

Abstract
The larger of two diuretic hormones of the tobacco hornworm, Manduca sexta, (Mas-DH) is a peptide of 41 residues. It is one of a family of seven currently known insect diuretic hormones that are similar to the corticotropin-releasing factor–urotensin–sauvagine family of peptides. We investigated the possible inactivation of Mas-DH by incubating it in vitro with larval Malpighian tubules (Mt), the target organ of the hormone. The medium was analyzed, and degradation products were identified, using on-line microbore reversed-phase liquid chromatography coupled to electrospray ionization mass spectrometry (RPLC-ESI-MS). This sensitive technique allows identification of metabolites of Mas-DH (present at an initial level of ≈1 μM). An accurate Mr value for a metabolite is usually sufficient for unambiguous identification. Mas-DH is cleaved by Mt proteases initially at L29–R30 and R30–A31 under our assay conditions; some Mas-DH is also oxidized, apparently at M2 and M11. The proteolysis can be inhibited by 5 mM EDTA, suggesting that divalent metals are needed for peptide cleavage. The oxidation of the hormone can be inhibited by catalase or 1 mM methionine, indicating that H2O2 or related reactive oxygen species are responsible for the oxidative degradation observed. RPLC-ESI-MS is shown here to be an elegant and efficient method for studying peptide hormone metabolism resulting from unknown proteases and pathways.
Keywords: proteolysis, endopeptidase, oxidation, lepidoptera, inactivation
Water balance and ion balance in insects are regulated by the actions of diuretic hormones and antidiuretic hormones. The larger of two diuretic hormones (DH) of the tobacco hornworm Manduca sexta (Mas-DH), RMPSLSIDLPMSVLRQKLSLEKERKVHALRAAANRNFLNDI-NH2, is an amidated peptide with 41 residues (1). It was the first peptide identified of a family of insect diuretic peptides related to corticotropin-releasing factor (CRF), sauvagine, and urotensin I (2, 3). Subsequently, a 30-amino acid DH homologous to Mas-DH was identified from the same species (4). Mas-DH activates urine production in M. sexta Malpighian tubules (Mt) by stimulating cAMP production and, eventually, KCl and NaCl cotransport, causing osmotically driven fluid excretion from hemolymph into the lumen of Mt (5, 6). A G-protein-coupled Mas-DH receptor with seven putative transmembrane domains has been cloned from M. sexta Mt and functionally expressed (7, 8). Termination of Mas-DH-activated diuresis is believed to be accomplished by the cessation of Mas-DH production coupled with its rapid removal from hemolymph by unknown mechanisms.
Here we report on the metabolism of Mas-DH by M. sexta Mt after incubation in vitro. Mt were chosen because they are the putative target organ of Mas-DH, and they have been found to decrease diuretic activities of crude tissue extracts in four insect species (9). In addition, Mt have been shown to degrade some other insect neuropeptides (10–13). Peptide hormone degradation products have been routinely analyzed by using separation by reversed-phase liquid chromatography (RPLC) and identified by comparing retention properties of the products to those of a number of synthetic peptide standards representing possible metabolites. The products have been detected by UV absorbance (14), by ELISA or RIA (15), by radioactivity (12, 16), from fluorescently labeled or chromogen-modified peptides (15), by amino acid analysis (AAA) of fractions (17), or by Edman degradation (18). We used RPLC coupled to electrospray ionization (ESI)-MS to identify metabolites of Mas-DH, a technique that can detect unlabeled, nonmodified peptide and its metabolites at low concentrations. RPLC-ESI-MS has become a powerful method for studying peptide metabolism, owing to the high sensitivity offered by soft ionization techniques and capillary RPLC systems (19–21). Mas-DH was incubated with Mt in vitro at initial levels of 40 or 1 μM. Metabolite identification was based on agreement of the Mr of metabolites with the calculated Mr. We found that Mas-DH can be metabolized via two overlapping pathways, proteolysis and oxidation.
MATERIALS AND METHODS
Insects.
The M. sexta colony was reared essentially as described by Yamamoto (22). The Mt were dissected from day 2 or 3 fifth-instar larvae. The insects were chilled on ice for 15 min before dissection.
Chemicals.
Synthetic Mas-DH was donated by Sandoz Agro. EDTA, malic acid, salts, and amino acids were from Sigma. Sucrose, glucose, and trifluoroacetic acid (TFA) were purchased from Fisher. HPLC grade CH3CN was from EM Science. Water was purified with a Milli-Q apparatus from Millipore. Human erythrocyte catalase (160,000 units/mg) was from ICN.
In Vitro Incubation.
The ascending portions of Mt were carefully dissected from M. sexta larvae bathed in M. sexta saline (5). Mt were separated from the fat body, tracheae, muscle, and attached gut tissue. After preincubation in saline at 30°C for 1 h, a Mt fragment (≈1 cm) was transferred into a 0.5-ml microcentrifuge tube containing 100 μl of saline with 1 μM Mas-DH (100 pmol). The incubations were performed at 30°C with the tubes capped and were terminated by removing the Mt and then adding 100 μl of TFA/CH3CN (2:98, vol/vol) to the medium. The samples were frozen in liquid N2 and stored at −20°C until assayed by RPLC-ESI-MS. In separate experiments, 5 mM EDTA, 25 units of catalase, or 1 mM methionine was also included. Incubations were for 1 h except as specified. Two controls were used for each experiment: Mas-DH in saline without Mt, and Mt in saline without Mas-DH. When the Mt luminal fluid was used as metabolizing agent, it was collected by cutting one end of a tubule, lifting the other end, and removing the murky yellowish-white fluid [containing crystals presumed to be uric acid (23)]. Only 2 μl of the fluid diluted in saline was used for each in vitro incubation. When a cleansed ascending Mt fragment (≈1 cm) was used as protease source, the luminal fluid was removed as described, and the Mt were rinsed five times in saline prior to preincubation. In an experiment studying Mas-DH degradation at 40 μM, a single Mt (≈10 cm, cut at the junction with the ampulla and the entry to the ileac plexus) was used; great care was taken not to damage the Mt during dissection. After a 1-h preincubation, the Mt were incubated in 50 μl of saline containing 40 μM Mas-DH for 2 h.
The half-life value (t1/2) for Mas-DH at 1 μM was derived from peak area values for Mas-DH determined in chromatographic analyses at 1-, 55-, 60-, 90-, and 120-min incubation times (three replicates per time point), analyzed with ez chrom 6.0 software (Scientific Software, Pleasanton, CA). The t1/2 calculated is based on the assumption that Mas-DH disappears from the medium as a first-order process.
Liquid Chromatography.
A Model UMA chromatograph with built-in UV detector set at 220 nm, the liquid chromatography columns, and the peptide trap cartridge were from Michrom BioResources (Auburn, CA). The solvents were 2% CH3CN in water with 0.1% TFA (A) and 90% CH3CN in water with 0.085% TFA (B) (all vol/vol measurements). A 0.5 × 150 mm (5 μm, 300 Å) Reliasil C18 column was eluted at 20 μl/min, with column temperature set at 50°C. Samples were diluted with 5 vol of solvent A, loaded onto a 1 × 10 mm peptide trap cartridge (Michrom), and rinsed with 1 ml of solvent A before beginning the gradient. The 30-min gradient was composed of three linear segments from 20% to 40% B in 7 min, 40–45% B in 15 min, and 45–65% B in 8 min. The chromatograph was controlled with ez chrom 6.0 software.
MS.
A Finnigan MAT SSQ 710 mass spectrometer interfaced with an ESI ion source (needle 60 μm inside diameter, from Analytica, Branford, CT) was used to determine the Mr of Mas-DH and its metabolites. Samples were introduced into the ESI source after splitting the column effluent with a 60 μm inside diameter polyaryletherketone (PEEK) tube coupled to a homemade PEEK splitter with ≈50:1 (waste/MS) split ratio. Other ESI conditions were as described previously (2), except that the cation (m/z) scan range was from 400 to 1,400 Da. Spectra were acquired every 1.5 sec. The optimal ESI needle voltage was −2 to −3 kV.
Metabolite Identification.
The Mr values of all possible metabolites of Mas-DH resulting from cleavage of a single peptide bond were calculated by using MacVector (International Biotechnologies); the measured Mr of Mas-DH and its metabolites was derived with the BioDeconvolution and BioMass Calculation algorithm (Finnigan). We also used our own software (written by H.W.) to visualize and align the envelope ion peaks of all metabolites eluted from liquid chromatography. The identification of a Mas-DH metabolite is based on matching the observed and calculated Mr. Each peak in the UV chromatogram was characterized both by its retention time and from the corresponding relative ion chromatogram (RIC).
AAA and Edman Sequencing.
Some purified Mas-DH metabolites were further identified by AAA. Metabolites were hydrolyzed to amino acids using 5.7 M HCl (Pierce) plus 1% phenol at 110°C for 20 h. Amino acids were analyzed as 6-aminoquinoline carbamate derivatives (24) by using a Hewlett–Packard 1090 chromatograph with a Hewlett–Packard 1046A fluorescence detector. When quantities sufficed, some purified metabolites were sequenced with a Porton Instruments (Tarzana, CA) PI 2090 gas-phase sequencer with an integral phenylthiohydantoin amino acid analyzer.
RESULTS
Degradation of 40 μM Mas-DH.
In an initial study, we investigated metabolism of Mas-DH by uncleansed Mt at the nonphysiological concentration of 40 μM. A very complex metabolic profile was seen (summarized in Fig. 1). Mas-DH was cleaved at several different peptide bonds and oxidized. Four major proteolytic metabolites were seen; they were identified by ESI-MS as Mas-DH-(1–29), -(30–41), -(1–30), and -(31–41). These suggest that the initial endoproteolytic sites are at L29–R30 and R30–A31. We also saw evidence at this concentration for another major endoproteolytic site at S6–I7 and eight minor endopeptidase sites, each with the corresponding fragments of the opposite portion of the peptide (Fig. 1). Such observation of both possible fragments from a single peptide bond is strongly supportive of the existence of specific endopeptidase activity. For other fragments detected, the corresponding piece was not found. The latter type of fragments occurred as a series of products having successively one less residue at their N or C terminus. They are most likely generated by the actions of nonspecific amino- or carboxypeptidases. Only the N terminus of intact Mas-DH and of certain endoproteolytic metabolites appeared to be susceptible to aminopeptidase activity. No fragments corresponding to carboxypeptidase-type enzyme activity on the blocked C terminus of Mas-DH were observed, but carboxypeptidase-like activity seemed to affect the C terminus of several endoproteolytic metabolites (Fig. 1). Some of these cleavages at this high Mas-DH concentration (40 μM) are likely due to proteases with low specificity for Mas-DH.
Figure 1.
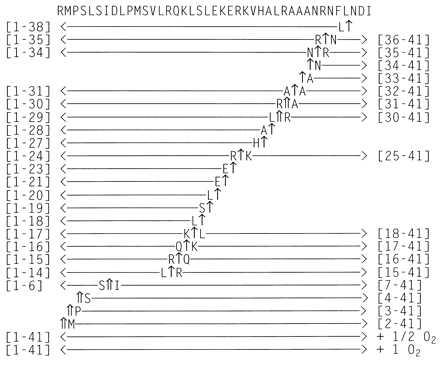
Scheme illustrating the sequences of the metabolites of Mas-DH identified from incubation with uncleansed Mt at 40 μM. Chromatographic conditions and methods of identification are described in the text. Major cleavage sites are indicated by ⇑ and minor cleavage sites are indicated by ↑.
Degradation of 1 μM Mas-DH.
Physiological concentrations of peptide hormones are usually in the order of nanomolar; we therefore lowered the MasDH concentration in the incubation. Because of the high sensitivity of RPLC-ESI-MS, we were able to study peptide metabolism at 1 μM initial concentration of unlabeled material, but only upon changing from a 1.0-mm diameter column to a 0.5-mm diameter column. The degradation of Mas-DH by Mt is relatively fast. When 100 pmol of Mas-DH in 100 μl of medium was incubated with a 1-cm piece of a freshly dissected Mt, the level of intact Mas-DH dropped to less than 30% in 60 min and to ≈5% in 90 min (Fig. 2). The t1/2 calculated for intact Mas-DH in this system is ≈22 min. As determined by comparing the calculated and measured Mr, the major metabolites observed were Mas-DH-(1–29) and -(1–30), Mas-DH plus two O atoms, and two different isomers of Mas-DH plus one O atom (Figs. 2 and 3).
Figure 2.
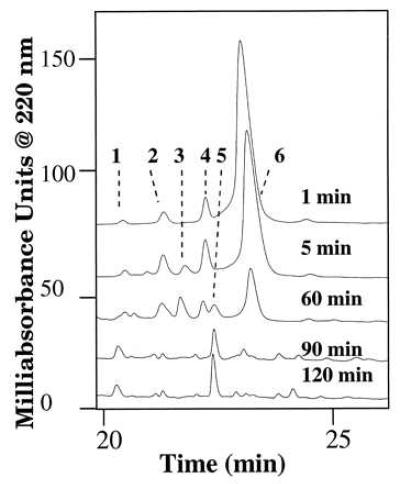
RPLC separation of degradation products of Mas-DH at initial concentration of 1 μM incubated with 1-cm pieces of Mt for 1, 5, 60, 90, or 120 min. Chromatographic conditions and identification of peaks are described in the text. Peak identities: 1, Mas-DH + 2O; 2 and 4, Mas-DH + O; 3, Mas-DH-(1–30); 5, Mas-DH-(1–29); and 6, Mas-DH. We observed a run-to-run variation in retention times, likely because of the 20 μl/min flow rate used with the 0.5-mm column. The Michrom UMA is designed for accurate delivery at flow rates of 50 μl/min and higher.
Figure 3.
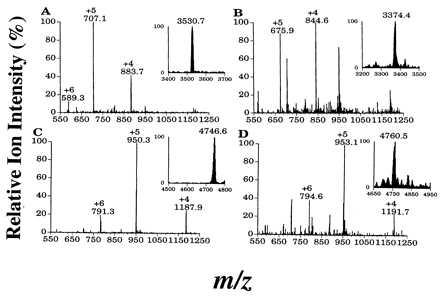
ESI mass spectra of major Mas-DH metabolites. Mass spectral conditions are described in the text. Insets show the deconvoluted spectra of the metabolites. (A) Mas-DH-(1–30). (B) Mas-DH-(1–29). (C) Mas-DH + O. (D) Mas-DH + 2O.
Two different Mas-DH + 16 atomic mass unit (amu) peaks and one Mas-DH+32 amu peak were identified (Figs. 2 and 3); we believe they represent mono- and bissulfoxides of the two Met (M2 and M11) residues. Small amounts of the sulfoxides were present in the saline control incubation without added Mt, due to the presence of traces of these as impurities in the stock of Mas-DH. We were unable to completely remove oxidized Mas-DH by RPLC purification, likely because of the notorious sensitivity of Met to oxidation in low-pH mobile phases from RPLC. However, incubation with Mt clearly increases the amount of oxidized Mas-DH. The amount of oxidized Mas-DH more than doubled between 1 and 5 min after the incubation was started (Fig. 2).
The first proteolytic product detected was Mas-DH-(1–30), which appeared 5 min after the beginning of the incubation, increased in abundance up to 60 min, then disappeared (Fig. 2). A second fragment, Mas-DH-(1–29), was detected after 1 h and was the main metabolite remaining after 2 h of incubation (Fig. 2). After a second RPLC purification of fractions containing each metabolite, they were analyzed by Edman sequencing; the results confirmed the MS identification (data not shown). None of the Mas-DH proteolytic products were detected in the control incubation of Mt, and only trace amounts of oxidation products were found in a control incubation of Mas-DH in saline. Fragments Mas-DH-(30–41) and Mas-DH-(31–41) were not detected, nor were any smaller fragments from the C-terminal portion of the peptide.
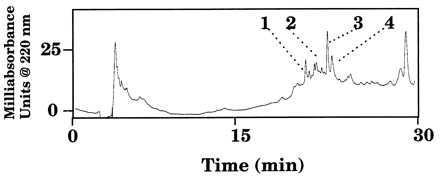
In the experiments described above, uncleansed Mt were used. The proteolytic or oxidative activities could be intrinsic to the tubules or could be components of the luminal fluid (urine) contained within the tubules. To further understand our findings, we studied metabolism of Mas-DH by Mt removed of their contents. Mt cleansed of the luminal fluid seemed to retain high proteolytic but low oxidative activity, as evidenced by the presence of a high yield of Mas-DH-(1–29) and a relatively low yield of other metabolites (Fig. 4). The luminal fluid from Mt seemed to contain all the factors needed to cleave and oxidize Mas-DH (Fig. 5). After incubating Mas-DH for 1 h with 2 μl of Mt luminal fluid diluted in 98 μl of saline, we detected the two C-terminal fragments of Mas-DH, Mas-DH-(31–41) and -(30–41) (identified by ESI-MS and AAA), along with all proteolytic and oxidative metabolites found with uncleansed Mt (Fig. 5). The discovery of these intact C-terminal fragments (and no others) verifies that the L29–R30 and R30–A31 sites are the primary Mas-DH sites cleaved by Mt and confirm that Mas-DH-(1–29) is not produced by degradation of Mas-DH-(1–30). In the same experiment, we also identified the previously undetected metabolites Mas-DH-(1–29)+O, -(1–30)+O, -(1–35), -(1–35)+O, and -(2–41). The discovery of oxidized proteolytic metabolites suggests that the oxidation and proteolysis are parallel processes. Oxidation occurs actively in luminal fluid even at this 50:1 dilution, but not in cleansed tubules.
Figure 4.
UV trace of RPLC analysis of 1 μM Mas-DH incubated with a cleansed Mt for 1 h. Peak identities: 1, unknown; 2, Mas-DH-(1–30); 3, Mas-DH-(1–29); and 4, Mas-DH.
Figure 5.
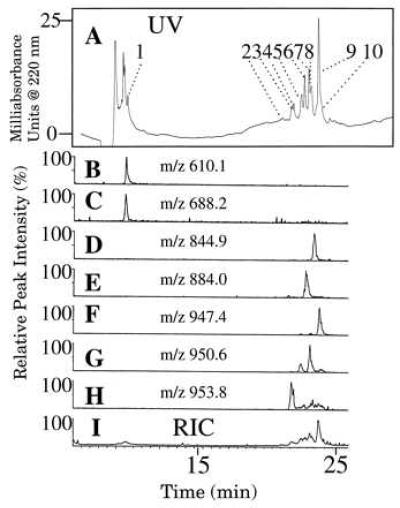
UV trace of RPLC analysis of 1 μM Mas-DH incubated with Mt luminal fluid for 1 h. Peak identities: 1, Mas-DH-(31–41) and Mas-DH-(30–41); 2, Mas-DH-(1–35) + O; 3, Mas-DH + 2O; 4, Mas-DH-(1–30) + O; 5, Mas-DH-(1–29) + O, Mas-DH + O, and Mas-DH-(1–35); 6, Mas-DH-(1–30); 7, Mas-DH + O; 8, Mas-DH-(1–29); 9, Mas-DH; and 10, Mas-DH-(2–41). (B–I) RIC. Single ions are plotted for the most abundant envelope ion of Mas-DH and its metabolites. The numbers of protons attached are indicated in the parentheses in the following: (B) Mas-DH-(31–41) (2H+). (C) Mas-DH-(30–41) (2H+). (D) Mas-DH-(1–29) (4H+). (E) Mas-DH-(1–30) (4H+). (F) Mas-DH (5H+). (G) Mas-DH + O (5H+). (H) Mas-DH + 2O (5H+). (I) Total RIC.
To demonstrate how we identified metabolites, we plotted reconstructed selected single-ion chromatograms for the most abundant ion of each metabolite produced by incubation with luminal fluid (Fig. 5 B–H); we have aligned the UV trace and the RIC traces (Fig. 5). These single-ion chromatograms allow measurement of subtle differences in the retention times of incompletely resolved peaks shown on the UV trace, thus enabling us to detect peptides that coelute or elute close together (e.g., peaks 1, 7, and 8 in Fig. 5A).
Inhibition of Proteolysis.
Because metalloproteases that require divalent cations for activity have been implicated in the degradation of other insect peptide hormones by Mt (10–12, 25), we studied the effect of a chelating agent, EDTA, on Mas-DH degradation. In a 2-h incubation with uncleansed Mt, 5 mM EDTA significantly inhibited proteolysis of Mas-DH, as evidenced by only a small quantity of the metabolite Mas-DH-(1–30) and complete absence of Mas-DH-(1–29) (Fig. 6). This result suggests that at least one of the Mt enzymes that degrades Mas-DH is a metalloprotease. Increased oxidation of Mas-DH occurred in the presence of EDTA (Fig. 6).
Figure 6.
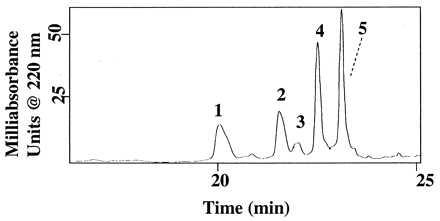
RPLC separation of metabolites from 2-h incubation of 1 μM Mas-DH with uncleansed Mt in the presence of 5 mM EDTA. Peak identities: 1, Mas-DH + 2O; 2 and 4, Mas-DH + O; 3, Mas-DH-(1–30); and 5, Mas-DH.
Inhibition of Oxidation.
Mas-DH oxidation by uncleansed Mt was inhibited by the inclusion of 25 units of catalase in the incubation (Fig. 7A), suggesting the involvement of H2O2. Addition of 1 mM methionine also prevented the oxidation of Mas-DH, because of the ability of methionine (present at 1,000-fold higher concentration than Mas-DH) to scavenge oxidant. This suggests that M2 and M11 were the targets of oxidation (Fig. 7B), because the peptide contains no Cys or Trp, only Met, as a readily oxidized residue. In both instances, the inhibition of Mas-DH oxidation favored the production of the proteolytic metabolite Mas-DH-(1–29) over -(1–30) (Fig. 7), although the latter is usually produced more rapidly (Fig. 2). This was also observed in the incubation with cleansed Mt (Fig. 4).
Figure 7.
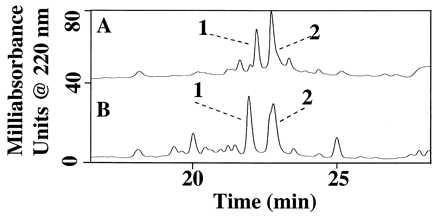
RPLC separation of metabolites from 1-h incubation of 1 μM Mas-DH with uncleansed Mt in the presence of 25 units of catalase (A) or 1 mM methionine (B). Peak identities: 1, Mas-DH-(1–29); and 2, Mas-DH.
DISCUSSION
Water and ion balance is crucial for insect survival. The function of Mt in insects is similar to that of the kidney in vertebrates. However, the mechanism of fluid transport is fundamentally different in Mt. The Mt actively transport salts and metabolites from hemolymph into the lumen, creating an osmotic gradient that drives primary urine excretion. For M. sexta and other terrestrial insects, diuresis provides a means to eliminate wastes, toxic metabolites, and water via the urine. Insects regulate fluid homeostasis with diuretic and antidiuretic hormones (9, 26). These chemical messengers should be relatively short-lived, so that hormonal stimulation of diuresis (or antidiuresis) occurs only to restore homeostasis. This is believed to be accomplished by the cessation of hormone production coupled with rapid hormone degradation. Early investigators showed evidence of DH-degrading activities present in hemolymph (27, 28) and in Mt of a few insect species (13, 29–31). In hemolymph diluted 1 to 50 with saline, we saw no significant degradation of Mas-DH at 1 μM after 2 h (data not shown). This is intuitively satisfying, because if hemolymph were to play a major role in degradation of DH, the hormone released from the neurohemal regions might have difficulty reaching its target receptors intact and biologically active. Another possibility for a degradation process is the internalization of Mas-DH bound to its receptor. This has not been investigated yet, in part because of the lack of radiolabeled Mas-DH. Mt, the target organ of DH, would seem a logical site for DH-degrading enzymes. Here we report a mechanism of DH degradation by Mt.
The proteolysis we observed of Mas-DH generates major peptide fragments Mas-DH-(1–29), -(1–30), -(30–41), and -(31–41) and their M2 or M11 oxidized derivatives. These metabolites are unlikely to be active for diuresis because they lack those structural components that other data (6, 8, 32) suggest are needed for the coordinated effects of receptor binding and activation. A previous study suggests that the N-terminal portion of Locusta migratoria DH (a molecule highly homologous to Mas-DH) is pivotal for receptor activation and the C-terminal portion for receptor binding (3). A receptor binding assay showed that Mas-DH-(13–41) had only 10-fold lower affinity for the receptor compared with Mas-DH, whereas Mas-DH-(31–41) even at 1 μM could not compete with [3H]propionyl-Mas-DH for the Mt plasma membrane receptor (32). In contrast, when Sf9 cells transfected with the Mas-DH receptor were treated with Mas-DH-(3–41), this analogue stimulated cAMP production with the same potency as Mas-DH. However, Mas-DH-(13–41) did not elicit cAMP production even at 1 μM (8). These results demonstrate that some portion of the N-terminal 12 residues of Mas-DH are required for receptor activation and that probably more than 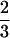 of the C-terminal portion of Mas-DH contains the determinants for receptor binding. Although no systematic structure–activity study on Mas-DH or its truncated fragments is available, it is known that Mas-DH with a free C terminus is 1/1000 as active as the native amidated peptide (1, 6). Because the fragments were formed only in the presence of Mt, our results indicate that Mt play an important role in inactivating Mas-DH.
of the C-terminal portion of Mas-DH contains the determinants for receptor binding. Although no systematic structure–activity study on Mas-DH or its truncated fragments is available, it is known that Mas-DH with a free C terminus is 1/1000 as active as the native amidated peptide (1, 6). Because the fragments were formed only in the presence of Mt, our results indicate that Mt play an important role in inactivating Mas-DH.
The absence of the C-terminal metabolites Mas-DH-(31–41) and -(30–41) in incubations with Mt, but their presence in incubations with Mt luminal fluid, suggests that these shorter metabolites are further degraded more quickly than their longer N-terminal counterparts after initial endoproteolytic cleavage. At least one of the initial Mas-DH-cleaving enzymes is EDTA sensitive, and it is probably present in both secreted (in urine) and plasma membrane-associated forms (in cleansed Mt).
Ectoenzymes (i.e., plasma membrane-bound and positioned extracellularly), especially metalloproteases, have long been implicated in regulating peptide hormone action (33). In vertebrates, a relatively few proteases are thought to be involved in the inactivation of a broad range of peptide hormones (34). However, such information in insect systems is quite limited. Adipokinetic hormones are degraded by metallopeptidases from tissue and membrane preparations of various species (10, 11, 35–37). Proctolin (14, 25) and pheromone biosynthesis-activating neuropeptide (38) are degraded by metalloproteases in different insect tissues. In vivo degradation studies have been reported for proctolin (16, 39), periplanetin CC-2 (12, 25), and adipokinetic hormones (40) after injection of labeled peptides into insects. Our study reveals unusual cleavage sites for M. sexta Mt endopeptidases at the bonds L29–R30 and R30–A31. This specificity is dissimilar to that observed in insect systems mentioned above. The major cleavage between L29–R30 cannot be attributed simply to the primary peptide sequence, as another L–R bond (L14–R15) in Mas-DH was not cleaved at 1 μM. The three-dimensional structure of Mas-DH may account for the observed major Mas-DH cleavage sites. A molecular modeling study of Mas-DH suggests that this hormone assumes a helix–loop–helix structure; L29–R30 is located in the putative loop region and L14–R15 is a constituent of the first helix (41).
Mas-DH oxidation by Mt at M2 and M11 was unexpected. However, the oxidation of Met residues in peptide hormones is well documented (42). The effects of Met oxidation on conformation and activity of those hormones is unknown (42). Although a low level of Mas-DH oxidation occurs spontaneously, Mt increased the relative level of oxidized forms (Fig. 2). A Mas-DH analogue with M2 and M11 replaced by norleucine was found to be more effective at stimulating fluid secretion by Acheta domesticus Mt than was Mas-DH (43). The authors attributed this difference to the prevention of oxidation by the amino acid substitution, but no data on the activity of oxidized Mas-DH are available.
Methionine is one of the most readily oxidized amino acid constituents of proteins; the most common oxidants present in biological systems are O2⨪ (superoxide), H2O2, HO• (hydroxyl), hypochlorite, chloramine, and peroxynitrite (44). Of these, O2⨪ normally does not oxidize methionine (42). Because catalase inhibits Mas-DH oxidation (Fig. 7A), H2O2 likely plays a role in the oxidation. Addition of EDTA significantly enhances oxidation in addition to inhibiting proteolysis (Fig. 6). This may be due to the production of the highly reactive HO• radical by means of the Fenton reaction (Fe2+EDTA + H2O2 → Fe3+EDTA + HO• + HO−). However, we were unable to detect an appreciable quantity of H2O2 in Mt luminal fluid or in medium from Mas-DH and Mt incubation, using a recent assay for H2O2 sensitive to submicromolar levels (44). This could be due to the presence of high levels of a known antioxidant, urate, in the Mt (23, 45). Phytophagous insects are known to employ superoxide dismutase (SOD), which rapidly converts O2⨪ into H2O2, as a major defense mechanism for oxidative stress (46). A study of the related lepidopteran Trichoplusia ni showed that catalase activity is completely absent from some tissues, including hemolymph and Mt, whereas SOD activity is relatively high in the same tissues (47). This evidence could explain the presence of oxidation products typical of H2O2 in the Mt. The inclusion of 20–1000 units of SOD did not affect the production of oxidized Mas-DH metabolites in the incubation (data not shown).
There are several possible sources of reactive oxygen species in Mt. The active transport required for the production of primary urine in insects demands high mitochondrial metabolism to generate ≈1 ATP per K+ or Na+ transported (48). Cytochrome oxidase is a well-known ubiquitous generator of O2⨪; the latter is converted to H2O2 by SOD. Uric acid or its metabolites allantoin and allantoic acid are the major nitrogenous wastes found in Mt (49). The synthesis of these compounds requires xanthine oxidase, urate oxidase, and allantoinase, all of which lack reducing cofactors and generate H2O2 as a product. The presence of xanthine oxidase in the fat body is well known (49). On the other hand, assay of a variety of tissues of several insects showed urate oxidase to be present only in the Mt of Calliphora, Musca domestica, and Dysdercus fasciatus and in the Mt and gut of Periplaneta (49). Transition metal ions, particularly Fe2+ and Cu2+, are important constituents of metalloproteins; at the same time they can act as catalysts for the production of reactive oxygen species (50). Indeed, a high level of the iron storage protein ferritin has been located in both the gut and the ascending region of larval Mt of M. sexta (51). Finally, a lipid carrier protein, lipophorin, has been reported to catalyze the production of O2⨪ in the hemolymph of Pseudaletia separata (52). The exact mechanism of Mas-DH oxidation warrants further investigation. The physiological relevance of the oxidation in vivo also warrants study by alternative methods. With knowledge of the major metabolic processes in hand, analysis of metabolic products of radiolabeled Mas-DH (when available) at submicromolar concentrations could determine if oxidation occurs in hemolymph, or whether by some process Mas-DH is transported into the lumen of the Mt and oxidized. Such studies could also allow addressing the question of internalization of Mas-DH bound to its receptor.
RPLC-ESI-MS is an elegant, efficient method for studying metabolism of peptide hormones by unknown pathways. A particularly powerful advantage is the ability to distinguish peptides that coelute on RPLC, which is impossible if the identity of a metabolite were determined only by comparison of its retention time with synthetic standards. This advantage is more crucial when the metabolism of a larger peptide such as Mas-DH is studied. If more classic approaches were used, standards of all metabolites from cleavage of each single peptide bond would have to be synthesized (≈80 peptides for Mas-DH) and a liquid chromatography assay capable of resolving them all would have to be developed! Variants of RPLC-ESI-MS have also been used for peptide degradation studies. For example, size-exclusion (53) and ion-exchange (54) liquid chromatography have been coupled with ESI instead of RPLC; and fast atom bombardment (54), matrix-assisted laser desorption ionization (55), and microdialysis (56) mass spectrometric techniques have been used instead of ESI-MS.
The use of capillary columns enhances the sensitivity of RPLC-ESI-MS because ESI-MS is a concentration-sensitive method. The sensitivity offered by a 0.5-mm inside diameter column and RPLC-ESI-MS technique allows detection of metabolites at an initial Mas-DH concentration of 1 μM. The main drawback of using such a column is that the low flow rate (20 μl/min) demands a long sample loading time. This can be overcome by using a peptide trap cartridge instead of a loop, which allows larger volumes of sample to be loaded quickly (<1 min).
In conclusion, Mas-DH appears to be degraded by at least one metalloendopeptidase in Mt of larval M. sexta. It is oxidized at its two Met residues by reactive oxygen species of Mt. The combined effect of proteolytic and oxidative metabolism may be responsible for inactivating Mas-DH.
Acknowledgments
We dedicate this paper to Prof. John H. Law on the occasion of his retirement. We thank Dr. R. S. Pardini for helpful discussions. This project was supported by National Institutes of Health Grant GM48172. The mass spectrometer was acquired with a Grant DIR 9102839 from the National Science Foundation. H.L. thanks the American Society for Mass Spectrometry, the 12th Montreux LC/MS Symposium Committee, and the Graduate Student Association at the University of Nevada, Reno, for financial support for presentation of these results at several conferences.
ABBREVIATIONS
- DH
diuretic hormone
- Mas-DH
Manduca sexta DH
- Mt
Malpighian tubules
- TFA
trifluoroacetic acid
- AAA
amino acid analysis
- ESI
electrospray ionization
- RPLC
reversed-phase liquid chromatography
- RPLC-ESI-MS
RPLC coupled to ESI mass spectrometry
- RIC
relative ion chromatogram
References
- 1.Kataoka H, Troetschler R G, Li J P, Kramer S J, Carney R L, Schooley D A. Proc Natl Acad Sci USA. 1989;86:2976–2980. doi: 10.1073/pnas.86.8.2976. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Furuya K, Schegg K M, Wang H, King D S, Schooley D A. Proc Natl Acad Sci USA. 1995;92:12323–12327. doi: 10.1073/pnas.92.26.12323. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Schooley D A. In: Structure and Function of Primary Messengers in Invertebrates: Insect Diuretic and Antidiuretic Peptides. Beyenbach K W, editor. Basel: Karger; 1993. pp. 22–37. [Google Scholar]
- 4.Blackburn M B, Kingan T G, Bodnar W, Shabanowitz J, Hunt D F, Kempe T, Wagner R M, Raina A K, Schnee M E, Ma M C. Biochem Biophys Res Commun. 1991;181:927–932. doi: 10.1016/0006-291x(91)92025-f. [DOI] [PubMed] [Google Scholar]
- 5.Audsley N, Coast G M, Schooley D A. J Exp Biol. 1993;178:231–243. [Google Scholar]
- 6.Audsley N, Kay I, Hayes T K, Coast G M. Comp Biochem Physiol A Physiol. 1995;110:87–93. doi: 10.1016/0300-9629(94)00132-d. [DOI] [PubMed] [Google Scholar]
- 7.Reagan J D. J Biol Chem. 1994;269:9–12. [PubMed] [Google Scholar]
- 8.Reagan J D. Insect Biochem Mol Biol. 1995;25:535–539. doi: 10.1016/0965-1748(95)00021-4. [DOI] [PubMed] [Google Scholar]
- 9.Phillips J E. In: Endocrinology of Insects. Downer R G H, Laufer H, editors. New York: Liss; 1983. pp. 411–425. [Google Scholar]
- 10.Rayne R C, O’Shea M. Insect Biochem Mol Biol. 1992;22:25–34. [Google Scholar]
- 11.Baumann E, Penzlin H. J Comp Physiol B. 1987;157:511–517. [Google Scholar]
- 12.Skinner W S, Quistad G B, Adams M E, Schooley D A. Insect Biochem. 1987;17:433–437. [Google Scholar]
- 13.Picquot M, Proux J. Regul Pept. 1990;31:139–156. doi: 10.1016/0167-0115(90)90001-d. [DOI] [PubMed] [Google Scholar]
- 14.Puiroux J, Loughton B G. Arch Insect Biochem Physiol. 1992;19:193–202. doi: 10.1002/arch.940200304. [DOI] [PubMed] [Google Scholar]
- 15.Smith A I. Peptidases and Neuropeptide Processing. San Diego: Academic; 1995. [Google Scholar]
- 16.Quistad G B, Adams M E, Scarborough R M, Carney R L, Schooley D A. Life Sci. 1984;34:569–576. doi: 10.1016/0024-3205(84)90490-9. [DOI] [PubMed] [Google Scholar]
- 17.Lamango N S, Isaac R E. Insect Biochem Mol Biol. 1993;23:801–808. doi: 10.1016/0965-1748(93)90068-4. [DOI] [PubMed] [Google Scholar]
- 18.Vogt-Schaden M, Gagelmann M, Hock D, Herbst F, Forssmann W G. Biochem Biophys Res Commun. 1989;161:1177–1183. doi: 10.1016/0006-291x(89)91366-1. [DOI] [PubMed] [Google Scholar]
- 19.Abraham L D, Chow D T, Breuil C. FEBS Lett. 1995;374:208–210. doi: 10.1016/0014-5793(95)01107-p. [DOI] [PubMed] [Google Scholar]
- 20.Brudel M, Kertscher U, Berger H, Mehlis B. J Chromatogr A. 1994;661:55–60. doi: 10.1016/0021-9673(94)85177-8. [DOI] [PubMed] [Google Scholar]
- 21.Kertscher U, Brudel M, Mehlis B, Sandow J, Berger H. J Pharm Exp Ther. 1995;273:709–715. [PubMed] [Google Scholar]
- 22.Yamamoto R T. J Econ Entomol. 1969;62:1427–1431. [Google Scholar]
- 23.Buckner J S, Caldwell J M, Reinecke J P. J Insect Physiol. 1980;26:7–12. [Google Scholar]
- 24.Cohen S A, Michaud D P. Anal Biochem. 1993;211:279–287. doi: 10.1006/abio.1993.1270. [DOI] [PubMed] [Google Scholar]
- 25.Schooley D A, Quistad G B, Skinner W S, Adams M E. In: Pesticide Science and Biotechnology. Greenhalgh R, Roberts T R, editors. Oxford: Blackwell Scientific; 1987. pp. 97–100. [Google Scholar]
- 26.Phillips J E, Audsley N. Am Zool. 1995;35:503–514. [Google Scholar]
- 27.Maddrell S H P. J Exp Biol. 1964;41:459–472. doi: 10.1242/jeb.41.3.459. [DOI] [PubMed] [Google Scholar]
- 28.Schwartz L M, Reynolds S E. J Insect Physiol. 1979;25:847–854. [Google Scholar]
- 29.Pilcher D E M. J Exp Biol. 1970;52:653–655. doi: 10.1242/jeb.52.3.653. [DOI] [PubMed] [Google Scholar]
- 30.Berridge M J. J Exp Biol. 1966;44:553–566. doi: 10.1242/jeb.44.3.553. [DOI] [PubMed] [Google Scholar]
- 31.Gee J D. J Exp Biol. 1975;63:391–401. doi: 10.1242/jeb.63.2.391. [DOI] [PubMed] [Google Scholar]
- 32.Reagan J D, Li J P, Carney R L, Kramer S J. Arch Insect Biochem Physiol. 1993;23:135–145. [Google Scholar]
- 33.Konkoy C S, Davis T P. Trends Pharm Sci. 1996;17:288–294. doi: 10.1016/0165-6147(96)10036-5. [DOI] [PubMed] [Google Scholar]
- 34.O’Cuinn G O, O’Connor B, Gilmartin L, Smyth M. In: Metabolism of Brain Peptides. O’Cuinn G O, editor. Boca Raton, FL: CRC; 1995. pp. 99–158. [Google Scholar]
- 35.Isaac R E. Biochem J. 1988;255:843–847. doi: 10.1042/bj2550843. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Fox A M, Reynolds S E. Peptides. 1991;12:937–944. doi: 10.1016/0196-9781(91)90041-m. [DOI] [PubMed] [Google Scholar]
- 37.Lamango N S, Isaac R E. Insect Biochem Mol Biol. 1993;23:801–808. doi: 10.1016/0965-1748(93)90068-4. [DOI] [PubMed] [Google Scholar]
- 38.Kuniyoshi H, Nagasawa H, Ando T, Suzuki A. Insect Biochem Mol Biol. 1992;22:399–403. [Google Scholar]
- 39.Starratt A N, Steele R W. Insect Biochem. 1984;14:97–102. [Google Scholar]
- 40.Oudejans R C H M, Vroemen S F, Jansen R F R, Van Der Horst D J. Proc Natl Acad Sci USA. 1996;93:8654–8659. doi: 10.1073/pnas.93.16.8654. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Copley K S, Welch W H, Schooley D A. In: Protein Society 10th Symposium. Neurath H, editor. San Jose, CA: Cambridge Univ. Press; 1996. p. 99. [Google Scholar]
- 42.Vogt W. Free Rad Biol Med. 1995;18:93–105. doi: 10.1016/0891-5849(94)00158-g. [DOI] [PubMed] [Google Scholar]
- 43.Coast G M, Hayes T K, Kay I, Chung J S. J Exp Biol. 1992;162:331–338. [Google Scholar]
- 44.Jiang Z Y, Woollard A C, Wolff S P. FEBS Lett. 1990;268:69–71. doi: 10.1016/0014-5793(90)80974-n. [DOI] [PubMed] [Google Scholar]
- 45.Souza A V G, Petretski J H, Oliveira P L. Free Rad Biol Med. 1997;22:209–214. doi: 10.1016/s0891-5849(96)00293-6. [DOI] [PubMed] [Google Scholar]
- 46.Ahmad S, Pardini R S. Free Rad Biol Med. 1990;8:401–413. doi: 10.1016/0891-5849(90)90107-t. [DOI] [PubMed] [Google Scholar]
- 47.Ahmad S, Duval D L, Weinhold L C, Pardini R S. Insect Biochem. 1991;21:563–572. [Google Scholar]
- 48.Azuma M, Harvey W R, Wieczorek H. FEBS Lett. 1995;361:153–156. doi: 10.1016/0014-5793(95)00146-z. [DOI] [PubMed] [Google Scholar]
- 49.Cochran D G. In: Comprehensive Insect Physiology, Biochemistry, and Pharmacology. Kerkut G A, Gilbert L I, editors. New York: Pergamon; 1985. pp. 467–506. [Google Scholar]
- 50.Stadtman E R. Free Rad Biol Med. 1990;9:315–325. doi: 10.1016/0891-5849(90)90006-5. [DOI] [PubMed] [Google Scholar]
- 51.Nichol H, Locke M. Tissue Cell. 1990;22:767–777. [Google Scholar]
- 52.Arakawa T, Kato Y, Hattori M, Yamakawa M. Insect Biochem Mol Biol. 1996;26:403–409. [Google Scholar]
- 53.Nylander I, Tan-No K, Winter A, Silberring J. Life Sci. 1995;57:123–129. doi: 10.1016/0024-3205(95)00253-3. [DOI] [PubMed] [Google Scholar]
- 54.Li Y-M, Brostedt P, Hjertén S, Nyberg F, Silberring J. J Chromatogr B. 1995;664:426–430. doi: 10.1016/0378-4347(94)00478-n. [DOI] [PubMed] [Google Scholar]
- 55.Deacon C F, Johnsen A H, Holst J J. J Clin Endocrinol Metab. 1995;80:952–957. doi: 10.1210/jcem.80.3.7883856. [DOI] [PubMed] [Google Scholar]
- 56.Andén P E, Caprioli R. J Mass Spec. 1995;30:817–824. [Google Scholar]