

Abstract
In response to infection or effective vaccination, naive antigen-specific CD8+ T cells undergo a dramatic highly orchestrated activation process. Initial encounter with an appropriately activated antigen-presenting cell leads to blastogenesis and an exponential increase in antigen-specific CD8+ T cell numbers. Simultaneously, a dynamic differentiation process occurs, resulting in formation of both primary effector and long-lived memory cells. Current findings have emphasized the heterogeneity of effector and memory cell populations with the description of multiple cellular subsets based on phenotype, function, and anatomic location. Yet, only recently have we begun to dissect the underlying factors mediating the temporal control of the development of distinct effector and memory CD8+ T cell sublineages. In this review we will focus on the requirements for mounting an effective CD8+ T cell response and highlight the elements regulating the differentiation of effector and memory subsets.
Keywords: memory, CD8 T cell, differentiation
Introduction
The practical concept of immunologic memory has existed for many centuries, from Thucydides’ (ca. 430 BC) observation that surviving plague victims did not secondarily contract the disease to the use of variolation to prevent smallpox by the ancient Chinese (ca. 1000 AD) and Edward Jenner’s observation that milk maids exposed to cowpox rarely acquired smallpox (1796 AD). Jenner’s vaccination scheme led to a scientific explosion in trying to understand the mechanisms behind acquired protection. To date, vaccines have eliminated or minimized many devastating diseases throughout the world. Most of the effective vaccines in use today rely on the generation of neutralizing antibody responses.1 However, for some pathogens, such as malaria and HIV, vaccines aimed at inducing neutralizing antibody responses have been largely unsuccessful.2,3 In addition, although the influenza virus vaccine provides a level of protection to some individuals, it is less than ideal because antigenic drift forces yearly vaccination with predicted emerging serotypes.4 For these infections and others, vaccines geared toward generation of CD8+ T-cell immunity may provide protection.5–8 This is true for two reasons: CD8+ T cells generally react to epitopes derived from proteins with less strain to strain variation9 and the life cycle of certain microbes does not lend itself to inhibition by antibody neutralization, while CD8+ T cells provide sterilizing immunity.10 Thus, CD8+ T cells play a crucial role in the control of infection by intracellular pathogens, such as certain viruses, bacteria, and protozoan parasites. Additionally, CD8+ T cells are able to limit tumor development and growth.11
Only in the past few decades have the cellular and molecular mechanisms behind memory T-cell development begun to be elucidated. Much of that work has been greatly aided by technological advances, such as flow cytometry, knockout and transgenic mice, gene arrays, and peptide: major histocompatibility complex p (MHC) tetrameric reagents as well as the more recent application of laser-based confocal and two-photon microscopy. These techniques have enabled us to quantify, characterize, localize, and isolate antigen-specific T cells throughout the entire immune response. Additionally, broad cell genetic analysis through the use of DNA microarrays and deep sequencing has given us large data sets that are indicative of either effective or ineffective CD8+ T-cell-mediated immune responses12,13 (K.M. Khanna et al., University of Connecticut Health Center, unpublished observation). As such, our understanding of the events leading to the successful generation of T-cell memory has been significantly enhanced in recent years, but much remains unresolved.
Phases of the T cell response
Mature mammals maintain a pool of naive antigen-specific T cells that is capable of responding to a vast array of peptide antigens. Recent studies have elucidated that in laboratory mice there are approximately between 15 and 1000 antigen-specific CD8+ T cells14–16 and 10 and 200 CD4+ T cells17 specific for any given MHC/peptide epitope combination. Although analysis of many more specificities is needed before generalizations can be made, naive T-cell frequencies in this range would enable the host to respond to approximately 106–107 different peptides. However, the total number of antigens to which mice can respond is likely larger as a result of cross-reactive T-cell receptors (TCRs).18,19 Activation of naive T cells is highly dependent on dendritic cells (DCs),20 but other antigen-presenting cells (APCs), such as macrophages, may also be used under certain conditions.21 The initiation of the T-cell response to a pathogen occurs in an exquisitely orchestrated process whereby the antigen is enriched in the T-cell zones of the lymphoid organs via capture and concentration by APCs. This process increases the odds of the rare antigen-specific T cell encountering its cognate antigen presented on MHC molecules expressed by DCs. Initial studies suggested that only a short encounter with antigen was necessary for optimal CD8 T-cell expansion and subsequent memory formation.22,23 However, more recent studies indicate that prolonged interactions with antigen-bearing APCs are necessary for the optimal generation of effector and memory T cells.24–28 The activation of CD8+ T cells is dependent on at least three signals: 1) antigenic stimulation through the TCR; 2) costimulation through molecules such as CD28, CD40, 4-1BB, CD27, ICOS, and/or OX40; and 3) stimulation through receptors for inflammatory cytokines, especially IL-12 and interferon (IFN)-α.29–31 Naive antigen-specific CD8+ T cells receiving all three of these signals will undergo robust expansion, generating a large effector CD8+ T-cell population. In addition to the exponential increase in cell numbers, activation results in the acquisition by the responding cells of several effector functions, such as the expression of cytokines (i.e., IFN-γ and tumor necrosis factor [TNF]-α) and perforin and granzyme molecules, which are necessary for cytolytic activity, and in some cases via nonlytic mechanisms for antiviral defense.32 Furthermore, at this point the armed effector T cells, whether CD8+ or CD4+ T cells, will migrate to virtually all the tissues of the body33,34 on a mission to patrol the body with the goal of removing all infected cells.
After this rapid expansion, the effector cell population undergoes a catastrophic contraction phase whereby approximately 95% of effector cells die by apoptosis, which is mediated largely by the ratio of the expression of survival versus apoptotic factors, such as Bcl-2, Bclx, and Bim and Fas pathways.35–41 Ultimately, the remaining pathogen-specific T cells form the memory T-cell population. These memory CD8+ T cells will then persist long term in the host in an antigen-independent42 and TCR-independent43 but cytokine-dependent manner.44,45 The cytokines responsible for the long-term maintenance of the memory CD8+ T-cell population are members of the γc-cytokine family, with IL-7 or perhaps thymic stromal lymphopoietin (TSLP) enhancing memory cell survival,46–48 while IL-15 is essential for the slow homeostatic turnover of the memory cells.44,45 Several salient features of memory cells enable them to rapidly respond when they are called back to duty. First, memory T cells may persist in relatively large numbers, compared to naive T-cell frequencies, in multiple tissues of the body,33,34 which positions them on the front line near potential points of pathogen entry into nonlymphoid tissues, such as the lung, intestines, and skin. Second, memory T cells are able to rapidly re-express effector molecules, such as cytokines and proteins of the lytic machinery, even prior to cell division in some cases.49,50 The ability to rapidly respond is a result of, at least in part, the hyper-responsive state of memory T cells compared to naive T cells.51–54 Thus, the proper training of naive CD8+ T-cell recruits results in the formation of an operational army of effector and memory CD8+ T cells that can protect the host from the present infection and be poised to win future battles with the same pathogen.
Definition of memory T cell subsets
Earlier studies demonstrated the existence of at least two memory T-cell subpopulations based on the expression of different homing molecules, specifically CD62L (L-selectin) and the receptor for the CCL19 and CCL21 chemokines CCR7.33,34,55,56 CD62L and CCR7 along with the β2 integrin, αxβ2, are required for entry of naive57 and presumably memory T cells to enter lymph nodes (LNs). However, memory T cells may enter a reactive LN in a CD62L- and CCR7-independent manner.58 CCR7 is also important for positioning of lymphocytes and APCs within the spleen59 and appears to be required for exit of cells from nonlymphoid tissues by controlling entry into afferent lymphatics.60,61 Memory T cells expressing both these receptors are termed central memory cells (TCM), while those lacking expression of both molecules are termed effector memory cells (TEM). Differential expression of both CD62L and CCR7 results in memory cell subpopulation localization to different anatomical sites with the TCM largely confined to the LNs, spleen, and blood, while TEM are localized in peripheral nonlymphoid tissues (e.g., lung, liver, intestine), spleen, and blood.33,34 TEM may also be present in LNs perhaps as a result of migration via afferent lymphatics from non-lymphoid tissues. Additionally, central and effector memory T cells exhibit distinct functional abilities,34,56,62 which intriguingly could play a role in the different protective abilities of each subset against future antigen encounters.63,64 Furthermore, tissue-resident CD8+ TEM are important in the maintenance of latent α-herpesvirus infection and undergo rapid expansion following viral reactivation.32,65–67
The origin of TEM versus TCM has been a matter of considerable study and debate. Early work demonstrates that, with increasing time after immunization, the proportion of CD62Lhigh cells increases within the memory population.68 Initially this phenomenon was thought to be a result of re-expression of CD62L by CD62Llow TEM converting to CD62Lhigh TCM, but this appears to be the result of artificially high naive CD8+ T-cell-precursor frequencies.14,69–71 More likely, on the basis of the available evidence, the increasing predominance of CD62Lhigh TCM in the memory population is a result of the increased turnover rate of those cells relative to the CD62Llow TEM, which divide at a slower rate.14,70 However, the strict classification of memory cells based on expression of CD62L and CCR7 alone is unlikely to be all inclusive as further phenotypic heterogeneity within the memory T-cell pool has been observed.72,73 Thus, classifications of memory T cells should also account for specific characteristics of the immune response related to the inciting infectious agent or immunization scheme. For example, following respiratory influenza virus infection subsets of cells in the lung and lung-draining mediastinal LNs express a unique pattern of activation markers as a result of the presence of low levels of persistent antigen.73–76 Furthermore, following γ-herpesvirus infections in both mice and humans, CD8+ T cells responding to either lytic-cycle or latent-cycle proteins have distinct activation profiles resembling TEM and TCM, respectively.77–79 Additionally, the level of antigenic persistence and localization of such persistence has a dramatic impact on the phenotype, functionality, and survival/maintenance of the antigen-specific CD8+ T cell responding to persistent infections.80–85 Thus, classifications of memory cell subsets should be made with the appropriate contextual considerations.
Models describing the origins of memory CD8+ T cells
When, where, and how memory T cells and their associated subsets are formed is an area of intense study. One important fact to consider when generating any model to explain effector and memory CD8+ T-cell differentiation is that a single naive antigen-specific CD8+ T cell of fixed avidity has the potential to form every effector and memory subset observed during the CD8+ T-cell response.86 This finding suggests that after activation the progeny of the initial cell must integrate distinct downstream signals, resulting in generation of heterogeneous populations of effector and memory cells. Although past debate has focused on whether memory T cells can develop without transiting an effector stage,87 recent results88,89 show that effector cells are the source of memory T cells. These studies used reporter mice in which CD8+ or CD4+ T cells exhibiting granzyme B or IFN-γ promoter activity were irreversibly labeled by means of Cre-mediated recombination to identify effector T cells and their offspring.88,89 The results support a linear differentiation model in which naive T cells are activated, expand, and differentiate into effector T cells, after which a small proportion of the effector cells survives to form the long-lived memory population (Fig. 1). Nevertheless, the heterogeneity found in both the effector and memory cell pools must also be accounted for in any differentiation scheme.56,90,91
Figure 1.

Early cell fate determination model of effector and memory CD8+ T-cell differentiation. In this model naive CD8+ T cells become activated and form an early effector CD8+ T-cell population, which is CD127low KLRG1low. Next, three populations of effector cells can be identified by the peak of the CD8+ T-cell response: short-lived effector cells (SLEC) that are CD127low KLRG1high CD62Llow, effector memory cells (TEM) memory precursor effector cells (MPEC) that are CD127high KLRG1low CD62Llow, and central memory cells (TCM) MPEC that are CD127high KLRG1low CD62Lhigh. Over time the SLEC population is lost through apoptosis, while the MPEC population remains long term in the host forming the memory CD8+ T-cell population. Additionally, with time the memory population transitions from being predominately TEM in phenotype to TCM in nature, and this is the result of the increased homeostatic proliferation rate of the TCM population. KLRG1, killer cell lectin-like receptor G1; Ag, antigen; CpG, unmethylated CpG containing oligonucleotide.
Here we propose an early fate determination model to explain the current data related to effector and memory cell differentiation (Fig. 1). After initial activation the early effector CD8+ T-cell population appears to be phenotypically homogeneous, being CD127low KLRG1low CD27high. In keeping with the genetic labeling experiments,88,89 this population of early effector CD8+ T cells will have gained full effector cell functions, expressing the effector molecules granzyme B and IFN-γ, and as a result all the subsequent effector and memory populations will be similarly genetically labeled. As the immune response continues, substantial heterogeneity within the effector cell population can be observed by the peak of the CD8+ T-cell response. The majority of the effector CD8+ T cells present at this time are terminally differentiated effector cells, which are relatively short lived, while a much smaller population of memory precursor effector cells also exist.90–93 Additionally, we have recently observed further heterogeneity within the memory precursor effector cell population, with both CD62Lhigh TCM and CD62Llow TEM precursors being present by 5 days after infection (J.J. Obar et al., manuscript submitted), which fits with our previous data indicating that the TCM and TEM represent fixed cell lineages, at least based on CD62L expression.14,70 However, it remains unknown whether a memory precursor cell upon further stimulation can give rise to short-lived effector cells or if a CD62Lhigh TCM memory precursor can become a CD62Llow TEM memory precursor.
Another controversial topic of research is the question of when and how the heterogeneity within the effector population is induced. The early fate determination model proposed here suggests that the early effector CD8+ T-cell population is homogeneous, but this seems unlikely given that within the next 24–48 h diversity becomes evident in the population. Thus, heterogeneity at the level of the earliest signaling events apparently controls long-term outcomes. Taken to the extreme, the model of Reiner and colleagues suggests that asymmetric division occurring during the first cell division results in proximal and distal daughter cells that have different lineage potential, producing effector and memory lineages, respectively.94 However, this model has not explained how such lineages may be maintained during the ensuing robust cellular division and ongoing acquisition of antigen- and cytokine-driven signals. In this scheme, genetic “imprinting” would occur during the initial T-cell–APC encounter. Thus, it remains possible that heterogeneity exists at a genetic level within the early effector CD8+ T-cell population and that deterministic events continue to occur downstream of initial activation. At this point, an early fate determination model, as described, best fits the available data and largely explains the cellular heterogeneity observed during both the effector and memory stages.
Requirements for generating effector cell heterogeneity
As proposed in the early fate determination model, the effector cell population contains both terminally differentiated effector cells and memory precursors at the peak of the CD8+ T-cell response. Originally, the memory precursor population of effector CD8+ T cells was identified based solely on their retained expression of IL-7Rα.46,90 More recently, such memory precursor cells were shown to lack expression of killer cell lectin-like receptor G1 (KLRG1), while the terminally differentiated effector cells express high levels of KLRG1.91–93 Additionally, CD27 expression remains high on the memory precursor population and decreases on the short-lived effector cell population95 (J.J. Obar & L. Lefrançois, unpublished observation). Recently the factors regulating the differentiation of these two effector cell subpopulations have begun to be elucidated. As previously mentioned, activation of a naive antigen-specific CD8+ T cell is highly dependent on three separate signals: antigenic stimulation through the TCR, costimulation, and stimulation via inflammatory cytokines. Each of these three signals alone or in combination has the potential to direct the responding CD8+ T cells to differentiation toward one cell fate or another: terminal effector cell or memory precursor.
Central to the activation cascade is the interaction of the TCR with cognate antigen presented by the appropriate MHC molecule. It has been demonstrated that the overall strength of TCR stimulation, an accumulation of the magnitude and duration of TCR: pMHC (peptide MHC) interaction, can direct the magnitude of the T-cell response,96–99 but this does not seem to drastically alter the differentiation pattern of the responding CD8+ T cells91 (J.J. Obar et al., manuscript submitted). After TCR engagement, CD8+ T cells upregulate numerous costimulatory molecules. It is well established that such receptors play an important role in the clonal expansion and survival of the responding antigen-specific CD8+ T-cell population,100,101 but much less is known about their impact on effector cell differentiation leading to memory subset development. Although more work is needed, CD27 or 4–1BB costimulation does not appear to dramatically alter the development of short-lived effector CD8+ T cells (J.J. Obar & L. Lefrançois, unpublished observation). One recent study indicates that following Listeria monocytogenes infection OX40 plays a significant role in the differentiation of the memory precursor pool, as OX40−/− CD8+ T cells generated reduced numbers of CD127high KLRG1low cells, which led to poor memory development.102 Given the large number of potential costimulators, the role of each costimulator in influencing effector and memory cell heterogeneity requires substantial further exploration.
The effects of cytokines on expansion and differentiation of effector and memory CD8+ T cells has also been a topic of considerable scrutiny. It is well documented that both type I and type II IFNs as well as IL-12 can act directly on responding CD8+ T cells to enhance their survival and/or expansion.31,103–108 Furthermore, some of these inflammatory cytokines have also been shown to have a dramatic effect on the differentiation of responding antigen-specific CD8+ T cells. For example, low levels of inflammation, such as are present during DC vaccination, shift the balance toward enhanced generation of memory precursors,91,109,110 while high levels of inflammation, such as during infections, lead to the rapid and preferential expansion of terminally differentiated KLRG1high effector cells.91,111,112 Specifically, IL-12 has been shown to influence the differentiation of terminally differentiated short-lived effector cells,91 and this inflammatory event must be directly coupled temporally with antigenic stimulation of the T cell.112 Additionally, IL-2 immune complexes given as a vaccine adjuvant will enforce differentiation of the KLRG1high short-lived effector cell population.93 One inflammatory pathway of note, which does not act directly on the responding antigen-specific CD8+ T cells, is the MyD88 pathway, which is crucial in mediating signals through Toll-like receptors and IL-1 cytokine family members.113 However, this pathway is likely important in CD8+ T-cell differentiation but may act upstream of the CD8+ T cell by directing the activation of the APC through modulation of the costimulatory and inflammatory environment. Thus, we have begun to understand the integration of the signaling pathways mediated by the three classes of activation signals leading to generation of short-lived apparently terminally differentiated effector cells. However, one glaring observation is the absence of the identification of the factors that are important in directing the differentiation of the memory precursor effector cell population and derivative memory subsets.
Recently, an elegant study by Palmer and colleagues demonstrated that the CART domain within the transmembrane region of the TCRβ chain is required for the formation of CD8+ T-cell memory.114 Further, this effect is likely mediated through formation of the immunological synapse and subsequent signaling through protein kinase C (PKC)-θ and nuclear factor (NF)-κB. However, this study did not determine whether memory precursors were ever formed or whether all antigen-specific CD8+ T cells failed to survive the contraction phase. Therefore, this work demonstrates that the type or “strength” of initial signaling received by the responding naive antigen-specific CD8+ T cell may have a drastic effect on the future outcome of the response, but further work is needed to definitively separate memory cell differentiation from cell survival.
One important insight into memory cell differentiation has come from the profiling of transcription factor expression in the different T-cell populations. The transcription factors that have been explored during CD8+ T-cell responses include T-bet, Eomes, Blimp1, Bcl6, and Id2. To date, the most extensively studied transcription factors during an antigen-specific CD8+ T-cell response have been the pair of T-box transcription factors, T-bet and Eomes. Reiner and colleagues have shown that T-bet and Eomes are critical in the regulation of effector and memory cell differentiation, respectively.115 Overexpression of T-bet in effector cells leads to the enhanced generation of short-lived terminal effector cells.91 Furthermore, the transcription factor Blimp1 has been shown to be highly expressed in those cells that also express T-bet.116 This suggests that T-bet and Blimp1 together may direct a transcriptional program that leads to the formation and expansion of the terminally differentiated short-lived effector cell population. Blimp1 activity can be opposed by the transcription factor Bcl6, which is important in directing the differentiation of germinal center B cells into either memory cells or plasma cells.117 Intriguingly, Bcl6 appears to also play a role in the differentiation of memory CD8+ T cells as Bcl6−/− mice have reduced numbers of antigen-specific memory CD8+ T cells, while the opposite is true of Bcl6 transgenic mice.118,119 Thus, by understanding the factors that regulate these pivotal transcription factors, we will garner a better understanding of the complex regulation inherent to the differentiation of distinct effector and memory CD8+ T-cell subsets.
Regulation of L-selectin expression
L-selectin (CD62L) plays a key role, together with CCR7, in enabling naive T cells to enter lymph nodes from the blood through the high endothelial venules.120,121 As such, CD62L and CCR7 expression are tightly regulated to enable the redistribution of effector T cells that downregulate these homing receptors following activation.122–126 Much of what is known about the control of CD62L expression has been determined using in vitro activation systems. In these studies, the regulation of CD62L after TCR engagement occurs in a three-step process.127,128 Initially, within 6–8 h of activation, CD62L is shed from the cell surface by proteolytic cleavage through the action of the protease TACE/Adam17.126 Next, between 24–48 h, CD62L is re-expressed on the cell surface of the activated T cells. Finally, relatively late after initial activation, sell gene activity can be modified by mechanisms that are only now being defined.
Recently, the minimal promoter region of the sell gene was identified and five transcription factors were found to play a role in mediating transcription driven by the sell promoter. These transcription factors are Ets1, Irf1, Klf2, Mzf1, and Sp1.129 Importantly, Klf2 is known to play a key role in regulating CD62L expression in naive T cells and thymocytes.130 Interestingly, recent work has begun to elucidate the factors that not only regulate promoter activity of the sell gene but also regulate activation of the protease TACE/Adam17, which is responsible for CD62L cleavage (Fig. 2). Elegant in vitro studies from Cantrell and colleagues show that the p110δ subunit of phosphatidylinositol-3-kinase (PI[3]K) is critical in not only the modulation of TACE/Adam17 activity but also the genetic regulation of sell promoter activity.131 Signaling through the TCR results in activation of the p110δ subunit of PI(3)K, resulting in subsequent activation of the Erk1/2 pathway. Active Erk1/2 can then phosphorylate TACE/Adam17, promoting translocation to the cell membrane where it can cleave the CD62L protein from the cell surface, resulting in the transient early loss of CD62L. Sustained or permanent genetic regulation of CD62L expression occurs at later time points after activation and appears to be mediated, at least in part, by members of the γc-cytokine family.131 Signaling through the IL-2R complex results in strong activation of the PI(3)K pathway, while signals mediated by the IL-15R complex result in apparently weaker activation of the PI(3)K pathway.132 Furthermore, in vitro studies show that antigen-mediated activation of CD8+ T cells in the presence of IL-2 or IL-15 results in the generation of TEM- or TCM-like effector cells, respectively.131,133,134 Activation of the PI(3)K path-way by γc-cytokines appears to regulate sell gene expression through regulation of mammalian target of rapamycin (mTOR) activity, which subsequently modulates the activity of the transcription factor Klf2. As such, the strong signal through PI(3)K initiated by IL-2 activates mTOR, resulting in inhibition of Klf2. In contrast, IL-15 signals will only weakly activate the PI(3)K pathway, resulting in inefficient mTOR activation, leaving Klf2 free to enhance sell gene expression as well as the genes encoding other receptors important to lymphocyte migration, including S1P1 and CCR7.130 More recent work has also found that the transcription factor Foxo1 can regulate CD62L expression on naive T cells perhaps via IL-7R signaling,135,136 but whether Foxo1 directly binds to the sell promoter or works indirectly through Klf2 is unknown. To date, the work using conditional knockout mice, in which the importance of Klf2 and Foxo1 in the regulation of sell gene expression was demonstrated, has been exclusively performed using naive T cells.130,135,136 However, gene-profiling studies have shown that early after activation Klf2 mRNA and protein levels decrease, while at later time points during the effector phase and in memory cells Klf2 levels increase.137–139 Therefore, it will be interesting to explore whether CD62L expression is regulated in a similar manner within the central memory CD8+ T cells and how the two transcription factors Klf2 and Foxo1 are regulated within the memory precursor subpopulations described earlier. It should also be kept in mind that epigenetic modification of the sell locus could also play an important role in regulation of CD62L expression, although that aspect has yet to be analyzed. In any case, a more thorough understanding of the factors regulating the Foxo1 and Klf2 transcription factors will provide insight into the process of central memory CD8+ T-cell development.
Figure 2.
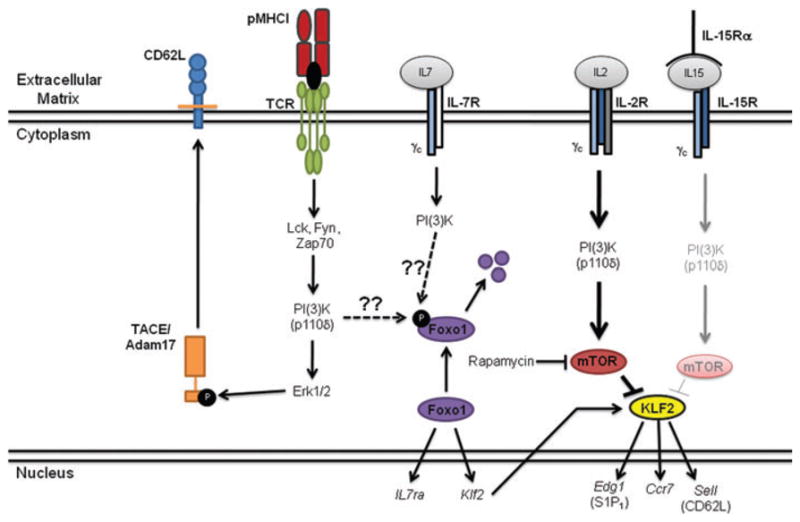
Model of CD62L regulation by T-cell receptor (TCR) stimulation and γc-cytokines. Antigen-specific CD8+ T cells are activated following engagement of the TCR with cognate peptide antigen presented in the context of major histocompatibility complex (MHC)-I. This leads to the activation of numerous signaling molecules, including the p110δ subunit of phosphatidylinositol-3-kinase (PI[3]K). This leads to activation of Erk1/2, which will then phosphorylate the TACE/Adam17 protease. Following phosphorylation the TACE/Adam17 protease will translocate to the cell membrane where it can cleave CD62L from the cell surface. This results in the transient early downregulation of CD62L. More sustained modulation of CD62L expression occurs at the genetic level and appears to be regulated, at least in part, by the γc-cytokines IL-2 and IL-15. IL-2 strongly activates the p110δ subunit of PI(3)K that will subsequently activate the mammalian target of rapamycin (mTOR) complex, while IL-15 only weakly activates this pathway. mTOR then regulates Klf2 activity, which is known to enhance the expression of CD62L, CCR7, and S1P1.
Molecular determinants of central and effector memory differentiation
Initially, in vitro studies indicated that the strength and/or duration of TCR engagement could play an important role in directing the differentiation of CD4+ and CD8+ T cells into either central or effector memory-like cells.140,141 Naive CD4+ T cells stimulated in vitro by a weak stimulus underwent proliferation but preferentially trafficked to the lymph nodes, indicative of TCM.142 Similarly, priming of CD8+ T cells in vitro at a low DC: T-cell ratio preferentially generates TCM, while higher ratios tend to generate TEM.143,144 In vivo studies using graded numbers of transgenic CD8+ T cells indicate that when high numbers of antigen-specific CD8+ T cells are transferred prior to activation, the CD62Lhigh central memory population emerges more rapidly.69–71 This finding also holds true when examining the endogenous, polyclonal, antigen-specific CD8+ T-cell populations of different naive T-cell frequencies.14 These data suggest that competition for antigen by the responding CD8+ T cells may direct the differentiation of central and effector memory lineages. Further support for this hypothesis comes from studies in which mice treated with Flt3L to increase the number of DCs are given high numbers of TCR transgenic CD8+ T cells.70 Following immunization, the resultant memory population contains fewer CD62Lhigh memory cells compared to controls, again supporting the concept that the antigen-bearing APC: T-cell ratio regulates CD62L expression. However, the results of these studies could be affected by the possibility that transfer of high numbers of antigen-specific CD8+ T cells may result in more rapid clearance of the pathogen, resulting in an altered inflammatory milieu.145 To circumvent this problem, we have established a system to modulate antigen availability by administration of a monoclonal antibody (25-D1.16) specific for the SIINFEKL peptide (derived from chicken ovalbumin) presented in the context of the class I MHC molecule H-2Kb.146 Using this protocol during vesicular stomatitis virus encoding ovalbumin (VSV-ova) infection resulted in an increase in CD62Lhigh central memory precursor cells early during the response (J.J. Obar et al., manuscript submitted). Furthermore, this was likely not because of an alteration of the inflammatory environment as the effect was observed only for CD8+ T cells specific for SIINFEKL/Kb; CD8+ T cells specific for the irrelevant epitope VSV-N/Kb were left unaffected. Thus, limiting antigen availability enhanced central memory T-cell development. Additionally, the temporal window for altering the phenotype of the resultant memory CD8+ T-cell population was brief. Antigen availability was necessary for up to 4 days for optimal expansion of the antigen-specific CD8+ T-cell population. However, competition for antigen only appeared to affect the Tem-Tcm lineage decision during the period up to approximately 72 h after infection (J.J. Obar et al., manuscript submitted). Thus, the present data strongly support the concept that the level of stimulation via the TCR regulates the differentiation of central and effector memory T-cell subsets.
It is also known that CD4+ T-cell help is important in mounting effective CD8+ T-cell responses in some cases.147–154 The provision of help to CD8+ T cells by CD4+ T cells is thought to act through the activation of the DCs, resulting in upregulation of costimulatory molecules and cytokine production,155,156 in addition to the direct production of inflammatory cytokines and chemokines by the CD4+ T cells.157,158 So-called “helpless” memory CD8+ T cells, those raised in the absence of CD4+ T cells, have an apparent defect in their long-term survival and recall capacity,149,151,159 suggesting that conditioning of memory precursors by CD4+ T cells in the primary response plays an important role in generating functional CD8+ memory T cells. The mechanism by which this occurs has yet to be defined, and our published and preliminary results do not support this model152 (J.J. Obar & L. Lefrançois, unpublished observation). Nonetheless, the memory CD8+ T-cell population produced in CD4+ T-cell-deficient animals contains an increased proportion of CD62Lhigh TCM.149 These results suggest that CD4+ T-cell help may impact the lineage decision directing central versus effector memory CD8+ T-cell development, although at what stage of the response this occurs is unknown. The two most likely downstream intermediates modulated by CD4+ T cells are the expression of costimulatory molecules on the APCs and the expression of inflammatory cytokines by the APCs and/or CD4+ T cells themselves. Once again, as with the generation of effector cell subpopulations, very little is known regarding the role of costimulatory molecules in the generation of memory cell heterogeneity. Of note is that the use of immature DCs or intercellular adhesion molecule 1 (ICAM1)−/− DCs for immunization results in the generation of a memory population that is skewed toward the central memory phenotype.160 Thus, not only do the large numbers of potential costimulators need to be studied but the extensive family of adhesion molecules that can alter the duration of T-cell–APC interactions also require additional exploration with respect to their potential roles in memory development.
On the other hand, the role of cytokines, specifically members of the γc-cytokine family, has been investigated in memory cell subset differentiation. In vitro studies have demonstrated that activation of CD8+ T cells through the TCR in the presence of IL-2 leads to accumulation of cells with a CD62Llow effector memory phenotype, while activation in the presence of IL-15 leads to the accumulation of CD62Lhigh central memory phenotype cells.131,133,134 Furthermore, our in vivo data using CD25−/− (IL-2Rα) mixed bone marrow chimeras or IL-15−/− mice support these observations, with more CD62Lhigh cells present in the memory precursor CD8+ T cells from the CD25-deficient population and less being found in the absence of IL-15 (J.J. Obar et al., manuscript submitted), which fits with previous findings showing that memory CD8+ T cells are skewed toward a TCM phenotype in CD25-deficient cells.161,162 The γc-cytokine family contains six members (IL-2, IL-4, IL-7, IL-9, IL-15, IL-21), all of which could impact the memory subset differentiation decision. Of note, in vitro studies using IL-21 as a growth factor have shown that cell-surface CD62L expression is maintained on the activated CD8+ T cells.163 Thus, a more thorough examination of the γc-cytokine family as well as other inflammatory cytokines, such as IFN-α/β, IFN-γ, and the IL-12 family, may shed light on the factors directing differentiation of the different memory subsets.
Future perspectives
Much has been learned about the origins of effector and memory CD8+ T cells over the past few years. However, many of the molecular mechanisms behind these processes remain ill defined. More specifically, we have readily been able to identify some of the factors important in the generation of terminally differentiated short-lived effector cells, but the factors regulating memory precursor effector cell differentiation have largely eluded us. An understanding of such factors will be critically important in the rational design of vaccination protocols that may induce larger numbers of long-lived and protective memory T cells. Furthermore, much of our knowledge base with respect to memory CD8+ T cells comes from primary infection, but most human vaccines require multiple administrations of attenuated pathogens or co-administration of protein antigens with adjuvants, such as alum. Thus, we must begin to examine the differentiation of memory cells after multiple antigenic encounters and what effect the adjuvant used has on the subsequent memory cell development.164 Furthermore, what relationship the peripheral memory cells in restricted environments, such as the skin, gut, and brain, has with the systemic population remains to be explored as these cells do not seem to be efficiently repopulated by the systemic pool.165 We are beginning to make progress toward the generation of rational vaccines, as evidenced by the development of the human papillomavirus vaccine, which uses virus-like particles,166 but much remains to be discovered and successfully applied to clinical settings.
Acknowledgments
The authors thank all the members of the Lefrançois lab for insightful discussions shaping this paper. This work was funded by the U.S. National Institutes of Health grants (Leo Lefrancois) and F32AI074277 (Joshua J. Obar).
Footnotes
Conflict of interest
The authors declare no conflicts of interest.
References
- 1.Plotkin SA. Vaccines: correlates of vaccine-induced immunity. Clin Infect Dis. 2008;47:401–409. doi: 10.1086/589862. [DOI] [PubMed] [Google Scholar]
- 2.Nabel GJ. Mapping the future of HIV vaccines. Nat Rev Microbiol. 2007;5:482–484. doi: 10.1038/nrmicro1713. [DOI] [PubMed] [Google Scholar]
- 3.Todryk SM, Hill AV. Malaria vaccines: the stage we are at. Nat Rev Microbiol. 2007;5:487–489. doi: 10.1038/nrmicro1712. [DOI] [PubMed] [Google Scholar]
- 4.Hikono H, et al. T-cell memory and recall responses to respiratory virus infections. Immunol Rev. 2006;211:119–132. doi: 10.1111/j.0105-2896.2006.00385.x. [DOI] [PubMed] [Google Scholar]
- 5.Hill AV. Pre-erythrocytic malaria vaccines: towards greater efficacy. Nat Rev Immunol. 2006;6:21–32. doi: 10.1038/nri1746. [DOI] [PubMed] [Google Scholar]
- 6.McMichael AJ. HIV vaccines. Annu Rev Immunol. 2006;24:227–255. doi: 10.1146/annurev.immunol.24.021605.090605. [DOI] [PubMed] [Google Scholar]
- 7.Brown LE, Kelso A. Prospects for an influenza vaccine that induces cross-protective cytotoxic T lymphocytes. Immunol Cell Biol. 2009;87:300–308. doi: 10.1038/icb.2009.16. [DOI] [PubMed] [Google Scholar]
- 8.Masopust D. Developing an HIV cytotoxic T-lymphocyte vaccine: issues of CD8 T-cell quantity, quality and location. J Intern Med. 2009;265:125–137. doi: 10.1111/j.1365-2796.2008.02054.x. [DOI] [PubMed] [Google Scholar]
- 9.Grebe KM, Yewdell JW, Bennink JR. Heterosubtypic immunity to influenza A virus: where do we stand? Microbes Infect. 2008;10:1024–1029. doi: 10.1016/j.micinf.2008.07.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Wong P, Pamer EG. CD8 T cell responses to infectious pathogens. Annu Rev Immunol. 2003;21:29–70. doi: 10.1146/annurev.immunol.21.120601.141114. [DOI] [PubMed] [Google Scholar]
- 11.De Visser KE, Eichten A, Coussens LM. Paradoxical roles of the immune system during cancer development. Nat Rev Cancer. 2006;6:24–37. doi: 10.1038/nrc1782. [DOI] [PubMed] [Google Scholar]
- 12.Gaucher D, et al. Yellow fever vaccine induces integrated multilineage and polyfunctional immune responses. J Exp Med. 2008;205:3119–3131. doi: 10.1084/jem.20082292. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Querec TD, et al. Systems biology approach predicts immunogenicity of the yellow fever vaccine in humans. Nat Immunol. 2009;10:116–125. doi: 10.1038/ni.1688. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Obar JJ, Khanna KM, Lefrancois L. Endogenous naive CD8+ T cell precursor frequency regulates primary and memory responses to infection. Immunity. 2008;28:859–869. doi: 10.1016/j.immuni.2008.04.010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Kotturi MF, et al. Naive precursor frequencies and MHC binding rather than the degree of epitope diversity shape CD8+ T cell immunodominance. J Immunol. 2008;181:2124–2133. doi: 10.4049/jimmunol.181.3.2124. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Haluszczak C, et al. The antigen-specific CD8+ T cell repertoire in unimmunized mice includes memory phenotype cells bearing markers of homeostatic expansion. J Exp Med. 2009;206:435–448. doi: 10.1084/jem.20081829. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Moon JJ, et al. Naive CD4(+) T cell frequency varies for different epitopes and predicts repertoire diversity and response magnitude. Immunity. 2007;27:203–213. doi: 10.1016/j.immuni.2007.07.007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Nahill SR, Welsh RM. High frequency of cross-reactive cytotoxic T lymphocytes elicited during the virus-induced polyclonal cytotoxic T lymphocyte response. J Exp Med. 1993;177:317–327. doi: 10.1084/jem.177.2.317. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Selin LK, Nahill SR, Welsh RM. Cross-reactivities in memory cytotoxic T lymphocyte recognition of heterologous viruses. J Exp Med. 1994;179:1933–1943. doi: 10.1084/jem.179.6.1933. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Jung S, et al. In vivo depletion of CD11c(+) dendritic cells abrogates priming of CD8(+) T cells by exogenous cell-associated antigens. Immunity. 2002;17:211–220. doi: 10.1016/s1074-7613(02)00365-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Pozzi LA, Maciaszek JW, Rock KL. Both dendritic cells and macrophages can stimulate naive CD8 T cells in vivo to proliferate, develop effector function, and differentiate into memory cells. J Immunol. 2005;175:2071–2081. doi: 10.4049/jimmunol.175.4.2071. [DOI] [PubMed] [Google Scholar]
- 22.Kaech SM, Ahmed R. Memory CD8+ T cell differentiation: initial antigen encounter triggers a developmental program in naive cells. Nat Immunol. 2001;2:415–422. doi: 10.1038/87720. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Van Stipdonk MJ, Lemmens EE, Schoenberger SP. Naive CTLs require a single brief period of antigenic stimulation for clonal expansion and differentiation. Nat Immunol. 2001;2:423–429. doi: 10.1038/87730. [DOI] [PubMed] [Google Scholar]
- 24.Obst R, Van Santen HM, Mathis D, Benoist C. Antigen persistence is required throughout the expansion phase of a CD4(+) T cell response. J Exp Med. 2005;201:1555–1565. doi: 10.1084/jem.20042521. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Celli S, Lemaitre F, Bousso P. Real-time manipulation of T cell-dendritic cell interactions in vivo reveals the importance of prolonged contacts for CD4+ T cell activation. Immunity. 2007;27:625–634. doi: 10.1016/j.immuni.2007.08.018. [DOI] [PubMed] [Google Scholar]
- 26.Khanna KM, McNamara JT, Lefrancois L. In situ imaging of the endogenous CD8 T cell response to infection. Science. 2007;318:116–120. doi: 10.1126/science.1146291. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Sarkar S, et al. Strength of stimulus and clonal competition impact the rate of memory CD8 T cell differentiation. J Immunol. 2007;179:6704–6714. doi: 10.4049/jimmunol.179.10.6704. [DOI] [PubMed] [Google Scholar]
- 28.Blair DA, Lefrancois L. Increased competition for antigen during priming negatively impacts the generation of memory CD4 T cells. Proc Natl Acad Sci USA. 2007;104:15045–15050. doi: 10.1073/pnas.0703767104. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Mescher MF, et al. Signals required for programming effector and memory development by CD8+ T cells. Immunol Rev. 2006;211:81–92. doi: 10.1111/j.0105-2896.2006.00382.x. [DOI] [PubMed] [Google Scholar]
- 30.Thompson LJ, Kolumam GA, Thomas S, Murali-Krishna K. Innate inflammatory signals induced by various pathogens differentially dictate the IFN-I dependence of CD8 T cells for clonal expansion and memory formation. J Immunol. 2006;177:1746–1754. doi: 10.4049/jimmunol.177.3.1746. [DOI] [PubMed] [Google Scholar]
- 31.Xiao Z, et al. Programming for CD8 T cell memory development requires IL-12 or type I IFN. J Immunol. 2009;182:2786–2794. doi: 10.4049/jimmunol.0803484. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Knickelbein JE, et al. Noncytotoxic lytic granule-mediated CD8+ T cell inhibition of HSV-1 reactivation from neuronal latency. Science. 2008;322:268–271. doi: 10.1126/science.1164164. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Reinhardt RL, et al. Visualizing the generation of memory CD4 T cells in the whole body. Nature. 2001;410:101–105. doi: 10.1038/35065111. [DOI] [PubMed] [Google Scholar]
- 34.Masopust D, Vezys V, Marzo AL, Lefrançois L. Preferential localization of effector memory cells in nonlymphoid tissue. Science. 2001;291:2413–2417. doi: 10.1126/science.1058867. [DOI] [PubMed] [Google Scholar]
- 35.Hildeman DA, et al. Activated T cell death in vivo mediated by proapoptotic bcl-2 family member bim. Immunity. 2002;16:759–767. doi: 10.1016/s1074-7613(02)00322-9. [DOI] [PubMed] [Google Scholar]
- 36.Wojciechowski S, et al. Bim mediates apoptosis of CD127(lo) effector T cells and limits T cell memory. Eur J Immunol. 2006;36:1694–1706. doi: 10.1002/eji.200635897. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Wojciechowski S, et al. Bim/Bcl-2 balance is critical for maintaining naive and memory T cell homeostasis. J Exp Med. 2007;204:1665–1675. doi: 10.1084/jem.20070618. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Weant AE, et al. Apoptosis regulators Bim and Fas function concurrently to control autoimmunity and CD8+ T cell contraction. Immunity. 2008;28:218–230. doi: 10.1016/j.immuni.2007.12.014. [DOI] [PubMed] [Google Scholar]
- 39.Hughes PD, et al. Apoptosis regulators Fas and Bim cooperate in shutdown of chronic immune responses and prevention of autoimmunity. Immunity. 2008;28:197–205. doi: 10.1016/j.immuni.2007.12.017. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Hutcheson J, et al. Combined deficiency of proapoptotic regulators Bim and Fas results in the early onset of systemic autoimmunity. Immunity. 2008;28:206–217. doi: 10.1016/j.immuni.2007.12.015. [DOI] [PubMed] [Google Scholar]
- 41.Green DR. Fas Bim boom! Immunity. 2008;28:141–143. doi: 10.1016/j.immuni.2008.01.004. [DOI] [PubMed] [Google Scholar]
- 42.Murali-Krishna K, et al. Persistence of memory CD8 T cells in MHC class I-deficient mice. Science. 1999;286:1377–1381. doi: 10.1126/science.286.5443.1377. [DOI] [PubMed] [Google Scholar]
- 43.Leignadier J, et al. Memory T-lymphocyte survival does not require T-cell receptor expression. Proc Natl Acad Sci USA. 2008;105:20440–20445. doi: 10.1073/pnas.0806289106. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Schluns KS, et al. Cutting Edge: Requirement for IL-15 in the generation of primary and memory antigen-specific CD8 T cells. J Immunol. 2002;168:4827–4831. doi: 10.4049/jimmunol.168.10.4827. [DOI] [PubMed] [Google Scholar]
- 45.Becker TC, et al. Interleukin 15 is required for proliferative renewal of virus-specific memory CD8 T cells. J Exp Med. 2002;195:1541–1548. doi: 10.1084/jem.20020369. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Schluns KS, Kieper WC, Jameson SC, Lefrançois L. Interleukin-7 mediates the homeostasis of naive and memory CD8 T cells in vivo. Nat Immunol. 2000;1:426–432. doi: 10.1038/80868. [DOI] [PubMed] [Google Scholar]
- 47.Schluns KS, Lefrancois L. Cytokine control of memory T-cell development and survival. Nat Rev Immunol. 2003;3:269–279. doi: 10.1038/nri1052. [DOI] [PubMed] [Google Scholar]
- 48.Klonowski KD, Williams KJ, Marzo AL, Lefrancois L. Cutting edge: IL-7-independent regulation of IL-7 receptor alpha expression and memory CD8 T cell development. J Immunol. 2006;177:4247–4251. doi: 10.4049/jimmunol.177.7.4247. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Tanchot C, et al. Modifications of CD8+ T cell function during in vivo memory or tolerance induction. Immunity. 1998;8:581–590. doi: 10.1016/s1074-7613(00)80563-4. [DOI] [PubMed] [Google Scholar]
- 50.Whitmire JK, Eam B, Whitton JL. Tentative T cells: memory cells are quick to respond, but slow to divide. PLoS Pathog. 2008;4:e1000041. doi: 10.1371/journal.ppat.1000041. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Veiga-Fernandes H, et al. Response of naive and memory CD8+ T cells to antigen stimulation in vivo. Nat Immunol. 2000;1:47–53. doi: 10.1038/76907. [DOI] [PubMed] [Google Scholar]
- 52.Hussain SF, Anderson CF, Farber DL. Differential SLP-76 expression and TCR-mediated signaling in effector and memory CD4 T cells. J Immunol. 2002;168:1557–1565. doi: 10.4049/jimmunol.168.4.1557. [DOI] [PubMed] [Google Scholar]
- 53.Veiga-Fernandes H, Rocha B. High expression of active CDK6 in the cytoplasm of CD8 memory cells favors rapid division. Nat Immunol. 2004;5:31–37. doi: 10.1038/ni1015. [DOI] [PubMed] [Google Scholar]
- 54.Chandok MR, Farber DL. Signaling control of memory T cell generation and function. Semin Immunol. 2004;16:285–293. doi: 10.1016/j.smim.2004.08.009. [DOI] [PubMed] [Google Scholar]
- 55.Hamann D, et al. Phenotypic and functional separation of memory and effector human CD8+ T cells. J Exp Med. 1997;186:1407–1418. doi: 10.1084/jem.186.9.1407. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Sallusto F, et al. Two subsets of memory T lymphocytes with distinct homing potentials and effector functions. Nature. 1999;401:708–712. doi: 10.1038/44385. [DOI] [PubMed] [Google Scholar]
- 57.Springer TA. Traffic signals for lymphocyte recirculation and leukocyte emigration: the multistep paradigm. Cell. 1994;76:301–314. doi: 10.1016/0092-8674(94)90337-9. [DOI] [PubMed] [Google Scholar]
- 58.Guarda G, et al. L-selectin-negative CCR7-effector and memory CD8+ T cells enter reactive lymph nodes and kill dendritic cells. Nat Immunol. 2007;8:743–752. doi: 10.1038/ni1469. [DOI] [PubMed] [Google Scholar]
- 59.Forster R, et al. CCR7 coordinates the primary immune response by establishing functional microenvironments in secondary lymphoid organs. Cell. 1999;99:23–33. doi: 10.1016/s0092-8674(00)80059-8. [DOI] [PubMed] [Google Scholar]
- 60.Bromley SK, Thomas SY, Luster AD. Chemokine receptor CCR7 guides T cell exit from peripheral tissues and entry into afferent lymphatics. Nat Immunol. 2005;6:895–901. doi: 10.1038/ni1240. [DOI] [PubMed] [Google Scholar]
- 61.Debes GF, et al. Chemokine receptor CCR7 required for T lymphocyte exit from peripheral tissues. Nat Immunol. 2005;6:889–894. doi: 10.1038/ni1238. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62.Geginat J, Sallusto F, Lanzavecchia A. Cytokine-driven proliferation and differentiation of human naive, central memory, and effector memory CD4(+) T cells. J Exp Med. 2001;194:1711–1719. doi: 10.1084/jem.194.12.1711. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.Lefrancois L. Development, trafficking, and function of memory T-cell subsets. Immunol Rev. 2006;211:93–103. doi: 10.1111/j.0105-2896.2006.00393.x. [DOI] [PubMed] [Google Scholar]
- 64.Lefrancois L, Marzo AL. The descent of memory T-cell subsets. Nat Rev Immunol. 2006;6:618–623. doi: 10.1038/nri1866. [DOI] [PubMed] [Google Scholar]
- 65.Khanna KM, Bonneau RH, Kinchington PR, Hendricks RL. Herpes simplex virus-specific memory CD8+ T cells are selectively activated and retained in latently infected sensory ganglia. Immunity. 2003;18:593–603. doi: 10.1016/s1074-7613(03)00112-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 66.Wakim LM, et al. Dendritic cell-induced memory T cell activation in nonlymphoid tissues. Science. 2008;319:198–202. doi: 10.1126/science.1151869. [DOI] [PubMed] [Google Scholar]
- 67.Gebhardt T, et al. Memory T cells in nonlymphoid tissue that provide enhanced local immunity during infection with herpes simplex virus. Nat Immunol. 2009;10:524–530. doi: 10.1038/ni.1718. [DOI] [PubMed] [Google Scholar]
- 68.Tripp RA, Hou S, Doherty PC. Temporal loss of the activated L-selectin-low phenotype for virus-specific CD8+ memory T cells. J Immunol. 1995;154:5870–5875. [PubMed] [Google Scholar]
- 69.Wherry EJ, et al. Lineage relationship and protective immunity of memory CD8 T cell subsets. Nat Immunol. 2003;4:225–234. doi: 10.1038/ni889. [DOI] [PubMed] [Google Scholar]
- 70.Marzo AL, et al. Initial T cell frequency dictates memory CD8+ T cell lineage commitment. Nat Immunol. 2005;6:793–799. doi: 10.1038/ni1227. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 71.Badovinac VP, Haring JS, Harty JT. Initial T cell receptor transgenic cell precursor frequency dictates critical aspects of the CD8(+) T cell response to infection. Immunity. 2007;26:827–841. doi: 10.1016/j.immuni.2007.04.013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 72.Unsoeld H, Pircher H. Complex memory T-cell phenotypes revealed by coexpression of CD62L and CCR7. J Virol. 2005;79:4510–4513. doi: 10.1128/JVI.79.7.4510-4513.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 73.Hikono H, et al. Activation phenotype, rather than central- or effector-memory phenotype, predicts the recall efficacy of memory CD8+ T cells. J Exp Med. 2007;204:1625–1636. doi: 10.1084/jem.20070322. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 74.Zammit DJ, et al. Residual antigen presentation after influenza virus infection affects CD8 T cell activation and migration. Immunity. 2006;24:439–449. doi: 10.1016/j.immuni.2006.01.015. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 75.Khanna KM, et al. In situ imaging reveals different responses by naive and memory CD8 T cells to late antigen presentation by lymph node DC after influenza virus infection. Eur J Immunol. 2008;38:3304–3315. doi: 10.1002/eji.200838602. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 76.Woodland DL, Kohlmeier JE. Migration, maintenance and recall of memory T cells in peripheral tissues. Nat Rev Immunol. 2009;9:153–161. doi: 10.1038/nri2496. [DOI] [PubMed] [Google Scholar]
- 77.Hislop AD, et al. EBV-specific CD8+ T cell memory: relationships between epitope specificity, cell phenotype, and immediate effector function. J Immunol. 2001;167:2019–2029. doi: 10.4049/jimmunol.167.4.2019. [DOI] [PubMed] [Google Scholar]
- 78.Catalina MD, Sullivan JL, Brody RM, Luzuriaga K. Phenotypic and functional heterogeneity of EBV epitope-specific CD8+ T cells. J Immunol. 2002;168:4184–4191. doi: 10.4049/jimmunol.168.8.4184. [DOI] [PubMed] [Google Scholar]
- 79.Obar JJ, Crist SG, Gondek DC, Usherwood EJ. Different functional capacities of latent and lytic antigen-specific CD8 T cells in murine gammaherpesvirus infection. J Immunol. 2004;172:1213–1219. doi: 10.4049/jimmunol.172.2.1213. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 80.Wherry EJ, et al. Viral persistence alters CD8 T cell immunodominance and tissue distribution and results in distinct stages of functional impairment. J Virol. 2003;77:4911–4927. doi: 10.1128/JVI.77.8.4911-4927.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 81.Obar JJ, Crist SG, Leung EK, Usherwood EJ. IL-15-independent proliferative renewal of memory CD8+ T cells in latent gammaherpesvirus infection. J Immunol. 2004;173:2705–2714. doi: 10.4049/jimmunol.173.4.2705. [DOI] [PubMed] [Google Scholar]
- 82.Wherry EJ, et al. Antigen-independent memory CD8 T cells do not develop during chronic viral infection. Proc Natl Acad Sci USA. 2004;101:16004–16009. doi: 10.1073/pnas.0407192101. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 83.Obar JJ, et al. Gammaherpesvirus persistence alters key CD8 T-cell memory characteristics and enhances antiviral protection. J Virol. 2006;80:8303–8315. doi: 10.1128/JVI.00237-06. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 84.Sheridan BS, Khanna KM, Frank GM, Hendricks RL. Latent virus influences the generation and maintenance of CD8+ T cell memory. J Immunol. 2006;177:8356–8364. doi: 10.4049/jimmunol.177.12.8356. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 85.Munks MW, et al. Four distinct patterns of memory CD8 T cell responses to chronic murine cytomegalovirus infection. J Immunol. 2006;177:450–458. doi: 10.4049/jimmunol.177.1.450. [DOI] [PubMed] [Google Scholar]
- 86.Stemberger C, et al. A single naive CD8+ T cell precursor can develop into diverse effector and memory subsets. Immunity. 2007;27:985–997. doi: 10.1016/j.immuni.2007.10.012. [DOI] [PubMed] [Google Scholar]
- 87.Seder RA, Ahmed R. Similarities and differences in CD4(+) and CD8(+) effector and memory T cell generation. Nat Immunol. 2003;4:835–842. doi: 10.1038/ni969. [DOI] [PubMed] [Google Scholar]
- 88.Harrington LE, et al. Memory CD4 T cells emerge from effector T-cell progenitors. Nature. 2008;452:356–360. doi: 10.1038/nature06672. [DOI] [PubMed] [Google Scholar]
- 89.Bannard O, Kraman M, Fearon DT. Secondary replicative function of CD8+ T cells that had developed an effector phenotype. Science. 2009;323:505–509. doi: 10.1126/science.1166831. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 90.Kaech SM, et al. Selective expression of the interleukin 7 receptor identifies effector CD8 T cells that give rise to long-lived memory cells. Nat Immunol. 2003;4:1191–1198. doi: 10.1038/ni1009. [DOI] [PubMed] [Google Scholar]
- 91.Joshi NS, et al. Inflammation directs memory precursor and short-lived effector CD8(+) T cell fates via the graded expression of T-bet transcription factor. Immunity. 2007;27:281–295. doi: 10.1016/j.immuni.2007.07.010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 92.Sarkar S, et al. Functional and genomic profiling of effector CD8 T cell subsets with distinct memory fates. J Exp Med. 2008;205:625–640. doi: 10.1084/jem.20071641. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 93.Rubinstein MP, et al. IL-7 and IL-15 differentially regulate CD8+ T cell subsets during contraction of the immune response. Blood. 2008;112:3704–3712. doi: 10.1182/blood-2008-06-160945. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 94.Chang JT, et al. Asymmetric T lymphocyte division in the initiation of adaptive immune responses. Science. 2007;315:1687–1691. doi: 10.1126/science.1139393. [DOI] [PubMed] [Google Scholar]
- 95.Stonier SW, Ma LJ, Castillo EF, Schluns KS. Dendritic cells drive memory CD8 T cell homeostasis via IL-15 transpresentation. Blood. 2008;112:4546–4554. doi: 10.1182/blood-2008-05-156307. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 96.Wherry EJ, Puorro KA, Porgador A, Eisenlohr LC. The induction of virus-specific CTL as a function of increasing epitope expression: responses rise steadily until excessively high levels of epitope are attained. J Immunol. 1999;163:3735–3745. [PubMed] [Google Scholar]
- 97.Wherry EJ, Mcelhaugh MJ, Eisenlohr LC. Generation of CD8(+) T cell memory in response to low, high, and excessive levels of epitope. J Immunol. 2002;168:4455–4461. doi: 10.4049/jimmunol.168.9.4455. [DOI] [PubMed] [Google Scholar]
- 98.Bullock TN, Mullins DW, Engelhard VH. Antigen density presented by dendritic cells in vivo differentially affects the number and avidity of primary, memory, and recall CD8+ T cells. J Immunol. 2003;170:1822–1829. doi: 10.4049/jimmunol.170.4.1822. [DOI] [PubMed] [Google Scholar]
- 99.Prlic M, Hernandez-Hoyos G, Bevan MJ. Duration of the initial TCR stimulus controls the magnitude but not functionality of the CD8+ T cell response. J Exp Med. 2006;203:2135–2143. doi: 10.1084/jem.20060928. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 100.Greenwald RJ, Freeman GJ, Sharpe AH. The B7 family revisited. Annu Rev Immunol. 2005;23:515–548. doi: 10.1146/annurev.immunol.23.021704.115611. [DOI] [PubMed] [Google Scholar]
- 101.Watts TH. TNF/TNFR family members in costimulation of T cell responses. Annu Rev Immunol. 2005;23:23–68. doi: 10.1146/annurev.immunol.23.021704.115839. [DOI] [PubMed] [Google Scholar]
- 102.Mousavi SF, et al. OX40 costimulatory signals potentiate the memory commitment of effector CD8+ T cells. J Immunol. 2008;181:5990–6001. doi: 10.4049/jimmunol.181.9.5990. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 103.Cousens LP, et al. Two roads diverged: interferon alpha/beta- and interleukin 12-mediated pathways in promoting T cell interferon gamma responses during viral infection. J Exp Med. 1999;189:1315–1328. doi: 10.1084/jem.189.8.1315. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 104.Badovinac VP, Tvinnereim AR, Harty JT. Regulation of antigen-specific CD8(+) T cell homeostasis by perforin and interferon-gamma. Science. 2000;290:1354–1358. doi: 10.1126/science.290.5495.1354. [DOI] [PubMed] [Google Scholar]
- 105.Curtsinger JM, Johnson CM, Mescher MF. CD8 T cell clonal expansion and development of effector function require prolonged exposure to antigen, costimulation, and signal 3 cytokine. J Immunol. 2003;171:5165–5171. doi: 10.4049/jimmunol.171.10.5165. [DOI] [PubMed] [Google Scholar]
- 106.Haring JS, Corbin GA, Harty JT. Dynamic regulation of IFN-gamma signaling in antigen-specific CD8+ T cells responding to infection. J Immunol. 2005;174:6791–6802. doi: 10.4049/jimmunol.174.11.6791. [DOI] [PubMed] [Google Scholar]
- 107.Kolumam GA, et al. Type I interferons act directly on CD8 T cells to allow clonal expansion and memory formation in response to viral infection. J Exp Med. 2005;202:637–650. doi: 10.1084/jem.20050821. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 108.Whitmire JK, Tan JT, Whitton JL. Interferon-gamma acts directly on CD8+ T cells to increase their abundance during virus infection. J Exp Med. 2005;201:1053–1059. doi: 10.1084/jem.20041463. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 109.Badovinac VP, Porter BB, Harty JT. CD8+ T cell contraction is controlled by early inflammation. Nat Immunol. 2004;5:809–817. doi: 10.1038/ni1098. [DOI] [PubMed] [Google Scholar]
- 110.Badovinac VP, et al. Accelerated CD8+ T-cell memory and prime-boost response after dendritic-cell vaccination. Nat Med. 2005;11:748–756. doi: 10.1038/nm1257. [DOI] [PubMed] [Google Scholar]
- 111.Wilson DC, Matthews S, Yap GS. IL-12 signaling drives CD8+ T cell IFN-gamma production and differentiation of KLRG1+ effector subpopulations during Toxoplasma gondii Infection. J Immunol. 2008;180:5935–5945. doi: 10.4049/jimmunol.180.9.5935. [DOI] [PubMed] [Google Scholar]
- 112.Cui W, Joshi NS, Jiang A, Kaech SM. Effects of Signal 3 during CD8 T cell priming: Bystander production of IL-12 enhances effector T cell expansion but promotes terminal differentiation. Vaccine. 2009;27:2177–2187. doi: 10.1016/j.vaccine.2009.01.088. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 113.Rahman AH, et al. MyD88 plays a critical T cell-intrinsic role in supporting CD8 T cell expansion during acute lymphocytic choriomeningitis virus infection. J Immunol. 2008;181:3804–3810. doi: 10.4049/jimmunol.181.6.3804. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 114.Teixeiro E, et al. Different T cell receptor signals determine CD8+ memory versus effector development. Science. 2009;323:502–505. doi: 10.1126/science.1163612. [DOI] [PubMed] [Google Scholar]
- 115.Intlekofer AM, et al. Effector and memory CD8+ T cell fate coupled by T-bet and eomesodermin. Nat Immunol. 2005;6:1236–1244. doi: 10.1038/ni1268. [DOI] [PubMed] [Google Scholar]
- 116.Intlekofer AM, et al. Requirement for T-bet in the aberrant differentiation of unhelped memory CD8+ T cells. J Exp Med. 2007;204:2015–2021. doi: 10.1084/jem.20070841. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 117.Benson MJ, Erickson LD, Gleeson MW, Noelle RJ. Affinity of antigen encounter and other early B-cell signals determine B-cell fate. Curr Opin Immunol. 2007;19:275–280. doi: 10.1016/j.coi.2007.04.009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 118.Ichii H, et al. Role for Bcl-6 in the generation and maintenance of memory CD8+ T cells. Nat Immunol. 2002;3:558–563. doi: 10.1038/ni802. [DOI] [PubMed] [Google Scholar]
- 119.Ichii H, Sakamoto A, Kuroda Y, Tokuhisa T. Bcl6 acts as an amplifier for the generation and proliferative capacity of central memory CD8+ T cells. J Immunol. 2004;173:883–891. doi: 10.4049/jimmunol.173.2.883. [DOI] [PubMed] [Google Scholar]
- 120.Campbell JJ, et al. 6-C-kine (SLC), a lymphocyte adhesion-triggering chemokine expressed by high endothelium, is an agonist for the MIP-3beta receptor CCR7. J Cell Biol. 1998;141:1053–1059. doi: 10.1083/jcb.141.4.1053. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 121.Forster R, et al. CCR7 coordinates the primary immune response by establishing functional microenvironments in secondary lymphoid organs. Cell. 1999;99:23–33. doi: 10.1016/s0092-8674(00)80059-8. [DOI] [PubMed] [Google Scholar]
- 122.Dailey MO, Gallatin WM, Weissman IL. The in vivo behavior of T cell clones: altered migration due to loss of the lymphocyte surface homing receptor. J Mol Cell Immunol. 1985;2:27–36. [PubMed] [Google Scholar]
- 123.Jung TM, Gallatin WM, Weissman IL, Dailey MO. Down-regulation of homing receptors after T cell activation. J Immunol. 1988;141:4110–4117. [PubMed] [Google Scholar]
- 124.Jung TM, Dailey MO. Reversibility of loss of homing receptor expression following activation. Adv Exp Med Biol. 1988;237:519–524. doi: 10.1007/978-1-4684-5535-9_79. [DOI] [PubMed] [Google Scholar]
- 125.Mobley JL, Dailey MO. Regulation of adhesion molecule expression by CD8 T cells in vivo. I. Differential regulation of gp90MEL-14 (LECAM-1), Pgp-1, LFA-1, and VLA-4 alpha during the differentiation of cytotoxic T lymphocytes induced by allografts. J Immunol. 1992;148:2348–2356. [PubMed] [Google Scholar]
- 126.Ley K, Laudanna C, Cybulsky MI, Nourshargh S. Getting to the site of inflammation: the leukocyte adhesion cascade updated. Nat Rev Immunol. 2007;7:678–689. doi: 10.1038/nri2156. [DOI] [PubMed] [Google Scholar]
- 127.Jung TM, Dailey MO. Rapid modulation of homing receptors (gp90MEL-14) induced by activators of protein kinase C. Receptor shedding due to accelerated proteolytic cleavage at the cell surface. J Immunol. 1990;144:3130–3136. [PubMed] [Google Scholar]
- 128.Chao CC, Jensen R, Dailey MO. Mechanisms of L-selectin regulation by activated T cells. J Immunol. 1997;159:1686–1694. [PubMed] [Google Scholar]
- 129.Dang X, Raffler NA, Ley K. Transcriptional regulation of mouse l-selectin. Biochim Biophys Acta. 2008;1789:146–152. doi: 10.1016/j.bbagrm.2008.10.004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 130.Carlson CM, Endrizzi BT, Wu J, et al. Kruppel-like factor 2 regulates thymocyte and T-cell migration. Nature. 2006;442:299–302. doi: 10.1038/nature04882. [DOI] [PubMed] [Google Scholar]
- 131.Sinclair LV, et al. Phosphatidylinositol-3-OH kinase and nutrient-sensing mTOR pathways control T lymphocyte trafficking. Nat Immunol. 2008;9:513–521. doi: 10.1038/ni.1603. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 132.Cornish GH, Sinclair LV, Cantrell DA. Differential regulation of T-cell growth by IL-2 and IL-15. Blood. 2006;108:600–608. doi: 10.1182/blood-2005-12-4827. [DOI] [PubMed] [Google Scholar]
- 133.Manjunath N, et al. Effector differentiation is not prerequisite for generation of memory cytotoxic T lymphocytes. J Clin Invest. 2001;108:871–878. doi: 10.1172/JCI13296. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 134.Klebanoff CA, et al. Central memory self/tumor-reactive CD8+ T cells confer superior antitumor immunity compared with effector memory T cells. Proc Natl Acad Sci USA. 2005;102:9571–9576. doi: 10.1073/pnas.0503726102. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 135.Kerdiles YM, et al. Foxo1 links homing and survival of naive T cells by regulating L-selectin, CCR7 and interleukin 7 receptor. Nat Immunol. 2009;10:176–184. doi: 10.1038/ni.1689. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 136.Ouyang W, Beckett O, Flavell RA, Li MO. An essential role of the Forkhead-box transcription factor Foxo1 in control of T cell homeostasis and tolerance. Immunity. 2009;30:358–371. doi: 10.1016/j.immuni.2009.02.003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 137.Schober SL, et al. Expression of the transcription factor Lung Kruppel-Like Factor is regulated by cytokines and correlates with survival of memory T cells in vitro and in vivo. J Immunol. 1999;163:3662–3667. [PubMed] [Google Scholar]
- 138.Grayson JM, Murali-Krishna K, Altman JD, Ahmed R. Gene expression in antigen-specific CD8+ T cells during viral infection. J Immunol. 2001;166:795–799. doi: 10.4049/jimmunol.166.2.795. [DOI] [PubMed] [Google Scholar]
- 139.Endrizzi BT, Jameson SC. Differential role for IL-7 in inducing lung Kruppel-like factor (Kruppel-like factor 2) expression by naive versus activated T cells. Int Immunol. 2003;15:1341–1348. doi: 10.1093/intimm/dxg133. [DOI] [PubMed] [Google Scholar]
- 140.Lanzavecchia A, Sallusto F. Progressive differentiation and selection of the fittest in the immune response. Nat Rev Immunol. 2002;2:982–987. doi: 10.1038/nri959. [DOI] [PubMed] [Google Scholar]
- 141.Sallusto F, Geginat J, Lanzavecchia A. Central memory and effector memory T cell subsets: function, generation, and maintenance. Annu Rev Immunol. 2004;22:745–763. doi: 10.1146/annurev.immunol.22.012703.104702. [DOI] [PubMed] [Google Scholar]
- 142.Iezzi G, Scheidegger D, Lanzavecchia A. Migration and function of antigen-primed nonpolarized T lymphocytes in vivo. J Exp Med. 2001;193:987–993. doi: 10.1084/jem.193.8.987. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 143.Weninger W, Crowley MA, Manjunath N, Von Andrian UH. Migratory properties of naive, effector, and memory CD8(+) t cells. J Exp Med. 2001;194:953–966. doi: 10.1084/jem.194.7.953. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 144.Gett AV, Sallusto F, Lanzavecchia A, Geginat J. T cell fitness determined by signal strength. Nat Immunol. 2003;4:355–360. doi: 10.1038/ni908. [DOI] [PubMed] [Google Scholar]
- 145.Wirth TC, Pham NL, Harty JT, Badovinac VP. High initial frequency of TCR-transgenic CD8 T cells alters inflammation and pathogen clearance without affecting memory T cell function. Mol Immunol. 2009;47:71–78. doi: 10.1016/j.molimm.2008.12.018. [DOI] [PubMed] [Google Scholar]
- 146.Porgador A, et al. Localization, quantitation, and in situ detection of specific peptide-MHC class I complexes using a monoclonal antibody. Immunity. 1997;6:715–726. doi: 10.1016/s1074-7613(00)80447-1. [DOI] [PubMed] [Google Scholar]
- 147.Janssen EM, et al. CD4+ T cells are required for secondary expansion and memory in CD8+ T lymphocytes. Nature. 2003;421:852–856. doi: 10.1038/nature01441. [DOI] [PubMed] [Google Scholar]
- 148.Shedlock DJ, et al. Role of CD4 T cell help and costimulation in CD8 T cell responses during Listeria monocytogenes infection. J Immunol. 2003;170:2053–2063. doi: 10.4049/jimmunol.170.4.2053. [DOI] [PubMed] [Google Scholar]
- 149.Sun JC, Bevan MJ. Defective CD8 T cell memory following acute infection without CD4 T cell help. Science. 2003;300:339–342. doi: 10.1126/science.1083317. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 150.Wang JC, Livingstone AM. Cutting edge: CD4+ T cell help can be essential for primary CD8+ T cell responses in vivo. J Immunol. 2003;171:6339–6343. doi: 10.4049/jimmunol.171.12.6339. [DOI] [PubMed] [Google Scholar]
- 151.Sun JC, Williams MA, Bevan MJ. CD4+ T cells are required for the maintenance, not programming, of memory CD8+ T cells after acute infection. Nat Immunol. 2004;5:927–933. doi: 10.1038/ni1105. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 152.Marzo AL, et al. Fully functional memory CD8 T cells in the absence of CD4 T cells. J Immunol. 2004;173:969–975. doi: 10.4049/jimmunol.173.2.969. [DOI] [PubMed] [Google Scholar]
- 153.Wilson EB, Livingstone AM. Cutting edge: CD4+ T cell-derived IL-2 is essential for help-dependent primary CD8+ T cell responses. J Immunol. 2008;181:7445–7448. doi: 10.4049/jimmunol.181.11.7445. [DOI] [PubMed] [Google Scholar]
- 154.Fuse S, et al. Recall responses by helpless memory CD8+ T cells are restricted by the up-regulation of PD-1. J Immunol. 2009;182:4244–4254. doi: 10.4049/jimmunol.0802041. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 155.Bevan MJ. Helping the CD8(+) T-cell response. Nat Rev Immunol. 2004;4:595–602. doi: 10.1038/nri1413. [DOI] [PubMed] [Google Scholar]
- 156.Williams MA, Bevan MJ. Effector and memory CTL differentiation. Annu Rev Immunol. 2007;25:171–192. doi: 10.1146/annurev.immunol.25.022106.141548. [DOI] [PubMed] [Google Scholar]
- 157.Castellino F, et al. Chemokines enhance immunity by guiding naive CD8+ T cells to sites of CD4+ T cell-dendritic cell interaction. Nature. 2006;440:890–895. doi: 10.1038/nature04651. [DOI] [PubMed] [Google Scholar]
- 158.Castellino F, Germain RN. Chemokine-guided CD4+ T cell help enhances generation of IL-6R{alpha}highIL-7R{alpha}high prememory CD8+ T cells. J Immunol. 2007;178:778–787. doi: 10.4049/jimmunol.178.2.778. [DOI] [PubMed] [Google Scholar]
- 159.Shedlock DJ, Shen H. Requirement for CD4 T cell help in generating functional CD8 T cell memory. Science. 2003;300:337–339. doi: 10.1126/science.1082305. [DOI] [PubMed] [Google Scholar]
- 160.Parameswaran N, et al. Lack of ICAM-1 on APCs during T cell priming leads to poor generation of central memory cells. J Immunol. 2005;175:2201–2211. doi: 10.4049/jimmunol.175.4.2201. [DOI] [PubMed] [Google Scholar]
- 161.Williams MA, Tyznik AJ, Bevan MJ. Interleukin-2 signals during priming are required for secondary expansion of CD8+ memory T cells. Nature. 2006;441:890–893. doi: 10.1038/nature04790. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 162.Bachmann MF, et al. Differential role of IL-2R signaling for CD8(+) T cell responses in acute and chronic viral infections. Eur J Immunol. 2007;37:1502–1512. doi: 10.1002/eji.200637023. [DOI] [PubMed] [Google Scholar]
- 163.Casey KA, Mescher MF. IL-21 promotes differentiation of naive CD8 T cells to a unique effector phenotype. J Immunol. 2007;178:7640–7648. doi: 10.4049/jimmunol.178.12.7640. [DOI] [PubMed] [Google Scholar]
- 164.Malherbe L, et al. Vaccine adjuvants alter TCR-based selection thresholds. Immunity. 2008;28:698–709. doi: 10.1016/j.immuni.2008.03.014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 165.Klonowski KD, et al. Dynamics of blood-borne CD8 memory T cell migration in vivo. Immunity. 2004;20:551–562. doi: 10.1016/s1074-7613(04)00103-7. [DOI] [PubMed] [Google Scholar]
- 166.Stanley M, Gissmann L, Nardelli-Haefliger D. Immunobiology of human papillomavirus infection and vaccination – implications for second generation vaccines. Vaccine. 2008;26(Suppl 10):K62–K67. doi: 10.1016/j.vaccine.2008.05.066. [DOI] [PubMed] [Google Scholar]