

Abstract
Iron (Fe) is an essential metal ion, required for basic cellular processes such as respiration, photosynthesis and cell division. Therefore, Fe has to be stored and distributed to several organelles to fulfill its roles. The molecular basis of Fe distribution is poorly understood. In this context, elemental imaging approaches are becoming essential for a better understanding of metal homeostasis in plants. Recently, several genes have been involved in Fe storage (VIT1) and remobilization (NRAMP3 and NRAMP4) in the seed of Arabidopsis, mostly with the help of sophisticated imaging techniques. We have adapted an histochemical procedure to detect Fe in plant tissues, based on Perls staining coupled to diaminobenzidine (DAB) intensification. The Perls/DAB technique, quick and inexpensive, was shown to be specific for Fe and highly sensitive. We have applied this procedure to Arabidopsis embryos and shown that Fe is stored in the vacuoles of a specific cell layer surrounding the pro-vascular system, the endodermis. Our results have revealed a new role for the endodermis in Fe storage in the embryo and established the Perls/DAB technique as a powerful tool to detect Fe in plant tissues and cells.
Key words: iron, vacuole, Arabidopsis, endodermis, embryo
Iron (Fe) is a very important essential metal for all living organisms. The function of Fe in biological processes relies on its ability to exist in two redox states (ferrous and ferric iron). Consequently, Fe is crucial for metabolic reactions of respiration and photosynthesis. Although at the whole plant level the molecular mechanisms of Fe acquisition and storage are now well documented, the control of Fe distribution at organ, cell and sub-cellular levels is extremely poorly understood and represents an important stake. Recent discoveries on Fe storage and remobilization in the embryo have unequivocally shown that genetic approaches are not sufficient to unravel the function of genes, unless coupled to high-resolution metal imaging. The use of energy dispersive X-ray spectroscopy (EDX) and inelastically scattered electrons (ESI) on electron micrographs clearly showed that in Arabidopsis seeds Fe is accumulated in vacuoles and remobilized during germination.1 Likewise, the analysis of Arabidopsis seeds by XRF and tomography has provided three-dimensional mapping of Fe that has showed that VIT1 (for Vacuolar Iron Transporter) is required for proper allocation of Fe in the vascular tissue of the embryo, a distribution that is altered in a vit1-1 mutant.2 Noteworthy is the fact that these alterations of Fe distribution do not induce a change in total Fe content and thus would not have been detected by measurements of total Fe concentration. There is therefore an increasing interest for imaging techniques that are becoming a must-have to understand the function of metal transporters in plants. Metal imaging technologies are mostly based on X ray absorption/emission/fluorescence, often requiring expensive and rare equipments (synchrotron for instance). Taking advantage on the high reactivity of metal ions for organic ligands, several reagents have been used as chromophores for histochemical staining of metals. Among those, potassium ferrocyanide, also known as Perls reagent, was used since the late XIXth century to produce the Prussian blue, after reaction with ferric iron. The Perls reagent has been widely used to stain Fe in tissues, but only occasionally in plants, due to its low sensitivity and poor penetration in hydrophobic tissues. Nevertheless, it is possible to increase the sensitivity of the staining by secondary reactions with diaminobenzidine (DAB) and hydrogen peroxide (H2O2). Indeed, since the Fe-Perls complex is redox active, the addition of DAB and H2O2 triggers the oxidative polymerization of DAB, producing brown pigments.3 This reaction is the basis of the intensification of the Perls staining (Perls/DAB from now on). To adapt this procedure to plants, we have first established that the staining was specific for Fe and did not practically cross-react with other metal ions. In doing so, we also showed that Perls/DAB could stain both FeII and FeIII. We have chosen Arabidopsis seeds as model, in order to compare our Fe staining procedure with the imaging already available by XRF.2 Compared to Perls stain alone, the Perls/DAB protocol appeared to be much more sensitive. Iron appeared to be concentrated around the provascular system of mature Arabidopsis embryos.
Staining of the vit1-1 mutant showed a modified pattern compatible with the available μXRF-tomography data, thus clearly establishing that the staining procedure is specific for Fe in vivo and represents a new, quick and simple tool to detect Fe in plant samples.4 Thin sections of Perls/DAB embryos uncovered that, in mature embryos, Fe was concentrated in a single cell layer, apparently corresponding to the endodermis. This observation was further confirmed by the analysis of longitudinal sections of the radicle-hypocotyl region. In this particular zone, a second periclinal division occurs, giving rise to a cortex cell layer and the endodermis,5 the latter one alone being intensively stained with Perls/DAB. Other developmental mutants were used, such as for example the vein patterning mutant SCARFACE (scf),6 which presents Fe staining in cotyledons as small segments corresponding to discontinuous veins characteristic of the scf mutant (Fig. 1). Thus, the Perls/DAB protocol represents a very useful tool not only to study Fe homeostasis but also in the field of developmental research, as a marker of endodermis and provascular system of the embryo. Finally, the Perls/DAB procedure can be greatly improved by staining directly the histological thin sections instead of whole embryos, thereby (i) increasing tremendously the resolution and (ii) solving the problem of low penetration of the dyes in hydrophobic plant samples. This modification enabled us to show that in endodermal cells Fe is actually located in vacuoles. Remobilization of the vacuolar pool of iron by AtNRAMP3 and AtNRAMP4 is crucial during germination.1 Since we found that Fe is blocked in the endodermis of the nramp3nramp4 mutant,4 we can now propose that in mature embryos Fe is mainly stored in the vacuoles of the endodermis.
Figure 1.
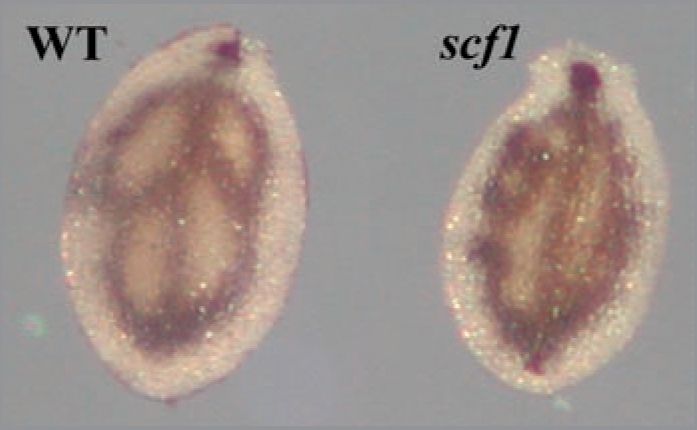
Iron distribution in cotyledons. Wild-type and scf1 dry seed embryos were dissected and stained with Perls/DA B according to Roschzttardtz et al.4
In conclusion, we have adapted an histochemical staining procedure, easy to set up and inexpensive, that fills the gap between the Perls reagent and X ray-based elemental imaging. The resolution, at the sub-cellular scale, makes it a valuable tool to investigate the distribution of Fe in plant tissues without employing electron microscopy or synchrotron X ray fluorescence. Furthermore, the possibility of direct staining on histological sections makes this technique applicable to virtually any plant material, after fixation and embedding in resin.
Acknowledgements
We wish to thank Leslie Sieburth for the gift of SCARFACE seeds. This work was supported by the Centre National de la Recherche Scientifique (CNRS), l’Institut National de la Recherche Agronomique (INRA). The work of H.R. is supported by a post-doctoral fellowship from the Agence Nationale pour la Recherche (ANR program 07-3-18-8-87 DISTRIMET).
Footnotes
Previously published online: www.landesbioscience.com/journals/psb/article/10159
References
- 1.Lanquar V, Lelievre F, Bolte S, Hames C, Alcon C, Neumann D, et al. Mobilization of vacuolar iron by AtNRAMP3 and AtNRAMP4 is essential for seed germination on low iron. EMBO J. 2005;24:4041–4051. doi: 10.1038/sj.emboj.7600864. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Kim SA, Punshon T, Lanzirotti A, Li L, Alonso JM, Ecker JR, et al. Localization of iron in Arabidopsis seed requires the vacuolar membrane transporter VIT1. Science. 2006;314:1295–1298. doi: 10.1126/science.1132563. [DOI] [PubMed] [Google Scholar]
- 3.Nguyen-Legros J, Bizot J, Bolesse M, Pulicani JP. “Diaminobenzidine black” as a new histochemical demonstration of exogenous iron (author’s transl) Histochemistry. 1980;66:239–244. doi: 10.1007/BF00495737. [DOI] [PubMed] [Google Scholar]
- 4.Roschzttardtz H, Conéjéro G, Curie C, Mari S. Identification of the endodermal vacuole as the iron storage compartment in the Arabidopsis embryo. Plant Physiol. 2009;151:1329–1338. doi: 10.1104/pp.109.144444. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Scheres B, Dilaurenzio L, Willemsen V, Hauser MT, Janmaat K, Weisbeek P, Benfey PN. Mutations affecting the radial organization of the Arabidopsis root display specific defects throughout the embryonic axis. Development. 1995;121:53–62. [Google Scholar]
- 6.Deyholos MK, Cordner G, Beebe D, Sieburth LE. The SCARFACE gene is required for cotyledon and leaf vein patterning. Development. 2000;127:3205–3213. doi: 10.1242/dev.127.15.3205. [DOI] [PubMed] [Google Scholar]