

Summary
The retromer is a heteropentameric complex that associates with the cytosolic face of endosomes and mediates retrograde transport of transmembrane cargo from endosomes to the trans-Golgi network. The mammalian retromer complex comprises a sorting nexin dimer composed of a still undefined combination of SNX1, SNX2, SNX5 and SNX6, and a cargo-recognition trimer composed of Vps26, Vps29 and Vps35. The SNX subunits contain PX and BAR domains that allow binding to PI(3)P enriched, highly curved membranes of endosomal vesicles and tubules, while Vps26, Vps29 and Vps35 have arrestin, phosphoesterase and α-solenoid folds, respectively. Recent studies have implicated retromer in a broad range of physiological, developmental and pathological processes, underscoring the critical nature of retrograde transport mediated by this complex.
Introduction
The retromer is a multi-subunit complex that associates with the cytosolic face of endosomes and mediates retrograde transport of transmembrane proteins from endosomes to the trans-Golgi network (TGN) [[1], [2] and [3]] (Fig. 1). The best-characterized transmembrane proteins sorted by retromer are the acid hydrolase receptors, vacuolar protein sorting 10 (Vps10) in the yeast Saccharomyces cerevisiae [[4] and [5]] and mannose 6-phosphate receptors (MPR) in mammals [[6], [7] and [8]]. The main role of these receptors is to carry newly-synthesized acid hydrolase precursors from the TGN to endosomes for eventual delivery to the vacuole and lysosomes, respectively (Fig. 1). Upon release of the acid hydrolase precursors into the endosomal lumen, the unoccupied receptors undergo retrograde transport to the TGN in a retromer-dependent manner (Fig. 1). Mutation of genes encoding retromer subunits in yeast [[4] and [5]] or RNA interference of retromer subunit expression in mammalian cells [[6], [7] and [8]] prevents the retrieval of acid hydrolase receptors to the TGN and causes their missorting to the vacuole or lysosomes, where they are degraded. As a consequence, acid hydrolase precursors are secreted and vacuolar/lysosomal degradation is impaired. Recent studies have shed light into the molecular mechanisms by which retromer mediates the sorting of acid hydrolase receptors, and have also revealed a requirement for retromer in many other physiological and developmental processes that depend on retrograde transport.
Figure 1.

Sorting of acid hydrolases mediated by mannose 6-phosphate receptors. The scheme represents the sorting of acid hydrolases by transmembrane, mannose 6-phosphate receptors (MPR) that cycle between the trans-Golgi network (TGN) and endosomes. Coats containing clathrin, GGA and AP-1 mediate exit of hydrolase-MPR complexes from the TGN, whereas retromer mediates the retrieval of unoccupied MPR from endosomes. Sequestration of MPR into recycling tubules by retromer prevents their delivery to the degradative, multivesicular body (MVB) pathway by the clathrin-ESCRT machinery. A fraction of MPR are transported from the TGN to the plasma membrane, after which they are internalized by coats containing clathrin and AP-2, joining the pool of recycling receptors in endosomes. See text for additional details and references. TEN: tubular endosomal network; H+ represents the acidic pH of the endosome lumen.
Sorting nexin subunits
Retromer is a heteropentameric complex consisting of a sorting nexin (SNX) dimer and a Vps26-Vps29-Vps35 trimer (Figs. 2 and 3). In yeast, the SNX dimer consists of Vps5 and Vps17 [[4] and [5]]. In mammals, the exact composition of the SNX dimer is less clear. SNX1 and SNX2 are largely interchangeable Vps5 orthologs [[9], [10*] and [11*]], while SNX5 and SNX6 appear be the functional mammalian orthologs of Vps17 [[12] and [13*]]. In mammals, the SNX dimer is required for the recruitment of the Vps26-Vps29-Vps35 trimer to endosomes [10]. The SNX dimer, on the other hand, can be recruited to endosomes independently of the Vps26-Vps29-Vps35 trimer [[6], [14], [15] and [16]].
Figure 2.

Domain organization of mammalian retromer subunits. Boxes indicate folded domains and lines largely disordered sequences. Number of amino acids and molecular masses of each subunit are indicated. The functions of each domain are denoted on top. All subunits occur as multiple splice forms, only one of which is represented here. The exact subunit composition of the mammalian retromer complex is not known with certainty, although a current view is that it comprises a dimer of SNX1/2 with SNX5/6, and a trimer composed of one copy each of Vps26A/B, Vps29 and Vps35.
Figure 3.

Structure of retromer. Retromer subunits are colored as in Fig. 2. The crystal structure of human Vps26A is shown in cyan (PDB 2FAU; [26*]), and those of human Vps29 and Vps35 are shown in green and red, respectively (PDB 2R17; [28**]). Residues surrounding the inactive metallophosphoesterase site of VPS29 are shown in a stick model. The structure of the SNX dimer is represented by the crystal structure of the SNX9 PX-BAR unit [24*]. The entire SNX dimer model is shown in purple, with the PI(3)P shown in a stick model. The structures are shown relative to a hypothetical model for the retromer coat bound to a membrane tubule (modified from ref. [28**]).
All SNX family members contain a PX (phox homology) domain (Figs. 2 and 3), which is responsible for binding to specific phosphoinositides [[17] and [18]]. The PX domains of SNX1 [[14], [15] and [19]], SNX2 [[20] and [21]], and SNX5 [16] have all been shown to bind phosphatidylinositol 3-phosphate (PI(3)P), and in some cases to other 3-phosphoinositides. The NMR structure of the SNX1 PX domain shows that the PI(3)P-binding pocket has fewer basic residues than that of high-affinity PI(3)P binding PX domains [19]. Because of its relatively low affinity binding, the SNX1 PX domain requires dimerization and/or association with additional domains to target to PI(3)P-containing membranes in cells.
SNX1, SNX2, SNX5, and SNX6 contain a BAR domain immediately C-terminal to their PX domain (Figs. 2 and 3). BAR domains are banana-shaped dimers with different degrees of curvature [[22] and [23]]. Some BAR domains induce, while others only sense, curvature. Although the BAR domain structures for the SNX subunits of retromer are not known, the structure of the closely related tandem PX and BAR domain portion of SNX9 was recently determined [24*]. SNX1 binds preferentially to highly curved liposomes, and at high concentrations can form high curvature membrane tubules in vitro [7]. The SNX dimer may therefore drive tubule formation and, by recruiting the cargo recognition complex, effectively coordinate cargo sorting with tubule carrier formation.
The cargo recognition complex
While the SNX dimer is responsible for the recruitment of retromer to endosomes, the Vps26-Vps29-Vps35 trimer (Figs. 2 and 3) is thought to participate in cargo binding and is therefore referred to as the ‘cargo recognition complex’. The yeast genome encodes one Vps26 species, whereas the human genome encodes two Vps26 isoforms, A and B [25]. X-ray crystallographic analyses have shown that the structures of both human Vps26 isoforms are very similar to each other, consistent with their high sequence homology [[26*] and [27*]]. Vps26 has a two-lobed structure with a polar core that is closely similar to the arrestin family of trafficking adaptors [[26*] and [27*]] (Fig. 3). The arrestins are best known for linking G-protein coupled receptors to clathrin. However, the binding sites for these receptors are not conserved in Vps26, and the overall structural similarity does not necessarily mean that these proteins have any ligands in common. Arrestins are thought to undergo a conformational change upon activation, in which the two lobes open up and expose the polar core, although the putative open state has not been directly visualized by X-ray crystallography. It also remains to be seen whether Vps26 is capable of undergoing such a conformational change. Vps26 is incorporated into the retromer complex via interactions with the Vps35 subunit [[26*] and [27*]]. Vps26 binds to Vps35 through its C-terminal lobe [[26*] and [27*]] (Fig. 3). Indeed, the two lobes of Vps26 are capable of folding independently when expressed in isolation, and the C-terminal lobe is sufficient to form a complex with Vps35 [[27*] and [28**]].
The crystal structures of human, murine, and Cryptosporidium parvum Vps29 reveal that this subunit has a metallophosphoesterase fold [[28**], [29] and [30]] (Fig. 3). The direct binding of divalent metal ions to the metallophosphoesterase-like active site has been confirmed structurally for murine Vps29 [30]. Active metallophosphoesterases contain a His residue that serves as a catalytic base and is required for activity. Vps29 contains instead a Phe residue at this position, consistent with the absence of catalytic activity. Based on the structure of the Vps29-Vps35 subcomplex [28**] and on mutational analyses [[27*] and [28**]], the metal-binding face of Vps29 appears to serve as a scaffold for the assembly of Vps35. Mutational studies also show that Vps29 contains a SNX binding site [27*], which is on the opposite face of the protein from the Vps35 binding site [28**] (Fig. 3).
The C-terminal ~40 % of the human Vps35 has been crystallized in complex with Vps29, revealing an α-solenoid fold that curves around the metal-binding face of Vps29 [28**] (Fig. 3). Bioinformatics analysis guided by the structure shows that the α-solenoid extends through the entire structure of Vps35 [28**]. This structure is reminiscent of that of the large adaptin subunits of the clathrin-associated adaptor protein (AP) complexes, AP-1 [31] and AP-2 [32], and of the clathrin heavy chain leg domains [[33] and [34]]. While the C-terminal ~300 residues of Vps35 bind to Vps29, the N-terminal ~150 are responsible for binding to Vps26 [28**] (Fig. 3). A PRLYL motif in this N-terminal portion of Vps35 is required for its interaction with Vps26 [[35*], [36*] and [37*]]. The location of the PRLYL motif suggests that Vps26 binds ‘in trans’ to Vps29, i.e., on the opposite face of the α-solenoid (Fig. 3). The retromer cargoes Vps10 and dipeptidyl aminopeptidase A (DPAP-A; also known as Ste13) bind to the elongated Vps35 structure near both ends [[38] and [39]] (Fig. 3). Binding sites for N-terminal sequences from the SNX subunits [[21] and [40]] are also located at both ends of the cargo recognition complex [[30] and [41]] (Fig. 3), suggesting that the Vps26-Vps29-Vps35 cargo recognition complex binds to membranes in a parallel manner [28**]. EM analysis shows that the cargo recognition complex is capable of curvature [28**]. This has led to the current structural model of retromer as a putative ‘coat’ for tubulovesicular structures [28**] (Figs. 3 and 4). Although this model is speculative, a newly described electron microscopic reconstruction of an F-BAR domain bound to a tubular vesicle is similar, providing an experimental precedent for such a structure [42**].
Figure 4.
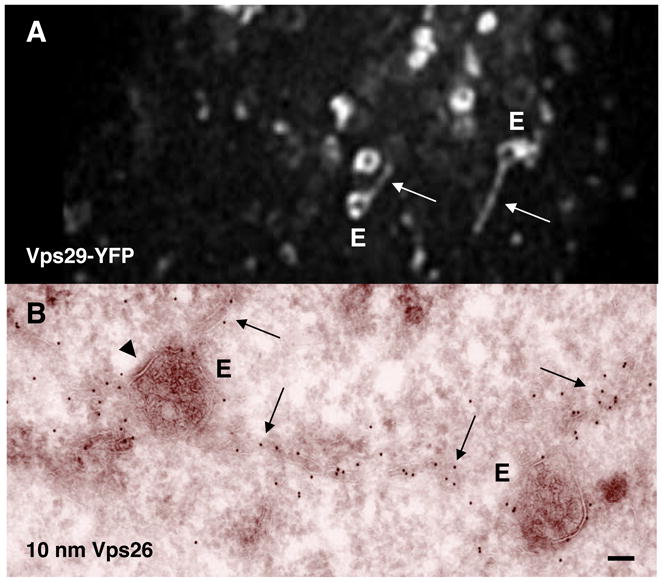
Presence of retromer on vacuolar (E) and tubular (arrows) endosomes. A. Imaging of the cytoplasm of a live cell expressing Vps29 tagged with the yellow fluorescent protein (Vps29-YFP) A video showing the dynamics of Vps29-YFP-containing endosomes can be seen at http://cellimages.ascb.org/cdm4/item_viewer.php?CISOROOT=/p4041coll12&CISOPTR=258&CISOBOX=1&REC=4. Image and video are courtesy of Gonzalo Mardones, NICHD, NIH. B. Immunoelectron microscopy of the localization of endogenous Vps26 labeled with 10 nm gold particles (reproduced with permission from ref. [6]). Arrowhead shows a flat, electron dense coat characteristic of coated endosomes. Bar: 100 nm.
Possible regulation by Rab7
The SNX and cargo recognition components of the yeast retromer form a tightly assembled heteropentameric complex that stays together upon extraction from cells and isolation by immunoprecipitation [[5] and [40]]. In contrast, the mammalian retromer complex is labile and fractionates as separate SNX and cargo recognition complexes under various conditions [10*]. Moreover, interactions of SNX1 and SNX2 with Vps26, Vps29 and Vps35 are extremely weak or undetectable when tested in isolation [[10*] and [30]]. Interactions of SNX5 and SNX6 with these Vps subunits have not been observed [13*]. It is possible that combinations of SNX1/2 with SNX5/6 might show stronger interactions, but this remains to be tested. Another possibility that has to be considered, however, is that assembly of the mammalian retromer complex on membranes involves other yet unidentified components. Interactions with the cytosolic tails of transmembrane cargoes might be a stabilizing factor. In addition, recent studies suggest that, like other protein coats, retromer might be regulated by a small GTPase of the Ras superfamily. Indeed, the GTP-bound form of Rab7 has been shown to interact with the retromer cargo recognition complex in Entamoeba histolytica [43]. Strikingly, the genome of this organism does not appear to encode any SNX orthologs. It therefore remains to be determined whether it is Rab7-GTP that promotes recruitment of the cargo recognition complex to endosomes in Entamoeba histolytica, and whether this function is conserved in mammals.
Association with vacuolar and tubular endosomes
In vivo, the mammalian retromer is associated with vacuoles that label for early endosomal markers [[6], [7] and [8]], contain intraluminal vesicles [[6], [7] and [20]], and have flat, bilayered clathrin coats [[6], [7] and [20]] (Fig. 4). These properties define the retromer-containing vacuoles as intermediates in the maturation from early to late endosomes [[44] and [45]]. In addition, retromer is found on long, highly dynamic tubules that emanate from specific foci on the vacuolar endosomes and extend for long distances along microtubules [[6], [7], [10*] and [20]] (Fig. 4). These tubules are likely intermediates in cargo transport to the TGN, and may be part of an extensive tubular endosomal network (TEN) that both recycles and sorts proteins to various destinations within the cell [2]. The existence of vesicular, endosome-to-TGN transport carriers (ETCs) emerging from SNX1-enriched regions of vacuolar endosomes has also been documented [46*]. The relationship of these vesicular carriers to the retromer-decorated tubules and the TEN remains to be determined. In any case, the function of retromer appears to be the sequestration of membrane-bound cargo proteins from the limiting membrane of the vacuolar endosomes into retrograde transport intermediates, thus preventing their default delivery to lysosomes. An important corollary of these findings is that retromer-mediated retrieval of acid hydrolase receptors starts at the level of early endosomes and proceeds as the endosomes mature to late endosomes.
Cargo recognition
Early studies in yeast identified residues in the cytosolic tails of Vps10 and DPAP-A that are important for endosome-to-TGN retrieval and that show genetic interactions with Vps35 in suppressor mutation analyses [[39], [47] and [48]]. One such set of residues from DPAP-A, FXFXD, was found to be required for binding of DPAP-A to retromer, as shown by cross-linking/co-immunoprecipitation analyses [39]. The mammalian Vps10 homologs, sortilin and SorLA, are also thought to undergo retromer-dependent transport from endosomes to the TGN [[46*], [49*] and [50]]. A recent study has identified short sequence motifs in the cytosolic tails of the cation-independent MPR (CI-MPR) (i.e., WLM) and sortilin (i.e., FLV) that interact with the retromer cargo recognition complex in co-immunoprecipitation assays and that are required for retrograde transport [49*]. Common features of the yeast and mammalian motifs are their overall hydrophobicity and presence of aromatic amino acid residues. However, it is still not known whether these motifs conform to a particular consensus. Importantly, they are distinct from other motifs that interact with the GGA adaptor proteins and with the AP-1 and AP-2 complexes [51], indicating a novel mode of recognition. Yeast two-hybrid analyses showed interaction of the cytosolic tail of the cation-independent MPR (CI-MPR) with mammalian Vps35, although the interaction was very weak and the sequences that mediate it were not identified [6]. Thus, the nature of the signal-retromer interactions that are involved in retrograde transport remains to be fully elucidated.
Other retromer-interacting proteins
Whereas the core components of retromer have probably all been identified, additional binding partners that might act as regulators or effectors of retromer function have recently been discovered. One of these partners is another yeast sorting nexin known as Grd19 or Snx3, which physically interacts with retromer in cross-linking/affinity purification analyses [52*]. Grd19 in turn recognizes the sequence GHLPFTKNLQ in the C-terminal tail of the Ftr1 component of the reductive iron transporter, Fet3-Ftr1 [52*]. These interactions are required for maintenance of the transporter at the plasma membrane under low-iron conditions, a process that involves continuous recycling between the plasma membrane, endosomes and the TGN [52*]. As for other cargoes, retromer, in conjunction with Grd19, is thought to function in the endosome-to-TGN part of this cycle. Interaction with Grd19 thus endows the retromer complex with additional cargo-recognition specificity and represents an example of binding of a retrograde transport signal by a sorting nexin instead of a Vps component.
Another retromer-binding partner, identified by co-immunoprecipitation analyses, is EHD1 [53*]. This protein has helical, nucleotide-binding/dimerization, and EH domains, and functions in tubule formation in a manner analogous to dynamin [[54] and [55]]. Moreover, EHD1 partially co-localizes with retromer on endosomal vacuoles and tubules, promotes retromer recruitment to membranes, maintains the integrity of retromer tubules, and is required for the retrieval to the TGN of chimeric proteins having the cytosolic tail of the CI-MPR [53*]. Interestingly, EHD1 had been previously implicated in the recycling of the transferrin receptor [56] and MHC class I molecules [54] from endosomes to the plasma membrane, also utilizing tubular intermediates [54]. It thus appears that EHD1 might be a common effector of tubular transport in various recycling pathways stemming from endosomes.
Involvement in multiple physiological processes
Although most of the early work on retromer focused on its role in the sorting of acid hydrolase receptors, a recent flurry of papers has uncovered an involvement of retromer in many other physiological and developmental processes (Table 1). Most of these processes are thought to rely on the retromer-mediated transport of an intracellular sorting receptor (e.g., the mammalian Vps10 homologs sortilin and SorLA [[46*], [49*] and [50]], the plant seed storage protein receptor AtVSR1/AtELP [[57*] and [58*]], the Wnt chaperone Wntless [[59*], [60*], [61*], [62*], [63*], [64*], [65*] and [66*]]), TGN or endosomal SNAREs (e.g., yeast Pep12 [67]), or a plasma membrane transporter (e.g., the auxin efflux carrier PIN [[68*] and [69*]], the reductive iron transporter Fet3-Ftr1 [52*]). All of these proteins have the property of cycling between the TGN and endosomes, or between the plasma membrane and the TGN, in the latter case with an intermediate station in endosomes. As is the case for the acid hydrolase receptors, retromer likely functions in the endosome-to-TGN leg of these cycling pathways (Fig. 1). This function of retromer is exploited by internalized bacterial exotoxins such as Shiga toxin, the B subunit of which requires retromer to travel from endosomes to the TGN on its way to the endoplasmic reticulum and the cytosol [[11*], [70*] and [71*]]. An interesting variation in this case is that Shiga toxin binds to the exofacial glycosphingolipid Gb3, which does not traverse the lipid bilayer and therefore cannot directly interact with retromer.
Table 1.
Diverse functions of retromer
| Process | Mechanism | Refs. |
|---|---|---|
| Cargo sorting mediated by intracellular receptors such as Vps10, MPRs, sortilin, SorLA, AtVSR1/AtELP, in various organisms | Retrieval of the receptors from endosomes to the TGN | [4], [5], [8], [6], [7], [10*], [46*], [49*], [50], [57*], [58*] |
| Recycling of the endosomal SNARE, Pep12 | Retrieval from endosomes to the TGN in conjunction with Grd19/Snx3 | [67] |
| Secretion of Wnt morphogens and generation of Wnt gradients during development in Caenorhabditis elegans and Drosophila melanogaster | Allows recycling to the TGN of the Wnt receptor, Wntless, so that it can participate in further rounds of Wnt transport to the plasma membrane | [59*], [60*], [61*], [62*], [63*], [64*], [65*], [66*] |
| Cell polarity and organ initiation mediated by the phytohormone, auxin, in Arabidopsis thaliana | Cycling and polarized delivery of the auxin efflux carriers, PIN | [68*], [69*] |
| Maintenance at the plasma membrane of the reductive iron transporter, Fet3-Ftr1, under low iron conditions in Saccharomyces cerevisiae | Retrieval of internalized transporter to the TGN mediated in conjunction with Grd19/Snx3 | [52*] |
| Bacterial (e.g., Shiga) toxin action | Transport of internalized toxin from endosomes to the TGN | [11*], [70*], [71*] |
| Regulation of amyloid- peptide production | Retrieval of amyloid precursor protein (APP), its sorting receptor (SorLA) or its processing peptidases (- or -secretase) from endosomes to the TGN | [50], [73], [74], [75] |
| Transcytosis of polymeric immunoglobulin receptor (pIgR) and its ligand, polymeric IgA (pIgA) | Prevents delivery of internalized pIgR-pIgA to lysosomes, thus allowing for its transport to the plasma membrane | [76], [81] |
| Endocytic trafficking, signaling and tumor suppression in Drosophila melanogaster | Inhibition of Rac1-dependent actin polymerization | [82*] |
Defects in retromer have also been proposed to underlie some forms of Alzheimer’s disease [72]. Indeed, retromer levels are reduced in affected areas of the brain of Alzheimer’s disease patients. Moreover, depletion of retromer by RNA interference increases production of amyloid-β peptide, a causative agent of Alzheimer’s disease [72]. Amyloid-β peptide is produced by proteolytic cleavage of the transmembrane amyloid precursor protein (APP) by two transmembrane endopeptidases, β-secretase and γ-secretase, a process that at least in part occurs in endosomes. The transmembrane protein SorLA has also been shown to be reduced in Alzheimer’s disease brains, to interact with APP and promote its localization to the TGN, and to traffic in a retromer-dependent manner [[50], [73], [74] and [75]]. Therefore, retromer could keep amyloid-β peptide production low by retrieving the amyloid precursor protein, SorLA, β-secretase, and/or γ-secretase from endosomes to the TGN.
Finally, retromer is also required for transport from the basolateral to the apical surface of polarized epithelial cells (i.e., ‘transcytosis’) of the polymeric immunoglobulin receptor (pIgR) and its cargo IgA [76]. This role is mediated by a specific interaction between the cytosolic tail of the pIgR and Vps35. As for many of the transmembrane proteins described above, depletion of retromer causes the pIgR to be targeted for lysosomal degradation. To date, transcytosis has not been shown to involve passage through the TGN. However, it does involve endosomal tubules [77] that could also be part of the TEN [2]. Thus, removal of transmembrane cargo from vacuolar to tubular endosomes, whether for transport to the TGN or to the apical plasma membrane, might be a more general role of retromer.
Concluding remarks
Retromer has recently come into sharp focus as a novel sorting device involved in the physiologically critical process of rescuing transmembrane proteins from the limiting membrane of vacuolar endosomes for recycling to the TGN. Its role thus counteracts that of the ESCRT machinery [[78] and [79]], which functions to sort transmembrane proteins from the limiting membrane of endosomes into intraluminal vesicles (Fig. 1). The distinct actions of these two sorting devices contribute to the biochemical and structural changes that occur during maturation of early endosomes to late endosomes. Because of the opposing effects of retromer and ESCRT complexes on endosomal membranes, it is not surprising that they have different structures and mechanisms of action. Topologically, the role of retromer is more akin to that of clathrin coats that function in protein sorting at the plasma membrane and TGN [80]. Indeed, as discussed above, retromer subunits share some common structural motifs and regulatory mechanisms with clathrin-associated proteins. However, the overall arrangement of the structural motifs and specific mechanistic details are quite distinct, indicating that retromer is in a class of its own.
Recent discoveries highlight the role of retromer in diverse biological pathways, from lysosome biogenesis to developmental signaling. These pathways in turn involve molecular interactions with a wide range of cargo molecules. Thus far no conserved retromer-interaction motif has come to light. However, the tools are now coming into place to address the molecular mechanisms for cargo selection, and to determine to what extent conserved vs. specialized motifs are utilized. Despite the diversity of biological functions and biochemical interactions of retromer, the concept of it as a BAR-domain based coat for tubulovesicular structures is consistent with the available data. The abundance of tubulovesicular structures in eukaryotic cells suggests retromer may have many yet-to-be discovered biological roles. Furthermore, the prevalence of BAR domains and their variants in eukaryotic proteomes suggests that there are likely to be other types of tubulovesicular coats based on a retromer-like structural paradigm. Experimentation aimed at these questions should provide new insights into the biological roles of retromer and tubulovesicular trafficking. On the mechanistic front, the recent structural and functional findings provide a foothold for analyses of tubulovesicular trafficking that may parallel but also diverge from paradigms established for conventional vesicle trafficking. The initiation, growth, scission, transport, docking and fusion of retromer-coated tubules or derived carriers will be fruitful ground for mechanistic studies in coming years.
Acknowledgments
We thank Raul Rojas, Yogikala Prabhu, Steve Nothwehr, Chris Burd and Pete Cullen for helpful discussions and comments on the manuscript, and Gonzalo Mardones for Fig. 4A. Work in our labs is supported by the Intramural Programs of NICHD and NIDDK, NIH.
Footnotes
Publisher's Disclaimer: This is a PDF file of an unedited manuscript that has been accepted for publication. As a service to our customers we are providing this early version of the manuscript. The manuscript will undergo copyediting, typesetting, and review of the resulting proof before it is published in its final citable form. Please note that during the production process errors may be discovered which could affect the content, and all legal disclaimers that apply to the journal pertain.
References and recommended reading
- 1.Seaman MN. Recycle your receptors with retromer. Trends Cell Biol. 2005;15:68–75. doi: 10.1016/j.tcb.2004.12.004. [DOI] [PubMed] [Google Scholar]
- 2.Bonifacino JS, Rojas R. Retrograde transport from endosomes to the trans-Golgi network. Nat Rev Mol Cell Biol. 2006;7:568–579. doi: 10.1038/nrm1985. [DOI] [PubMed] [Google Scholar]
- 3.Verges M. Retromer and sorting nexins in development. Front Biosci. 2007;12:3825–3851. doi: 10.2741/2355. [DOI] [PubMed] [Google Scholar]
- 4.Horazdovsky BF, Davies BA, Seaman MN, McLaughlin SA, Yoon S, Emr SD. A sorting nexin-1 homologue, Vps5p, forms a complex with Vps17p and is required for recycling the vacuolar protein-sorting receptor. Mol Biol Cell. 1997;8:1529–1541. doi: 10.1091/mbc.8.8.1529. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Seaman MN, McCaffery JM, Emr SD. A membrane coat complex essential for endosome-to-Golgi retrograde transport in yeast. J Cell Biol. 1998;142:665–681. doi: 10.1083/jcb.142.3.665. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Arighi CN, Hartnell LM, Aguilar RC, Haft CR, Bonifacino JS. Role of the mammalian retromer in sorting of the cation-independent mannose 6-phosphate receptor. J Cell Biol. 2004;165:123–133. doi: 10.1083/jcb.200312055. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Carlton J, Bujny M, Peter BJ, Oorschot VM, Rutherford A, Mellor H, Klumperman J, McMahon HT, Cullen PJ. Sorting nexin-1 mediates tubular endosome-to-TGN transport through coincidence sensing of high- curvature membranes and 3-phosphoinositides. Curr Biol. 2004;14:1791–1800. doi: 10.1016/j.cub.2004.09.077. [DOI] [PubMed] [Google Scholar]
- 8.Seaman MN. Cargo-selective endosomal sorting for retrieval to the Golgi requires retromer. J Cell Biol. 2004;165:111–122. doi: 10.1083/jcb.200312034. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Griffin CT, Trejo J, Magnuson T. Genetic evidence for a mammalian retromer complex containing sorting nexins 1 and 2. Proc Natl Acad Sci U S A. 2005;102:15173–15177. doi: 10.1073/pnas.0409558102. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10*.Rojas R, Kametaka S, Haft CR, Bonifacino JS. Interchangeable but essential functions of SNX1 and SNX2 in the association of retromer with endosomes and the trafficking of mannose 6-phosphate receptors. Mol Cell Biol. 2007;27:1112–1124. doi: 10.1128/MCB.00156-06. In agreement with genetic studies in mice (ref. [9]), cellular studies show that the sorting nexins SNX1 and SNX2 are interchangeable components of the mammalian retromer complex. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11*.Utskarpen A, Slagsvold HH, Dyve AB, Skanland SS, Sandvig K. SNX1 and SNX2 mediate retrograde transport of Shiga toxin. Biochem Biophys Res Commun. 2007;358:566–570. doi: 10.1016/j.bbrc.2007.04.159. Together with refs. [70*] and [71*], this study demonstrates an involvement of retromer in the transport of Shiga toxin from early endosomes to the TGN. Since Shiga toxin binds to an exofacial glycosphingolipid, this transport does not depend on a direct interaction of the toxin or its receptor with retromer. [DOI] [PubMed] [Google Scholar]
- 12.Liu H, Liu ZQ, Chen CX, Magill S, Jiang Y, Liu YJ. Inhibitory regulation of EGF receptor degradation by sorting nexin 5. Biochem Biophys Res Commun. 2006;342:537–546. doi: 10.1016/j.bbrc.2006.01.179. [DOI] [PubMed] [Google Scholar]
- 13*.Wassmer T, Attar N, Bujny MV, Oakley J, Traer CJ, Cullen PJ. A loss-of-function screen reveals SNX5 and SNX6 as potential components of the mammalian retromer. J Cell Sci. 2007;120:45–54. doi: 10.1242/jcs.03302. The mammalian sorting nexins SNX5 and SNX6 are proposed to be the functional equivalents of the yeast Vps17 subunit of retromer. [DOI] [PubMed] [Google Scholar]
- 14.Cozier GE, Carlton J, McGregor AH, Gleeson PA, Teasdale RD, Mellor H, Cullen PJ. The phox homology (PX) domain-dependent, 3-phosphoinositide-mediated association of sorting nexin-1 with an early sorting endosomal compartment is required for its ability to regulate epidermal growth factor receptor degradation. J Biol Chem. 2002;277:48730–48736. doi: 10.1074/jbc.M206986200. [DOI] [PubMed] [Google Scholar]
- 15.Zhong Q, Lazar CS, Tronchere H, Sato T, Meerloo T, Yeo M, Songyang Z, Emr SD, Gill GN. Endosomal localization and function of sorting nexin 1. Proc Natl Acad Sci U S A. 2002;99:6767–6772. doi: 10.1073/pnas.092142699. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Merino-Trigo A, Kerr MC, Houghton F, Lindberg A, Mitchell C, Teasdale RD, Gleeson PA. Sorting nexin 5 is localized to a subdomain of the early endosomes and is recruited to the plasma membrane following EGF stimulation. J Cell Sci. 2004;117:6413–6424. doi: 10.1242/jcs.01561. [DOI] [PubMed] [Google Scholar]
- 17.Worby CA, Dixon JE. Sorting out the cellular functions of sorting nexins. Nat Rev Mol Cell Biol. 2002;3:919–931. doi: 10.1038/nrm974. [DOI] [PubMed] [Google Scholar]
- 18.Carlton JG, Cullen PJ. Sorting nexins. Curr Biol. 2005;15:R819–820. doi: 10.1016/j.cub.2005.10.012. [DOI] [PubMed] [Google Scholar]
- 19.Zhong Q, Watson MJ, Lazar CS, Hounslow AM, Waltho JP, Gill GN. Determinants of the endosomal localization of sorting nexin 1. Mol Biol Cell. 2005;16:2049–2057. doi: 10.1091/mbc.E04-06-0504. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Carlton JG, Bujny MV, Peter BJ, Oorschot VM, Rutherford A, Arkell RS, Klumperman J, McMahon HT, Cullen PJ. Sorting nexin-2 is associated with tubular elements of the early endosome, but is not essential for retromer-mediated endosome-to-TGN transport. J Cell Sci. 2005;118:4527–4539. doi: 10.1242/jcs.02568. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Gullapalli A, Garrett TA, Paing MM, Griffin CT, Yang Y, Trejo J. A role for sorting nexin 2 in epidermal growth factor receptor down-regulation: evidence for distinct functions of sorting nexin 1 and 2 in protein trafficking. Mol Biol Cell. 2004;15:2143–2155. doi: 10.1091/mbc.E03-09-0711. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Gallop JL, McMahon HT. BAR domains and membrane curvature: bringing your curves to the BAR. Biochem Soc Symp. 2005:223–231. doi: 10.1042/bss0720223. [DOI] [PubMed] [Google Scholar]
- 23.Itoh T, De Camilli P. BAR, F-BAR (EFC) and ENTH/ANTH domains in the regulation of membrane-cytosol interfaces and membrane curvature. Biochim Biophys Acta. 2006;1761:897–912. doi: 10.1016/j.bbalip.2006.06.015. [DOI] [PubMed] [Google Scholar]
- 24*.Pylypenko O, Lundmark R, Rasmuson E, Carlsson SR, Rak A. The PX-BAR membrane-remodeling unit of sorting nexin 9. Embo J. 2007;26:4788–4800. doi: 10.1038/sj.emboj.7601889. This structure of the SNX9 PX-BAR dimer provides a useful model for the SNX dimer component of retromer. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Kerr MC, Bennetts JS, Simpson F, Thomas EC, Flegg C, Gleeson PA, Wicking C, Teasdale RD. A novel mammalian retromer component, Vps26B. Traffic. 2005;6:991–1001. doi: 10.1111/j.1600-0854.2005.00328.x. [DOI] [PubMed] [Google Scholar]
- 26*.Shi H, Rojas R, Bonifacino JS, Hurley JH. The retromer subunit Vps26 has an arrestin fold and binds Vps35 through its C-terminal domain. Nat Struct Mol Biol. 2006;13:540–548. doi: 10.1038/nsmb1103. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27*.Collins BM, Norwood SJ, Kerr MC, Mahony D, Seaman MN, Teasdale RD, Owen DJ. Structure of Vps26B and mapping of its interaction with the retromer protein complex. Traffic. 2007 doi: 10.1111/j.1600-0854.2007.00688.x. The structures of Vps26A and Vps26B reported in the above two papers reveal an unexpected structural similarity to the arrestins. While the functional significance of this similarity is still unclear, the structures provide a key building block for understanding retromer assembly. [DOI] [PubMed] [Google Scholar]
- 28*.Hierro A, Rojas AL, Rojas R, Murthy N, Effantin G, Kajava AV, Steven AC, Bonifacino JS, Hurley JH. Functional architecture of the retromer cargo-recognition complex. Nature. 2007;449:1063–1067. doi: 10.1038/nature06216. The structure of Vps35 and the structural basis for its interactions with Vps29 was revealed. A low-resolution molecular structure for the entire Vps26-Vps29-Vps35 cargo recognition complex was developed, and a model for the docking of retromer to tubular endosomes was proposed. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Wang D, Guo M, Liang Z, Fan J, Zhu Z, Zang J, Li X, Teng M, Niu L, Dong Y, et al. Crystal structure of human vacuolar protein sorting protein 29 reveals a phosphodiesterase/nuclease-like fold and two protein-protein interaction sites. J Biol Chem. 2005;280:22962–22967. doi: 10.1074/jbc.M500464200. [DOI] [PubMed] [Google Scholar]
- 30.Collins BM, Skinner CF, Watson PJ, Seaman MN, Owen DJ. Vps29 has a phosphoesterase fold that acts as a protein interaction scaffold for retromer assembly. Nat Struct Mol Biol. 2005;12:594–602. doi: 10.1038/nsmb954. [DOI] [PubMed] [Google Scholar]
- 31.Heldwein EE, Macia E, Wang J, Yin HL, Kirchhausen T, Harrison SC. Crystal structure of the clathrin adaptor protein 1 core. Proc Natl Acad Sci U S A. 2004;101:14108–14113. doi: 10.1073/pnas.0406102101. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Collins BM, McCoy AJ, Kent HM, Evans PR, Owen DJ. Molecular architecture and functional model of the endocytic AP2 complex. Cell. 2002;109:523–535. doi: 10.1016/s0092-8674(02)00735-3. [DOI] [PubMed] [Google Scholar]
- 33.ter Haar E, Musacchio A, Harrison SC, Kirchhausen T. Atomic structure of clathrin: a beta propeller terminal domain joins an alpha zigzag linker. Cell. 1998;95:563–573. doi: 10.1016/s0092-8674(00)81623-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Ybe JA, Brodsky FM, Hofmann K, Lin K, Liu SH, Chen L, Earnest TN, Fletterick RJ, Hwang PK. Clathrin self-assembly is mediated by a tandemly repeated superhelix. Nature. 1999;399:371–375. doi: 10.1038/20708. [DOI] [PubMed] [Google Scholar]
- 35*.Gokool S, Tattersall D, Reddy JV, Seaman MN. Identification of a conserved motif required for Vps35p/Vps26p interaction and assembly of the retromer complex. Biochem J. 2007;408:287–295. doi: 10.1042/BJ20070555. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36*.Restrepo R, Zhao X, Peter H, Zhang BY, Arvan P, Nothwehr SF. Structural features of vps35p involved in interaction with other subunits of the retromer complex. Traffic. 2007;8:1841–1853. doi: 10.1111/J.1600-0854.2007.00659.X. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37*.Zhao X, Nothwehr S, Lara-Lemus R, Zhang BY, Peter H, Arvan P. Dominant-Negative Behavior of Mammalian Vps35 in Yeast Requires a Conserved PRLYL Motif Involved in Retromer Assembly. Traffic. 2007;8:1829–1840. doi: 10.1111/j.1600-0854.2007.00658.x. The preceding three papers report the identification of a conserved structural motif, PRLYL, on Vps35 that is involved in interaction with Vps26 and assembly of the retromer complex. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Nothwehr SF, Bruinsma P, Strawn LA. Distinct domains within Vps35p mediate the retrieval of two different cargo proteins from the yeast prevacuolar/endosomal compartment. Mol Biol Cell. 1999;10:875–890. doi: 10.1091/mbc.10.4.875. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Nothwehr SF, Ha SA, Bruinsma P. Sorting of yeast membrane proteins into an endosome-to-Golgi pathway involves direct interaction of their cytosolic domains with Vps35p. J Cell Biol. 2000;151:297–310. doi: 10.1083/jcb.151.2.297. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Seaman MN, Williams HP. Identification of the functional domains of yeast sorting nexins Vps5p and Vps17p. Mol Biol Cell. 2002;13:2826–2840. doi: 10.1091/mbc.02-05-0064. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Haft CR, de la Luz Sierra M, Bafford R, Lesniak MA, Barr VA, Taylor SI. Human orthologs of yeast vacuolar protein sorting proteins Vps26, 29, and 35: assembly into multimeric complexes. Mol Biol Cell. 2000;11:4105–4116. doi: 10.1091/mbc.11.12.4105. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42*.Frost A, Perera R, Roux A, Spasov K, Destaing O, Egelman EH, De Camilli P, Unger VM. Structural Basis of Membrane Invagination by F-BAR Domains. Cell. 2008;132:807–817. doi: 10.1016/j.cell.2007.12.041. The EM reconstruction of a BAR domain coat for a tubular vesicle provides the first experimental model for what the SNX-BAR portion of the retromer coat might look like. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Nakada-Tsukui K, Saito-Nakano Y, Ali V, Nozaki T. A retromer like complex is a novel Rab7 effector that is involved in the transport of the virulence factor cysteine protease in the enteric protozoan parasite Entamoeba histolytica. Mol Biol Cell. 2005;16:5294–5303. doi: 10.1091/mbc.E05-04-0283. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Raposo G, Tenza D, Murphy DM, Berson JF, Marks MS. Distinct protein sorting and localization to premelanosomes, melanosomes, and lysosomes in pigmented melanocytic cells. J Cell Biol. 2001;152:809–824. doi: 10.1083/jcb.152.4.809. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Peden AA, Oorschot V, Hesser BA, Austin CD, Scheller RH, Klumperman J. Localization of the AP-3 adaptor complex defines a novel endosomal exit site for lysosomal membrane proteins. J Cell Biol. 2004;164:1065–1076. doi: 10.1083/jcb.200311064. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46*.Mari M, Bujny MV, Zeuschner D, Geerts WJ, Griffith J, Petersen CM, Cullen PJ, Klumperman J, Geuze HJ. SNX1 defines an early endosomal recycling exit for sortilin and mannose 6-phosphate receptors. Traffic. 2007 doi: 10.1111/j.1600-0854.2007.00686.x. Together with ref. [49*], this paper shows that retromer participates in retrograde transport of sortilin from endosomes to the TGN. [DOI] [PubMed] [Google Scholar]
- 47.Cooper AA, Stevens TH. Vps10p cycles between the late-Golgi and prevacuolar compartments in its function as the sorting receptor for multiple yeast vacuolar hydrolases. J Cell Biol. 1996;133:529–541. doi: 10.1083/jcb.133.3.529. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Nothwehr SF, Roberts CJ, Stevens TH. Membrane protein retention in the yeast Golgi apparatus: dipeptidyl aminopeptidase A is retained by a cytoplasmic signal containing aromatic residues. J Cell Biol. 1993;121:1197–1209. doi: 10.1083/jcb.121.6.1197. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49*.Seaman MN. Identification of a novel conserved sorting motif required for retromer-mediated endosome-to-TGN retrieval. J Cell Sci. 2007;120:2378–2389. doi: 10.1242/jcs.009654. This study identifies novel sequences within the cytosolic tails of the cation-independent mannose 6-phosphate receptor and sortilin that interact with the mammalian retromer and function as signals for retrograde transport from endosomes to the TGN. [DOI] [PubMed] [Google Scholar]
- 50.Nielsen MS, Gustafsen C, Madsen P, Nyengaard JR, Hermey G, Bakke O, Mari M, Schu P, Pohlmann R, Dennes A, et al. Sorting by the cytoplasmic domain of the amyloid precursor protein binding receptor SorLA. Mol Cell Biol. 2007;27:6842–6851. doi: 10.1128/MCB.00815-07. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Bonifacino JS, Traub LM. Signals for sorting of transmembrane proteins to endosomes and lysosomes. Annu Rev Biochem. 2003;72:395–447. doi: 10.1146/annurev.biochem.72.121801.161800. [DOI] [PubMed] [Google Scholar]
- 52*.Strochlic TI, Setty TG, Sitaram A, Burd CG. Grd19/Snx3p functions as a cargo-specific adapter for retromer-dependent endocytic recycling. J Cell Biol. 2007;177:115–125. doi: 10.1083/jcb.200609161. Grd19/Snx3, a yeast sorting nexin distinct from the retromer components Vps5 and Vps17, is shown to interact with the retromer complex, conferring additional cargo-recognition specificity on the complex. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53*.Gokool S, Tattersall D, Seaman MN. EHD1 Interacts with Retromer to Stabilize SNX1 Tubules and Facilitate Endosome-to-Golgi Retrieval. Traffic. 2007;8:1873–1886. doi: 10.1111/j.1600-0854.2007.00652.x. The tubule-forming protein EHD1 is shown to interact with retromer and to cooperate with it in retrograde transport from endosomes to the TGN. [DOI] [PubMed] [Google Scholar]
- 54.Caplan S, Naslavsky N, Hartnell LM, Lodge R, Polishchuk RS, Donaldson JG, Bonifacino JS. A tubular EHD1-containing compartment involved in the recycling of major histocompatibility complex class I molecules to the plasma membrane. Embo J. 2002;21:2557–2567. doi: 10.1093/emboj/21.11.2557. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Daumke O, Lundmark R, Vallis Y, Martens S, Butler PJ, McMahon HT. Architectural and mechanistic insights into an EHD ATPase involved in membrane remodelling. Nature. 2007;449:923–927. doi: 10.1038/nature06173. [DOI] [PubMed] [Google Scholar]
- 56.Lin SX, Grant B, Hirsh D, Maxfield FR. Rme-1 regulates the distribution and function of the endocytic recycling compartment in mammalian cells. Nat Cell Biol. 2001;3:567–572. doi: 10.1038/35078543. [DOI] [PubMed] [Google Scholar]
- 57*.Shimada T, Koumoto Y, Li L, Yamazaki M, Kondo M, Nishimura M, Hara-Nishimura I. AtVPS29, a putative component of a retromer complex, is required for the efficient sorting of seed storage proteins. Plant Cell Physiol. 2006;47:1187–1194. doi: 10.1093/pcp/pcj103. [DOI] [PubMed] [Google Scholar]
- 58*.Fuji K, Shimada T, Takahashi H, Tamura K, Koumoto Y, Utsumi S, Nishizawa K, Maruyama N, Hara-Nishimura I. Arabidopsis vacuolar sorting mutants (green fluorescent seed) can be identified efficiently by secretion of vacuole-targeted green fluorescent protein in their seeds. Plant Cell. 2007;19:597–609. doi: 10.1105/tpc.106.045997. Refs. [57*] and [58*] report an involvement of the plant retromer in sorting of seed storage protein to granules through its role in recycling of the sorting receptor AtVSR1/AtELP from endosome to the TGN. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59*.Belenkaya TY, Wu Y, Tang X, Zhou B, Cheng L, Sharma YV, Yan D, Selva EM, Lin X. The Retromer Complex Influences Wnt Secretion by Recycling Wntless from Endosomes to the Trans-Golgi Network. Dev Cell. 2007 doi: 10.1016/j.devcel.2007.12.003. [DOI] [PubMed] [Google Scholar]
- 60*.Pan CL, Baum PD, Gu M, Jorgensen EM, Clark SG, Garriga GC. elegans AP-2 and Retromer Control Wnt Signaling by Regulating MIG-14/Wntless. Dev Cell. 2007 doi: 10.1016/j.devcel.2007.12.001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61*.Yang PT, Lorenowicz MJ, Silhankova M, Coudreuse DY, Betist MC, Korswagen HC. Wnt Signaling Requires Retromer-Dependent Recycling of MIG-14/Wntless in Wnt-Producing Cells. Dev Cell. 2007 doi: 10.1016/j.devcel.2007.12.004. [DOI] [PubMed] [Google Scholar]
- 62*.Prasad BC, Clark SG. Wnt signaling establishes anteroposterior neuronal polarity and requires retromer in C. elegans. Development. 2006;133:1757–1766. doi: 10.1242/dev.02357. [DOI] [PubMed] [Google Scholar]
- 63*.Coudreuse DY, Roel G, Betist MC, Destree O, Korswagen HC. Wnt gradient formation requires retromer function in Wnt-producing cells. Science. 2006;312:921–924. doi: 10.1126/science.1124856. [DOI] [PubMed] [Google Scholar]
- 64*.George A, Leahy H, Zhou J, Morin PJ. The vacuolar-ATPase inhibitor bafilomycin and mutant VPS35 inhibit canonical Wnt signaling. Neurobiol Dis. 2007;26:125–133. doi: 10.1016/j.nbd.2006.12.004. [DOI] [PubMed] [Google Scholar]
- 65*.Franch-Marro X, Wendler F, Guidato S, Griffith J, Baena-Lopez A, Itasaki N, Maurice MM, Vincent JP. Wingless secretion requires endosome-to-Golgi retrieval of Wntless/Evi/Sprinter by the retromer complex. Nat Cell Biol. 2008 doi: 10.1038/ncb1678. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 66*.Port F, Kuster M, Herr P, Furger E, Banziger C, Hausmann G, Basler K. Wingless secretion promotes and requires retromer-dependent cycling of Wntless. Nat Cell Biol. 2008 doi: 10.1038/ncb1687. The preceding eight papers illustrate the critical role of retromer in organismal development by demonstrating its indirect involvement in the secretion of the Wnt morphogen through recycling of the transmembrane Wnt chaperone, Wntless, from endosomes to the TGN. [DOI] [PubMed] [Google Scholar]
- 67.Hettema EH, Lewis MJ, Black MW, Pelham HR. Retromer and the sorting nexins Snx4/41/42 mediate distinct retrieval pathways from yeast endosomes. Embo J. 2003;22:548–557. doi: 10.1093/emboj/cdg062. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 68*.Jaillais Y, Fobis-Loisy I, Miege C, Rollin C, Gaude T. AtSNX1 defines an endosome for auxin-carrier trafficking in Arabidopsis. Nature. 2006;443:106–109. doi: 10.1038/nature05046. [DOI] [PubMed] [Google Scholar]
- 69*.Jaillais Y, Santambrogio M, Rozier F, Fobis-Loisy I, Miege C, Gaude T. The retromer protein VPS29 links cell polarity and organ initiation in plants. Cell. 2007;130:1057–1070. doi: 10.1016/j.cell.2007.08.040. Refs. [68*] and [69*] report that the plant retromer mediates dynamic cycling and polarization of the PIN auxin efflux transporters, thus mediating the formation of new axes of development. [DOI] [PubMed] [Google Scholar]
- 70*.Bujny MV, Popoff V, Johannes L, Cullen PJ. The retromer component sorting nexin-1 is required for efficient retrograde transport of Shiga toxin from early endosome to the trans Golgi network. J Cell Sci. 2007;120:2010–2021. doi: 10.1242/jcs.003111. [DOI] [PubMed] [Google Scholar]
- 71*.Popoff V, Mardones GA, Tenza D, Rojas R, Lamaze C, Bonifacino JS, Raposo G, Johannes L. The retromer complex and clathrin define an early endosomal retrograde exit site. J Cell Sci. 2007;120:2022–2031. doi: 10.1242/jcs.003020. See annotation to ref. [11*]. [DOI] [PubMed] [Google Scholar]
- 72.Small SA, Kent K, Pierce A, Leung C, Kang MS, Okada H, Honig L, Vonsattel JP, Kim TW. Model-guided microarray implicates the retromer complex in Alzheimer’s disease. Ann Neurol. 2005;58:909–919. doi: 10.1002/ana.20667. [DOI] [PubMed] [Google Scholar]
- 73.Scherzer CR, Offe K, Gearing M, Rees HD, Fang G, Heilman CJ, Schaller C, Bujo H, Levey AI, Lah JJ. Loss of apolipoprotein E receptor LR11 in Alzheimer disease. Arch Neurol. 2004;61:1200–1205. doi: 10.1001/archneur.61.8.1200. [DOI] [PubMed] [Google Scholar]
- 74.Andersen OM, Reiche J, Schmidt V, Gotthardt M, Spoelgen R, Behlke J, von Arnim CA, Breiderhoff T, Jansen P, Wu X, et al. Neuronal sorting protein-related receptor sorLA/LR11 regulates processing of the amyloid precursor protein. Proc Natl Acad Sci U S A. 2005;102:13461–13466. doi: 10.1073/pnas.0503689102. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 75.Offe K, Dodson SE, Shoemaker JT, Fritz JJ, Gearing M, Levey AI, Lah JJ. The lipoprotein receptor LR11 regulates amyloid beta production and amyloid precursor protein traffic in endosomal compartments. J Neurosci. 2006;26:1596–1603. doi: 10.1523/JNEUROSCI.4946-05.2006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 76.Verges M, Luton F, Gruber C, Tiemann F, Reinders LG, Huang L, Burlingame AL, Haft CR, Mostov KE. The mammalian retromer regulates transcytosis of the polymeric immunoglobulin receptor. Nat Cell Biol. 2004;6:763–769. doi: 10.1038/ncb1153. [DOI] [PubMed] [Google Scholar]
- 77.Verges M, Havel RJ, Mostov KE. A tubular endosomal fraction from rat liver: biochemical evidence of receptor sorting by default. Proc Natl Acad Sci U S A. 1999;96:10146–10151. doi: 10.1073/pnas.96.18.10146. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 78.Saksena S, Sun J, Chu T, Emr SD. ESCRTing proteins in the endocytic pathway. Trends Biochem Sci. 2007;32:561–573. doi: 10.1016/j.tibs.2007.09.010. [DOI] [PubMed] [Google Scholar]
- 79.Hurley JH. ESCRT complexes and the biogenesis of multivesicular bodies. Curr Opin Cell Biol. 2008;20:4–11. doi: 10.1016/j.ceb.2007.12.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 80.Edeling MA, Smith C, Owen D. Life of a clathrin coat: insights from clathrin and AP structures. Nat Rev Mol Cell Biol. 2006;7:32–44. doi: 10.1038/nrm1786. [DOI] [PubMed] [Google Scholar]
- 81.Verges M, Sebastian I, Mostov KE. Phosphoinositide 3-kinase regulates the role of retromer in transcytosis of the polymeric immunoglobulin receptor. Exp Cell Res. 2007;313:707–718. doi: 10.1016/j.yexcr.2006.11.010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 82*.Korolchuk VI, Schutz MM, Gomez-Llorente C, Rocha J, Lansu NR, Collins SM, Wairkar YP, Robinson IM, O’Kane CJ. Drosophila Vps35 function is necessary for normal endocytic trafficking and actin cytoskeleton organisation. J Cell Sci. 2007;120:4367–4376. doi: 10.1242/jcs.012336. Drosophila retromer is shown to inhibit Rac1-induced actin polymerization, thus regulating receptor endocytosis and signaling. [DOI] [PubMed] [Google Scholar]