

Abstract
Sugar transport by some permeases in Escherichia coli is allosterically regulated by the phosphorylation state of the intracellular regulatory protein, enzyme IIAglc of the phosphoenolpyruvate:sugar phosphotransferase system. A sensitive radiochemical assay for the interaction of enzyme IIAglc with membrane-associated lactose permease was used to characterize the binding reaction. The binding is stimulated by transportable substrates such as lactose, melibiose, and raffinose, but not by sugars that are not transported (maltose and sucrose). Treatment of lactose permease with N-ethylmaleimide, which blocks ligand binding and transport by alkylating Cys-148, also blocks enzyme IIAglc binding. Preincubation with the substrate analog β-d-galactopyranosyl 1-thio-β-d-galactopyranoside protects both lactose transport and enzyme IIAglc binding against inhibition by N-ethylmaleimide. A collection of lactose permease replacement mutants at Cys-148 showed, with the exception of C148V, a good correlation of relative transport activity and enzyme IIAglc binding. The nature of the interaction of enzyme IIAglc with the cytoplasmic face of lactose permease was explored. The N- and C-termini, as well as five hydrophilic loops in the permease, are exposed on the cytoplasmic surface of the membrane and it has been proposed that the central cytoplasmic loop of lactose permease is the major determinant for interaction with enzyme IIAglc. Lactose permease mutants with polyhistidine insertions in cytoplasmic loops IV/V and VI/VII and periplasmic loop VII/VIII retain transport activity and therefore substrate binding, but do not bind enzyme IIAglc, indicating that these regions of lactose permease may be involved in recognition of enzyme IIAglc. Taken together, these results suggest that interaction of lactose permease with substrate promotes a conformational change that brings several cytoplasmic loops into an arrangement optimal for interaction with the regulatory protein, enzyme IIAglc. A topological map of the proposed interaction is presented.
Bacteria use a wide range of carbon sources that are transported across the cytoplasmic membrane by a variety of specific transport systems. The phosphoenolpyruvate:sugar phosphotransferase system (PTS), a multifunctional system consisting of several protein components, is widespread in bacteria. The primary function of the PTS is the concomitant phosphorylation and translocation of a variety of sugar substrates from the medium (1, 2). In the case of glucose transport in Gram-negative bacteria, three soluble proteins (enzyme I, HPr, and enzyme IIAglc) and one membrane-bound protein (enzyme IIB, Cglc) are required for transport.
In Escherichia coli, the PTS also regulates uptake of various non-PTS sugars such as lactose, maltose, melibiose, galactose, and raffinose (inhibition of non-PTS inducer transport by PTS sugars is termed “inducer exclusion”), as well as the phosphorylation of glycerol and the synthesis of cyclic AMP (catabolite repression) (2–6). These regulatory roles of the PTS have been proposed to be mediated by the glucose-specific enzyme IIAglc, and the currently accepted model suggests that the phosphorylation state of IIAglc determines whether or not it binds to a target protein (2).
E. coli lactose permease (lac permease) is an important model for the study of membrane transport proteins. Lac permease is one of the most extensively investigated secondary active transport proteins that transduce free energy stored in an electrochemical ion gradient into a solute concentration gradient (7–9). It is a hydrophobic, polytopic membrane protein that catalyzes the coupled stoichiometric translocation of β-galactosides and H+. All available evidence indicates strongly that lac permease is comprised of a bundle of 12 transmembrane α-helices that traverse the membrane in zigzag fashion connected by hydrophilic loops with the N and C termini on the cytoplasmic face (7, 10). Moreover, the arrangement of the helices in the tertiary structure has been determined (11).
It has been shown that lac permease is allosterically regulated by direct binding of IIAglc (12, 13). Although a lac permease triple mutant in the N-terminal tail was reported to lack IIAglc binding (14), it has been suggested (15, 16) that the allosteric regulation of lac permease is mediated mainly by an interaction of IIAglc with the central cytoplasmic loop.
In the present work we describe a direct method that uses [3H]-labeled IIAglc to quantitate binding of IIAglc to lac permease. The requirement for a substrate-induced conformational change in the permease for IIAglc binding is established. Membrane preparations enriched in lac permease (17, 18) with mutations or insertions in each loop were used to survey sites essential for binding to IIAglc. The results, in conjunction with the recently proposed helix packing model, provide the basis for a proposed topological map of the region of lac permease that interacts with IIAglc.
MATERIALS AND METHODS
Bacterial Strains and Plasmids.
Membrane vesicles were prepared from E. coli K12 strain T184[lacI+O+ Z-Y-(A) rpsL Met-Thr- recA hsdM hsdR/F′ lacIqO+ZD118(Y+A+)] (19) harboring given plasmids. IIAglc was prepared by using E. coli strain HK759 transformed with pRK248 and pEL110 as described (20). Use of plasmids pT7–5/lacY (Ampr, T7 φ10 promoter J) for overproduction of wild type, the 6-histidine insertion mutants, and a C-terminal deletion mutant of lac permease were described (17) (see Fig. 4A). Plasmids used for expression of lac permease harboring site-directed mutations of Cys-148 were described by Jung et al. (18).
Figure 4.
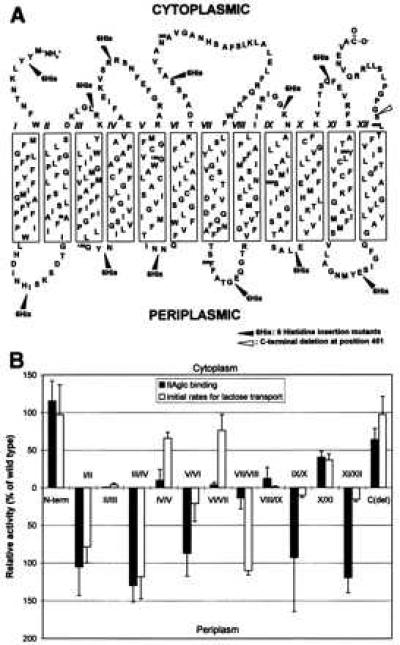
Binding of IIAglc to lactose permease insertion and deletion mutants. In A, the secondary-structure model of E. coli lac permease is shown with 12 6-histidine insertion sites (solid arrowheads) and one C-terminal deletion site (open arrowhead) in the polypeptide chain. The single-letter amino acid code is used, and the 12 putative transmembrane helices are shown in boxes. The model is based on the combination of the hydropathy plot of the primary amino acid sequence of lac permease (10) and some experimental refinements (11). In B, the effect of mutations of lactose permease on the TDG-dependent binding of IIAglc (▪) was compared with that on the initial rates of lactose transport (not normalized) of the permease (□). Expression levels of lac permease from each strain (except the C-terminal deletion mutation) were determined and the binding activity to EIIAglc was corrected as described in the legend to Fig. 3. Because the antibody used for quantitation of expression of lac permease is directed against the C terminus (21), it was not possible to quantitate permease expression in the C-terminal deletion. Consequently, the binding observed in that construct (63% of the control) was not corrected for the level of expression. Therefore, it is likely that the C-terminal deletion is unaffected with respect to IIAglc binding. Binding assays were done three to five times and SDs were plotted. The initial rates for lactose transport of the lac permease mutants came from a previous report (17) and are also expressed as the percentage of wild-type activity.
Membrane Preparations.
Cells grown overnight at 37°C were used to inoculate Luria–Bertani medium supplemented with 50 μg/ml of ampicillin and 20 μg/ml of streptomycin. When the culture reached an A600 = 0.5, isopropyl 1-thio-β-d-galactopyranoside was added to a final concentration of 1 mM and growth was continued for 4 h. Cells were harvested by centrifugation (6,000 × g for 10 min at 4°C), washed twice in 50 mM Na⋅phosphate buffer (pH 6.3) and either stored at −20°C until use or used immediately. For the preparation of membrane vesicles, harvested cells were suspended in 50 mM Na⋅phosphate buffer (pH 6.3) and broken by two passages through a French pressure cell at 11,000 psi. Cell debris and unbroken cells were removed by centrifugation at 10,000 × g for 10 min. The membranes then were collected by centrifugation at 100,000 × g for 90 min, washed once with 50 mM Na⋅phosphate (pH 6.3), and pelleted at 100,000 × g for 90 min. The resulting membrane vesicles were resuspended in the same buffer (20 mg protein/ml), and stored frozen at −80°C until use. Protein was estimated by using bicinchoninic acid protein assay reagents (Pierce). Expression of lac permeases was determined by quantitative Western blot analysis with anti-C-terminal antibody, as described (21).
Preparation of [3H]-Labeled E. coli enzyme IIAglc.
E. coli HK759 transformed with pRK248 and pEL110 was used for the expression of IIAglc. The inoculation medium (medium M) contained: 1× M9 salts, 1 mM MgSO4, 1% glucose, 30 μg/ml thiamine, 50 μg/ml arginine, 10 μg/ml isoleucine, 10 μg/ml valine, 20 μg/ml threonine, 20 μg/ml leucine, 1 mM MgCl2, 0.1 mM CaCl2, 60 μg/ml FeSO4⋅7H2O, 12 μg/ml MnCl2⋅4H2O, 8 μg/ml CoCl2⋅6H2O, 7 μg/ml ZnSO4⋅7H2O, 3 μg/ml CuCl2⋅2H2O, 0.2 μg/l H3BO3, 2.5 μg/ml (NH4)6Mo7O24⋅4H2O, and 50 μg/ml EDTA. Cells were grown in a total volume of 500 ml at 30°C in medium M and induction was initiated by shifting the temperature to 41°C when the A600 reached about 0.5. Threonine (10 μg/ml) and [3H]leucine (10 μg/ml, 2 mCi; 1 Ci = 37 GBq) were added again after the temperature shift to label the expressed protein; 6 h after induction, the cells were harvested and the protein was purified as described (20). Specific radioactivity of the purified [3H]IIAglc was 1,080 cpm/μg of protein (counting efficiency in Beckman LS6500 scintillation system ≈60%).
Binding Assays.
Incubations were carried out at room temperature in polyallomer tubes (5 × 20 mm) designed for use in the Beckman Airfuge. Incubation mixtures contained (total volume of 100 μl): 50 mM Na⋅phosphate (pH 6.3), 2 mM DTT, 2 mM MgCl2, 0.5 mg of membrane protein unless otherwise indicated, and 1.2 μg of [3H]-labeled IIAglc. Sugars (2 mM) were added as indicated. After incubation for 5 min, the membranes were collected by centrifugation at 100,000 × g for 15 min in an Airfuge. After discarding the supernatants, the tubes were rinsed with 100 μl of 50 mM Na⋅phosphate (pH 6.3) containing 1 mM DTT, the pellets were resuspended in 100 μl of 1% Triton X-100, and membrane-bound IIAglc was quantitated by liquid scintillation spectrometry after adding 10 ml of Aquasol (DuPont). All binding assays were done at least two times and all results were reproducible.
RESULTS
Binding of IIAglc to Lac Permease Requires Substrate.
The previously described assays (12, 13) for the interaction of IIAglc with membrane-bound permeases were indirect. Bound IIAglc was eluted from pelleted membranes and was then quantitated by rocket electrophoresis (12) or assays for complementation of PTS activity (13). In the experiments reported here, the more direct approach of measuring membrane-bound radioactive IIAglc is used. The data in Fig. 1 validate this method. Although membrane vesicles prepared from E. coli T184 cells transformed with expression vector pT7–5 (labeled control) do not exhibit sugar-dependent binding, vesicles prepared from cells transformed with pT7–5 expressing the wild-type lac permease (labeled LacY) bind IIAglc in a substrate-dependent manner. When 1.2 μg of IIAglc is incubated with these membrane vesicles (0.5 mg protein), TDG addition results in five times as much binding of IIAglc and ≈40% of the added IIAglc is bound. Of several sugars tested, melibiose yields the greatest stimulation of IIAglc binding (6× the control value), raffinose and lactose also show considerable stimulation (340 and 400% of control, respectively), but sucrose and maltose (that are not substrates of lac permease) do not stimulate IIAglc binding. TDG-dependent binding is proportional to the concentration of membrane protein added, as shown in the inset of Fig. 1. These data are consistent (12, 13) with the model that a substrate-induced conformational change in lac permease is required for interaction with IIAglc.
Figure 1.
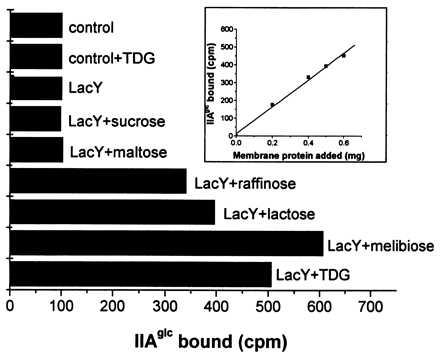
Binding of IIAglc to lactose permease in the presence of various sugars. Membrane vesicles were prepared from strains T184/pT7–5 (control) and T184/pT7–5/cassette lacY (LacY) harvested 4 h after induction with 1 mM isopropyl 1-thio-β-d-galactopyranoside as described in Materials and Methods. Membrane vesicles (0.5 mg protein) were mixed with 1.2 μg of [3H]IIAglc (1080 cpm/μg of protein) in 50 mM Na⋅phosphate buffer, pH 6.3, containing 2 mM DTT and 2 mM MgCl2 in total volume of 100 μl in the presence or absence of 2 mM sugars as indicated. After incubation at room temperature for 5 min, the membranes were separated in an Airfuge and the bound and free IIAglc were determined as described in Materials and Methods. The inset shows the TDG-dependent binding between IIAglc and membrane vesicles enriched in lac permease as a function of membrane protein concentration. TDG-dependent binding was determined by subtracting the 3H counts in the membrane pellet of the incubation mixture without added sugar from the 3H counts of membrane-bound [3H]IIAglc in the presence of TDG. Binding assays were done more than twice and SD was <5%.
IIAglc Binding Is Sensitive to N-Ethylmaleimide (NEM).
Early studies showed that NEM, a sulfhydryl-specific reagent, inactivates lactose transport activity and that the inactivation is blocked by TDG (22). It was later shown that Cys-148 is the residue protected against NEM blockade by substrate and that this residue interacts hydrophobically with the galactosyl moiety of the substrate (18, 23). As shown in Fig. 2, TDG-dependent IIAglc binding (column labeled −NEM) is essentially completely eliminated by preincubation with NEM (column labeled NEM) (12) and, as expected, inactivation of NEM with DTT (column labeled DTT) abolishes blockade of IIAglc binding. Importantly, preincubation of IIAglc with NEM (column labeled IIAglc) does not affect its capability to bind to the permease, demonstrating that NEM acts on the permease rather than IIAglc. The possibility that substrate protects the permease against NEM-dependent inhibition of IIAglc binding (12) was also tested. TDG gives ≈80% protection, whereas melibiose (column labeled mel) and raffinose (column labeled raf) afford less protection (30% and 25%, respectively). Sucrose, maltose and lactose (columns labeled suc, mal, and lac, respectively) are ineffective. These experiments are consistent with the argument that Cys-148 is part of the substrate binding region. Modification of this region by NEM prevents lac permease from assuming a substrate-induced conformation that permits binding of IIAglc.
Figure 2.
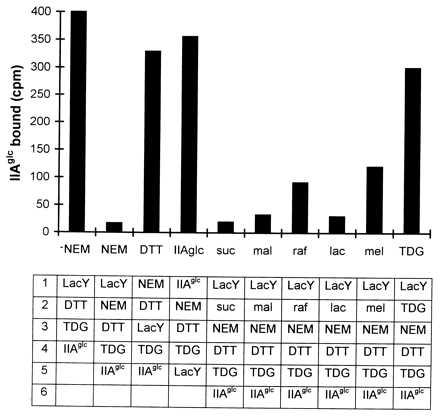
Effect of N-ethylmaleimide on the binding of IIAglc to lactose permease. The effect of modification of cysteine residues on the interaction between lac permease and IIAglc was tested by treating the protein with 1 mM NEM. The table shows the order of addition to each incubation mixture containing 50 mM Na⋅phosphate buffer, pH 6.3 and 2 mM MgCl2. For efficient modification of cysteine residues and deactivation of NEM, the reaction mixture was incubated for 5 min at room temperature after each addition of NEM (1 mM) and DTT (10 mM), respectively. -NEM, no NEM treatment; NEM, membranes enriched in lac permease were treated with NEM first; DTT, NEM was deactivated with DTT before adding proteins; IIAglc, IIAglc was treated with NEM first. The capability of a variety of sugars (each at 2 mM concentration) to block the NEM-dependent inhibition of the interaction of IIAglc and lac permease was tested: suc, sucrose; mal, maltose; raf, raffinose; lac, lactose; mel, melibiose. Binding assays were done more than twice and the findings were reproducible.
Cys-148 Is Not Essential for IIAglc Binding to Lac Permease.
Lac permease mutants in which Cys-148 was replaced with other amino acid residues (18, 23) were tested for IIAglc binding before and after treatment with NEM (Fig. 3). Consistent with the data of Fig. 2, IIAglc binding to wild-type permease (columns labeled WT) is inhibited by NEM. Replacement of Cys-148 by alanine results in an ≈40% increase in TDG-dependent IIAglc binding that is essentially insensitive to NEM. Other replacements at Cys-148 exhibit varying levels of IIAglc binding activity. Only the alanine and valine replacements have IIAglc binding activities near that of wild-type lac permease. None of the Cys-148 replacements are sensitive to NEM. Thus, Cys-148 is the site of NEM inactivation of IIAglc binding to lac permease.
Figure 3.
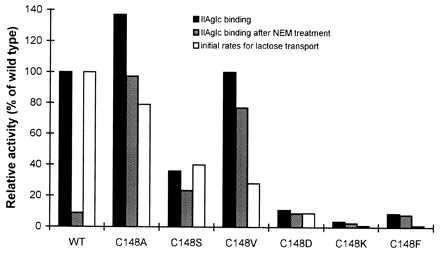
Effect of replacements of Cys-148 of lactose permease on the binding to IIAglc. Several Cys-148 replacement mutants of lac permease were tested for binding to IIAglc without (▪) or with (▨) NEM treatment (the data are corrected for IIAglc binding in the absence of TDG), and a comparison was made with the initial rate of lactose transport (□) determined previously (18). The interaction was measured as described in Fig. 2. Expression levels of lac permease from each strain were determined by quantitative phosphoimaging using polyclonal antibody against the C terminus of the permease (21) and the relative binding activity of each mutant to IIAglc was corrected for expression level using the concentration curve shown in the inset to Fig. 1 and compared with that of wild-type lac permease. Binding experiments were done at least two times and the results were reproducible.
The pattern of IIAglc binding activity in the lac permease variants was compared with the previously studied (18) pattern for the initial rates of lactose transport (Fig. 3). With one exception, activity for lactose transport parallels IIAglc binding activity. In the case of the valine substitution, IIAglc binding activity is preserved, but lac transport activity is reduced to ≈30% of the control. It is interesting to note that this mutant has a higher apparent affinity for substrate than the wild-type lac permease (18). These data further support the notion that substrate binding to lac permease results in a conformation suitable for interaction with IIAglc.
Some 6-His Insertion Mutants of lac Permease Are Defective in IIAglc Binding.
It has been reported (16, 24) that the central cytoplasmic loop (loop VI/VII) of lac permease is important for IIAglc binding. The possible involvement of different cytoplasmic loops of lac permease in IIAglc binding was evaluated by examining TDG-dependent IIAglc binding to membranes containing mutants of lac permease with 6 contiguous His residues inserted into each loop (17) or the N terminus; a deletion of the C-terminal tail also was examined (see Fig. 4A). The results are expressed as a percentage of wild-type activity in Fig. 4B. Interestingly, four out of seven cytoplasmic loop insertion mutants (loops II/III, IV/V, VI/VII, and VIII/IX) show markedly diminished IIAglc binding (<20% of wild type), whereas only one of six periplasmic loop insertion mutants (loop VII/VIII) shows diminished binding (Fig. 4B). It is noteworthy that, based on the helix-packing model (11), all of the IIAglc-binding defective mutants have insertions in loops located close to one another. In contrast, the N-terminal hydrophilic domain tolerates a 6-histidine insertion with no effect on IIAglc binding, and deletion of the C-terminal tail has only a small effect.
For comparative purposes, previously reported data (17, 25) on the initial rates of transport for these preparations are shown. The pattern of sensitivity to loop insertions for IIAglc binding and transport activity differs markedly. Activity is severely diminished by insertions in two cytoplasmic (loops II/III and VIII/IX) and three periplasmic loops (V/VI, IX/X and XI/XII) (Fig. 4B). In contrast, IIAglc binding is defective in four cytoplasmic (loops II/III, IV/V, VI/VII and VIII/IX) and one periplasmic (loop VII/VIII) loops. The results suggest that multiple loops of lac permease are involved in the interaction with IIAglc.
DISCUSSION
The use of radiolabeled IIAglc provides an important technological advance for studies of the PTS-dependent regulation of permeases. It is now possible to quickly and accurately measure the binding of IIAglc to membrane vesicle preparations directly. The studies presented here use this approach that should produce results less subject to experimental variability than those previously presented (12, 13). The efficacy of the direct Airfuge method is demonstrated by the observation (Fig. 1) that only substrates that are transported by lac permease (raffinose, lactose, melibiose, and TDG) stimulate IIAglc binding; sugars that are not lac permease substrates (sucrose and maltose) do not stimulate binding. Protection by sugars against NEM inhibition of IIAglc binding (Fig. 2) also mirrors the same pattern of substrates. Furthermore, IIAglc binding is linearly related to permease concentration over a reasonable range of permease concentrations.
NEM alkylates Cys-148 of lac permease (18, 23). The alkylated permease is defective with respect to both transport and IIAglc binding. The obvious explanation for this behavior is that permease alkylated at Cys-148 no longer binds substrate, a property necessary for both transport and IIAglc binding.
Cys-148 in lac permease makes direct contact with substrate, interacting hydrophobically with the galactosyl end of the substrate (18, 23). Replacements of Cys-148 by other amino acids (Fig. 3) lead to varying degrees of loss of both transport activity and IIAglc binding, with the exception of the Cys-148-Val mutant. Because this mutant has a higher affinity for substrate (18), it appears likely that the property determining the interaction of IIAglc with lac permease is substrate binding rather than rate of transport. This is consistent with the idea that the binding of substrate to lac permease induces a conformational change that allows effective interaction with IIAglc.
The data in Fig. 4B suggest strongly that the interaction of IIAglc with lac permease involves contacts with at least one loop (IV/V) in addition to VI/VII. Insertions in these two loops result in retention of transport activity, but loss of IIAglc binding. We propose that the relative alignment of these two loops with respect to one another is important for effective IIAglc binding.
Insertions in loops II/III and VIII/IX lead to loss of both transport and IIAglc binding (Fig. 4B). Because it is not clear whether or not these mutants bind TDG, the possible requirement for those loops in IIAglc binding cannot yet be evaluated. Experiments with site-directed mutants localized in these loops should clarify this issue.
Interestingly, loop VII/VIII is the only periplasmic loop where a 6-histidine insertion markedly decreases IIAglc binding. Previous studies demonstrate that this loop contains the epitope for mAb4B1 binding (21) that has been shown to cause conformational changes at the cytoplasmic end of helix X (26).
Based on the current studies and the recently presented (11) helix packing scheme for lac permease, it is possible to devise a topological model (Fig. 5) for the association of IIAglc with lac permease. It is proposed that IIAglc binding requires interaction with residues in loops IV/V and VI/VII (and perhaps the flanking loops as well). For effective protein-protein interaction, the multiple sites must be in a defined orientation with respect to one another. Insertions in the cytoplasmic loops comprising portions of the interaction surface are expected to disrupt the normal arrangement and result in decreased IIAglc binding. The requirement of a specific arrangement of the helices for IIAglc binding may explain why it has not been possible to show any interaction of IIAglc with purified solubilized lac permease by surface plasmon resonance (unpublished data).
Figure 5.
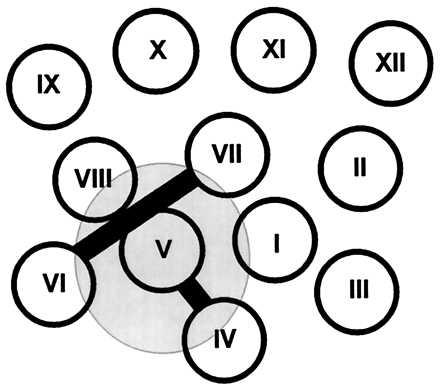
Helix packing model of lactose permease viewed from cytoplasm showing the possible interaction region with IIAglc. The transmembrane helices are depicted as circles, and the proposed interaction region with IIAglc is shown in gray. Loops IV/V and VI/VII, proposed to be sites of interaction with IIAglc, are represented as solid bars.
Some studies have focused attention on the possible importance of the N-terminal region (Fig. 4A) of lac permease for IIAglc binding. Hoischen et al. (16) showed that lac permease deleted for residues 2–7 or 2–8 retains regulation by the PTS. The present studies show that insertion of 6-histidine residues between Tyr-2 and Tyr-3 does not diminish IIAglc binding. In contrast, a lac permease mutant (Thr-7-Ile, Met-11-Ile) does not exhibit IIAglc binding (12, 14). It should be noted that Met-11 is located in transmembrane helix I; modification of this residue might result in a change in helix packing and consequently loss of IIAglc binding. On the other hand, modification of the N-terminal region not found in the helix should not affect the relationship of one helix to another.
From rotational diffusion measurements, the effective surface area of lac permease in the plane of the membrane is estimated to be ≈1,200 Å2 (27). The model presented in Fig. 5 suggests that a significant portion of this surface can be covered by interaction with IIAglc. NMR analysis (28) and modeling studies (29) of the complex of IIAglc with HPr indicate that a hydrophobic patch on IIAglc occupying ≈500 Å2 interacts with HPr. The model of Fig. 5 suggests that a similarly sized region of the IIAglc structure may be involved in complex formation with lac permease. X-ray crystallographic analysis of the regulatory complex of IIAglc with glycerol kinase indicated that the interface buries ≈1300 Å2 of molecular surface including the hydrophobic patch of IIAglc.
Mutants in IIAglc that abolish only binding to lac permease but not phosphotransferase activity have been isolated and characterized (30). The residues mutated were Gly-47, Ala-76, and Ser-78; these residues are on the same face of IIAglc as is the hydrophobic patch that interacts with HPr. It is also noteworthy that the hydrophobic patch of IIAglc surrounds the active site histidine (His-90) that, when phosphorylated, results in a form of IIAglc that does not bind to lac permease. These observations suggest strongly that there is a common interface on IIAglc that can interact with HPr (28, 29), glycerol kinase (4) or lac permease, although there are no obvious sequence similarities among these three proteins. It might be predicted that overexpression of HPr or glycerol kinase might reduce the sensitivity of lac permease to inhibition by IIAglc.
In summary, we have presented evidence that the interaction of IIAglc with lac permease is dependent on a conformational change induced by the binding substrate. The interaction requires a precise alignment of IIAglc on the cytoplasmic face of lac permease to allow multiple specific contacts in which at least 30% of the cytoplasmic surface of lac permease may be covered by IIAglc.
Footnotes
This paper was submitted directly (Track II) to the Proceedings Office.
Abbreviations: lac permease, E. coli lactose permease; TDG, β-d-galactopyranosyl 1-thio-β-d-galactopyranoside; IIAglc, E. coli enzyme IIAglc; PTS, phosphoenolpyruvate:sugar phosphotransferase system; NEM, N-ethylmaleimide.
References
- 1.Kaback H R. J Biol Chem. 1968;243:3711–3724. [PubMed] [Google Scholar]
- 2.Postma P W, Lengeler J W, Jacobson G R. Microbiol Rev. 1993;57:543–594. doi: 10.1128/mr.57.3.543-594.1993. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Titgemeyer F, Mason R E, Saier M H., Jr J Bacteriol. 1994;176:543–546. doi: 10.1128/jb.176.2.543-546.1994. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Hurley J H, Faber H R, Worthylake D, Meadow N D, Roseman S, Pettigrew D W, Remington S J. Science. 1993;259:673–677. [PubMed] [Google Scholar]
- 5.Peterkofsky A, Reizer A, Reizer J, Gollop N, Zhu P, Amin N. Prog Nucleic Acid Res Mol Biol. 1993;44:31–65. doi: 10.1016/s0079-6603(08)60216-0. [DOI] [PubMed] [Google Scholar]
- 6.Peterkofsky A, Seok Y-J, Amin N, Thapar R, Lee S Y, Klevit R E, Waygood E B, Anderson J W, Gruschus J, Huq H, Gollop N. Biochemistry. 1995;34:8950–8959. doi: 10.1021/bi00028a003. [DOI] [PubMed] [Google Scholar]
- 7.Kaback H R. In: Handbook of Biological Physics: Transport Processes in Eukaryotic and Prokaryotic Organisms. Konings W N, Kaback H R, Lolkema J S, editors. Amsterdam: Elsevier; 1996. pp. 203–227. [Google Scholar]
- 8.Kaback H R. J Membr Biol. 1983;76:95–112. doi: 10.1007/BF02000610. [DOI] [PubMed] [Google Scholar]
- 9.Kaback H R, Frillingos S, Jung H, Jung K, Privé G G, Ujwal M L, Weitzman C, Wu J, Zen K. J Exp Biol. 1994;196:183–195. doi: 10.1242/jeb.196.1.183. [DOI] [PubMed] [Google Scholar]
- 10.Foster D L, Boublik M, Kaback H R. J Biol Chem. 1983;258:31–34. [PubMed] [Google Scholar]
- 11.Kaback H R, Voss J, Wu J. Curr Opin Struct Biol. 1997;7:537–542. doi: 10.1016/s0959-440x(97)80119-4. [DOI] [PubMed] [Google Scholar]
- 12.Nelson S O, Wright J K, Postma P W. EMBO J. 1983;2:715–720. doi: 10.1002/j.1460-2075.1983.tb01490.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Osumi T, Saier M H., Jr Proc Natl Acad Sci USA. 1982;79:1457–1461. doi: 10.1073/pnas.79.5.1457. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Overath P, Weigel U, Neuhaus J-M, Soppa J, Seckler R, Riede I, Bocklage H, Muller-Hill B, Aichele G, Wright J K. Proc Natl Acad Sci USA. 1987;84:5535–5539. doi: 10.1073/pnas.84.16.5535. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Wilson T H, Yunker P L, Hansen C L. Biochim Biophys Acta. 1990;1029:113–116. doi: 10.1016/0005-2736(90)90443-r. [DOI] [PubMed] [Google Scholar]
- 16.Hoischen C, Levin J, Pitaknarongphorn S, Reizer J, Saier M H., Jr J Bacteriol. 1996;178:6082–6086. doi: 10.1128/jb.178.20.6082-6086.1996. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.McKenna E, Hardy D, Kaback H R. Proc Natl Acad Sci USA. 1992;89:11954–11958. doi: 10.1073/pnas.89.24.11954. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Jung H, Jung K, Kaback H R. Biochemistry. 1994;33:12160–12165. doi: 10.1021/bi00206a019. [DOI] [PubMed] [Google Scholar]
- 19.Teather R M, Bramhall J, Riede I, Wright J K, Furst M, Aichele G, Wilhelm U, Overath P. Eur J Biochem. 1980;108:223–231. doi: 10.1111/j.1432-1033.1980.tb04715.x. [DOI] [PubMed] [Google Scholar]
- 20.Reddy P, Fredd-Kuldell N, Liberman E, Peterkofsky A. Protein Expression Purif. 1991;2:179–187. doi: 10.1016/1046-5928(91)90069-u. [DOI] [PubMed] [Google Scholar]
- 21.Sun J, Wu J, Carrasco N, Kaback H R. Biochemistry. 1996;35:990–998. doi: 10.1021/bi952166w. [DOI] [PubMed] [Google Scholar]
- 22.Fox C F, Kennedy E P. Proc Natl Acad Sci USA. 1965;54:891–899. doi: 10.1073/pnas.54.3.891. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Wu J, Kaback H R. Biochemistry. 1994;33:12166–12171. doi: 10.1021/bi00206a020. [DOI] [PubMed] [Google Scholar]
- 24.Saier M H, Jr, Ramseier T M, Reizer J. In: Escherichia coli and Salmonella: Cellular and Molecular Biology. 2nd Ed. Neidhardt F C, editor. Washington, DC: Am. Soc. Microbiol.; 1996. pp. 1325–1343. [Google Scholar]
- 25.Kaback H R. Int Rev Cytol. 1992;137:97–125. doi: 10.1016/s0074-7696(08)62674-1. [DOI] [PubMed] [Google Scholar]
- 26.Frillingos S, Wu J, Venkatesan P, Kaback H R. Biochemistry. 1997;36:6408–6414. doi: 10.1021/bi970233b. [DOI] [PubMed] [Google Scholar]
- 27.Dornmair K, Corin A F, Wright J K, Jähnig F. EMBO J. 1985;4:3633–3638. doi: 10.1002/j.1460-2075.1985.tb04127.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Chen Y, Reizer J, Saier M H, Jr, Fairbrother W J, Wright P E. Biochemistry. 1993;32:32–37. doi: 10.1021/bi00052a006. [DOI] [PubMed] [Google Scholar]
- 29.Herzberg O. J Biol Chem. 1992;267:24819–24823. [PubMed] [Google Scholar]
- 30.Zeng G Q, De Reuse H, Danchin A. Res Microbiol. 1992;143:251–261. doi: 10.1016/0923-2508(92)90017-i. [DOI] [PubMed] [Google Scholar]