2002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Brandt R, Keston AS. Synthesis of dichlorodihydrofluorescin: a stable reagent for fluorometric analysis. Anal Biochem. 1965;11:6–9. doi: 10.1016/0003-2697(65)90035-7. [DOI] [PubMed] [Google Scholar]
- 24.Nalvarte I, Damdimopoulos AE, Nystom C, et al. Overexpression of enzymatically active human cytosolic and mitochondrial thioredoxin reductase in HEK-293 cells: Effect on cell growth and differentiation. J Biol Chem. 2004;279(52):54510–54517. doi: 10.1074/jbc.M408494200. [DOI] [PubMed] [Google Scholar]
- 25.Gasdaska JR, Harney JW, Gasdaska PY, Powis G, Berry MJ. Regulation of human thioredoxin reductase expression and activity by 3'-untranslated region selenocysteine insertion sequence and mRNA instability elements. J Biol Chem. 1999;274(36):25379–25385. doi: 10.1074/jbc.274.36.25379. [DOI] [PubMed] [Google Scholar]
- 26.Inano H, Onoda M. Radioprotective action of curcumin extracted from curcuma longa LINN: Inhibitory effect on formation of urinary 8-hydroxy-2′-deoxyguanosine, tumorigenesis, but not mortality, induced by γ-ray irradiation. Int J Radiat Oncol Biol Phys. 2002;53(3):735–743. doi: 10.1016/s0360-3016(02)02794-3. [DOI] [PubMed] [Google Scholar]
- 27.Thresiamma KC, George J, Kuttan R. Protective effect of curcumin, ellagic acid and bixin on radiation induced toxicity. Indian J Exp Biol. 1996;34(9):845–847. [PubMed] [Google Scholar]
- 28.Kunnumakkara AB, Diagaradjane P, Guha S, et al. Curcumin sensitizes human colorectal cancer xenografts in nude mice to {gamma}-radiation by targeting nuclear factor-{kappa}B-regulated gene products. Clin Cancer Res. 2008;14(7):2128–2136. doi: 10.1158/1078-0432.CCR-07-4722. [DOI] [PubMed] [Google Scholar]
- 29.Li M, Zhang Z, Hill DL, Wang H, Zhang R. Curcumin, a dietary component, has anticancer, chemosensitization, and radiosensitization effects by down-regulating the MDM2 oncogene through the PI3K/mTOR/ETS2 pathway. Cancer Res. 2007;67(5):1988–1996. doi: 10.1158/0008-5472.CAN-06-3066. [DOI] [PubMed] [Google Scholar]
- 30.Kunnumakkara AB, Diagaradjane P, Guha S, et al. Curcumin Sensitizes Human Colorectal Cancer Xenografts in Nude Mice to {gamma}-Radiation by Targeting Nuclear Factor-{kappa} B-Regulated Gene Products. Clin Cancer Res. 2008;14(7):2128–2136. doi: 10.1158/1078-0432.CCR-07-4722. [DOI] [PubMed] [Google Scholar]
- 31.Dhandapani KM, Mahesh VB, Brann DW. Curcumin suppresses growth and chemoresistance of human glioblastoma cells via AP-1 and NFkappaB transcription factors. J Neurochem. 2007;102(2):522–538. doi: 10.1111/j.1471-4159.2007.04633.x. [DOI] [PubMed] [Google Scholar]
- 32.Toyokuni S, Okamoto K, Yodoi J, Hiai H. Persistent oxidative stress in cancer. FEBS Lett. 1995;358:1–3. doi: 10.1016/0014-5793(94)01368-b. [DOI] [PubMed] [Google Scholar]
- 33.Pervaiz S, Clement MV. Tumor intracellular redox status and drug resistance — serendipity or a causal relationship? Curr Pharm Des. 2004;10:1969–1977. doi: 10.2174/1381612043384411. [DOI] [PubMed] [Google Scholar]
- 34.Tiligada E. Chemotherapy: induction of stress responses. Endocr Relat Cancer. 2006;13 Suppl. 1:S115–S124. doi: 10.1677/erc.1.01272. [DOI] [PubMed] [Google Scholar]
- 35.Yoo M, Xu X, Carlson BA, Gladyshev VN, Hatfield DL. Thioredoxin reductase 1 deficiency reverses tumor phenotype and tumorigenicity of lung carcinoma cells. J Biol Chem. 2006;281(19):13005–13008. doi: 10.1074/jbc.C600012200. [DOI] [PubMed] [Google Scholar]
- 36.Kirkpatrick DL, Kuperus M, Dowdeswell M, et al. Mechanisms of inhibition of the thioredoxin growth factor system by antitumor 2-imidazolyl disulfides. Biochem Pharmacol. 1998;55(7):987–994. doi: 10.1016/s0006-2952(97)00597-2. [DOI] [PubMed] [Google Scholar]
- 37.Magda D, Lepp C, Gerasimchuk N, et al. Redox cycling by motexafin gadolinium enhances cellular response to ionizing radiation by forming reactive oxygen species. Int J of Radiat Oncol Biol Phys. 2001;51(4):1025–1036. doi: 10.1016/s0360-3016(01)01810-7. [DOI] [PubMed] [Google Scholar]
- 38.Rockwell S, Donnelly ET, Liu Y, Tang L. Preliminary studies of the effects of gadolinium texaphyrin on the growth and radiosensitivity of EMT6 cells in vitro. Int J of Radiat Oncol Biol Phys. 2002;54(2):536–541. doi: 10.1016/s0360-3016(02)02962-0. [DOI] [PubMed] [Google Scholar]
- 39.Gius D. Redox-sensitive signaling factors and antioxidants: How tumor cells respond to ionizing radiation. J Nutr. 2004;134(11):3213S–3214S. doi: 10.1093/jn/134.11.3213S. [DOI] [PubMed] [Google Scholar]
- 40.Von Roretz C, Gallouzi I. Decoding ARE-mediated decay: Is microRNA part of the equation? J Cell Biol. 2008;181(2):189–194. doi: 10.1083/jcb.200712054. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Moos PJ, Edes K, Mullally JE, Fitzpatrick FA. Curcumin impairs tumor suppressor p53 function in colon cancer cells. Carcinogenesis. 2004;25(9):1611–1617. doi: 10.1093/carcin/bgh163. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.