

Abstract
The major histocompatibility complex class I complex consists of a heavy chain and a light chain (β2-microglobulin, β2m), which assemble with a short endogenously derived peptide in the endoplasmic reticulum. The class I peptide can be directly exchanged, either at the cell surface or, as recently described, in vesicles of the endocytic compartments, thus allowing exogenous peptides to enter the class I presentation pathway. To probe the interactions between the components of the class I molecule, we analyzed the exchange of peptide and β2m by using purified, recombinant H2-Kb/peptide complexes in a cell-free in vitro system. The exchange of competitor peptide was primarily dependent on the off-rate of the original peptide in the class I binding groove. Peptide exchange was not enhanced by the presence of exogenous β2m, as exchange occurred to the same extent in its absence. Thus, the exchange of peptide and β2m are independent events. The exchange rate of β2m also was not affected by the dissociation rates of the original peptides. Furthermore, peptides could substantially exchange into class I molecules over a pH range of 5.5 to 7.5, conditions prevalent in certain endocytic compartments. We conclude that the dynamic properties of the components of class I molecules explain its function as a highly peptide-receptive molecule. The major histocompatibility complex class I can readily receive peptides independent of the presence of exogenous β2m, even at a low pH. Such properties are relevant to class I peptide acquisition, which can occur at the cell surface, as well as in specialized endosomes.
In the cell-mediated immune response, the function of the major histocompatibility complex (MHC) class I molecule is to present antigenic peptides to CD8+ cytotoxic T cells. The MHC class I molecule is comprised of a polymorphic 45-kDa class I heavy chain (H chain), a noncovalently associated 12-kDa class I light chain [also called β2-microglobulin (β2m)], and a short peptide of 8–10 amino acids in length (1, 2). In the conventional route of class I antigen presentation, these peptides are derived from endogenous cellular or viral proteins after proteolytic degradation, then transport into the endoplasmic reticulum where binding to class I occurs (3, 4). However, the class I molecule also has been postulated to bind and present exogenous peptides through alternative routes (5, 6). In the cytosolic pathway, exogenous antigens are internalized into phagosomes via an endocytic mechanism and then transferred to the cytosol where they re-enter the classical pathway for peptide antigen presentation (7–10). In the noncytosolic pathway, recycled class I is thought to bind exogenous peptides in endocytic compartments, a process described in macrophages and in several cases for T cells (11–14).
The mechanism of peptide exchange onto MHC class I and the role of β2m has been explained by several models. The free H chain model has suggested that β2m enhances peptide binding through stabilizing free H chains (15–19), whereas the cooperative exchange model has proposed that the exchange of β2m liberates the bound peptide and thus allows binding of exogenous peptide (16, 18, 20), although contradictory observations of a noncooperative exchange of peptide and β2m recently have been presented (21). Despite the wide range of different methodological approaches addressing the dynamics of peptide and β2m exchange, a uniform mechanism has not yet evolved.
In view of the different cellular contexts in which MHC class I can function as a peptide-receptive molecule, we have re-examined the inherent dynamics of the class I components in an in vitro study using detergent-free, purified, and soluble H2-Kb complexes. This system uniquely allows us to analyze the process of peptide and β2m exchange on a molecular level. We tried to approach the questions as to how the nature of the peptide or the availability of free β2m may influence this exchange process, and how peptide exchange could take place at low pH, as it might occur in endocytic vesicles. The results show that the exchange of peptide and β2m are independent events and furthermore, that peptide exchange occurs even in the absence of exogenous β2m. Our findings support the idea that the class I molecule is a versatile peptide-receptive molecule able to exert its functions in a variety of cellular and cell-surface conditions.
MATERIALS AND METHODS
Complex Formation by Dialysis.
Recombinant MHC class I complexes were formed and purified as described earlier in detail (22), except that reduced glutathione was added during the refolding/dialysis step. Complexes were purified by gel-exclusion chromatography (Superdex-75) equilibrated in 10 mM potassium phosphate, pH 7.0, 150 mM NaCl operating at a flow rate of 0.75 ml/min, and further judged by SDS/PAGE, and stored in aliquots at −70°C.
Purification and Iodination of β2m.
Details for the cloning and expression of recombinant murine β2m have been published elsewhere (20). Briefly, β2m was produced as inclusion bodies in Escherichia coli, solubilized in 8.0 M urea/10 mM Tris, pH 8.5, 50 mM reduced glutathione, and then dialyzed (500 molecular weight cutoff) against 10 mM Tris, pH 8.5 with several changes over 48 hr. The β2m was concentrated and purified by gel-exclusion chromatography (Superdex-75) operating at 10 mM potassium phosphate, pH 7.0. Purified β2m (100 μg) in 160 μl of PBS was incubated with 500 μCi of 125I-NaI and one Iodo-Bead for 2 min at 25°C. Unbound iodine was removed by passage over a GF-5 desalting column.
Isoelectric Focusing (IEF).
Native IEF was performed on Ampholine PAGplate gels (pH 4.0–6.5 or 5.5–8.5) for 2.5 hr at 10°C. Sample volumes were typically 10–40 μl. Gels were fixed with trichloroacetic acid (10%) and stained with Coomassie brilliant blue G-250. Protein density and radioactive intensity were quantitated by using a Molecular Dynamics densitometer or PhosphorImager.
Exchange Incubations.
For a typical exchange incubation, purified Kb/peptide complex (2.6–4.4 nmols) assembled either with wild-type (Wt), PolyI, or PolyG peptide, as listed in Table 1, was incubated with a 1× molar excess of purified “cold” β2m, plus 1–200× molar excess of competitor peptide (E6, K6, PolyG, and PolyI as listed in Table 1). Radiolabeled I125-β2m was included where indicated (140,000–250,000 dpm/1 nmol β2m). The incubation mixture was aliquoted and final Kb/peptide complex concentrations varied between 0.3–0.8 nmol/time point. Aliquots were withdrawn at specified times (0–18 hr) at either 4°C, 25°C, or 37°C.
Table 1.
Sequences, relative binding affinities, and isoelectric points of peptides used in this study
| VSV-peptide analogs | Sequence | Relative binding affinity* | Isoelectric point (IP) |
|---|---|---|---|
| WtN52-59† | RGYVYQGL | Medium | 10.3 |
| K6 | RGYVYKGL | Medium | 10.7 |
| E6 | RGYVYEGL | Medium | 8.2 |
| PolyG | GGYGYGGL | Low | 8.4 |
| PolyI | IGYIYIGL | High | 8.2 |
Relative binding affinity was assessed in competitive folding experiments of Kb H chain and β2m together with a mixture of K6, E6, PolyG, and PolyI peptide against the Wt peptide (data not shown). The high relative binding affinity is approximately eight times higher than the medium; and the medium relative binding affinity is approximately 15 times higher than the low relative binding affinity.
The KD = 171 nmolar (0.17 μM) for the VSV Wt peptide at 37°C (35).
Materials.
Biochemicals were obtained from Sigma unless otherwise stated. Sequenal-grade urea and guanidine, BCA-protein assay kit, Iodo-Beads, and GF-5 desalting columns were obtained from Pierce. Ampholine PAGplate gels were obtained from Pharmacia. All water used was distilled before passage through a MilliQ+ apparatus.
Peptide Synthesis and Purification.
Peptides were synthesized by solid-phase method that used fluorenylmethoxycarbonyl chemistry on an Applied Biosystems 433A peptide synthesizer. All peptides were purified by reverse-phase HPLC to >95% purity with a Vydac C-18 column (2.1 or 4.6 mm × 25 cm, 300 Å) on a Hewlett Packard HP-1090M instrument. Peptides were analyzed by electrospray mass spectroscopy on a PE-Sciex API-III instrument.
RESULTS
To examine the interaction of peptide and β2m during the peptide exchange process of MHC class I molecules, we used an in vitro system, with purified and solubilized Kb H chain, radiolabeled β2m, and synthetic peptides. This system has the advantage that these characteristics can be studied in the absence of other cellular constituents or detergents. We used an IEF technique, in which the individual Kb/peptide complexes could be separated according to the net charge difference placed on the entire molecule by either the original or the competitor peptide and be visualized by Coomassie. On the same gel, by using autoradiography, labeled β2m exchange could be monitored by its binding to the preformed or newly formed Kb/peptide complexes. In contrast to previous approaches, this method allowed us to distinguish with which Kb/peptide complex the labeled β2m had exchanged.
Exchange of Competitor Peptide and Exogenous β2m Occur Simultaneously, But at Different Rates.
To study the exchange of competitor peptide with the peptides bound to the Kb complex, we used analogs of the vesicular stomatitis virus (VSV) peptide, varying in their relative affinity of binding (high, medium, and low) to the Kb class I molecule and in their net charge (isoelectric point) because of amino acid replacements of nonanchor amino acid residues of VSVN52–59 peptide. In Fig. 1A, Kb/Wt complex was incubated with the E6 competitor peptide of relatively equal affinity to the Wt. When a time course was analyzed by IEF, the Coomassie-stained bands in the upper lane showed a time-dependent decrease of Kb/Wt complex, and a concomitant increase of newly formed Kb/E6 complex (lower lane), which migrated closer toward the anode, because of the net charge difference of the Wt vs. the E6 peptide. Analysis of the identical IEF gel by autoradiography (Fig. 1B) revealed that exogenous I125-β2m rapidly exchanged with the β2m of the initial Kb/Wt complex (upper lane). As the amount of Kb/E6 complex increased (>90 min), labeled β2m was increasingly found to be associated with this newly formed complex.
Figure 1.
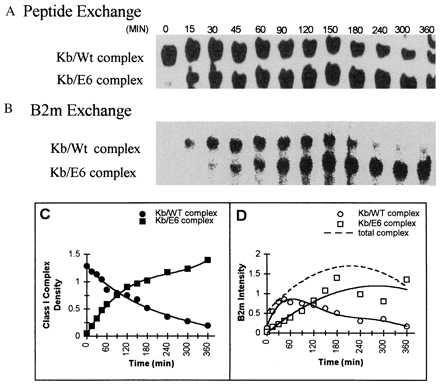
Exchange of peptide and β2m occur simultaneously but at different rates. Purified Kb/Wt complex was incubated with exogenous 125I-β2m (complex β2m/125I-β2m; 1:1) and with E6 competitor peptide (complex peptide/competitor; 1:>200) in PBS at 37°C. Aliquots were withdrawn at the specified times, analyzed by IEF gel chromatography, then visualized by Coomassie staining (A) and by phosphoimage detection (B) of the same gel. (C) The densities of the Coomassie-stained bands for Kb/Wt complex (•) and for Kb/E6 complex (▪) illustrate the E6 peptide exchange over the incubation time. The t1/2 for the Wt peptide dissociation from Kb/Wt complex is indicated to be 90 min. (D) The phosphoimage intensities for 125I-β2m, associated either to Kb/Wt complex (○) or to Kb/E6 complex (□) are compared. The t1/2 for the exchange of β2m onto the total amount of Kb/peptide complex is indicated to be ≈30 min (dotted line).
From the same IEF gel, the Coomassie bands were plotted in density units (Fig. 1C), and the autoradiograph was plotted in phosphoimage intensity units (Fig. 1D). Peptide exchanged with a t1/2 of 90 min. Assuming first-order kinetics, this exchange is a measure of the dissociation rate of the original peptide bound to the Kb/peptide complex (Fig. 1C). The exchange rate of exogenous β2m (Fig. 1D) showed a t1/2 of approximately 30 min. Significantly, the differences of the t1/2 rates of β2m and peptide exchange, as shown here for the exchange of peptides with an equal affinity, indicated that the exchange of β2m has occurred at a faster rate than peptide exchange. Thus, β2m and peptide exchange are kinetically uncoupled events.
Different Dissociation Rates of Class I Bound Peptides Have No Effect on the Exchange Rate of Exogenous β2m.
Although we found in the first set of experiments that the rate of β2m and peptide exchange were different for peptides of equal affinity to MHC, we further probed the question if the dissociation rate of peptides preloaded to class I molecules could influence the exchange rate of exogenous β2m. In these analyses, we used Kb/peptide complexes preformed with a low- or high-affinity peptide and exchanged them with a competitor peptide of medium affinity. As illustrated in Fig. 2A, Kb complex loaded with the high-affinity PolyI peptide was replaced by the medium-affinity K6 peptide with a t1/2 of 360 min. This rate is approximately 4-fold slower than the t1/2 measured in the previous experiment (Fig. 1C) where Kb/Wt was exchanged by the E6 competitor peptide. Because the competitor peptide was in excess and of equal affinity in both experiments (Figs. 1C and 2A), it is evident that the dissociation rate of the peptide prebound to the class I cleft is determining the peptide exchange rate, confirming the results of others (23). The phosphoimage intensity data (Fig. 2B), illustrating the exchange of labeled β2m, indicated a rapid exchange of exogenous β2m into the original Kb/PolyI complex with the identical t1/2 of approximately 30 min, as obtained for the Kb/Wt complex (Fig. 1D). Thus, despite the finding that the dissociation rate of the individual peptides preloaded to the class I can exhibit substantial differences, the exchange rate of β2m remains invariant.
Figure 2.
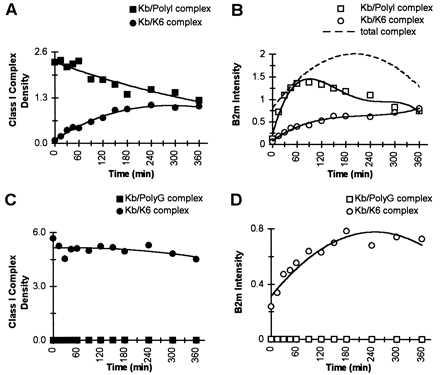
Varying dissociation rates of the class I peptides do not affect the β2m exchange rate. Either purified Kb/PolyI complex (A and B) or Kb/PolyG complex (C and D) were incubated with exogenous 125I-β2m (complex β2m/125I-β2m; 1:1) and with K6 competitor peptide (complex peptide/competitor; 1:>200) in PBS at 37°C. Aliquots were withdrawn at the specified times and analyzed by IEF gel chromatography and visualized by Coomassie staining and by phosphoimage detection of the same gel. (A) The densities of the Coomassie-stained bands for Kb/PolyI complex (▪) and the Kb/K6 complex (•) illustrate K6 peptide exchange over the incubation time; and the t1/2 for the dissociation of PolyI peptide from Kb/PolyI complex is indicated to be 6 hr. (B) The phosphoimage intensities for 125I-β2m, associated either to Kb/PolyI complex (□) or to Kb/K6 complex (○) are compared. The t1/2 for the β2m exchange onto the total amount of Kb/peptide complex is indicated to be ≈30 min (dotted line). (C) The densities of the Coomassie-stained bands for the Kb/PolyG complex (▪) and the Kb/K6 complex (•) illustrate K6 peptide exchange over the incubation time. The t1/2 for the exchange of K6 onto Kb/PolyG complex could not be measured because of the instantaneous dissociation of the prebound PolyG peptide. (D) The phosphoimage intensities for 125I-β2m, associated either to Kb/PolyG complex (□) or to Kb/K6 (○) complex are compared. As the Kb/PolyG complex was instantaneously exchanged into Kb/K6 complex, the data reflect the exchange of labeled β2m onto Kb/K6 complex with a t1/2 of ≈30 min.
When the Kb/peptide complex is preformed with the low-affinity PolyG peptide (Fig. 2C), the PolyG peptide dissociates instantaneously in the presence of K6 competitor peptide at 37°C with a concomitant formation of Kb/K6 complex, which is stable for the duration of the time course. The phosphoimage intensity data show that labeled β2m continues to associate into Kb/K6 complex until saturation is reached (Fig. 2D). The t1/2 for the β2m exchange was about 30 min. Significantly, for all the studies described so far, which used a high-, medium-, and low-affinity peptide preloaded to the Kb complex (Figs. 1D and 2 B and D), there was no variation in the exchange rate for exogenous β2m (t1/2 = 30 min). Moreover, these data reveal that the β2m dissociation can proceed at a slower, as well as faster, rate than the peptide dissociation. Thus, it appears that the dissociation of the preloaded peptide bound to the Kb groove determines the rate of exchange of competitor peptide.
Peptide Exchange Occurs in the Absence of Exogenous β2m.
Considerable data has accumulated that suggests a role for exogenous β2m in promoting peptide loading on MHC class I molecules (16–18). We further investigated if the availability of exogenous β2m would affect the peptide exchange in our system. We incubated three different Kb/peptide complexes, preloaded with a high-, medium-, and low-affinity peptide (PolyI, Wt, and PolyG, respectively), together with their corresponding competitor peptides (K6, E6, and K6) and in the presence or absence of exogenous β2m for 90 min at 37°C (Fig. 3). The Coomassie density units revealed that peptide exchange had taken place for all three Kb/peptide complexes in spite of the lack of exogenous β2m. These results again strengthen the findings that peptide and β2m exchange are independent events. Thus, peptide exchange can effectively occur in the absence of exogenous β2m.
Figure 3.
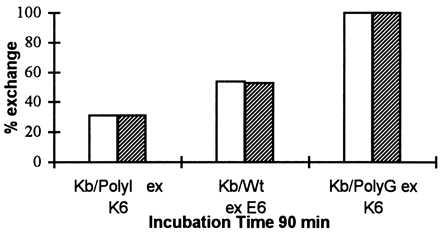
Peptide exchange occurs to the same extent in the presence or absence of exogenous β2m. Purified Kb/Wt, Kb/PolyI, and Kb/PolyG complexes were incubated with the competitor peptides E6, K6, and K6, respectively (complex peptide/competitor; 1:200), in the presence (empty bars) or absence (filled bars) of exogenous β2m (complex/β2m; 1:1) in PBS at 37°C. Aliquots were withdrawn after 90-min incubation time, analyzed by IEF gel chromatography, and visualized by Coomassie staining. The amount of the newly formed Kb/peptide complex (Kb/K6, Kb/E6, and Kb/K6) at 90 min is indicated as percentage of exchange (%), as compared with their corresponding starting material (100%) of Kb/peptide complex (Kb/PolyI, Kb/Wt, and Kb/PolyG, respectively) at time zero.
Peptide Exchange Process Can Occur Over a pH Range of 5.5 to 7.0.
To assess the effect of pH on the peptide exchange onto class I molecules, we analyzed two different Kb/peptide complexes preloaded with either the high-affinity PolyI peptide or with the medium-affinity Wt peptide over a pH range of 5.5 to 7.5 at 37°C and 3 hr of incubation. (Fig. 4). All experiments were done in the absence of exogenous β2m.
Figure 4.
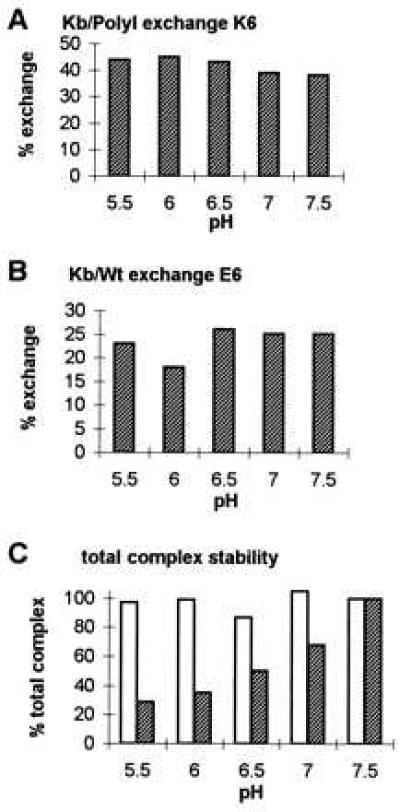
Peptide exchange occurs over a pH range from 5.5 to 7.5 for two different Kb/peptide complexes. Purified Kb/PolyI and Kb/Wt complexes were incubated with K6 (A) or E6 (B) competitor peptide, respectively (complex peptide/competitor; 1:10) in the absence of exogenous β2m in 10 mM Tris adjusted to pH values from 5.5 to 7.5. To assess Kb/PolyI and Kb/Wt complex stability over the tested pH range, the Kb/peptide complexes were incubated without competitor peptide and without β2m for the same incubation time (data not shown). Aliquots were withdrawn at time 0 (data not shown) and 3 hr of incubation at 37°C, analyzed by IEF gel chromatography, and visualized by Coomassie staining. (A) Kb/PolyI complex was exchanged by K6 competitor peptide. The amount of the newly formed Kb/K6 complex is indicated as percentage of exchange (%), as compared with the starting amount of Kb/PolyI complex (100%) at time zero. (B) The exchange of E6 competitor peptide onto Kb/Wt complex is illustrated. The percentage of exchange of newly formed Kb/E6 complex after 3 hr of incubation time is referred to the starting Kb/Wt complex (100%) at time zero. (C) The densities of the Coomassie-stained bands for the total amounts of Kb/peptide complexes, thus Kb/PolyI added to Kb/K6 (empty bars) and Kb/Wt added to Kb/E6 (filled bars), after 3 hr of incubation at 37°C illustrate complex stability over the pH range from 5.5 to 7.5. The maximal values for the two complex combinations at pH 7.5 were set at 100%. For the total Kb/Wt added to Kb/E6 complex, stability was decreased at the lower pH range (5.5 and 6.0). A similar instability was observed for the control Kb/Wt complex incubated in the absence of competitor peptide over the pH range tested (data not shown).
After incubating the Kb/PolyI complex with the K6 competitor peptide (Fig. 4A), peptide exchange took place and Kb/K6 complex was formed over the indicated pH range. Relative to the total amount of Kb/peptide complex (Kb/PolyI + Kb/K6), the percentage of peptide exchange varied only from 38% to 44% under the conditions used. The results showed that individual pH had no significant effect on the peptide exchange process and that these Kb/peptide complexes showed no instability at low pH.
As was found for the Kb/PolyI/K6 exchange, now starting with the Kb/Wt complex (Fig. 4B), peptide exchange occurred over the entire pH range to form Kb/E6 complex. The percentage of peptide exchange varied only from 18% to 26% relative to the total amount of Kb/peptide complex (Kb/Wt + Kb/E6). Although, some of the dissociated Kb from the Kb/Wt complex precipitated at pH 5.5, 6.0, and 6.5, causing a decreased amount of complex to exchange with peptide (Fig. 4C), there was no significant impact on the overall peptide exchange rate, which remained constant throughout the entire pH range. Thus, the results indicate that peptide exchange onto two differently stable Kb/peptide complexes was not affected within the physiological pH range from 5.5 to 7.5.
DISCUSSION
The classical role of the MHC class I molecule is to bind endogenously derived peptides during its biosynthetic maturation, and after translocation to the cell surface, to display an update of this peptide pool on the cell surface to cytotoxic T cells (reviewed in refs. 1 and 2). In general, peptides derived from exogenous protein sources are excluded from the class I antigen presentation pathway. However, recent data have shown that a subset of antigen-presenting cells (certain macrophages and dendritic cells) are able to display peptides derived from exogenous protein sources on MHC class I molecules and further to elicit a potent cytotoxic T cell response (reviewed in refs. 5 and 6). These exogenous peptides or their precursors can enter the cell through phagocytic uptake, and further they can gain access to the cytosol and be expressed either in a transporter associated with antigen processing (TAP)-dependent or -independent way on the cell surface. Thus, the MHC class I has to function as a peptide-receptive molecule in a number of different environments. In the endoplasmic reticulum, the H chain can assemble with β2m and peptide to form a stable trimolecular complex. Alternatively, class I molecules, in the form of trimolecular complexes or “empty” H chains, can acquire peptides through peptide exchange (14, 24–26). This peptide exchange can occur at the cell surface or, as recently proposed, in distinct vesicular compartments. Given such requirements, the question arises as to how the MHC class I molecule copes with the different amounts of free β2m, different pH levels, and peptides of different binding affinities.
To examine the interactions of the components of the MHC class I complex and their relevance to the functioning of the class I as a peptide-receptive molecule, we developed an in vitro system, free of cellular constituents, by using detergent-free, soluble recombinant H2-Kb complexes bound with different peptides (Table 1). Because of the net charge difference of complexes containing different peptides, we were able to directly measure both the incorporation of competitor peptide and the incorporation of labeled β2m into the individual Kb/peptide complexes by an IEF technique.
Peptide/H2-Kb class I combinations with three VSV peptide analogs displaying low, medium, and high affinity were probed for their capacity to exchange medium-affinity competitor peptides (Figs. 1 and 2). We found that the exchange of peptide is determined primarily by the dissociation rate of the pre-existing peptide-MHC complex. Peptides showing a high off-rate could be readily displaced from the class I binding groove by an exogenous peptide, a capability similar to the phenomenon described for the “empty” class I molecules (25, 26). This finding is in agreement with Ojcius et al. (23), who also determined the peptide off-rate to be the rate-limiting step for the binding of exogenous peptides by using an in vitro system with a soluble, truncated Kd H chain covalently linked to β2m. However, in our studies, soluble H chain and soluble β2m were not linked, permitting us to analyze the interactions of both peptide and β2m during the exchange process. We observed that the exchange of peptide was not influenced by the presence or absence of exogenous β2m (Fig. 3). Further, although each VSV peptide analog displayed a different off-rate (t1/2= 0–360 min), the exchange of peptide did not affect the exchange of β2m, which remained constant throughout (t1/2 = 30 min). We also found that the exchange of β2m was unaffected by the absence of competitor peptide (data not shown). Therefore, these two processes, the exchange of peptide and β2m, are shown to be independent events.
Our findings are in disagreement with the general perception of a cooperative nature of the exchange process in which the exchange of β2m permits the release of a bound peptide to create a free binding site (16–18, 20, 27). They also are not consistent with the idea that the t1/2 of β2m depends on the peptide already bound into the class I cleft (27). However, some peptides used in this similar study by Parker et al. (27) were extended beyond the optimal length for class I binding, whereas the peptides used in our studies were analogs of the optimal-length VSV8 mer peptide with amino acid substitutions at nonanchor residues only. Despite the different findings on the exchange rate of β2m, both studies showed exogenous peptides could be replaced independently of the β2m exchange (27). In contrast to reports using T cell-based assays, our cell- and detergent-free system allowed us to examine the dynamics of peptide and β2m exchange on a molecular level. Our data fit a model of the MHC class I as a dynamic trimolecular complex in which peptide and β2m are able to constantly associate and disassociate, and the exchange of peptide does not require a coordinate exchange of β2m. Therefore, the role of β2m seems to be to maintain the functional integrity of the MHC class I molecule.
In light of the recently described finding that exogenous peptides can assemble with internalized (11–14) or differently routed (28) MHC class I molecules in endocytic compartments, the question arises as to how pH and β2m availability influence the peptide exchange into intact class I molecules. Conventionally, class I proteins are folded together at neutral pH in the endoplasmic reticulum or they can exchange peptides on the cell surface at physiological pH. For the above reasons, we chose to analyze the effects of pH on the peptide exchange process by using either a slow (Kb/PolyI) or medium (Kb/Wt) dissociating peptide-MHC class I complex in our detergent- and cell-free system at 37°C (Fig. 4). As the amount of free β2m in such acidic vesicles is unknown, our studies were carried out in the absence of β2m. For these two different Kb/complexes, we found that the exchange of peptide occurred to the same extent for each individual Kb/peptide complex over the pH range 5.5–7.5. Although the individual Kb/peptide complexes showed a different acid sensitivity, allowing a different amount of class I molecules to become available for peptide exchange, the rate of exchange remained constant for each Kb/complex over the pH range tested. This result is in contrast to previously described studies that reported an enhanced peptide exchange at the more acidic pH environments by using detergent-solubilized, affinity-purified MHC class I complexes (29) or baculovirus-derived class I molecules (30, 31). We think possibly that the different results may be because of the use of detergents that could have a destabilizing effect on proteins at low pH (32), the use of lower temperatures (18°C) to increase complex stability (29), or that class I molecules expressed in baculovirus systems are known to already contain endogenous peptides, which could alter the dynamics of peptide exchange.
From our results, we conclude that the capability of the class I molecules to exchange peptides at the pH range 5.5–7.5 would allow the acquisition of exogenous peptides at the lower pH levels found in endosomes. Considering that early endosomes (pH 6.2) and late endocytic compartments (pH 4.5–5.5) display a different pH (33), and moreover, exhibit different peptide processing capacities (34), this capability could provide an opportunity for class I complexes of different stability to exchange exogenous peptides in these vesicles. Thus, the eventual fate of each individual MHC class I complex depends on the nature of the peptide bound into the binding groove, the class I complex stability, the pH environment of the vesicles the class I is passing through, and moreover, the range of peptides available for the exchange.
Our studies using homogenous components of the MHC class I in a soluble in vitro system have demonstrated specific dynamic properties of the class I molecule, which are important for its function as a versatile peptide-receptive molecule. The MHC class I can readily receive competitor peptides independent of the presence of exogenous β2m and even at a low pH, a process that can occur at the cell surface, as well as in specialized endosomes. These dynamic features provide an explanation for the uptake of exogenous peptides in the alternative pathway of MHC class I presentation.
Acknowledgments
We thank Drs. T. P. DiLorenzo, A. Davidson, D. C. Roopenian, M. D. Scharff, and P. E. Scherer for critical reading of the manuscript. This work was supported by National Institutes of Health Grant 5R37AI07289. H.H. was supported by a postdoctoral fellowship of the Cancer Research Institute, New York. N.J.P. was a recipient of National Institutes of Health Training Grant 5T32CA09173.
ABBREVIATIONS
- MHC
major histocompatibility complex
- H chain
class I heavy chain
- β2m
β2-microglobulin
- IEF
isoelectric focusing
- Wt
wild type
- VSV
vesicular stomatitis virus
References
- 1.Heemels M T, Ploegh H. Annu Rev Biochem. 1995;64:463–491. doi: 10.1146/annurev.bi.64.070195.002335. [DOI] [PubMed] [Google Scholar]
- 2.York I A, Rock K L. Annu Rev Immunol. 1996;14:369–396. doi: 10.1146/annurev.immunol.14.1.369. [DOI] [PubMed] [Google Scholar]
- 3.Lehner P J, Cresswell P. Curr Opin Immunol. 1996;8:59–67. doi: 10.1016/s0952-7915(96)80106-3. [DOI] [PubMed] [Google Scholar]
- 4.Hansen T H, Lee D R. Adv Immunol. 1997;64:105–137. doi: 10.1016/s0065-2776(08)60888-3. [DOI] [PubMed] [Google Scholar]
- 5.Rock K L. Immunol Today. 1996;17:131–137. doi: 10.1016/0167-5699(96)80605-0. [DOI] [PubMed] [Google Scholar]
- 6.Jondal M, Schirmbeck R, Reimann J. Immunity. 1996;5:295–302. doi: 10.1016/s1074-7613(00)80255-1. [DOI] [PubMed] [Google Scholar]
- 7.Kovacsovics-Bankowski M, Clark K, Benacerraf B, Rock K L. Proc Natl Acad Sci USA. 1993;90:4942–4946. doi: 10.1073/pnas.90.11.4942. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Kovacsovics-Bankowski M, Rock K L. Science. 1995;267:243–246. doi: 10.1126/science.7809629. [DOI] [PubMed] [Google Scholar]
- 9.Shen Z, Reznikoff G, Dranoff G, Rock K L. J Immunol. 1997;158:2723–2730. [PubMed] [Google Scholar]
- 10.Mazzaccaro R J, Gedde M, Jensen E R, van Santen H M, Ploegh H L, Rock K L, Bloom B R. Proc Natl Acad Sci USA. 1996;93:11786–11791. doi: 10.1073/pnas.93.21.11786. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Pfeifer J D, Wick M J, Roberts R L, Findlay K, Normark S J, Harding C V. Nature (London) 1993;361:359–362. doi: 10.1038/361359a0. [DOI] [PubMed] [Google Scholar]
- 12.Bachmann M F, Oxenius A, Pircher H, Hengartner H, Ashton-Richardt P A, Tonegawa S, Zinkernagel R M. Eur J Immunol. 1995;25:1739–1743. doi: 10.1002/eji.1830250637. [DOI] [PubMed] [Google Scholar]
- 13.Dasgupta J D, Watkins D, Slayter H, Yunis E J. J Immunol. 1988;141:2577–2580. [PubMed] [Google Scholar]
- 14.Schirmbeck R, Reimann J. Eur J Immunol. 1996;26:2812–2822. doi: 10.1002/eji.1830261204. [DOI] [PubMed] [Google Scholar]
- 15.Rock K L, Gamble S, Rothstein L, Gramm C, Benacerraf B. Cell. 1991;65:611–620. doi: 10.1016/0092-8674(91)90093-e. [DOI] [PubMed] [Google Scholar]
- 16.Rock K L, Gamble S, Rothstein L, Benacerraf B. Proc Natl Acad Sci USA. 1991;88:301–304. doi: 10.1073/pnas.88.1.301. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Rock K L, Rothstein L, Gamble D, Benacerraf B. Proc Natl Acad Sci USA. 1990;87:7517–7521. doi: 10.1073/pnas.87.19.7517. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Vitiello A, Potter T A, Sherman L A. Science. 1990;250:1423–1426. doi: 10.1126/science.2124002. [DOI] [PubMed] [Google Scholar]
- 19.Day P M, Esquivel F, Lukszo J, Bennink J R, Yewdell J W. Immunity. 1995;2:137–147. doi: 10.1016/s1074-7613(95)80014-x. [DOI] [PubMed] [Google Scholar]
- 20.Kozlowski S, Takeshita T, Boehncke W-H, Takahashi H, Boyd L F, Germain R N, Berzofsky J A, Margulies D H. Nature (London) 1991;349:74–77. doi: 10.1038/349074a0. [DOI] [PubMed] [Google Scholar]
- 21.Cook J R, Myers N B, Hansen T H. J Immunol. 1996;157:2256–2261. [PubMed] [Google Scholar]
- 22.Zhang W, Young A C M, Imarai M, Nathenson S G, Sacchettini J C. Proc Natl Acad Sci USA. 1992;89:8403–8407. doi: 10.1073/pnas.89.17.8403. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Ojcius D M, Abastado J-P, Casrouge A, Mottez E, Cabanie L, Kourilsky P. J Immunol. 1993;151:6020–6026. [PubMed] [Google Scholar]
- 24.Ojcius D M, Godeau F, Abastado J P, Casanova J L, Kourilsky P. Eur J Immunol. 1993;23:1118–1124. doi: 10.1002/eji.1830230521. [DOI] [PubMed] [Google Scholar]
- 25.Smith J D, Lie W-R, Gorka J, Myers N B, Hansen T H. Proc Natl Acad Sci USA. 1992;89:7767–7771. doi: 10.1073/pnas.89.16.7767. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Ljunggren H G, Stam N J, Ohlen C, Neefjes J J, Hoglund P, Heemels M T, Bastin J, Schumacher T N, Townsend A, Karre K. Nature (London) 1990;346:476–480. doi: 10.1038/346476a0. [DOI] [PubMed] [Google Scholar]
- 27.Parker K C, DiBrino M, Hull L, Coligan J E. J Immunol. 1992;149:1896–1904. [PubMed] [Google Scholar]
- 28.Sugita M, Brenner M B. J Biol Chem. 1995;270:1443–1532. doi: 10.1074/jbc.270.3.1443. [DOI] [PubMed] [Google Scholar]
- 29.Stryhn A, Pedersen L O, Romme T, Olsen A C, Nissen M H, Thorpe C J, Buus S. J Immunol. 1996;156:4191–4197. [PubMed] [Google Scholar]
- 30.Ojcius D M, Gapin L, Kourilsky P. Biochem Biophys Res Comm. 1993;197:1216–1222. doi: 10.1006/bbrc.1993.2606. [DOI] [PubMed] [Google Scholar]
- 31.Reich Z, Altman J D, Boniface J J, Lyons D S, Kozono H, Ogg G, Morgan C, Davis M M. Proc Natl Acad Sci USA. 1997;94:2495–2500. doi: 10.1073/pnas.94.6.2495. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Reay P A, Wettstein D A, Davis M M. EMBO J. 1992;11:2829–2839. doi: 10.1002/j.1460-2075.1992.tb05350.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Maxfield F R, Yamashiro D J. Adv Exp Med Biol. 1987;225:189–198. doi: 10.1007/978-1-4684-5442-0_16. [DOI] [PubMed] [Google Scholar]
- 34.Griffin J P, Chu R, Harding C V. J Immunol. 1997;158:1523–1532. [PubMed] [Google Scholar]
- 35.Saito Y, Peterson P A, Matsumura M. J Biol Chem. 1993;268:21309–21317. [PubMed] [Google Scholar]