

Abstract
A bioactive macrophage factor, the polypeptide daintain/allograft inflammatory factor 1 (AIF1), has been isolated from porcine intestine. It was discovered when searching for intestinal peptides with effects on insulin release, and its purification was monitored by the influence of the peptide fractions on pancreatic glucose-induced insulin secretion. Daintain/AIF1 is a 146-aa residue polypeptide with a mass of 16,603 Da and an acetylated N terminus. An internal 44-residue segment with the sequence pattern –KR–KK–GKR– has a motif typical of peptide hormone precursors, i.e., dibasic sites for potential activation cleavages and at the sequentially last such site, the structure GKR. The latter is a signal for C-terminal amide formation in the processing of peptide hormones. Daintain/AIF1 is immunohistochemically localized to microglial cells in the central nervous system and to dendritic cells and macrophages in several organs. A particularly dense accumulation of daintain/AIF1-immunoreactive macrophages was observed in the insulitis affecting the pancreatic islets of prediabetic BB rats. When injected intravenously in mice, daintain/AIF1 at 75 pmol/kg inhibited glucose (1 g/kg)-stimulated insulin secretion, with a concomitant impairment of the glucose elimination, whereas at higher doses (7.5 and 75 nmol/kg), daintain/AIF1 potentiated glucose-stimulated insulin secretion and enhanced the glucose elimination. Its dual influence on insulin secretion in vivo at different peptide concentrations, and the abundance of macrophages expressing daintain/AIF1 in the pancreatic islets of prediabetic rats, suggest that daintain/AIF1 may have a role in connection with the pathogenesis of insulin-dependent diabetes mellitus.
Several polypeptides initially isolated from the intestines have been shown to modulate the secretion of pancreatic hormones. Peptides stimulating insulin secretion, like GIP and GLP-1, have been proposed as candidate incretins (1, 2), whereas peptides inhibiting insulin secretion, like somatostatin, galanin, DBI, and PEC-60, have been suggested to be involved in the intra-islet regulation of insulin secretion (3–6). However, the majority of these peptides have other role(s) as well. During our continued search for bioactive gastrointestinal peptides, we discovered a polypeptide in porcine intestines, and initially called it daintain from a combination of Chinese and English: da, big; in, influencing; tai, peptide; in, insulin-secretion, thus “a well-sized peptide influencing insulin secretion.” Purification and some data were preliminarily reported at a conference (7). Later, it has become obvious that the amino acid sequence is similar to that of allograft inflammatory factor-1 (AIF1) characterized by cDNA cloning (8–10). Therefore, we call the peptide daintain/AIF1.
We now describe the isolation and characterization of porcine daintain/AIF1 and show that it has a dual influence on glucose-induced insulin secretion in vivo: inhibition at low concentration and stimulation at high concentrations. We have raised antibodies against the peptide and demonstrate that daintain/AIF1-like immunoreactivity is widely present in cells of the macrophage lineage. Because of its dual influence on glucose-induced insulin release in vivo, and its presence in macrophages, we analyzed the presence of daintain/AIF1 immunoreactivity in the pancreas of prediabetic and newly diabetic rats of the diabetes-prone strain BB. These animals frequently are used as a model of human insulin-dependent diabetes mellitus (IDDM) (11). IDDM is an organ-specific autoimmune disease with destruction of β cells as a consequence of tissue infiltration by macrophages, lymphocytes, and other mononuclear cells (12). The daintain/AIF1 immunoreactivity found makes the peptide of interest in the pathogenesis of IDDM. In addition, we notice that daintain/AIF1 contains a –KR–KK–GKR– motif typical of precursors to hormonally active peptides (13–16) and therefore also may have hormone precursor functions.
MATERIALS AND METHODS
Materials.
Porcine intestines were processed to yield a concentrate of thermostable intestinal polypeptides (5, 6), which was further purified as described (17) until an exclusion chromatography step on Sephadex G-25. The material corresponding to fraction 2 (11.5 liters) from that chromatogram (17) was collected and saturated with NaCl (320 g/liter) to produce a crude peptide precipitate (55 g) as the initial material for this study.
Purification and Structural Analysis of Daintain/AIF1.
A portion (15 g) of the initial material was dissolved in water (150 ml). After addition of ethanol (300 ml), adjustment to pH 7.2 with basic ethanol (1 vol 1 M NaOH/2 vol ethanol), and stirring for 5 min, the suspension was held at −20°C for 48 hr. A precipitate was collected by suction and was washed with ethanol/ether at −20°C. After evaporation under vacuum, 7.08 g of dry material was recovered. An aliquot (350 mg) was suspended in 10 ml 0.01 M NH4HCO3, pH 8.0. After centrifugation, this supernatant was applied to a DEAE-cellulose column (25 × 3 cm) in the same buffer. Elution was performed by stepwise increments to 0.02, 0.04, 0.1, 0.2 M NH4HCO3, and finally 0.2 M NH4OH. The fraction eluted with 0.1 M NH4HCO3 displayed a suppression of insulin release in our bioassay and was purified further. After lyophilization, this fraction (130 mg) was applied in aliquots to reverse-phase HPLC on Pharmacia-LKB TSK ODS 120-T (10 μm; 300 × 7.8 mm) by using a linear gradient of 24–56% acetonitrile in 0.1% trifluoroacetic acid (54 min, 1.5 ml/min) for elution. The fractions from this step, which suppressed insulin release, were purified further by rechromatography on Vydac 218TP54 (5 μm; 250 × 4.6 mm) with a linear gradient of 47–55% acetonitrile in 0.1% heptafluorobutyric acid (32 min, 1 ml/min) for elution.
The purity of the isolated polypeptide was ascertained by capillary zone electrophoresis in 50 mM phosphate buffer, pH 2.5, with a Beckman P/ACE 2100 system, by using fused silica capillaries (inner diameter 75 μm, length 57 cm). Molecular masses were determined with matrix-assisted laser desorption/ionization mass spectrometry in a Finnigan MAT Lasermat 2000 instrument, by using α-cyano-4-hydroxycinnamic acid (Sigma) as matrix added in aqueous 70% acetonitrile, 0.1% trifluoroacetic acid (10 mg/ml). Bovine cytochrome c (Sigma) was used as internal standard. Total compositions were determined with a Pharmacia-LKB Alpha Plus 4151 amino acid analyzer, after hydrolysis for 22 hr at 110°C in evacuated tubes with 6 M HCl/0.5% phenol (wt/vol). For N-terminal deblocking, a sample (1.5 nmol) was treated as described (18) and subjected to sequence analysis. Fragmentations were produced with trypsin and endoproteinases Lys-C and Glu-C (the products from Boehringer Mannheim) at enzyme to substrate ratios of 1:15, by weight, and with CNBr in 70% formic acid for 24 hr at room temperature, or with just 70% formic acid for 48 hr at 35°C. All fragments obtained were separated by reverse-phase HPLC on Vydac 218TP54 (5 μm; 250 × 4.6 mm) in 0.1% trifluoroacetic acid with a 0–60% gradient (90 min, 1 ml/min) of acetonitrile. Amino acid sequences of overlapping fragments were determined with an Applied Biosystems 477A instrument coupled to a 120 A analyzer, or with a 470 A instrument by using separate reverse-phase HPLC for detection.
Bioactivity Measurements.
Screening for peptides influencing insulin secretion was performed as described (17) by using isolated rat pancreatic islets (19).
For bioactivity analysis in vivo, nonfasted NMRI mice were anesthetized with an intraperitoneal injection of midazolam (Dormicum, Hoffman–La Roche, 0.4 mg/mouse) and a combination of fluanison (0.9 mg/mouse) and fentanyl (0.02 mg/mouse, Hypnorm, Janssen). After this injection, a blood sample was taken into heparinized tubes, from the retro-orbital venous plexus as described (20). d-Glucose (BDH) was injected intravenously (1 g/kg), alone or combined with daintain/AIF1 at dose levels 75 pmol/kg, 750 pmol/kg, 7.5 nmol/kg, and 75 nmol/kg. Blood samples were taken after 1, 5, 20, and 50 min. The samples were immediately centrifuged, and the plasma was separated and stored at −20°C until analysis.
Insulin in plasma was determined radioimmunochemically with the use of a guinea pig anti-rat insulin antibody, 125I-labeled porcine insulin as tracer, and a mix of rat insulin I and II as standard (Linco Research Immunoassay, St. Charles, MO). Free and bound radioactivity was separated by use of an anti-IgG (goat anti-guinea pig) antibody (Linco). The sensitivity of the assay was 12 pmol/liter, and the coefficiency of variation was less than 7% at both low and high levels. Glucose was determined with the glucose oxidase method (21).
In statistics, means ± SEM are shown. Statistical analyses were performed with the program spss for Windows. Comparisons between groups were performed with the Student’s paired t test. The area under the curve for insulin levels was calculated for suprabasal insulin levels during the first 20 min after glucose injection by the trapezoid rule. The glucose elimination rate (k value) after glucose injection was calculated by using t1/2 after logarithmic transformation of the individual plasma glucose values, as described (22).
Peptide Localization.
Biobreeding diabetes-prone and diabetes-resistant BB-Wistar rats (200 ± 20 g body weight, Karolinska Hospital, Stockholm, Sweden) were perfusion-fixed under hexobarbital anesthesia (100 mg/kg) with 200 ml of ice-cold fixative, consisting of 4% paraformaldehyde and 0.2% picric acid in 0.1 M phosphate buffer, pH 6.9. All organs were dissected and postfixed for 2 hr. After rinsing overnight in buffered 10% sucrose, the organs were cut in 14-μm sections with a cryostat. Thaw-mounted sections were incubated overnight with a polyclonal rabbit antiserum against daintain/AIF1, diluted 1:1,000 in 0.3% Triton (Sigma) and 0.5% BSA (Sigma). This antiserum was produced by the National Veterinary Institute, Sweden, and it was confirmed with a standard immune blot test. As specificity control of the daintain/AIF1-like immunoreactivity, closely adjacent sections were incubated with the anti-daintain/AIF1 antibody preabsorbed with the daintain/AIF1 polypeptide (10 mg/ml diluted antiserum). In double immunolabeling experiments, sections were incubated with both the antibody against daintain/AIF1 and each of the following mouse mAbs (Serotec): OX-42 (diluted 1/200), which labels CD11b/c antigen present in microglia; OX-18 (diluted 1/500) against rat major histocompatibility complex class I, OX-6 against rat MHC class II (diluted 1/200), OX-8 against a cytotoxic T cell marker (diluted 1/200) and ED-1, which labels an antigen expressed by the majority of tissue macrophages (diluted 1/200). Immunoreactivities were detected by secondary antibodies, fluorescein isothiocyanate- conjugated goat anti-rabbit and CY-3 (indocarbocyanine)-conjugated goat anti-mouse Igs (Jackson ImmunoResearch), diluted 1:100 and 1:400, respectively.
RESULTS
Purification.
The early steps of the daintain purification were carried out as described (5, 6, 17). After DEAE-cellulose ion-exchange chromatography, the active fractions suppressing insulin secretion in vitro were combined and purified further. Aliquots (2.5 mg) were subjected to reverse-phase HPLC on TSK ODS 120-T, and the components absorbing at 214 nm were collected and lyophilized (Fig. 1A). The bioactive fraction, indicated by a bar, was submitted to a second HPLC on Vydac 218TP54 (Fig. 1B). The final preparation was homogeneous as judged by capillary electrophoresis (Fig. 1C) and the mass was determined as 16,607 ± 3 Da by matrix-assisted laser desorption/ionization mass spectrometry. From 1 kg of boiled, wet intestinal tissue, 5 μg of pure polypeptide was obtained.
Figure 1.
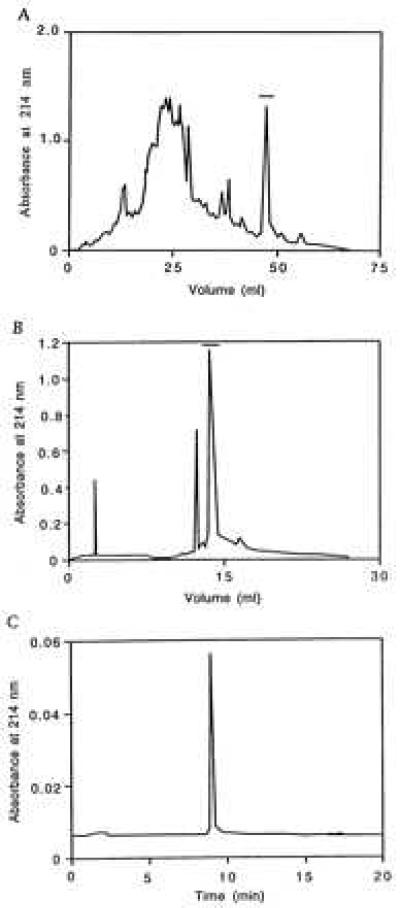
Purification of porcine daintain/AIF1. (A) HPLC on Pharmacia-LKB TSK ODS 120-T, the active fraction is indicated by a bar. (B) Additional HPLC on Vydac 218TP54, the main peak indicated by a bar corresponds to daintain/AIF1. (C) Two micrograms of the purified sample applied to capillary zone electrophoresis at 25 kV, 25°C in 50 mM phosphate, pH 2.5, showing a symmetrical peak.
Structure.
Direct sequencer analysis indicated that daintain/AIF1 is N-terminally blocked. It was determined to be a 146-residue polypeptide (Fig. 2) by fragment sequence analysis and by direct analysis after alcoholytic deacetylation. This sequence (including the N-terminal acetyl group) corresponds to a total mass of 16,602.5 Da, which is close to the mass value above actually established by mass spectrometry. An internal 44-residue segment with the sequence pattern –KR–KK–GKR– has a motif typical of peptide hormone precursors, i.e., double basic amino acid sites for potential activation cleavages during prohormone processing. At the sequentially last such site, the tripeptide GKR has the additional property of a signal structure for formation of a C-terminal amide.
Figure 2.
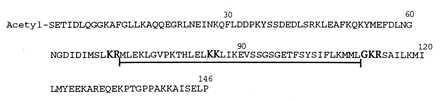
Structure of porcine daintain/AIF1. The sequence motif typical of posttranslational processing of peptide hormones is underlined. The cleavage sites for prohormone convertases and peptide amidation enzymes are boldfaced.
Bioactivity.
The purification of daintain/AIF1 was guided by the determination of the inhibitory effect of chromatographic fractions on insulin release from isolated pancreatic islets in the presence of 16.7 mM glucose. This activity in vitro of the highly purified peptide was, however, inconsistent and less marked than mentioned in a preliminary conference report (see above).
To further establish the effect of daintain/AIF1 on the pancreatic β cell, we studied acute plasma insulin and glucose responses in vivo after peptide administration intravenously in mice. The intravenous injection of glucose (1 g/kg) into nonfasted NMRI mice rapidly raised plasma insulin levels, with a peak obtained already after 1 min, after which plasma insulin levels returned toward baseline values. Concomitantly, plasma glucose levels also returned to baseline values from the peak value with first-order kinetics. At the lowest dose level of 75 pmol/kg, daintain/AIF1 inhibited glucose-induced insulin secretion. This result was evident in two ways. One was by the lower plasma insulin levels (at 1 or 5 min after glucose injection) between the glucose alone and the glucose plus 75 pmol/kg daintain/AIF1 groups (P < 0.05). The other was by calculation of the area under the curve for insulin (Table 1). Concomitantly, the glucose elimination was inhibited by daintain/AIF1 at this low dose level, as evidenced from the lowered KG (Table 1).
Table 1.
Effects of daintain/AIF1 on the insulin and glucose elimination rates
| AUC, nmol/liter in 20 min | KG, % per min | n | |
|---|---|---|---|
| Experiment series 1 | |||
| Glucose alone | 10.2 ± 1.7 | 2.4 ± 0.1 | 17 |
| Glucose + peptide, | 17.2 ± 2.4* | 2.7 ± 0.1* | 15 |
| 75 nmol/kg | |||
| Experiment series 2 | |||
| Glucose alone | 9.2 ± 0.6 | 2.2 ± 0.1 | 17 |
| Glucose + peptide, | 15.7 ± 3.0* | 2.6 ± 0.1* | 17 |
| 7.5 nmol/kg | |||
| Experiment series 3 | |||
| Glucose alone | 11.4 ± 1.7 | 2.4 ± 0.2 | 12 |
| Glucose + peptide, | 13.2 ± 2.1 | 2.3 ± 0.1 | 12 |
| 750 pmol/kg | |||
| Experiment series 4 | |||
| Glucose alone | 10.1 ± 1.2 | 2.7 ± 0.1 | 12 |
| Glucose + peptide, | 6.2 ± 1.1* | 2.1 ± 0.1† | 12 |
| 75 pmol/kg |
The area under the curve for insulin (AUC) and the glucose elimination rate (KG) during 20 min after the intravenous injection of glucose (1 g/kg) with or without addition of daintain/AIF1 at four different dose levels in anesthetized NMRI mice is shown. Means ± SEM are given. n indicates number of animals, ∗ indicates P < 0.05, and
indicates P < 0.01 between the groups.
In contrast, at the two high-dose levels of 7.5 and 75 nmol/kg, daintain/AIF1 potentiated the glucose-induced increase in plasma insulin concentration (Fig. 3 and Table 1) with a concomitant enhancement of the glucose elimination (Table 1). At the medium-dose level of 750 pmol/kg, daintain/AIF1 did not significantly affect the glucose-induced increase in plasma insulin concentration or in glucose elimination (Fig. 3 and Table 1).
Figure 3.
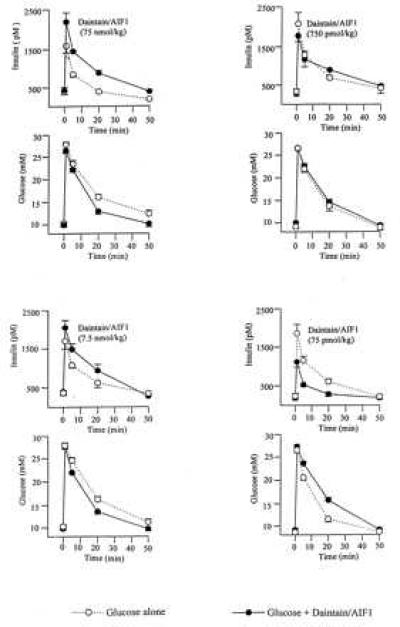
Effects of porcine daintain/AIF1 on glucose-induced insulin release. Plasma levels of insulin and glucose before and after an intravenous injection of glucose (1 g/kg) with or without the peptide at four different dose levels in anesthetized NMRI mice (12–17 animals in each group).
Distribution.
By means of immunohistochemical techniques that used a rabbit antiserum against purified daintain/AIF1, we have been able to demonstrate in rat tissues a large population of daintain/AIF1-like immunoreactive cells resembling macrophages and dendritic cells. Daintain/AIF1-like immunoreactive cells were located in the lamina propria of the intestinal mucosa (dendritic cells, Fig. 4 a and b); the liver (Kupffer cells, not shown); the central nervous system (Fig. 4c); the lymphatic organs, such as lymph nodes and the cortex of the thymus (dendritic cells, Fig. 4d); the red pulp of the spleen (Fig. 4e); and the pancreas (macrophages, Fig. 4g). Double immunolabeling analysis with the marker OX-42 revealed the microglial nature of the daintain/AIF1-like immunoreactive cells found over the entire central nervous system.
Figure 4.
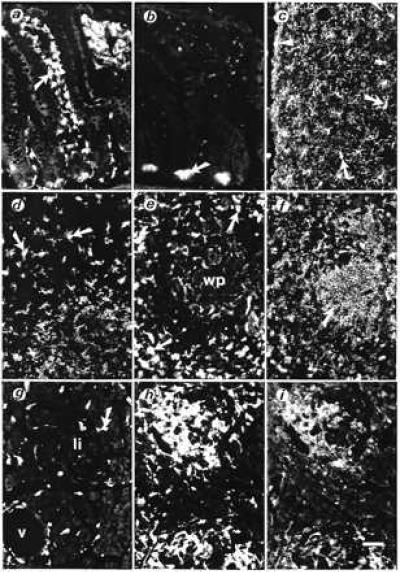
Cellular localization of porcine daintain/AIF1-like immunoreactivity in tissues from normal BB-Wistar rats (a–e and g) and from diabetes-prone BB rats (without hyperglycemia, f, h, and i). (a) In sections of the ileum, strong daintain/AIF1-like immunoreactivity is found in a large number of dendritic cells beneath the epithelial layer (arrow), within the lamina propria of the tunica mucosa, and in scattered cells in the tunica submucosa. (b) Preabsorption of the daintain/AIF1 antibody with the purified peptide (10 μg/ml diluted antibody) greatly prevented the appearance of daintain/AIF1 immunoreactivity. Only autofluorescent cell profiles, such as Paneth cells (arrow), are present after absorption. (c) Daintain/AIF1-like immunoreactivity in ramified microglial cells (arrows) located over the entire cerebral cortex and concentrated beneath the pia mater (arrowhead). These cells also were labeled after incubation with an antibody OX-42 against CD11b/c antigen present in microglia. (d) Daintain/AIF1-like immunoreactivity in dendritic cells within the cortex of the thymus (arrows) and (e) in the spleen, present in scattered cells within the white pulp (wp) and abundantly in macrophages (arrows) of the red pulp. (g) Daintain/AIF1 immunoreactive cells scattered in the exocrine pancreas (arrow), mainly in the vicinity of vessels (v), and occasionally within pancreatic islets (Langerhan’s islets, li). (h) In prediabetic BB rats, strongly daintain/AIF1-like immunoreactive cells are found around and within pancreatic islets. (i) Daintain/AIF1 immunoreactivity colocalizes with MHC class II immunoreactivity in the same section of the pancreas of a prediabetic BB rat. In the spleen of the prediabetic BB rat, MHC class II immunoreactivity is present in lymphocytes of the white pulp (arrow, f), whereas daintain/AIF1 immunoreactivity, is more concentrated to the red pulp (compare with e). The magnification bar represents 50 μm, as shown in i.
Because daintain/AIF1 was found to be a component of macrophages in the pancreas and to have an effect on insulin release, we decided to study daintain/AIF1 immunoreactivity in the pancreas of Biobreeding BB rats, which constitute an animal model for autoimmune insulin-dependent diabetes mellitus. Diabetes-prone BB rats develop acute hyperglycemia between 50 and 100 days of age, and the clinical onset is preceded by infiltration of mononuclear cells to form an insulitis, first by macrophages, then by both B and T lymphocytes (23, 24). We found daintain/AIF1 immunoreactivity in macrophages over the entire pancreatic parenchyma in the prediabetic BB rats. Especially, cells infiltrating the islets show strong immunoreactivity (compare Fig. 4 h and g). The amount of β cells within the islets was reduced, but to a variable degree. In contrast, β cells were almost absent in the pancreas of already hyperglycemic BB rats, whereas the α cells, producing glugacon, are present. Double labeling of pancreas sections, both with the antiserum against daintain/AIF1 and with mAbs against the same immunological markers, revealed that daintain/AIF1 was colocalized with OX-18 (rat MHC class I), OX-6 (rat MHC class II), (Fig. 4 h and i), and ED-1, a marker for macrophages. It also was found that MHC class II was present mainly in lymphocytes of the white pulp of the spleen (Fig. 4f), but that those lymphocytes do not show daintain/AIF1-like immunoreactivity.
DISCUSSION
AIF1 originally was cloned and identified in rat cardiac allografts with chronic rejection states (8). A similar study was carried out in transplanted human hearts (9). It also was found that AIF1 mRNA is present in injured rat carotid arteries (10). In our study, we have purified daintain in large quantity from porcine intestines and found that it is identical to AIF1. By determination of the porcine daintain/AIF1 primary structure, we find that it is N-terminally acetyl-blocked and that the amino acid sequence is 87% and 90% identical to that of rat and human AIF1, respectively. Interestingly, an internal 44 residue segment, with the conserved sequence motif –KR–KK–GKR– (Fig. 2), suggests that daintain/AIF1 might be a precursor protein. That is because most characterized peptide hormones are flanked, in their precursors, by pairs of basic amino acid residues, constituting cleavage sites for the processing enzymes (13, 14). The sequence GKR is a signal for the formation of a C-terminal amide during the processing (15, 16).
In this study, we have further found that daintain/AIF1 exerts a dual influence on glucose-stimulated insulin secretion in normal mice in vivo. The dual pattern is dependent on the peptide concentration and suggests that several effects are induced by the peptide in vivo. Hypothetically, indirect effects could be mediated by neural actions of daintain/AIF1, because sympathetic nerve activation inhibits insulin secretion, whereas parasympathetic nerve activation stimulates it (25). Alternatively, daintain/AIF1 as a macrophage polypeptide may influence insulin secretion through cytokines, the effects of which have been shown to be both stimulatory and inhibitory under different conditions (26). We have at present no explanation why the purified daintain/AIF1 only weakly and inconsistently inhibits glucose-induced insulin secretion from isolated rat pancreatic islets in vitro, despite the fact that this effect was instrumental in the purification and discovery of daintain/AIF1.
Monocytes, macrophages, microglia, and dendritic cells form an extended family of cells derived from bone marrow precursors (27). These cells are resident or appear on activation by pathological processes and express a large set of immunological markers (28). Microglia, dendritic cells, and Langerhan’s cells of the skin have antigen-presenting activities. Macrophages, besides their phagocytic function, also have antigen-presenting activity and exhibit autocrine and paracrine properties related to the release of both inflammatory and anti-inflammatory factors (29).
Cytokines, released from infiltrating lymphocytes and macrophages, have been implicated in autoimmune diabetes by exerting direct effects on β cell dysfunction and by recruitment of lymphocytes and macrophages to the site of inflammation (30, 31). Our results confirm that daintain/AIF1 cells, densely located in the prediabetic pancreas, infiltrate the islets, which suggest that daintain/AIF1 has properties of a cytokine-associated factor. Furthermore, it has been found that daintain/AIF1 mRNA expression is strongly detectable in the intestine, that is in the tissue of our preparation, and more strongly in the spleen, but very weakly in the normal pancreas (10). Especially, we prepared a crude peptide sample from pancreas glands of a prediabetic BB rat, checked it with the standard Western blot analysis, and found that indeed a high amount of daintain was present in the pancreas of the prediabetic BB rat, which is consistent with the results from our immunohistochemical experiments.
In conclusion, we report on the properties of a novel pig polypeptide, daintain/AIF1, purified from intestine, chemically characterized as an N-terminally acetylated, 146-residue polypeptide, with a hormone precursor motif. Its cellular localization in the macrophage lineage, its dual effects on β cell function in vivo, and its abundant presence in prediabetic pancreas, suggest that daintain/AIF1 is not only associated with allograft inflammation, but also could play a role in cell-mediated autoimmune responses, e.g., in connection with the mechanisms by which the diabetogenic β cell lesion is induced.
Acknowledgments
We are grateful to Dr. Rannar Sillard, Lilian Bengtsson, Irene Byman, Ulrika Gustavsson, Lena Kvist, Gunilla Lundquist, and Åke Norberg for discussion and assistance. This research was supported by the Swedish Society for Medical Research and the Swedish Medical Research Council (Projects 04x-715, 13x-00034, 13x-1010, 13x-3532, 13x-10832, and 14x-6834), the Swedish Cancer Foundation, Novo Nordic Foundation, Albert Påhlsson Foundation, Crafoord Foundation, Ernhold Lundström Foundation, the Swedish Diabetes Association, and the Faculty of Medicine, Lund University.
ABBREVIATIONS
- AIF1
allograft inflammatory factor 1
- MHC
major histocompatibility complex
Footnotes
Data deposition: The sequence reported in this paper has been deposited in the GenBank database (accession no. P81076).
References
- 1.Dupre J, Ross S A, Watson D, Brown J C. J Clin Endocrinol. 1973;37:826–828. doi: 10.1210/jcem-37-5-826. [DOI] [PubMed] [Google Scholar]
- 2.Kreymann B, Williams G, Ghatei M A, Bloom S R. Lancet. 1987;2:1300–1304. doi: 10.1016/s0140-6736(87)91194-9. [DOI] [PubMed] [Google Scholar]
- 3.Arimura A, Sato H, Dupont A, Nishi N, Schally A V. Science. 1975;189:1007–1009. doi: 10.1126/science.56779. [DOI] [PubMed] [Google Scholar]
- 4.Tatemoto K, Rökaeus Å, Jörnvall H, McDonald T J, Mutt V. FEBS Lett. 1983;164:124–128. doi: 10.1016/0014-5793(83)80033-7. [DOI] [PubMed] [Google Scholar]
- 5.Chen Z-W, Agerberth B, Gell K, Andersson M, Mutt V, Östenson C-G, Efendic S, Darros-Söderling J, Persson B, Jörnvall H. Eur J Biochem. 1988;174:239–245. doi: 10.1111/j.1432-1033.1988.tb14088.x. [DOI] [PubMed] [Google Scholar]
- 6.Agerberth B, Söderling-Barros J, Jörnvall H, Chen Z-W, Östenson C-G, Efendic S, Mutt V. Proc Natl Acad Sci USA. 1989;86:8590–8594. doi: 10.1073/pnas.86.21.8590. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Chen, Z.-W., Östenson, C.-G., Bergman, T. & Efendic, S. (1994) Eur. J. Endocrinol. 130, Suppl. 2, 32 (abstr.).
- 8.Utans U, Arceci R J, Yamashita Y, Russell M E. J Clin Invest. 1995;95:2954–2962. doi: 10.1172/JCI118003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Utans U, Quist W C, McManus B M, Wilson J E, Arceci R J, Wallace A F, Russell M E. Transplantation. 1996;62:1387–1392. doi: 10.1097/00007890-199605150-00018. [DOI] [PubMed] [Google Scholar]
- 10.Autieri M-V. Biochem Biophys Res Comm. 1996;228:29–37. doi: 10.1006/bbrc.1996.1612. [DOI] [PubMed] [Google Scholar]
- 11.Nakhooda A F, Like A A, Chappel C I, Wei C N, Merliss E B. Diabetologia. 1978;14:199–207. doi: 10.1007/BF00429781. [DOI] [PubMed] [Google Scholar]
- 12.Rossini A A, Greiner D L, Friedman H P, Mordes J P. Diabetes Rev. 1993;1:43–75. [Google Scholar]
- 13.Steiner D F, Smeekens S P, Ohagi S, Chan S J. J Biol Chem. 1992;267:23435–23438. [PubMed] [Google Scholar]
- 14.Seidah N G, Day R, Chretien M. Biochem Soc Transact. 1993;21:685–691. doi: 10.1042/bst0210685. [DOI] [PubMed] [Google Scholar]
- 15.Bradbury A F, Smyth D G. Eur J Biochem. 1987;169:579–584. doi: 10.1111/j.1432-1033.1987.tb13648.x. [DOI] [PubMed] [Google Scholar]
- 16.Katopodis A G, Ping D-S, Smith C E, May S W. Biochemistry. 1991;30:6189–6194. doi: 10.1021/bi00239a016. [DOI] [PubMed] [Google Scholar]
- 17.Chen Z-W, Bergman T, Östenson C-G, Höög A, Näslund J, Norberg Å, Carlquist M, Efendic S, Mutt V, Jörnvall H. FEBS Lett. 1994;341:239–243. doi: 10.1016/0014-5793(94)80464-8. [DOI] [PubMed] [Google Scholar]
- 18.Gheorghe M T, Jörnvall H, Bergman T. Anal Biochem. 1997;254:119–125. doi: 10.1006/abio.1997.2380. [DOI] [PubMed] [Google Scholar]
- 19.Östenson C-G, Grill V. Mol Cell Endocrinol. 1986;45:215–221. doi: 10.1016/0303-7207(86)90150-4. [DOI] [PubMed] [Google Scholar]
- 20.Rerup C, Lundquist I. Acta Endocrinol. 1966;52:357–367. [PubMed] [Google Scholar]
- 21.Bruss M L, Black A L. Anal Biochem. 1978;84:309–312. doi: 10.1016/0003-2697(78)90514-6. [DOI] [PubMed] [Google Scholar]
- 22.Ikkos D, Luft R. Acta Endocrinol. 1957;25:312–334. doi: 10.1530/acta.0.0250312. [DOI] [PubMed] [Google Scholar]
- 23.Lee K U, Kim M Y, Amano K, Pak C Y, Jaworski M A, Mehta J G, Yoon J-W. Diabetes. 1988;37:1053–1058. doi: 10.2337/diab.37.8.1053. [DOI] [PubMed] [Google Scholar]
- 24.Walker R, Bone A J, Cooke A, Baird J D. Diabetes. 1988;37:1301–1304. doi: 10.2337/diab.37.9.1301. [DOI] [PubMed] [Google Scholar]
- 25.Ahren B, Taborsky G J, Jr, Porte D., Jr Diabetologia. 1986;29:827–836. doi: 10.1007/BF00870137. [DOI] [PubMed] [Google Scholar]
- 26.Mehta V K, Hao W, Brooks-Worrell B M, Palmer J P. Eur J Endocrinol. 1994;130:208–214. doi: 10.1530/eje.0.1300208. [DOI] [PubMed] [Google Scholar]
- 27.Perry V H, Hume A, Gordon S. Neuroscience. 1985;15:313–326. doi: 10.1016/0306-4522(85)90215-5. [DOI] [PubMed] [Google Scholar]
- 28.Flaris N A, Densmore T L, Molleston M C, Hickey W F. Glia. 1993;7:34–40. doi: 10.1002/glia.440070108. [DOI] [PubMed] [Google Scholar]
- 29.Gordon S. BioEssays. 1995;17:977–986. doi: 10.1002/bies.950171111. [DOI] [PubMed] [Google Scholar]
- 30.Voorbij H A M, Jeucken P H M, Kabel P J, de Haan M, Drexhage H A. Diabetes. 1989;38:1623–1629. doi: 10.2337/diab.38.12.1623. [DOI] [PubMed] [Google Scholar]
- 31.Bach J-F. Endocrinol Rev. 1994;15:516–542. doi: 10.1210/edrv-15-4-516. [DOI] [PubMed] [Google Scholar]