

Abstract
The monoclonal a-70-kDa heat shock protein (hsp70) antibody recognizes in crude extracts from watermelon (Citrullus vulgaris) cotyledons two hsps with molecular masses of 70 and 72 kDa. Immunocytochemistry on watermelon cotyledon tissue and on isolated glyoxysomes identified hsp70s in the matrix of glyoxysomes and plastids. Affinity purification and partial amino acid determination revealed the 70-kDa protein to share high sequence identity with cytosolic hsp70s from a number of plant species, while the 72 kDa protein was very similar to plastid hsp70s from pea and cucumber. A full-length cDNA clone encoding the 72-kDa hsp70 was isolated and identified two start methionines in frame within the N-terminal presequence leading either to an N-terminal extension of 67 amino acids or to a shorter one of 47 amino acids. The longer presequence was necessary and sufficient to target a reporter protein into watermelon proplastids in vitro. The shorter extension starting from the second methionine within the long version harbored a consensus peroxisomal targeting signal (RT-X5-KL) that directed in vivo a reporter protein into peroxisomes of the yeast Hansenula polymorpha. Peroxisomal targeting was however prevented, when the 67-residue presequence was fused to the reporter protein, indicating that the peroxisomal targeting signal 2 information is hidden in this context. We propose that the 72-kDa hsp70 is encoded by a single gene, but targeted alternatively into two organelles by the modulated use of its presequence.
Keywords: peroxisomes, microbodies, protein targeting, peroxisomal targeting signal 2, Citrullus vulgaris
Seventy-kilodalton heat shock proteins (hsp70s) serve as ubiquitous molecular chaperones in DNA replication, protein folding, and transport. Organelle hsp70s (e.g., in plastids, mitochondria, and endoplasmic reticulum) characteristically contain N-terminal presequences with the organelle-specific targeting information; the endoplasmic reticulum hsp70 is also provided with a C-terminal retention signal. In eukaryotic cells cytosolic hsp70s play an essential role in the delivery of proteins to their target organelle by binding to nascent polypeptide chains and thereby keeping them in a translocation competent conformation. Organellar hsp70s located, for example, in the plastid or mitochondrial matrix and in the endoplasmic reticulum lumen, are involved in the protein translocation process (1–3). So far, hsp70s have not been encountered in microbodies (peroxisomes/glyoxysomes), though cytosolic hsp70s have been implicated to stimulate peroxisomal protein import in in vitro studies with permeabilized cells (4) and are associated with the outside of the microbody membrane (5, 26). Their specific in vivo role is still a matter of debate since the peroxisomal protein import machinery seems capable of transferring oligomeric folded proteins (6).
In this paper, we describe the isolation of a watermelon (Citrullus vulgaris) hsp70 that occurs in its mature form in both plastids and glyoxysomes. Analysis of mRNA molecules for this hsp70 reveals two start methionine in frame within the presequence leading to two types of precursor proteins: the longer polypeptide exhibits a 67-amino acid N-terminal presequence that functions in plastid import. The shorter version of the presequence directs the hsp70 into peroxisomes with a PTS2, which becomes functional when translation starts with the methionine located at position 21 of the plastidal presequence.
MATERIALS AND METHODS
Organisms and Growth Conditions.
Watermelon seeds (C. vulgaris Schrad., var. Stone Mountain, harvest 1992; Landreth Seed, Baltimore) were germinated under sterile conditions in the dark at 30°C for 3 days. Hansenula polymorpha A16 was grown in flasks in mineral medium on methanol (0.5% vol/vol) as C-source and (NH4)2SO4 (0.25% wt/vol) or methylamine (0.20% wt/vol) as N-source (7).
Affinity Chromatography of hsp70s.
Cotyledons were homogenized to yield a protein extract (8) and hsp70s isolated (9). Watermelon glyoxysomal malate dehydrogenase (gMDH) (10) was immobilized on cyanogen bromide-activated Sepharose 4B (Sigma), denatured with 8 M urea and used as an affinity matrix for binding hsp70s from the extract in the presence of 1 mM Mg-ATP. After washing proteins were eluted using five column volumes of 10 mM Mg-ATP.
Recombinant DNA Methods and Biochemical Procedures.
Recombinant DNA manipulations, DNA-sequencing, immunoscreening of a cDNA-library with monoclonal anti-hsp70 antibody (Sigma, catalog no. H5147), SDS/PAGE and Western blot analysis were performed as usual. For amino acid sequencing, hsp70s were separated by SDS/PAGE and cleaved in the gel using endoprotease Lys-C (Boehringer Mannheim). The resulting peptides were microsequenced (11). Escherichia coli DH5α was transformed by the Ca2+ method and H. polymorpha by electroporation (12).
Isolation of a Full-Length hsp70 cDNA Clone.
A cDNA library from mRNA of 2-day-old dark grown watermelon cotyledons was cloned into the phage vector Lambda ZAP Express (Stratagene). After screening of 9.5 × 105 plaques with a-hsp70 antibodies 18 positive clones were isolated and the phages converted by in vivo excision into the plasmid vector pBluescript K-cytomegalovirus. Sequencing of the 18 clones revealed inserts of different lengths derived from the same mRNA, the longest insert consisting of 2,126 bp. Computer analysis of the deduced amino acid sequence of the ORF showed >90% identity to the sequences of chloroplast hsp70s and predicted a deficiency of ≈300 bp at the 5′ end. The region encoding the N terminus of the protein was cloned by PCR with an antisense primer covering bases 378–395 of the final full-length clone (Fig. 3) and a sense primer corresponding to the T3-RNA-polymerase promoter in the vector. The amplified DNA separated into three bands with a length of 400, 350, or 300 bp and was subcloned and sequenced. Eight clones with 392, 378, 376, 324, 323, 321, 317, or 316 bp contained the required overlap of the antisense primer and the 5′ end of the existing 2,126-bp clone (i.e., nucleotides 304–395 in the final full-length clone). The 5′ end extended DNA clones fell into two groups: The three longer ones contained two potential ATG translation start codons in frame and the longest one a stop codon 24 bp upstream of the first start codon (Fig. 3). The five shorter clones contained only the second ATG codon; their 5′ ends being located 24, 25, 29, 31, and 32 bp upstream from this second start codon. Two full-length clones encoding the mature protein with the long and short presequence, respectively, were constructed with gene splicing by overlap extension (13).
Figure 3.

Nucleotide sequence of 8 hsp70 cDNA clones and deduced amino acid sequence. The putative N-terminal presequence is in bold letters, the PTS2 signal boxed. The cleavage site is suggested from homology to the N terminus of bacterial hsp70 proteins lacking a presequence. The sequenced internal peptide is underlined. The two start codons within the presequence and the stop codons in the 5′- and 3′-untranslated regions are in bold letters. Arrows indicate 5′ ends of cDNA clones.
Construction of Hybrid Genes Encoding Different hsp70-Presequences Fused to gMDH as a Reporter Protein.
Hybrid genes encoding gMDH fused to two wild-type versions of the hsp70 presequence and one containing a mutation in the PTS2 (−38RT → DD), were generated with gene splicing by overlap extension (13, 14): The long version of the hsp70-presequence (coded by bases 37–249; Fig. 3) or the short version of the hsp70-presequence (coded by bases 97–249; Fig. 3) in its wild-type or mutated form were combined with the mature subunit of gMDH (encoded in bases 181–1334; see figure 1 in ref. 15). Since the cleavage site for the presequence after amino acid 67 is not proven, we added the code for the four N-terminal amino acids of the mature hsp70 subunit (encoded in bases 238–249; Fig. 3) between the presequence and the gMDH polypeptide. The hybrid genes, pre-hsp70long::gMDH and pre-hsp70short::gMDH and pre-hsp70short(−38RT → DD)::gMDH, were inserted as HindIII-SalI fragments in pGEMEX-1 (Promega) under the control of the Sp6 promoter and in the H. polymorpha expression vector pHIPX4 (14) under the control of the strong methanol inducible alcohol oxidase promoter.
Isolation of Proplastids.
Isolation of proplastids was adapted from the procedure for etioplasts (16). Cotyledons were homogenized in 500 mM sucrose, 30 mM Tricine, 1 mM EDTA, 1 mM MgCl2, 0.1% BSA, pH 7.2 (isolation buffer), with five short (3–5 s) bursts in a Waring blender. The homogenate was filtered through Miracloth (Calbiochem) and centrifuged for 5 min at 1,500 × g to remove cell debris. The supernatant was centrifuged for 10 min at 5,000 × g. The organellar pellets were resuspended in isolation buffer and pipetted onto Percoll gradients [7 ml 80% (vol/vol) Percoll and 28 ml 30% (vol/vol) Percoll in isolation buffer without BSA]. The gradients were centrifuged for 1 h at 10,000 × g. After centrifugation two organellar bands were evident. The lower band representing intact proplastids was collected and washed in 500 mM sucrose, 50 mM Hepes, pH 8.0. The purified proplastids were used for protein import studies (17). The purity and integrity of the proplastids were confirmed by the presence of the marker enzyme NADP+-linked glyceraldehyde-3-phosphate dehydrogenase (18) and the absence of isocitrate lyase (marker for glyoxysomes; ref. 19) and fumarase (marker for mitochondria; ref. 20).
In Vitro Import into Isolated Proplastids.
The two fusion genes pre-hsp70long::gMDH or pre-hsp70short::gMDH, cloned in pGEMEX-1, were in vitro transcribed using Sp6-RNA-polymerase and translated in the reticulocyte lysate system (Promega) in the presence of 35S-methionine (DuPont/NEN). The precursor of gMDH (pre-gMDH; ref. 15) was used as a control. For import experiments (16) 1 μl translated proteins was diluted 1:10 with import buffer (17) and incubated with 20 μl proplastids at 25°C for 5–15 min. To check uptake of the labeled proteins by the organelles, the suspension was treated with trypsin (10 μg ml−1). The digestion was terminated by adding loading buffer for SDS/PAGE and incubation for 5 min at 100°C. The reaction was analyzed by SDS/PAGE and fluorography.
Electron Microscopy.
Cotyledons from 3-day-old watermelon seedlings (cut into 0.5 mm sections), glyoxysomes isolated on a Percoll gradient (21) and H. polymorpha cells were fixed and embedded in Lowicryl K4M (Polysciences) as described (7). Thin sections were cut with a diamond knife and picked up on formvar-coated 100-mesh grids. Immunocytochemistry was performed using antibodies against hsp70 (Sigma H5147) and gMDH (10) as primary antibodies and 15 nm gold conjugated goat anti-mouse (monoclonal anti-hsp70 antibody) and 5 nm gold conjugated goat anti-rabbit (polyclonal anti-gMDH antibody) as secondary antibodies.
RESULTS AND DISCUSSION
Identification and Purification of hsp70s from Watermelon Cotyledons.
In germinating watermelon seeds lipid mobilization proceeds actively in the cotyledons and hence, they contain many glyoxysomes. The a-hsp70 antibody recognized in Western blots of protein extracts from cotyledons two distinct bands of 70- and 72-kDa (Fig. 1B, lane 1). Double-ImmunoGold-labeling with a-hsp70- (secondary antibody conjugated to 15 nm gold) and a-gMDH-antibodies (secondary antibody conjugated to 5 nm gold) on ultrathin section of the cotyledons (Fig. 2 A and B) and of isolated glyoxysomes (Fig. 2C) identified hsp70s in the matrix of glyoxysomes and proplastids. A low magnification of a cell showing the overall labeling pattern (Fig. 2A) demonstrates hsp70 in plastids and in glyoxysomes (arrows pointing to 15 nm gold particles) whereas hsp70 labeling was insignificant in other parts of the cell. Labeling with a-gMDH antibodies revealed gMDH only within glyoxysomes. Fig. 2B shows a magnification of a glyoxysome in the cytoplasm and Fig. 2C an isolated glyoxysome: in both cases hsp70 (15 nm gold particles) and gMDH (5 nm gold particles) are visualized indicating the presence of a hsp70 in the glyoxysomal matrix.
Figure 1.
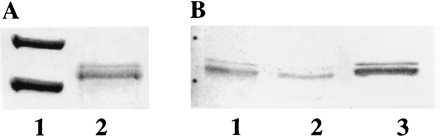
Affinity chromatography with denatured gMDH of hsp70s in protein extracts of watermelon cotyledons. (A) SDS/PAGE and Coomassie blue staining; (B) SDS/PAGE and decoration with a-hsp70 antibodies. A1, molecular mass markers 97 and 66 kDa; A2, Mg-ATP eluate; B1, crude protein extract; B2, run through; B3, Mg-ATP eluate.
Figure 2.
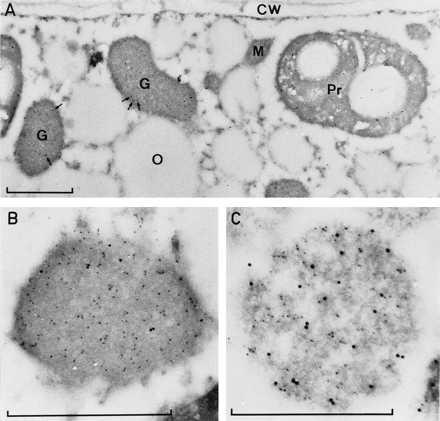
Electron micrographs of aldehyde-fixed cells from 3-day-old watermelon cotyledons (A and B) or isolated glyoxysomes (C). The thin sections were treated with a-hsp70 antibodies and these visualized by 15 nm gold-conjugated goat anti-mouse antibodies; they were at the same time treated with a-gMDH antibodies that were visualized with 5 nm gold-conjugated goat anti-rabbit antibodies. Arrows indicate 15 nm gold particles in glyoxysomes (A). G, glyoxysome; Pr, proplastid; M, mitochondria; O, oil body; CW, cell wall. Bars = 0.5 μm.
Purification of the two hsp70 proteins from the crude extracts was therefore attempted with an affinity procedure, which is based on the affinity of hsp70s for unfolded proteins and their ATP-dependent dissociation of the formed complex (9). Purified watermelon gMDH was coupled to cyanogen bromide-activated Sepharose 4B and two proteins were isolated with molecular masses of 70 and 72 kDa (Fig. 1A, lane 2). These were recognized by the a-hsp70 antibody (Fig. 1B, lane 3). As the N-termini of both proteins were blocked, internal peptides were generated and sequenced. The peptide obtained from the 72-kDa band and subsequently located to position 297–309 of Fig. 3 (underlined) was highly homologous to plastid hsp70s from Pisum sativum (accesion no. L032999; ref. 23) and Cucumis sativus (accession no. X73961). Two different peptides generated from the 70-kDa protein (sequence 1: EIAEAYLGSTVK; sequence 2: DAGVIAGLNVMRIINEPT) were almost 100% identical to cytosolic plant hsp70s and also significantly homologous to cytosolic counterparts from yeast and mammals.
Isolation of cDNA Clones Encoding the 72-kDa hsp70.
As detailed in Materials and Methods, two versions of cDNAs encoding the same 72-kDa hsp were obtained. The three longer cDNAs encoded a precursor with an N-terminal presequence of 67 amino acids, whereas the five shorter cDNAs encoded a protein with a presequence of 47 amino acids, starting at the 2. methionine codon present in the longer version of the sequence (Fig. 3). Sequence comparisons of the derived translation product revealed high identity to hsp70s of the chloroplast stroma of C. sativus (94% identity; accession no. X73961) or P. sativum (91%; accession no. L032999 (22) and algae (64–70%). The identity with other prokaryotic or eukaryotic hsp70s was much lower (<60%). The determined peptide sequence of the 72-kDa protein (see Fig. 1) was present in the translation product of the isolated cDNA clone (Fig. 3, underlined), while those from the 70-kDa protein were absent.
The long version of the presequence showed high homology in length and composition to presequences of plastid hsp70s from C. sativus and P. sativum and therefore might serve as a targeting sequence for plastids. In both presequences a motif with the characteristics of the N-terminal PTS2 was found (Fig. 3, boxed). This motif (RT-X5-KL) is similar to the consensus sequence of the PTS2 signal (RL/I/Q-X5-H/QL) as it is present, for example, in the presequence of the watermelon gMDH (14).
The Long Version of the hsp70 Presequence, But Not the Shorter One, Targets a Reporter Protein to Plastids.
We used an in vitro import assay with isolated intact proplastids from watermelon cotyledons to elucidate the targeting information of the long and the short versions of the hsp70 presequence (Fig. 4). Hybrid proteins were used, comprising the short or long version of the hsp70 presequence fused to gMDH as a reporter protein. As a control, in vitro translated pre-gMDH was used. Incubation of in vitro translated pre-hsp70long::gMDH with isolated proplastids revealed a processing intermediate and the processed gMDH (Fig. 4, lane 2). The precursor and the processing intermediate were sensitive to treatment of the organelles with trypsin, whereas the mature gMDH was protected against protease action (Fig. 4, lane 3). In controls, in which plastids were omitted, processing was not observed (Fig. 4, lane 1). Processing of pre-hsp70short::gMDH was not observed after incubation with plastids (Fig. 4, lane 8), suggesting that the short version of the hsp70 presequence contains no targeting information for plastids. Also, no protease protection was achieved (Fig. 4, lane 9). The same holds true for pre-gMDH (Fig. 4, lanes 4–6). Taken together, the long version of the hsp70 presequence contains the targeting information to guide a reporter protein into proplastids. In the short version the plastid targeting information is absent.
Figure 4.
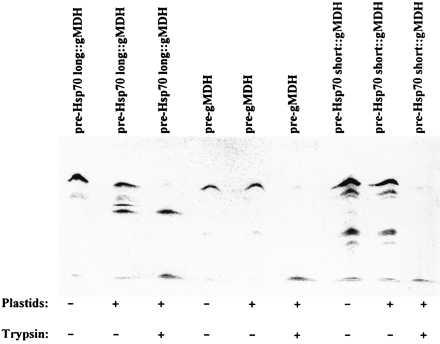
Translocation and processing of pre-hsp70 long::gMDH by isolated proplastids from 3-day-old watermelon cotyledons and protection of the imported gMDH from proteolytic degradation by trypsin (lane 1–3). The 35S-labeled in vitro translation product was incubated for 15 min at 25°C with isolated proplastids (lanes 2 and 3) or with import medium alone (lane 1). Subsequently the import assay was treated with trypsin (10 μg ml−1; lane 3) or with buffer as a control for 5 min on ice (lanes 1 and 2). The same assay was carried out with pre-hsp70short::gMDH (lanes 7–9) and pre-gMDH (lanes 4–6) as a control. The proteins were analyzed by SDS/PAGE and fluorography.
The PTS2 Signal in the Short Version of the hsp70 Presequence is able to direct a Reporter Protein to Peroxisomes.
To test whether the short version of the hsp70 presequence contains a functional PTS2, we carried out in vivo targeting studies into peroxisomes using the yeast H. polymorpha as host. This approach was chosen because the current microbody purification methods from watermelon cotyledons invariably lead to intact glyoxysomes that are contaminated with proplastids. Such a preparation is suitable for immunocytochemistry (Fig. 2), but makes dual localization studies by in vitro import impossible. Pure glyoxysomes can be obtained from a discontinuous sucrose gradient (at the interface between 50 and 57% sucrose); but they are not intact and therefore not reliable as an in vitro import systems.
We have developed the methylotrophic yeast H. polymorpha as a heterologous import system, using mature watermelon gMDH as reporter protein (14): The presequence of gMDH containing a PTS2 in the form of 10RI-X5-17HL is essential and sufficient for peroxisomal targeting. Substitutions of 10RI into DD or 17HL into DE destroyed the topogenic information. Control experiments revealed that mature gMDH, lacking any presequence, was localized to the cytosol. In this yeast the PTS2 import machinery can be induced by specific organic N-sources, e.g., primary amines (7). This has opened a way to analyze the microbody targeting information of watermelon (pre)sequences in vivo. Since the a-hsp70 antibodies cross-reacted with hsp70s from H. polymorpha, we analyzed fusion proteins comprising the short (wild-type and mutated form) or the long watermelon hsp70 presequence fused to gMDH.
Immunocytochemical experiments using a-gMDH antibodies were performed on transformants expressing pre-hsp70 short::gMDH and grown on (NH4)2SO4 or methylamine as N-source (Fig. 5 A and B). They revealed that predominantly mitochondria were labeled; in addition, significant labeling was observed on peroxisomes. A slight induction of the PTS2 import pathway by methylamine could be observed. A stronger enhancement might have been prevented, because the PTS2 signal in the watermelon hsp70 protein (RT-X5-KL) cannot be efficiently recognized by the H. polymorpha PTS2 receptor that is adapted to RL-X5-QA as it is present in H. polymorpha amino oxidase (23). However, both immunolocalizations suggest that the import of the hybrid protein was indeed mediated by the PTS2 import pathway. In a further attempt to establish whether the putative PTS2 signal in the short version of the presequence was functional, we mutated the RT-X5-KL motif into DD-X5-KL thereby destroying the topogenic information. The a-gMDH specific labeling was now confined to mitochondria; peroxisomes were never labeled (Fig. 5C). In cells expressing pre-hsp70long::gMDH the protein was targeted completely to mitochondria; the long version of the hsp70 presequence achieved no peroxisomal sorting (Fig. 5D). Targeting to yeast mitochondria by a plastid presequence has been observed (24). The cleavable presequences of imported chloroplast and mitochondrial proteins have several features in common (25), and yeast seemed to direct proteins lacking a correct destination into mitochondria. Even in case of correct peroxisomal targeting as it is observed for pre-hsp70short::gMDH, mitochondria are likely to be labeled due to overexpression under the control of the strong alcohol oxidase promoter and thus an overload of the peroxisomal import apparatus.
Figure 5.
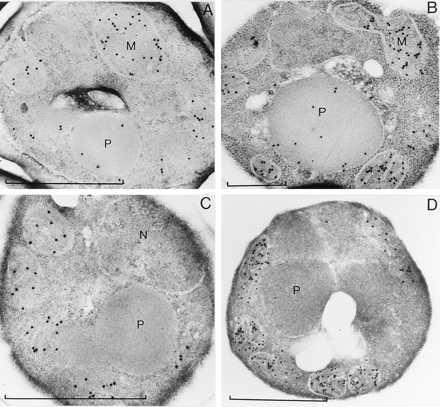
Electron micrographs of aldehyde fixed H. polymorpha transformants, labeled with a-gMDH antibodies and gold-coupled goat anti-rabbit antibodies. (A and B) Cells expressing pre-hsp70short::gMDH, grown on (NH4)2SO4 (A) or methylamine (B) as N-source; peroxisomes are labeled. (C and D) Peroxisomes are not labeled in cells grown on methylamine and expressing pre-hsp70short(RT → DD)::gMDH (C) or pre-hsp70long::gMDH (D). P, peroxisome; M, mitochondrion; N, nucleus. Bars = 1 μm.
We conclude that the short version of the hsp70 presequence contains a functional PTS2 motif. This signal is not recognized when presented to the peroxisomal import apparatus in the context of the long version of the hsp70 presequence. In transformants expressing hybrid proteins without a functional peroxisomal targeting signal, the gMDH is mistargeted to mitochondria.
Further support for this view was obtained from biochemical data. Yeasts have no peroxisomal higher molecular weight precursor proteins and no peroxisomal processing protease. PTS2-containing presequences such as the watermelon pre-gMDH were not cleaved during translocation into H. polymorpha peroxisomes, in contrast to watermelon mitochondrial precursors that were normally processed (14). An intact peroxisomal presequence is another indication for translocation into yeast peroxisomes, where it is protected from processing. We therefore prepared crude protein extracts from identically grown transformants expressing pre-hsp70short::gMDH (wild-type and mutated form) and pre-hsp70long::gMDH and performed a Western blot analysis using a-gMDH antibodies (Fig. 6). Protein extracts from cells expressing pre-gMDH or gMDH were used as a control. In protein extracts prepared from transformants expressing pre-hsp70short::gMDH and grown on (NH4)2SO4 or methylamine as N-source (Fig. 6, lanes 1 and 2) two protein bands were found. The upper band represents the full-length pre-hsp70short::gMDH hybrid protein. Based on its location in the gel, this protein is slightly larger than pre-gMDH (Fig. 6, lanes 4 and 8), as might be predicted from the length of the respective presequences (37 amino acids in case of pre-gMDH, 51 amino acids in case of the short hsp70 presequence fused to mature gMDH). The lower band in the blot represents the processed form of the gMDH protein; it is slightly larger than gMDH (Fig. 5, lanes 3 and 7), since four amino acids of the mature hsp70 protein were included N-terminal to gMDH to preserve the putative hsp70 cleavage site. The most likely explanation for our results is that part of the pre-hsp70short::gMDH hybrid protein is targeted to peroxisomes, but not processed. The majority of the protein, however, is targeted to mitochondria and processed. In Western blots prepared from protein extracts of cells expressing either pre-hsp70short(RT → DD)::gMDH that exhibits the destroyed PTS2-signal (Fig. 6, lane 5) or pre-hsp70long::gMDH where the PTS2 in the context of the longer presequence is not functional (Fig. 6, lane 6), the full-length precursor was not detectable. Instead the processed gMDH band and an intermediate form were found. These results suggest that import of such hybrid gMDH proteins into peroxisomes was effectively prevented, while sorting and processing in mitochondria took place.
Figure 6.
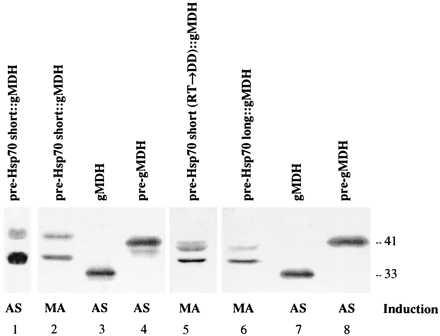
Western blots of protein extracts of transformed cells of H. polymorpha synthesizing hybrid proteins, which combine the mature subunit of gMDH with different hsp70 presequences: the short wild-type version (lanes 1 and 2), the short version with the mutant RT → DD PTS2 (lane 5) and the long version (lane 6). The cells are grown in the presence of (NH4)2SO4 (AS) or methylamine (MA) as N-source. In each lane 40 μg of protein was loaded. The blot was decorated with a-gMDH antibodies. Protein extracts of transformed H. polymorpha cells synthesizing watermelon pre-gMDH (lanes 4 and 8) or gMDH (lanes 3 and 7) provide molecular mass markers (in kDa).
Perspectives.
Several examples exist in which a single gene encodes a protein that is targeted to two distinct compartments (reviewed in ref. 27). In most cases, such a protein is located in an organelle and the cytosol. However, few other examples are known, in which a protein is targeted to two different organelles. Such dual locations were found for marmoset and rat alanine:glyoxylate aminotransferase (28) and for carnithine acetyltransferase from Saccharomyces cerevisiae (29) and Candida tropicalis (30); all of them are sorted to both mitochondria and peroxisomes. The alanine:glyoxylate aminotransferase and carnithine acetyltransferase genes encode N-terminal mitochondrial targeting signals and C-terminal peroxisomal targeting signal 1. Two transcripts are synthesized from the same gene. Polypeptides encoded by the longer transcripts, which include the mitochondrial targeting signal in the ORF, are targeted to the mitochondria, whereas those encoded by the short transcripts, in which the mitochondrial targeting signal is excluded from the ORF, are targeted to the peroxisomes.
The identification of a 72-kDa hsp70 in the glyoxysomal matrix permits to embark on a study of its function in glyoxysomes and peroxisomes of plants. Since the hsp70 is targeted to both plastids and glyoxysomes, it may participate in the ATP-dependent interaction of imported precursor molecules to drive protein translocation across the organellar membrane in the two organelles.
Acknowledgments
We thank Ineke Keizer for expert assistance in electron microscopy. This work was supported by a grant from the Deutsche Forschungsgemeinschaft.
ABBREVIATIONS
- gMDH
glyoxysomal malate dehydrogenase
- hsp70
70-kDa heat shock protein
- PTS2
peroxisomal targeting signal 2
Footnotes
Data deposition: The sequence reported in this paper has been deposited in the GenBank database (accession no. U92815).
References
- 1.Ryan K R, Jensen R E. Cell. 1995;83:517–519. doi: 10.1016/0092-8674(95)90089-6. [DOI] [PubMed] [Google Scholar]
- 2.Gray J C, Row P E. Trends Cell Biol. 1995;5:243–247. doi: 10.1016/s0962-8924(00)89018-2. [DOI] [PubMed] [Google Scholar]
- 3.Luebeck J, Heins L, Soll J. Physiol Plant. 1997;100:53–64. [Google Scholar]
- 4.Wendland M, Subramani S. J Cell Biol. 1993;120:675–685. doi: 10.1083/jcb.120.3.675. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Walton P A, Wendland M, Subramani S, Rachubinski R A, Welch W J. J Cell Biol. 1994;125:1037–1046. doi: 10.1083/jcb.125.5.1037. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.McNew J A, Goodman J M. J Cell Biol. 1994;127:1245–1257. doi: 10.1083/jcb.127.5.1245. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Faber K N, Haima P, Gietl C, Harder W, AB, G, Veenhuis M. Proc Natl Acad Sci USA. 1994;91:12985–12989. doi: 10.1073/pnas.91.26.12985. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Gietl C, Hock B. Plant Physiol. 1982;70:483–487. doi: 10.1104/pp.70.2.483. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Evers M E, Huhse B, Titorenko V I, Kunau W H, Hartl F U, Harder W, Veenhuis M. FEBS Lett. 1993;321:32–36. doi: 10.1016/0014-5793(93)80615-2. [DOI] [PubMed] [Google Scholar]
- 10.Gietl C, Seidel C, Svendson I. Biochim Biophys Acta. 1996;1274:48–58. doi: 10.1016/0005-2728(96)00009-6. [DOI] [PubMed] [Google Scholar]
- 11.Eckerskorn C, Lottspeich F. Chromatographia. 1989;28:92–94. [Google Scholar]
- 12.Faber K N, Haima P, Harder W, Veenhuis M, AB G. Curr Genet. 1994;25:305–310. doi: 10.1007/BF00351482. [DOI] [PubMed] [Google Scholar]
- 13.Horton R M, Hunt H D, Ho S N, Pullen J K, Pease L R. Gene. 1989;77:61–68. doi: 10.1016/0378-1119(89)90359-4. [DOI] [PubMed] [Google Scholar]
- 14.Gietl C, Faber K N, van der Klei I J, Veenhuis M. Proc Natl Acad Sci USA. 1994;91:3151–3155. doi: 10.1073/pnas.91.8.3151. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Gietl C. Proc Natl Acad Sci USA. 1990;87:5773–5777. doi: 10.1073/pnas.87.15.5773. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Soll J, Wanner G, Henkelmann G, Röper U, Schulze M. In: Regulation of Chloroplast Differentiation. Akoyunoglou G, editor. New York: Liss; 1986. pp. 229–234. [Google Scholar]
- 17.Waegemann K, Soll J. Methods Cell Biol. 1995;50:255–267. doi: 10.1016/s0091-679x(08)61035-3. [DOI] [PubMed] [Google Scholar]
- 18.Lendzian K J, Ziegler H. Biochem Physiol Pflanzen. 1978;173:500–504. [Google Scholar]
- 19.Dixon G H, Kornberg H L. Biochem J. 1959;72:3P. [Google Scholar]
- 20.Racker E. Biochim Biophys Acta. 1950;4:211–214. doi: 10.1016/0006-3002(50)90026-6. [DOI] [PubMed] [Google Scholar]
- 21.Kato A, Hayashi M, Mori H, Nishimura M. Plant Mol Biol. 1995;27:377–390. doi: 10.1007/BF00020191. [DOI] [PubMed] [Google Scholar]
- 22.Marshall J S, Keegstra K. Plant Physiol. 1992;100:1048–1054. doi: 10.1104/pp.100.2.1048. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Gietl C. Physiol Plant. 1996;97:599–608. [Google Scholar]
- 24.Hurt E C, Soltanifar N, Goldschmidt-Clermont M, Rochaix J D, Schatz G. EMBO J. 1986;5:1343–1350. doi: 10.1002/j.1460-2075.1986.tb04365.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Von Heijne G, Stepphuhn J, Herrmann R. Eur J Biochem. 1989;180:535–545. doi: 10.1111/j.1432-1033.1989.tb14679.x. [DOI] [PubMed] [Google Scholar]
- 26.Corpas F J, Trelease R N. Eur J Cell Biol. 1997;73:49–57. [PubMed] [Google Scholar]
- 27.Danpure C J. Trends Cell Biol. 1995;5:230–238. doi: 10.1016/s0962-8924(00)89016-9. [DOI] [PubMed] [Google Scholar]
- 28.Motley A, Lumb M J, Oatey P B, Jennings P R, De Zoysa P A, Wanders R J A, Tabak H F, Danpure C J. J Cell Biol. 1995;131:95–109. doi: 10.1083/jcb.131.1.95. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Elgersma Y, van Roermund C W T, Wanders R J A, Tabak H F. EMBO J. 1995;14:3472–3479. doi: 10.1002/j.1460-2075.1995.tb07353.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Kawachi H, Atomi H, Ueda M, Tanaka A. Eur J Biochem. 1996;238:845–852. doi: 10.1111/j.1432-1033.1996.0845w.x. [DOI] [PubMed] [Google Scholar]