

Abstract
Rates of mitochondrial superoxide anion radical ( ) generation are known to be inversely correlated with the maximum life span potential of different mammalian species. The objective of this study was to understand the possible mechanism(s) underlying such variations in the rate of generation. The hypothesis that the relative amounts of the ubiquinones or coenzyme Q (CoQ) homologues, CoQ9 and CoQ10, are related with the rate of generation was tested. A comparison of nine different mammalian species, namely mouse, rat, guinea pig, rabbit, pig, goat, sheep, cow, and horse, which vary from 3.5 to 46 years in their maximum longevity, indicated that the rate of generation in cardiac submitochondrial particles (SMPs) was directly related to the relative amount of CoQ9 and inversely related to the amount of CoQ10, extractable from their cardiac mitochondria. To directly test the relationship between CoQ homologues and the rate of generation, rat heart SMPs, naturally containing mainly CoQ9 and cow heart SMPs, with high natural CoQ10 content, were chosen for depletion/reconstitution experiments. Repeated extractions of rat heart SMPs with pentane exponentially depleted both CoQ homologues while the corresponding rates of generation and oxygen consumption were lowered linearly. Reconstitution of both rat and cow heart SMPs with different amounts of CoQ9 or CoQ10 caused an initial increase in the rates of generation, followed by a plateau at high concentrations. Within the physiological range of CoQ concentrations, there were no differences in the rates of generation between SMPs reconstituted with CoQ9 or CoQ10. Only at concentrations that were considerably higher than the physiological level, the SMPs reconstituted with CoQ9 exhibited higher rates of generation than those obtained with CoQ10. These in vitro findings do not support the hypothesis that differences in the distribution of CoQ homologues are responsible for the variations in the rates of mitochondrial generation in different mammalian species.
A current hypothesis of aging postulates that oxidative stress/damage is a major causal factor in the attrition of functional capacity occurring during the aging process (1–6). The basic tenet of this hypothesis is that there is an intrinsic imbalance between the reactive oxygen species (ROS),1 that are incessantly generated in the aerobic cells and the antioxidative defense against them, thereby resulting in the accrual of steady-state levels of oxidative molecular damage. The direct evidence in support of this hypothesis is that the augmentation of antioxidative defenses by simultaneous overexpression of Cu/Zn superoxide dismutase, which converts superoxide anion radicals ( ) into H2O2, and catalase, which removes H2O2, retards the age-associated increase in the levels of molecular oxidative damage and extends the life span of Drosophila melanogaster by one-third (7, 8).
Although there are several intracellular loci for the generation of (the first molecule in the ROS series), it is widely accepted that the mitochondrial electron transport chain is the main source of (9, 10). Previous studies in this laboratory have indicated that the rate of mitochondrial generation varies greatly, even in the same type of tissue, among different mammalian species and is inversely related to the maximum life span potential (MLSP) of the species (11, 12). The inverse relationship between the rate of generation and MLSP was found to hold in a sample of mammalian species as well as a group of dipteran insect species (11–13).
The question that arose out of these studies and that is also the subject of this investigation is what is the underlying mechanism for the variations in the rates of mitochondrial generation in different species? Although opinions vary (14), a number of experimental studies in the literature suggest that ubiquinones modulate the rate of mitochondrial generation (10, 15–18). Ubiquinones (2,3-dimethoxy-5-methyl-6-multiprenyl-1,4-benzoquinone), or coenzyme Q (CoQ), is a qui-none derivative with a chain of 1–12 isoprene units in the different homologue forms (CoQn) occurring in nature. Relatively short-lived mammalian species such as the mouse and the rat primarily contain CoQ9, whereas the larger long-lived mammals such as man predominantly exhibit CoQ10 (19). The present study tests the hypothesis that variations in the rate of by cardiac submitochondrial particles (SMPs) in different mammalian species are related to the relative CoQ9 and/or CoQ10 content. The hypothesis was prompted by the fact that longevity of non-primate mammalian species tends to be inversely correlated with the rate of mitochondrial generation and directly correlated with the body mass.
EXPERIMENTAL PROCEDURES
Materials
All solvents used were of HPLC grade (Fisher). Ubiqui-none-9, ubiquinone-10, (±)-tocopherol and ferricytochrome c (Type Vl), superoxide dismutase, rotenone, antimycin A, and 4,4,4-trifluoro-1-(2-thienyl)-1,3-butanedione (TTFA) were purchased from Sigma.
Animals
Hearts were obtained from mouse (Swiss), rat (Harlan Sprague Dawley), guinea pig (Hartley Albino), rabbit (New Zealand White), pig (Yorkshire), goat (Angora), sheep (Rambouillet), cow (Holstein), and horse (mixed) which range from 3.5 to 46 years in MLSP (21, 29). All the animals were young, healthy, sexually mature adult males. The approximate ages of the animals were: mouse, rat, and guinea pig, 4 months; rabbit, 7 months; pig, 6–7 months; goat and sheep, 1 year; cow and horse, 3 years. In smaller animals the entire heart was processed; however, in the pig, cow, and horse the hearts were cut into smaller pieces, and representative samples were selected. The values for the rates of generation in different species are partially based on the result of previous studies in this laboratory (12, 22). MLSP values for different species, obtained from the literature (20, 21), in years, are: mouse, 3.5; rat, 4.5; guinea pig, 7.5; rabbit 13; goat, 18; sheep, 20; pig, 27; cow, 30; horse, 46.
Isolation of Mitochondria and Preparation of SMPs
Mitochondria were isolated by differential centrifugation as described by Arcos et al. (23). Briefly, pieces of the heart were homogenized in 10 volumes (w/v) of isolation buffer containing 180 mM KCl, 0.5% bovine serum albumin, 10 mM MOPS, 10 mM EGTA Tris base, pH 7.2, at 4 °C. The homogenate was centrifuged at 1,000 × g for 10 min, and the supernatant was recentrifuged at 17,500 × g. The resulting mitochondrial pellet was washed and resuspended in 0.25 M sucrose, 1 mM EGTA, 10 mM MOPS, pH 7.2. To prepare SMPs, the mitochondrial pellet was resuspended in 30 mM potassium phosphate buffer, pH 7.0, and sonicated three times, each consisting of a 30-s pulse burst, at 1-min intervals at 4 °C. The sonicated mitochondria were centrifuged at 8,250 × g for 10 min to remove the unbroken organelles; the supernatant was recentrifuged at 80,000 × g for 45 min, and the resulting pellet was washed and resuspended in 0.1 M phosphate buffer, pH 7.4, as described previously (12).
Extraction and Quantitation of Coenzyme Q
CoQ was extracted from mitochondria using a hexane:ethanol mixture as described by Takada et al. (24). Briefly, 50 μl of mitochondrial suspension, containing ~100 μg of protein and 50 μl of double-distilled H2O were mixed with 750 μl of hexane:ethanol (5:2) for 1 min using a vortex mixture. The mixture was centrifuged for 3 min at 1,200 × g, and 450 μl of the hexane layer was collected, dried under helium, and dissolved in 100 μl of ethanol. Quantitation of ubiquinones was performed by HPLC by the method of Katayama et al. (25). The ethanol extract (10–20 μl) was chromatographed on a reverse phase C18 HPLC column (25.0 × 0.46 cm, 5 μm, Supelco), using a mobile phase consisting of 0.7% NaClO4 in ethanol:methanol:70% HClO4 (900:100:1) at a flow rate of 1.2 ml. The electrochemical and UV detectors consisted of an ESA Coulochem II and a Waters Associates Model 440 absorbance detector at a wavelength of 280 nm. The setting of the electrochemical detector was as follows: guard cell (upstream of the injector) at +200 mV, conditioning cell at −550 mV (downstream of the column), followed by the analytical cell at +150 mV. The concentrations of ubiquinones were estimated by comparison of the peak area with those of standard solutions of known concentration.
Coenzyme Q Depletion and Repletion
Submitochondrial particles were depleted of native CoQ by pentane extraction, as described by Maguire et al. (26) and selectively repleted with exogenous CoQ9or CoQ10. Aliquots of SMPs (100 μl containing ~250 μg of protein) were freeze-dried and extracted 6 times, each for 45 min at 4 °C, with 1 ml of pentane containing 15 μM α-tocopherol (for the last extraction pure pentane was used). The pentane layer was removed by centrifugation and discarded. In some cases the pentane layer was collected and brought to dryness under a stream of helium and resuspended in 100 μl of ethanol, and the CoQ content was measured by HPLC. Various amounts of CoQ9 or CoQ10 were added to pentane-extracted and/or freeze-dried SMPs, dried under helium, resuspended in 100 mM potassium phosphate buffer (pH 7.4), and sonicated for up to 3 s in a Branson 2200 sonicator. CoQ depletion and incorporation of exogenous CoQ9 or CoQ10 into SMPs membranes were confirmed by hexane:ethanol extraction and HPLC analysis, as described above.
Measurement of Superoxide Anion Radical Generation ( )
Rate of generation by SMPs was measured as superoxide dismutase-inhibitable reduction of acetylated ferricytochrome c (27), as described previously (12). The reaction mixture contained 10 μM acetylated ferricytochrome c, 6 μM rotenone, 1.2 μM antimycin A, 100 units of superoxide dismutase/ml (in the reference cuvette), and 10–100 μg of SMP protein in 100 mM potassium phosphate buffer, pH 7.4. The reaction was started by adding 7.5 mM succinate, and the reduction of acetylated ferricytochrome c was followed at 550 nm.
Measurement of Oxygen Consumption
The rate of respiration of submitochondrial particles was measured polarographically using a Clark-type electrode at 37 °C. The incubation mixture, to measure state 4 respiration, consisted of buffer (154 mM KCl, 3 mM MgCl2, 10 mM KPO4, 0.1 mM EGTA, pH 7.4) and 30–100 μg of SMP protein; 7 mM succinate and/or 7 mM NADH were used as substrates, and 1.2 μM antimycin A, 6 μM rotenone, or 0.5 mM TTFA were employed as specific respiratory inhibitors.
RESULTS
Variations in the Distribution of CoQ Homologues in Mitochondria of Different Species
Comparisons of the concentrations of CoQ9 and CoQ10 extracted from the heart mitochondria were made in nine different mammalian species, namely mouse, rat, guinea pig, rabbit, pig, goat, sheep, cow, and horse. The data, presented in Table I and Fig. 1, indicate that both the total as well as the relative concentrations of CoQ9 and CoQ10 in heart mitochondria vary greatly in different species. The total concentration of mitochondrial CoQ, i.e. CoQ9 + CoQ10, varied about 2-fold in different species with the rank order: horse > mouse > cow > sheep = goat > rat > pig > rabbit > guinea pig. Although all nine species examined in this study contained both CoQ9 and CoQ10 the ratio of CoQ10/CoQ9 varied > 600-fold. In species such as the mouse and the rat almost 90% of mitochondrial CoQ occurred as CoQ9 while in the guinea pig CoQ9 and CoQ10 were present in roughly equal amounts. In mitochondria from rabbit, pig, goat, sheep, cow, and horse, CoQ10 was the predominant form, with CoQ9 constituting ~1.3 to 4.0% of the total CoQ content.
Table I.
Comparison of CoQ9 and CoQ10 content of heart mitochondria between nine different mammalia species
| Species | Coenzyme Q |
||
|---|---|---|---|
| CoQ9 | CoQ10 | CoQ9 + CoQ10 | |
| nmol/mg mitochondrial protein | |||
| Mouse | 6.0 ± .04 | 0.72 ± .04 | 6.72 |
| Rat | 5.2 ± .11 | 0.63 ± .02 | 5.83 |
| Guinea pig | 1.9 ± .006 | 1.7 ± .02 | 3.6 |
| Rabbit | 0.11 ± .01 | 4.8 ± .10 | 4.91 |
| Pig | 0.19 ± .007 | 4.8 ± .04 | 4.99 |
| Goat | 0.25 ± .03 | 6.2 ± .05 | 6.45 |
| Sheep | 0.15 ± .01 | 6.3 ± .11 | 6.45 |
| Cow | 0.11 ± .006 | 6.5 ± .18 | 6.51 |
| Horse | 0.17 ± .01 | 6.6 ± .08 | 6.77 |
Values are average ± S.E. of three determinations.
Fig. 1. Relationship between the rates of generation by cardiac submitochondrial particles from different species and the amounts of CoQ9 (A) and CoQ10 (B), extractable from heart mitochondria.
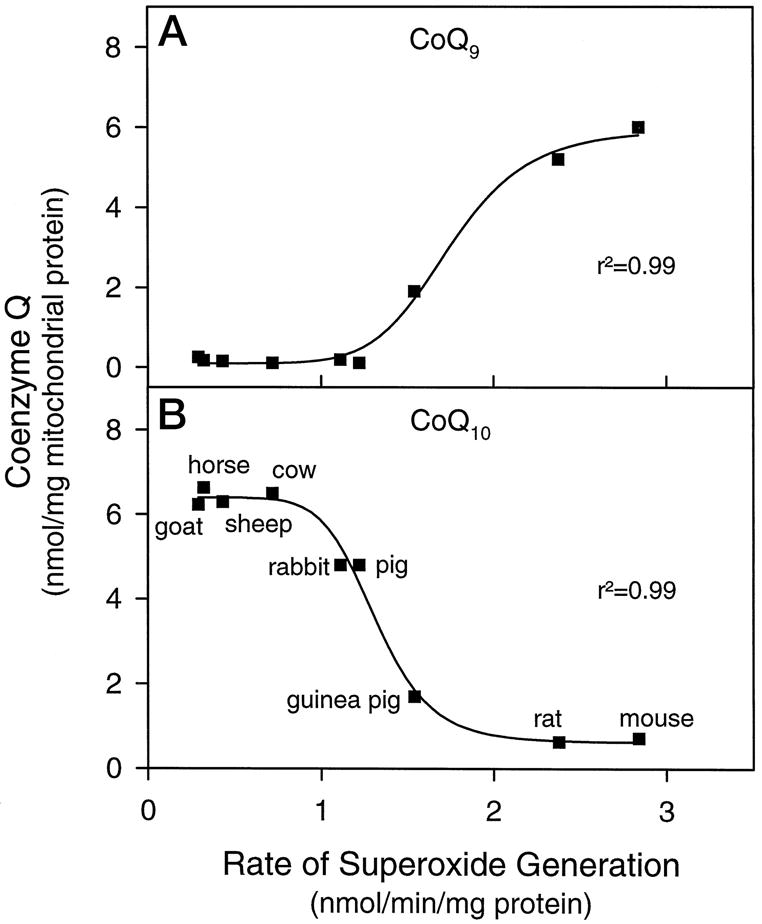
The rates of generation were measured as superoxide dismutase-inhibitable reduction of acetylated ferricytochrome c, as reported previously (12, 22). The reaction mixture contained 10 μM acetylated ferricytochrome c, 6 μM rotenone, 1.2 μM antimycin A, and 20 μg of SMP protein. Nonlinear regression analysis for the sigmoid curve fits was performed using 4-parameter logistic function (SigmaPlot 2.0; Jandel Corp., San Rafael, CA).
Correlation between CoQ and Superoxide Anion Radical Generation
To determine the relationship between mitochondrial CoQ content and the rate of generation in different species, the amounts of CoQ9 and of CoQ10 were plotted against the average rates of generation by SMPs, partially determined in the context of previous studies (12). As shown in Fig. 1A, the amount of CoQ9 was directly correlated and that of CoQ10 was inversely correlated (Fig. 1B) with the rate of generation in different species. There was no correlation between the total amount of CoQ content and the rate of generation (data not presented).
The results of the correlational study, presented in Fig. 1, led to the question of whether the relationship between mitochondrial CoQ homologues and the rate of generation was purely coincidental or causally related. To investigate the possible existence of a causal relationship, SMPs were experimentally depleted of native CoQ and reconstituted with either CoQ9 or CoQ10. These studies were conducted on cardiac SMPs of rat, a short-lived species containing relatively high amounts of CoQ9, and of cow, a representative of the long-lived species containing relatively high CoQ10 content.
Effect of Depletion of CoQ Homologues on the Rates of Oxygen Consumption and Generation in Rat Heart SMPs
Repeated extractions with pentane were found to exponentially deplete the amount of native CoQ9 from the rat heart SMPs (Fig. 2, inset); the amount remaining after six serial extractions was about 4.5% of the total amount extractable by hexane. In contrast, apparently due to the much lower natural content of CoQ10, only three extractions with pentane were sufficient to deplete SMPs of CoQ10 to a level below the detection threshold of 0.2 μM (i.e. 0.015 nmol/mg of SMP protein).
Fig. 2. Rates of oxygen consumption in rat heart SMPs after partial depletion of CoQ.
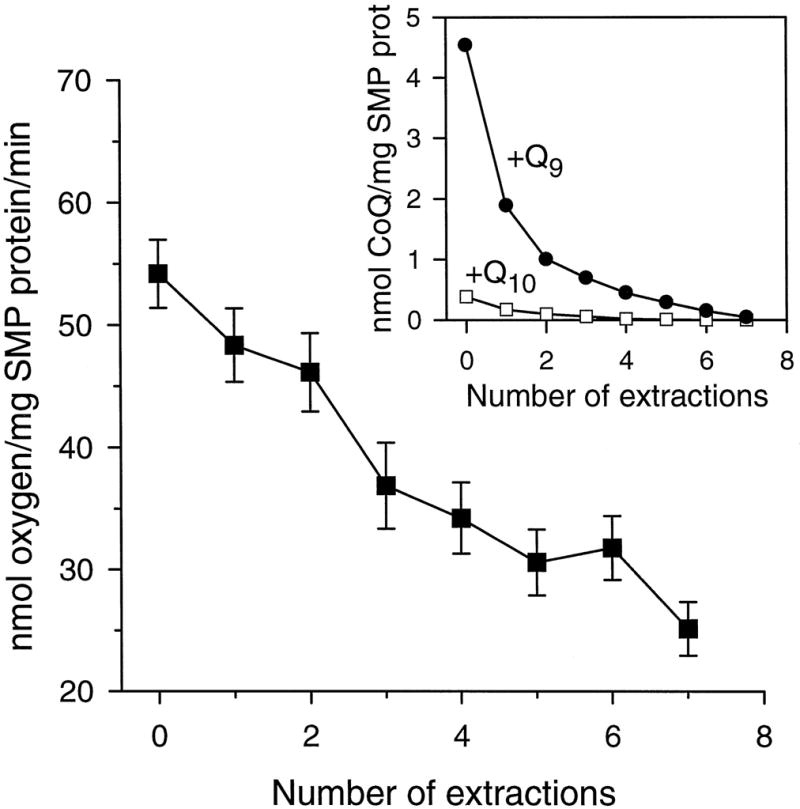
Freeze-dried SMPs were depleted of their native CoQ homologues by repeated pentane extractions. After each extraction procedure, the rate of oxygen consumption was measured polarographically with a Clark-type electrode using 7 mM succinate as a substrate. The inset shows the SMP CoQ content remaining after each extraction procedure with pentane. The (remaining) CoQ content was determined in hexane:ethanol extracts by HPLC. Data are mean ± S.E. of three independent experiments. S.E. is not shown in the inset.
To determine the effect of pentane extractions on the functional state of the SMPs, rates of oxygen consumption and generation were determined after each extraction procedure. The rate of succinate-supplemented oxygen consumption was highest in the unextracted SMPs, decreasing linearly following each extraction procedure, reaching 25% of the initial value after seven successive extraction procedures (Fig. 2). Addition of antimycin A and TTFA greatly reduced (to <2%) the rate of oxygen consumption by the depleted SMPs, whereas rotenone had no effect, indicating that O2 consumption observed was specifically due to succinate oxidase activity. NADH did not, in most instances, stimulate the rate of oxygen consumption by the depleted SMPs.
A similar study was conducted on the effect of various pentane extractions on the rate of generation by the SMPs. Again, the rate of generation was highest in the unextracted SMPs and progressively declined with each sequential pentane extraction, reaching 45% of the control value after six extraction procedures, where less than 5% of the original CoQ was present (Fig. 3).
Fig. 3. Rates of generation in rat heart SMPs following CoQ depletion.
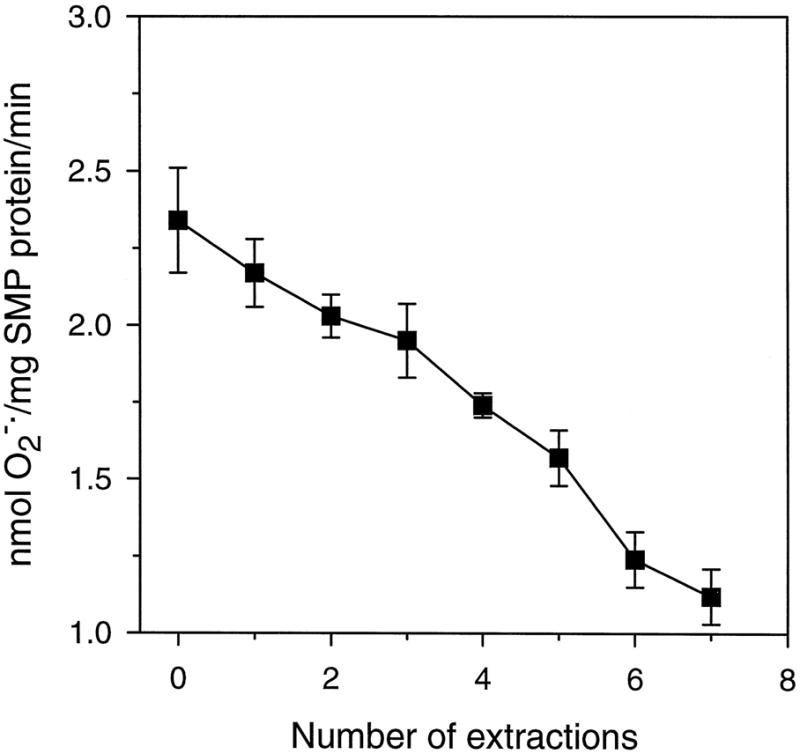
Freeze-dried SMPs were depleted of their native CoQ homologues by repeated pentane extractions. After each extraction procedure, the rate of generation was measured as superoxide dismutase-inhibitable reduction of acetylated ferricytochrome c as described in the legend of Fig. 1. CoQ content of SMPs, after successive extractions, is shown in Fig. 2, inset. Data are mean ± S.E. of three independent experiments.
Overall the results of the depletion experiments indicated that even after six or seven serial extractions with pentane the SMPs exhibited succinate oxidase activity and were able to generate albeit at rates lower than the unprocessed SMPs.
Effects of Reconstitution of Rat Heart SMPs with CoQ Homologues
Rat heart SMPs that had been extracted with pentane six times, as described above, were reconstituted with different amounts of CoQ9 and CoQ10. Reconstitution with increasing amounts of CoQ9 or CoQ10 caused an initial steep increase in the succinate-supplemented rate of oxygen consumption, which was followed by a plateau (Fig. 4B, inset). No significant differences in the rates of oxygen consumption were observed between the SMPs reconstituted with equal amounts of CoQ9 or CoQ10.
Fig. 4. Rates of oxygen consumption and generation in CoQ-depleted/reconstituted rat heart SMPs.
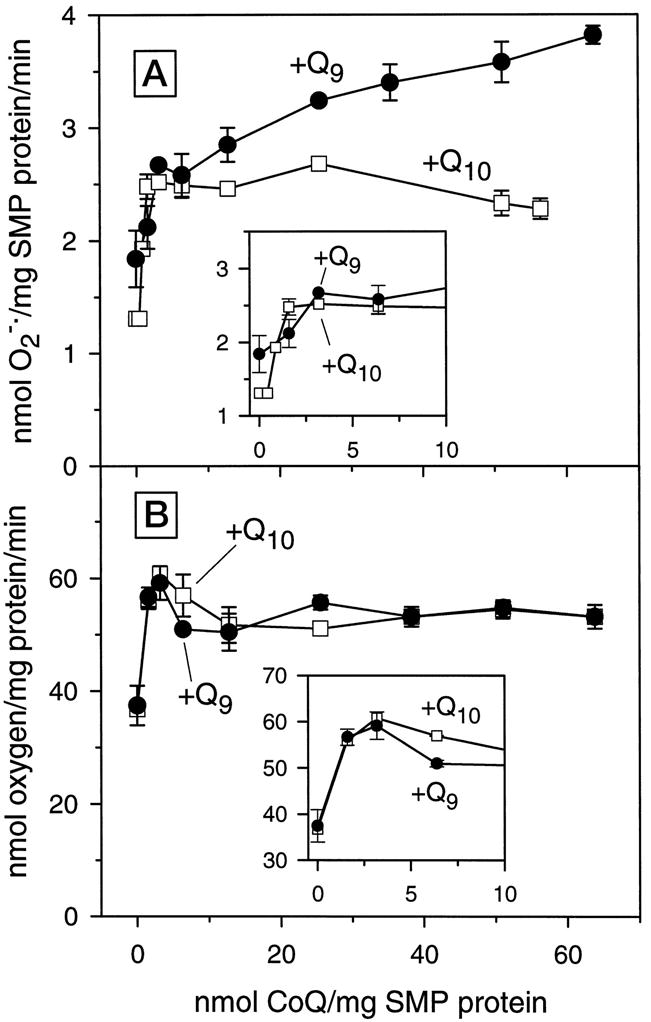
Freeze-dried SMPs were depleted of native CoQ homologues by six repeated pentane extractions and reconstituted with specific amounts of CoQ9 or CoQ10 in pentane. The reconstituted SMPs were dried and suspended in phosphate buffer, and rates of generation, shown in A, were measured as superoxide dismutase-inhibitable reduction of acetylated ferricytochrome c. Rates of oxygen consumption, shown in B, were determined polarographically with a Clark-type electrode using 7 mM succinate as a substrate. The insets depict the relationship between rates of generation and oxygen consumption by SMPs and CoQ concentrations, within the physiological range. Data are mean ± S.E. of three independent experiments.
Reconstitution of the depleted rat heart SMPs with increasing amounts of CoQ9 resulted in an initial sharp rise in the rate of generation, followed by a more gradual increase (Fig. 4A, inset). At the highest concentration of repleted CoQ9 used in this study, the rate of generation increased about 2-fold as compared with the depleted SMPs. Addition of increasing amounts of CoQ10 to the depleted SMPs also caused an initial steep rise in the rate of generation, but unlike CoQ9 further additions did not result in correspondingly increased rates of generation (Fig. 4A). For example, rat heart SMPs reconstituted with 50 nmol of CoQ9 exhibited a rate of generation that was 40% higher than when an equal amount of CoQ10 was used for reconstitution. The rate of succinate-supplemented oxygen consumption and generation by SMPs, reconstituted with different amounts of CoQ9 or CoQ10, were virtually undetectable following the addition of TTFA, demonstrating the involvement of electron transport chain and succinate oxidase activity in these two functions. It should however be noted that up to and within the physiological range of CoQ concentrations there were no differences in the rates of generation between SMPs reconstituted with CoQ9 or CoQ10 (Fig. 4A, inset). The differences between CoQ9- and CoQ10-reconstituted SMPs emerged only at concentrations considerably greater than the in vivo level (Table I).
Effects of Reconstitution with CoQ9 and CoQ10 on Bovine Heart SMPs
In contrast to the rat, bovine cardiac SMPs contain a relatively high amount of CoQ10 and a small amount of CoQ9 (see Table I). Depletion of bovine SMPs by six serial extractions with pentane achieved a 96% extraction of CoQ10 and virtually the entire amounts of the detectable CoQ9. Reconstitution of these depleted SMPs with varying concentrations of CoQ9 or CoQ10 indicated different patterns for the two omologues for the rate of oxygen consumption and of generation. Augmentation of SMPs with relatively low amounts of CoQ9 or CoQ10 caused a sharp increase in both the rate of oxygen consumption (Fig. 5B, inset) and generation (Fig. 5A, inset), but at higher concentrations these rates leveled off. Within the physiological range of CoQ content (Table I), there were no differences in the rates of generation between SMPs reconstituted with CoQ9 or CoQ10 (Fig. 5A, inset). However, the maximal rates of generation were greater for CoQ9 than for CoQ10. For example, in the bovine SMPs, reconstituted with 50 nmol of CoQ9, the rate of generation was 35% greater than in the SMPs reconstituted with an equal amount of CoQ10 (Fig. 5A).
Fig. 5. Rates of oxygen consumption and generation in CoQ-depleted/reconstituted bovine heart SMPs.
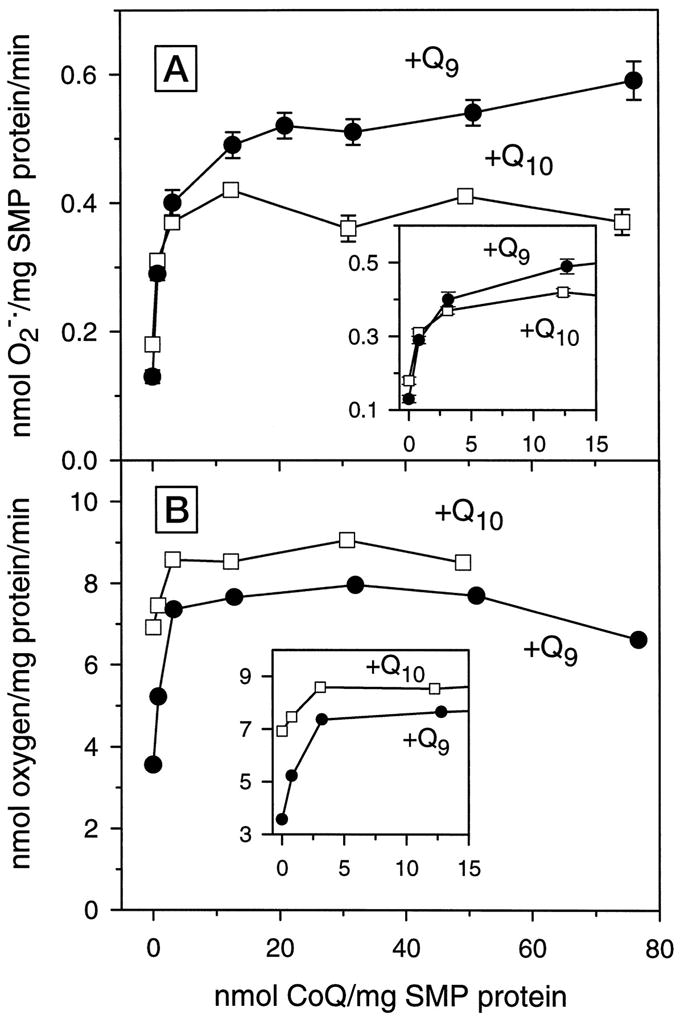
Freeze-dried SMPs were depleted of native CoQ by six repeated extractions with pentane and reconstituted with specific amounts of CoQ homologues as described in Fig. 4, and the rates of generation, shown in A, were measured. Plot B shows the rates of succinate-supplemented oxygen consumption of the SMPs from the same set of experiments. The insets depict the relationship between rates of generation and oxygen consumption by SMPs and CoQ concentrations, within the physiological range. Data are mean ± S.E. of three independent experiments; S.E. is not shown in the inset.
To further determine whether CoQ9 and CoQ10 content above the in vivo level had a different effect on the rate of generation, freeze-dried unextracted bovine SMPs were augmented with CoQ9 or CoQ10. As shown in Fig. 6, the rates of generation were stimulated to a greater extent by the addition of CoQ9 than CoQ10. The differences in the rate of generation tended to increase with the augmented amounts of CoQ9 or CoQ10 in the SMPs.
Fig. 6. Rates of generation in CoQ-augmented bovine heart SMPs.
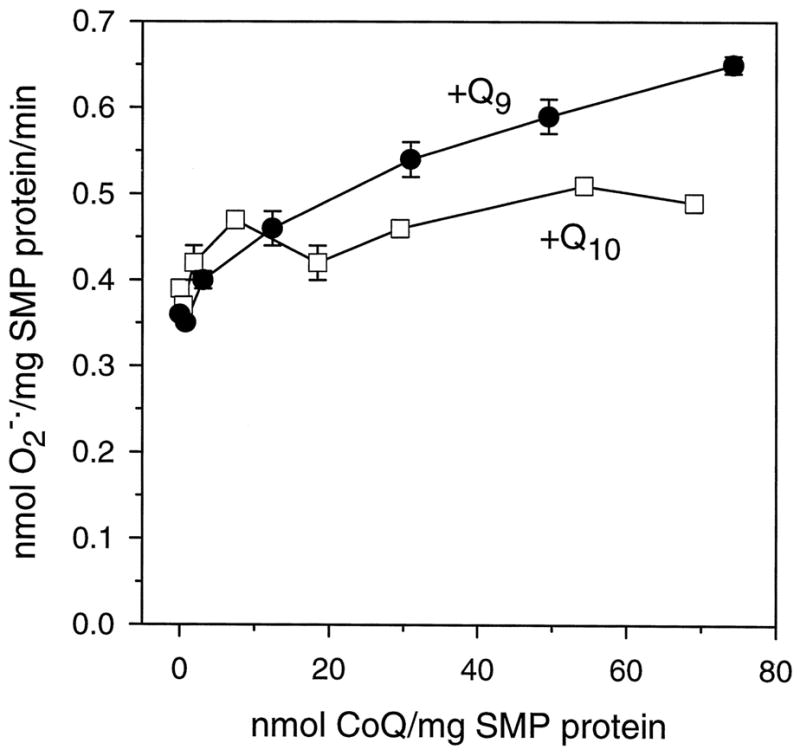
Freeze-dried unextracted SMPs were supplemented with known amounts of CoQ9 or CoQ10 in pentane, dried, and suspended in phosphate buffer. Rates of generation were measured as superoxide dismutase-inhibitable reduction of acetylated ferricytochrome c. Data are mean ± S.E. of three independent experiments.
DISCUSSION
Results of this study indicated that rates of generation in different mammalian species are directly correlated with the amounts of mitochondrial CoQ9 and are inversely related to the CoQ10 content. The in vitro depletion/reconstitution studies do not, however, support the hypothesis that variations in the relative concentrations of CoQ9 and CoQ10 are directly involved in the modulation of rates of generation in mitochondria. Although it is widely believed that the components of the mitochondrial respiratory chain, located within the domains of NADH- and succinate-cytochrome c reductase, are the main sites of generation (9, 10), there are at least two different schools of thought about whether or not autoxidation of ubisemiquinone is indeed the actual source of generation. The view, implicating ubisemiquinone (15–18), is based on the following evidence: (i) rates of generation by SMPs and/or mitochondria are highest in the presence of rotenone and antimycin A with succinate as the substrate (9); (ii) bovine heart SMPs, depleted of endogenous CoQ and reconstituted with variable amounts of exogenous CoQ, exhibit a linear relationship with the amount of quinone added (15, 16); (iii) rates of generation by isolated NADH-ubiquinone reductase particles, supplemented with different CoQ homologues, were found to be linearly dependent on the amount of exogenous CoQ (17); and (iv) in reconstituted SMPs, activities of succinate dehydrogenase and succinate-cytochrome c reductase reached a plateau at relative low concentrations of reducible CoQ, whereas the rate of H2O2 generation was linearly related to a higher range of CoQ concentration (16). In the opposing view (14), such observations do not necessarily establish a direct association between autoxidation of ubisemiquinone and generation. It is argued that such findings are also compatible with the interpretation that the iron-sulfur centers in mitochondrial respiratory complex I (NADH- ubiquinone reductase), II (succinate-ubiquinone reductase), or III (ubiquinol-cytochrome c reductase) may be the sources of generation. While the results of the present study are insufficient to resolve this controversy, they however support the view that CoQ is directly or indirectly associated with the modulation of the rates of generation, since experimental variations in its content have a clear effect on the rate of generation.
Results of this study should be interpreted in light of the fact that the preparatory procedures involving freeze-drying and pentane extractions, although widely employed (15–17, 26, 27), have an irreversible effect on the functional state of SMPs. For example, freeze-drying of SMPs followed by depletion and reconstitution with the original (natural) amount of CoQ decreased the rate of generation by about 30% and of oxygen consumption by about 60%. However, the rates of generation and oxygen consumption of pentane-extracted SMPs, reconstituted with relatively high concentrations of CoQ9 or CoQ10, reached the level comparable to the unextracted freeze-dried preparations. In both the rat and the bovine heart SMPs there were no differences in the rates of generation between SMPs reconstituted with the in vivo amounts of CoQ9 or CoQ10. Only at concentrations higher than those present under physiological conditions relatively higher rates of generation were obtained with CoQ9 than with CoQ10. The same tendency was observed when unextracted bovine SMPs were augmented with CoQ9 or CoQ10. Altogether results of depletion/reconstitution and/or augmentation studies suggest that CoQ9 and CoQ10 differ in their in vitro effect on the rate of generation only at concentrations that exceed the in vivo amounts.
A question arising from this study and also having some bearing on the interpretation of the present results concerns the nature of the structural differences between CoQ9 and CoQ10 molecules that apparently exert an effect on the rate of generation, albeit at high concentrations. Indeed, the molecular structural differences between CoQ9 (C54H82O4; Mr 794; melting point 44–45 °C) and CoQ1O (C59H90O4; Mr 862; melting point 49 °C) (28) are relatively minor. Nevertheless, the relative length of the polyisoprenoid chain and the resultant effects on the hydrophobicity of the molecule have been shown to have an effect on the location of the molecule within the phospholipid bilayer of the cell membrane. Although the relative position of CoQ9 and CoQ10 in the phospholipid bilayer has not been precisely determined, the CoQ homologues with relatively short polyisoprenoid chains are believed to lie closer to the surface of the bilayer, whereas the long chained ones are thought to be nearer to the center of the bilayer (29–32). For example, studies by Kagan et al. (33) have shown that short chain ubiquinols are relatively more efficient in inhibiting Fe2+-ascorbate-induced lipid peroxidation, suggesting that the polyisoprenoid chain length has an effect on the interaction between the quinols and the ROS present in the aqueous phase.
Studies by Matsura et al. (34) on rat and guinea pig hepatocytes also implicate major differences in antioxidant efficiency between the reduced CoQ9 and the reduced CoQ10. In response to the hydrophilic radical initiator, 2–2-azobis-(2-amidinopropane) dihydrochloride, CoQ9 was found to be preferentially oxidized as compared with CoQ10 homologue and thus may be more accessible to ROS present in the surrounding aqueous phase. This mechanism, albeit hypothetical, may underlie the relatively higher rates of generation in highly CoQ9-rich SMPs observed in this study.
Functional differences between CoQ9 and CoQ10 have also been reported by Edlund et al. (35), who found that treatment of mumps virus-infected cultured neurons with CoQ10 protected the cells from degeneration, whereas no effects were observed in response to CoQ9 treatment. Results of the present in vitro studies demonstrate that CoQ9 and CoQ10 homologues can differentially affect the rate of generation at high concentrations; however, the in vivo variations in rates of mitochondrial generation among different mammalian species cannot be explained on the basis of relative CoQ9 or CoQ10 content.
Footnotes
The abbreviation used are: ROS, reactive oxygen species; CoQ9, coenzyme Q9; CoQ10, coenzyme Q10; MLSP, maximum life span potential; , superoxide anion radical; SMPs, submitochondrial particles; TTFA, 4,4,4-trifluoro-1-(2-thienyl)-1,3-butanedione; HPLC, high performance liquid chromatography; MOPS, 4-morpholinepropanesulfonic acid.
This work was supported by the NIA, National Institutes of Health Grants R01 AG7657 and RO1 AG13563.
References
- 1.Harman D. J Gerontol. 1956;11:298–300. doi: 10.1093/geronj/11.3.298. [DOI] [PubMed] [Google Scholar]
- 2.Sohal RS, Weindruch R. Science. 1996;273:59–63. doi: 10.1126/science.273.5271.59. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Sohal RS, Orr WC. In: Molecular Aspects of Aging. Esser K, Martin GM, editors. John Wiley; Chichester, UK: 1995. pp. 109–127. [Google Scholar]
- 4.Stadtman ER. Science. 1992;257:1220–1224. doi: 10.1126/science.1355616. [DOI] [PubMed] [Google Scholar]
- 5.Stadtman ER, Starke-Reed PE, Oliver CN, Carney JM, Flyod RA. In: Free Radicals and Aging. Emerit I, Chance B, editors. Birkhauser; Basel: 1992. pp. 64–72. [Google Scholar]
- 6.Ames BN, Shigenaga MK, Hagen TM. Proc Natl Acad Sci U S A. 1993;90:7915–7922. doi: 10.1073/pnas.90.17.7915. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Orr WC, Sohal RS. Science. 1994;263:1128–1130. doi: 10.1126/science.8108730. [DOI] [PubMed] [Google Scholar]
- 8.Sohal RS, Agarwal A, Agarwal S, Orr WC. J Biol Chem. 1995;270:15671–15674. doi: 10.1074/jbc.270.26.15671. [DOI] [PubMed] [Google Scholar]
- 9.Chance B, Sies H, Boveris A. Physiol Rev. 1979;59:527–603. doi: 10.1152/physrev.1979.59.3.527. [DOI] [PubMed] [Google Scholar]
- 10.Turrens JF, Alexandre A, Lehinger AL. Arch Biochem Biophys. 1985;237:408–414. doi: 10.1016/0003-9861(85)90293-0. [DOI] [PubMed] [Google Scholar]
- 11.Sohal RS, Svensson l, Sohal BH, Brunk UT. Mech Ageing Dev. 1989;49:129–135. doi: 10.1016/0047-6374(89)90096-1. [DOI] [PubMed] [Google Scholar]
- 12.Ku HH, Brunk UT, Sohal RS. Free Radical Biol Med. 1993;15:621–627. doi: 10.1016/0891-5849(93)90165-q. [DOI] [PubMed] [Google Scholar]
- 13.Sohal RS, Sohal BH, Orr WC. Free Radical Biol Med. 1995;19:499–504. doi: 10.1016/0891-5849(95)00037-x. [DOI] [PubMed] [Google Scholar]
- 14.Beyer RE. Free Radical Biol Med. 1990;8:545–565. doi: 10.1016/0891-5849(90)90154-b. [DOI] [PubMed] [Google Scholar]
- 15.Boveris A, Chance B. Biochem J. 1973;134:707–716. doi: 10.1042/bj1340707. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Boveris A, Cadenas E, Stoppani AOM. Biochem J. 1976;156:435–444. doi: 10.1042/bj1560435. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Cadenas E, Boveris A, Ragan CL, Stoppani AOM. Arch Biochem Biophys. 1977;180:248–257. doi: 10.1016/0003-9861(77)90035-2. [DOI] [PubMed] [Google Scholar]
- 18.Turrens JF, McCord JM. In: Free Radicals, Lipoproteins, and Membrane Lipids. Paulet AC, Douste-Blazy L, Paoletti R, editors. Plenum Press; New York: 1990. pp. 203–212. [Google Scholar]
- 19.Ramasarma T. In: Coenzyme Q. Lenaz G, editor. John Wiley; London: 1985. pp. 67–81. [Google Scholar]
- 20.Comfort A. The Biology of Senescence. Elsevier Science Inc; New York: 1979. pp. 60–63. [Google Scholar]
- 21.Altman PL, Dittmer DS. Biology Data Book. 2. Vol. 1. Federation of American Societies for Experimental Biology; Bethesda, MD: 1972. pp. 229–233. [Google Scholar]
- 22.Ku HH, Sohal RS. Mech Ageing Dev. 1993;72:67–76. doi: 10.1016/0047-6374(93)90132-b. [DOI] [PubMed] [Google Scholar]
- 23.Arcos JC, Sohal RS, Sun SC, Argus MF, Bruch GE. Exp Mol Pathol. 1968;8:49–65. doi: 10.1016/0014-4800(68)90005-1. [DOI] [PubMed] [Google Scholar]
- 24.Takada M, Ikenoya S, Yuzuriha T, Katayama K. Methods Enzymol. 1984;105:147–155. doi: 10.1016/s0076-6879(84)05020-5. [DOI] [PubMed] [Google Scholar]
- 25.Katayama K, Takada M, Yuzuriha T, Abe K, Ikenoya S. Biochem Biophys Res Commun. 1980;95:971–977. doi: 10.1016/0006-291x(80)91568-5. [DOI] [PubMed] [Google Scholar]
- 26.Maguire JT, Kagan V, Ackress BAC, Serbinova E, Packer L. Arch Biochem Biophys. 1992;292:47–53. doi: 10.1016/0003-9861(92)90049-3. [DOI] [PubMed] [Google Scholar]
- 27.Ernster L, Dallner G. Biochim Biophys Acta. 1995;1271:195–204. doi: 10.1016/0925-4439(95)00028-3. [DOI] [PubMed] [Google Scholar]
- 28.Hatefi Y. Adv Enzymol. 1963;25:275–328. doi: 10.1002/9780470122709.ch5. [DOI] [PubMed] [Google Scholar]
- 29.Katsikas H, Quinn PJ. Biochim Biophys Acta. 1982;689:363–369. doi: 10.1016/0005-2736(82)90270-x. [DOI] [PubMed] [Google Scholar]
- 30.Fato R, Battino M, Castelli GP, Lenaz G. FEBS Lett. 1985;179:238–242. doi: 10.1016/0014-5793(85)80526-3. [DOI] [PubMed] [Google Scholar]
- 31.Lenaz G, Espoti MD. In: Biochemistry, Bioenergetics and Clinical Applications of Ubiquinone. Lenaz G, editor. John Wiley; New York: 1985. pp. 83–106. [Google Scholar]
- 32.Ulrich EL, Girvin ME, Cramer WA, Markley JL. Biochemistry. 1985;24:2501–2508. doi: 10.1021/bi00331a016. [DOI] [PubMed] [Google Scholar]
- 33.Kagan V, Serbinova EA, Koynova GM, Kitanova SA, Tyurin VA, Stoytchev TS, Quinn PJ, Packer L. Free Radical Biol Med. 1990;9:117–126. doi: 10.1016/0891-5849(90)90114-x. [DOI] [PubMed] [Google Scholar]
- 34.Matsura T, Yamada K, Kawaski T. Biochim Biophys Acta. 1992;1123:309–315. doi: 10.1016/0005-2760(92)90012-k. [DOI] [PubMed] [Google Scholar]
- 35.Edlund C, Holmberg K, Dallner G, Norrby E, Kristensson K. J Neurochem. 1994;63:634–639. doi: 10.1046/j.1471-4159.1994.63020634.x. [DOI] [PubMed] [Google Scholar]