

Abstract
This study investigated the use of motor abundance during the transport and placing of objects that required either precise or minimal orientation to the target. Analyses across repetitions of the structure of joint configuration variance relative to the position and orientation constraints were performed using the Uncontrolled Manifold (UCM) approach. Results indicated that the orientation constraint did not affect stability of the hand's spatial trajectory. Orientation was weakly stabilized during the late transport phase independent of the orientation constraint, indicating no default synergy stabilizing orientation. Stabilization of orientation for conditions most requiring it for successful insertion of the object was present primarily during the adjustment phase. The results support the hypothesis that a major advantage of a control scheme that utilizes motor abundance is the ability to resolve multiple task constraints simultaneously without undue interference among them.
Introduction
Motor redundancy or, when considered more positively, motor abundance (Gelfand and Latash 1998) makes it possible for multiple variations of joint and muscle coordination to be used to achieve a given task performance. That motor abundance is actually used by the central nervous system (CNS) when coordinating the motor elements (i.e., muscles, joints, finger forces, etc.) has been established in numerous studies of a variety of functional tasks (Danna-Dos-Santos et al. 2007; Hsu et al. 2007; Freitas et al. 2007; Scholz et al. 2007; Krishnamoorthy et al. 2007; Olafsdottir et al. 2007; Tseng et al. 2006; Tseng and Scholz 2005a, 2005b; Krisnamoorthy et al. 2005; Shinohara et al. 2004; Krishnamoorthy et al. 2003; Scholz et al. 2003; Latash et al. 2002; Tseng et al. 2002; Scholz et al. 2002; Latash et al. 2001; Scholz et al. 2001; Scholz et al. 2000; Scholz and Schöner 1999). These results are consistent with Bernstein's intuition that task performance typically involves “repetition without repetition” (Bernstein 1967). Although the role of motor abundance in decreasing performance variability (“error compensation”) has been emphasized in past work (e.g. Latash et al. 2007), we hypothesize that a major advantage is that it facilitates the performance of multiple tasks, or the solution of multiple task constraints, simultaneously with minimal interference among tasks or among components of a given task (Scholz and Kubo 2008). Many of the above-cited studies have investigated differences in indices of variability of various task-relevant variables when performing a particular task. However, none of those studies directly examined the effect of manipulating one task constraint on the stability of another, leaving this hypothesized advantage of motor abundance largely untested. In this paper, by “stabilization” of a performance variable, we imply relatively low variability across trials compared to what one could expect if all elemental variables affecting that performance variable varied independently of each other.
One exception was a recent study of finger force production (Zhang et al. 2008). In that study, the control of total force (primary task) and the moment of force about an axis of rotation (secondary task) were studied in a set of fingers. The authors found that stabilization of the moment of force was accomplished without adversely affecting total force stabilization by using different strategies to coordinate the finger forces compared to an isolated force-production task. The results from Zhang et al. (2008) suggested that participants exploited variable solutions for coordinating the effectors in order to maintain performance accuracy when the same effectors were involved in resolving another task constraint simultaneously. However, beyond this example, it is unclear how typical it is for the nervous system to use motor abundance to resolve multiple task constraints simultaneously. The present experiment investigated whether this hypothesis could be supported in kinematic tasks that required either reaching to a target location without an explicit constraint on hand orientation or reaching to the same target location when precise orientation of the hand to the target was required. A second goal of this work was to investigate whether the control of hand/object orientation was limited to the final phase of reaching, when the hand approached the target, or was initiated earlier in the reach. This question has been a point of some controversy (Desmurget et al. 1996; Soechting and Flanders 1993; Wang 1999).
Numerous studies have examined the control of hand/object orientation during the performance of skilled upper extremity tasks (Cuipjers, Smeets & Brenner, 2004). Those studies have not considered explicitly the role of motor abundance in resolving various constraints on task performance. Given the availability of motor abundance, simultaneous solution of these constraints is theoretically possible without mutual interference. Results of studies suggesting independent control of position and orientation could be consistent with this capacity (Fan et al. 2006; Soechting and Flanders 1993; Wang 1999). The method of the Uncontrolled Manifold (UCM) hypothesis (Schöner 1995; Scholz and Schöner 1999) provides a powerful tool to address such questions compared to the regression or correlation methods typically used. This method is based on the quantitative analysis of variance within two sub-spaces that form the space of elemental variables (joint rotations in studies of movement kinematics). The first sub-space (the UCM) corresponds to a certain desired value of a performance variable. This performance variable's magnitude changes within the other, complementary sub-space. By identifying how variance of the configuration of all joint motions affects the stability of specific task-relevant variables, such as the hand's spatial position and its orientation to the target, the method can be used to investigate whether different task constraints interfere with one another. This is the approach adopted in the current investigation.
We predicted that motor abundant solutions to joint coordination would be used to stabilize the hand's movement path regardless of the orientation constraint, and that the strength of the joint synergy affecting this stabilization would not differ among the conditions. In addition, it was predicted that the synergy related to stability of the hand's orientation to the target would appear only for those conditions where an explicit orientation constraint was present. We also sought to determine whether the synergy stabilizing hand-target orientation in those conditions requiring it was present throughout the transport phase or only when approaching the target.
Methods
Participants
Ten healthy adult volunteers participated in the study (age 26.5± 4.4 years). All participants signed the informed consent form approved by the Human Subject Review Board at the University of Delaware. All subjects were right-hand dominant, confirmed by 10-point Edinburgh Handedness Inventory (Oldfield, 1972) and were naive to the purpose of the study.
Participant set-up
Participants sat on a height-adjustable, high-back chair at a round table that had a rectangle cut out of one side into which the chair was inserted to provide for lateral support of the arm in the initial position. The participant's abdomen pressed firmly against the front and right side of the table's indentation to ensure stability across the experiment. A wide strap attached to the chair restricted trunk movement. Chair height was adjusted so that the participant's upper arm and forearm were at a 90° angle, with the forearm resting parallel to the table surface in a neutral position. The wrist was positioned in neutral flexion-extension. The initial arm and hand position was maintained by fitting a vacuum air bag to the underside, laterally and medially to the elbow, forearm and hand. The air was then vacuumed out to form a trough. The arrangement restricted only pure lateral or medial arm movement from the initial position.
Reflective markers were placed on the participants' right arm and used to track their movement. Rigid bodies, each comprised of an array of four reflective markers, were mounted on rigid shells and fitted to the participants at the following locations: 2/3 of the distance between the neck and the acromion process of the right shoulder; on the lateral side of the upper arm, 2/3 of the distance from the elbow to the wrist on the forearm, and on the back of the hand. Individual markers were used to locate approximate joint centers during an initial participant calibration trial. They were placed 1) immediately inferior to the sternum notch, 2) 2-cm below the acromion process on the lateral shoulder, 3) on the lateral and medial epicondyles of the humerus (the mean of where were used to compute the approximate joint center), and 4) on the radial and ulnar styloid processes of the forearm (the mean of which was used to compute the approximate joint center). The sternum marker served as the origin of the body-centered coordinate system used in the computations. In addition, individual markers were placed and remained on the subject's glove, just proximal to thumb interphalangeal joint and on the proximal interphalangeal joint of the index finger.
Experiment Conditions and Setup
Participants wore a batting glove in their right hand, the palm of which was made of the loop-side of sticky backed Velcro. For conditions 1-3 that required the insertion of an object into a slotted target (see below), a 7.6-cm diameter wooden ball that was covered with the hook-side of Velcro and attached firmly to the loop-side Velcro of the glove's palm as the subject grasped it. This ensured that a constant relationship between the ball's initial orientation and its position with respect to the hand could be maintained throughout the experiment (Figure 1). Once this ball was placed in the hand, it was not removed during any of these three conditions.
Figure 1.

Experimental Setup. Subjects wore a tight-fitting glove with loop sided Velcro attached to the palm while the ball had hook sided Velcro attached to it. The object to be inserted into the target screwed tightly into the large ball, controlling its initial orientation. The trapezoid shape (TRAP) is shown. For the ONLY condition, only an identically sized large ball was held, but without the Velcro to allow its release into the same container with a large circular cutout.
The side of the large ball opposite to the hand had a bolt inserted into it that faced away from the participant. One of the three different shaped objects was screwed tightly onto this bolt for Conditions 1-3. For Condition 4, which had no explicit orientation constraint, participants held in their gloved-hand an identical 7.6-cm diameter wooden ball without a Velcro cover, which allowed them to release the ball at the target.
The target consisted of a coffee can to which different plastic lids were attached depending on the experimental condition (Figure 1). The target was placed vertically at the body's midline on the table at which participants sat, at a distance from the body that was 85% of each participant's functional arm length, determined as the distance from the sternum to the midpoint of the web space between the thumb and index finger when the arm was fully extended.
For Condition 4, which we refer to here as the ONLY condition because the participant held only the large ball without Velcro in the hand, the coffee can lid had a large circular cutout slightly larger than the ball. The task was for the participant to reach with the large ball to the can and drop the ball into the can in one continuous motion. For Conditions 1-3, three different plastic coffee-can lids were used. Each lid had a different shape cut out of its center of the exact shape and slightly larger than the object that was attached to the large Velcro-covered ball held in the participants' hand.
The under surface of all lids were reinforced with a thick cardboard cutout to stiffen the edge of each shape. This required relatively precise orientation of the object to fit it into the slot. However, it was difficult to eliminate plasticity of the cutout completely, particularly for rotation about the anterior-posterior and medial-lateral axes.
The task for Conditions 1-3 was to reach to the target and insert the object into the slot in one continuous motion. The three objects came from a typical wooden infant toy. For the BALL condition (not to be confused with the large ball which we call the ONLY condition), a 5-cm diameter ball was attached to the large Velcro-covered ball and inserted into a 5.2-cm diameter circular cutout in the target lid. To insert the small ball into the cutout in the lid required that the hand be oriented so that the 5-cm diameter ball was below the hand; otherwise the large ball to which it was attached would hit the can and prevent insertion. This condition provided, therefore, some constraint on hand-object orientation along the roll and pitch axes. However, there was no explicit constraint on rotation about the vertical, or yaw axis of the target. For the other two conditions, the objects attached to the large ball were a trapezoid (TRAP condition) and a five-pointed star (STAR condition). These two conditions required more precise control of orientation about all axes, particularly the vertical or yaw axis of the target compared to the BALL condition. The objects' size deviated from the size of the target cutouts by only 1-2 millimeters across their largest dimension. However, once the object was inserted into the cutout a small amount of orientation variability was possible, particularly along pitch and roll, because the lid was not rigid despite the lids' reinforcement with cardboard and masking tape (about 1-mm thick).
Fifty-two trials for each condition were collected in two blocks of 26 trials each. Blocks of 26 trials were used to ensure consistency in the initial orientation with respect to the hand and global coordinate system of the attached shapes on the large ball. All experiments began and ended with participants performing twenty-six trials of the ONLY condition because the large ball used in this condition was not covered with Velcro and there was no additionally attached shape. Thus, rotation of the large ball in the hand from trial to trial had not effect on the experimental condition. Moreover, the experimenter checked to ensure that subjects held the ball firmly in the palm of the hand with fingers wrapped around the ball before each trial so that the position in the hand did not vary. After the initial block of the ONLY condition, the Velcro covered large ball was grasped by the subject, creating a firm attachment to the hand. Thereafter, six blocks of 26 trials of combined Conditions 1-3 were performed. The two blocks of 26 trials for each of the BALL, TRAP and STAR conditions were presented randomly. For each block of 26 trials, the shape was screwed into the large ball's bolt until it locked, ensuring that each object's orientation with the hand did not differ across blocks of trials. The experiment finished with the Velcro covered ball being removed and replaced by the smooth ball of the same diameter for the final block of the ONLY condition. Approximately 30-sec occurred between each trial of a block, which was the time required for participants to return to the starting location and align the arm within the vacuum bag. Ten minutes rest was provided between blocks of trials.
Participant Instructions
Participants were instructed to initiate their reach toward the target after they received a verbal ‘go’ signal from the experimenter, and to reach to the target in one continuous movement. It was emphasized that this was not a reaction time task, i.e. they were to move after the go signal but they did not need to react to that signal as fast as possible. Participants were told to remain at the target until the experimenter gave the instruction to return. They were asked to produce one smooth, continuous movement to the target and then to either drop it into the target can (ONLY condition) or insert the attached shape into the slot in the target can's lid (BALL, TRAP or STAR conditions).
We attempted to control movement time (MT) by using 10 test trials prior to the main experiment where participants were instructed to move as quickly as possible to the target and to accurately insert the STAR into its slot. The average MT of the ten test trials was obtained and used as the target MT. The STAR condition was used because in pilot studies, participants tended to perform this condition more slowly. Subjects were asked to keep their MT consistent with this target MT across trials and conditions. Because of the number of trials required for each condition, we could not formally monitor and provide feedback about MT after each trial. However, we monitored MT for groups of trials and provided the participant with feedback about how well they were maintaining the target MT. In practice, however, participants more consistently controlled the transport time component of MT, or the time required to reach the target after movement onset, with more difference in the time of adjustment once reaching the target across conditions (see Results).
Data Collection and Processing
Data Capture
Three-dimensional arm and scapula kinematics were recorded at a sampling rate of 120 Hz using a VICON™ M13 motion measurement system with eight pole-mounted cameras arranged in a circle around the subject. The VICON cameras were calibrated before each data collection.
Static Calibration
A static calibration with the upper arm positioned forward, parallel to the table and perpendicular to the trunk, the elbow fully extended, the upper arm and forearm in mid-position rotation about their long axes, and the wrist positioned in neutral flexion-extension and abduction-adduction, the thumb pointing upward. This position was treated as the zero position for all joint angles and used as the reference position for the computation of joint angles from experimental trials (see below). The X-coordinate of each joint's local coordinate system in this position was directed from medial to lateral, the Y-coordinate pointed anterior to posterior, parallel to the axis of the upper arm and forearm, and the Z-coordinate pointed vertically upward, all axes being parallel to the corresponding axes of the global coordinate system. Because the target can was aligned with the global coordinate system, orientation of the hand with respect to the global system corresponded to the same orientation angles with the target.
Preliminary marker processing
Marker positions were low-pass filtered at 5-Hz using a bidirectional second-order Butterworth filter before computing other kinematic variables. Programs written with Matlab™ 7.1 software were used to perform all analyses.
Joint angle calculation
The computation of joint angles from the rigid body arrays was done in Matlab. The rigid bodies, located on the body segments, at each time frame of an experimental trial were rotated back into their position obtained during the static calibration trial (see above) by using the Sodervist and Wedin method (1993) to compute the associated rotation matrices. The product of two such adjacent rotation matrices was used to extract the Euler angles between adjacent segments in Z-X-Y order. The result yielded ten rotational degrees of freedom (DOFs): three at the clavicle/scapula (abduction-adduction about the Z-axis; elevation-depression about the X-axis and upward-downward rotation about the Y-axis), and shoulder (horizontal abduction-adduction about the Z-axis; flexion-extension about the X-axis and internal-external rotation about the Y-axis), and two DOFs at the elbow (flexion-extension about the Z-axis; forearm pronation-supination about the Y-axis) and wrist (flexion-extension about the Z-axis; abduction-adduction about the X-axis). Rodrigues' rotation formula was used to rotate the elbow flexion-extension axis from the X-axis of the global coordinate frame to the axis formed by markers placed at the medial and lateral epicondyles of the humerus (Murray, Li, & Sastry, 1994). The accuracy and reliability of the motion analysis system for estimating smaller joint angle changes than occurred in this experiment has been established previously (Hsu, Scholz, Schöner, Jeka, & Kiemel, 2007).
Kinematic Variables
The following kinematic variables were computed from the motion of the hand or joint angles:
1. Transport and Adjustment Times
The peak value of resultant velocity of the hand markers was used to define the onset of the hand trajectory as the time when a backward-search algorithm from the time of peak velocity encountered 2% of that peak value. The termination of hand transport phase (and, consequently, the onset of hand adjustment phase) was also automatically selected as 6% of that value after the peak resultant velocity was reached. Termination of the adjustment phase was defined as 10% of the maximum velocity after the termination of the transport phase.
These three selected points were visually checked using the 3D path of the reflective marker placed on the index finger interphalangeal joint. If the points did not reflect the correct selection for each phase, they were manually selected for correctness.
The time from movement onset to the termination of the transport phase was considered transport time. The time from transport phase termination to the end of movement was considered adjustment time.
The transport and adjustment phases were each time-normalized separately in order to compute variances across trials at each percentage of the trajectory. We initially considered three sub-phases of the transport phase: early (1-30%), middle (41-60%) and late (81-100%) transport phase, but focus here on the late transport phase. The adjustment phase was divided in half into early and late adjustment phases.
2. Range of joint excursion
For each subject, the range of joint excursion was determined as the average across trials of each joint's excursion for each condition, computed separately for the transport and adjustment phases. The joint excursions for each DOF are illustrated in the Results section. However, for statistical analyses, we combined the proximal and distal joint DOFs to examine their differential contributions to investigate differences that might occur with a strong or weak orientation constraint.
3. Hand Path Variability
The variance of the hand's position in each dimension was computed and then the norm of these three measures was computed, at each percentage of the movement. The norm was then averaged across each of the early, mid and late transport and early and late adjustment phases for statistical analyses.
4. Hand-target orientation
To compute the hand's orientation with the target, we first computed the unit vectors defining a local coordinate system embedded in the hand (i.e., the local x, y and z axes) using the rigid body model on the hand. The rotation matrix taking this coordinate system into the coordinate system of the target, which was aligned with the global system, was then estimated using Södeqvist and Wedin's (1993) approach at each point in the movement. Euler angles corresponding to pitch (rotation about the x-axis), roll (rotation about the y-axis) and yaw (rotation about the z-axis) were then extracted from the matrix. The mean orientation angle and its SD across trials for pitch, roll and yaw then was computed for each subject. We predicted that changes in the orientation angles would occur early in the transport phase, not restricted to when the target was being approached. The SD of the hand's orientation about the z-dimension of the target was predicted to distinguish most between the experimental conditions, with variability being smallest for the TRAP and STAR conditions because of the need to align the shape about the yaw axis to insert it. Of course, this fact does not preclude the participants from also controlling rotation about this axis in the BALL or even the ONLY condition.
As with joint space variability, we compared the difference between the maximum SD of hand-to-target orientation and the SD at the end of the adjustment phase for differences among conditions.
5. Uncontrolled Manifold (UCM) analysis
The UCM approach was implemented to analyze the structure of joint configuration variance relative to stabilization of the movement variables of interest (Scholz and Schöner 1999). The structure of variance is defined by the amount of joint variance that leads to a change in the value of a task variable, compared to the amount of joint variance that reflects motor abundance, i.e., different ways of combining the joints to achieve the same value of the task variable of interest. Considering the space of joint configurations, at any point in the time-normalized movement a manifold can be defined in that space within which changes of the joint configuration across repetitions yield an identical value of the task variable to which the joint configuration is being related. This component of variance is referred to as UCM variance (VUCM) and may be considered in some contexts as reflecting compensations for errors in the output of some joints' motion by adjustments in the motion of other joints (Scholz et al. 2000; Latash et al. 2007). Trial-to-trial variations of the joint configuration that do not lie within this manifold lead to unwanted changes in the value of the task parameter of interest, and is referred to as orthogonal variance (VORT; i.e., variance in the subspace of joint space that is orthogonal to the uncontrolled manifold.).
In the present study, the variance of the ten recorded joint angles (10-DOF) was partitioned into these two variance components to test the following hypotheses: that the nervous system stabilizes the value of 1) the hand's three-dimensional (3D) position along its path to the target and 2) the hand's orientation with respect to the three vectors defining the target position in external space. The Jacobian matrix forms a link between small changes in the joint configuration at each point in the movement and changes of the task parameters. The Jacobian for the 3D position control hypothesis was obtained by differentiating the geometric map relating changes in each of the ten joint angles to changes in the 3D hand position. Orientation was described using two orthogonal vectors based on the hand markers that were constant in the wrist frame. Two projections of one vector and one projection of the other vector onto the three axes of the global coordinate frame, with which the target's coordinate system was aligned, provide the constraint equations from which the Jacobian was computed by differentiating with respect to the joint angles. The null space of each Jacobian matrix was computed in Matlab™ representing linear approximations to UCMs for control of hand position or hand orientation. Then, at each point in normalized-time ‘i’ of a given trial, the configuration of joints (θi) was referenced to the mean joint configuration (θij) across all trials at the same time point, ‘j’, i.e., (θi - θij), and projected onto the null space computed based on the mean joint configuration at that time (i.e., null[J(θij)]), and into the orthogonal subspace. The variance of the projections into each subspace across trials is then computed and normalized to the number of DOFs of each subspace to allow comparisons across the two subspaces for each control hypothesis. For analyses related to stabilization of either the 3D hand position or 3D hand orientation, variance within the UCM was divided by 7 (VUCM), whereas variance in the subspace orthogonal to the UCM was divided by 3 (VORT). The normalized variances are reported here as VUCM and VORT, respectively. Mathematical details of the method can be found in (Reisman and Scholz 2003).
If VUCM is found to be significantly larger than VORT for either or both hypotheses, this would be consistent with previous studies indicating that stable control of a particular movement variable is accomplished by using motor abundant solutions to coordinate the motor elements, here the joint motions. If the stability of the 3D position, identified by differences between VUCM and VORT, is unaffected when a change in the task requires the precise control of hand orientation, this will suggest that the presence of motor abundance allows for simultaneous stabilization of both movement variables. In addition, we expect to see greater differences between VUCM and VORT related to the stabilization of orientation when more precise orientation is required.
We also examined the relative variance difference between the variance components, i.e. RVDIFF = (VUCM − VORT)/(VUCM+VORT), which is sometimes helpful to see more clearly how the two components differ. The closer the relative difference is to +1.0, the more that the joint variance reflects the use of motor abundance to stabilize a given performance variable. When this difference is close to zero or negative, this indicates that more of the variance of the joint combinations leads to variability of the performance variable.
Statistical Analyses
Either repeated measures analyses of variance (ANOVA) or repeated measures multiple analyses of variance were performed to test for the statistical significance of the experimental variables, as outlined in Table 1. Condition refers to the experimental condition, i.e. ONLY, BALL, TRAP and STAR. Phase refers to the transport and adjustment phases. The variance components for the analysis of joint configuration variance were VUCM and VORT. The level of significance was set at p<0.05 for all comparisons. If the interaction was significant, the condition effect was examined using another ANOVA or MANOVA, depending on the variable (see Table 1), performed on experimental condition, separately for each phase. Further investigations of the results were performed with post-hoc pairwise comparisons with Bonferroni corrections, as indicated in Table 1.
Table 1.
Statistical procedure for each experimental variable
| Two-Way ANOVA | Three-Way ANOVA | MANOVA Condition by Phase | |
|---|---|---|---|
| Movement Time | Condition by # Phase | ||
| Range of Joint Excursion | Condition by # Phase a | ||
| Hand Path Variability | Condition by * Phase | ||
| Hand-Target Orientation | Condition by # Phase b | ||
| Hand-Target Orientation Variability | Condition by # Phase b | ||
| Joint Configuration Variance (VUCM & VORT) | Condition by * Phase by Variance Component | ||
| Relative Difference of Joint Variance Components (VUCM − VORT)/(VUCM + VORT) | Condition by * Phase |
Post-Hoc Tests: SPSS pairwise comparisons table with Bonferroni corrections for all variables; For some comparisons following the MANOVA, the within-subjects contrast table in SPSS was used to investigate significant results.
Movement Phases: Two levels of the factor “phase” were included in these analyses, the transport and adjustment phases;
Movement Phases: Five levels of the factor “phase” was included in these analyses, three phases during hand/object transport (early, middle and late) and two phases during adjustment to the target (early and late).
Dependent Measures for MANOVA: scapula, shoulder, and combined elbow & wrist joint angles;
Dependent Measures for MANOVA: yaw, roll and pitch orientation angles
Results
Transport and Adjustment Time components of Movement Time (MT)
The two-way ANOVA revealed significant effects of experimental condition (F3, 27 = 21.5, p<0.0001) and phase of the task (F1, 9 = 8.5, p<0.05), as well as a significant condition by phase interaction (F3, 27 = 45.6, p<0.00001). Figure 3 shows that differences in the transport component of MT among the conditions were small. However, the ANOVA performed at this phase revealed that this difference was significant (F3, 27 =19.8, P<0.00001). Post-hoc tests found significant differences between the ONLY and BALL conditions (p<0.05) and between these two conditions compared to both the STAR and TRAP conditions (p<0.05). The transport time for the STAR AND TRAP conditions did not differ significantly (p=1.0).
Figure 3.
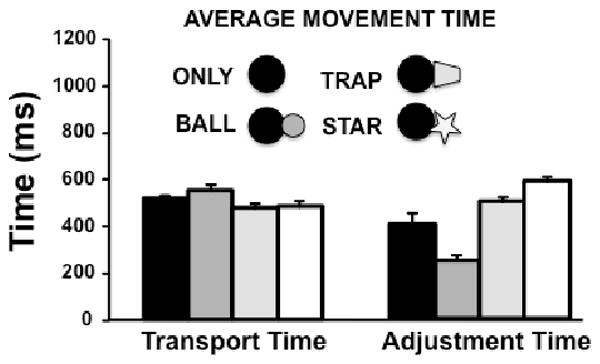
Average time ±standard error of the mean (SEM) encompassed by each phase of the task, hand transport and hand adjustment at the target.
The separate ANOVA performed on the adjustment phase revealed that this MT component also differed significantly among the conditions (F3, 27 = 36.7, p<0.000001). Post-hoc analyses showed that Adjustment time for the BALL condition was shorter than for the ONLY condition (p<0.05), and the TRAP and STAR conditions (p<0.00001). Adjustment time for the STAR condition was longest, on average, greater than for the TRAP or BALL (both p<0.01) or ONLY (p<0.05) conditions. The adjustment time for the ONLY condition was not significantly different, however, from that of the TRAP condition (p>0.45).
Range of Joint Excursion
Figure 4 illustrates the range of joint excursions during (a) the transport and (b) the adjustment phases for the four experimental conditions. Differences across conditions appeared to be relatively small for the transport phase (Figure 4a), although. For the repeated measures MANOVA, we combined excursions for the three scapula joint angles, the three shoulder joint angles and those of the four distal joint angles (i.e. elbow flexion-extension, pronation-supination, and wrist flexion-extension, abduction-adduction), treating them as dependent measures. The overall MANOVA revealed significant differences among the experimental conditions (F9, 81 = 6.06, p<0.00001), between the movement phases (F3, 7 = 81.2, p<0.00001), and a significant condition by phase interaction (F9, 81 = 4.9, p<0.0001). The univariate effects for all joint complexes were also significant.
Figure 4.
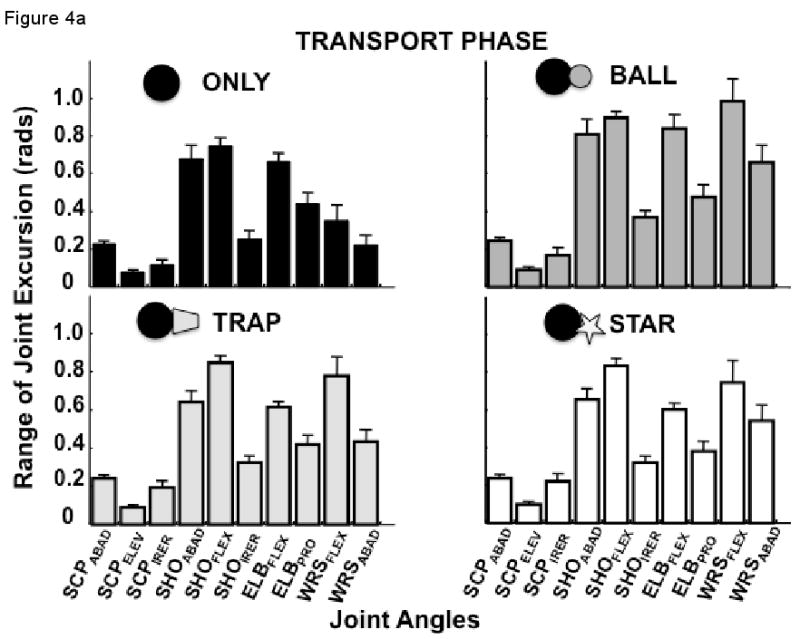
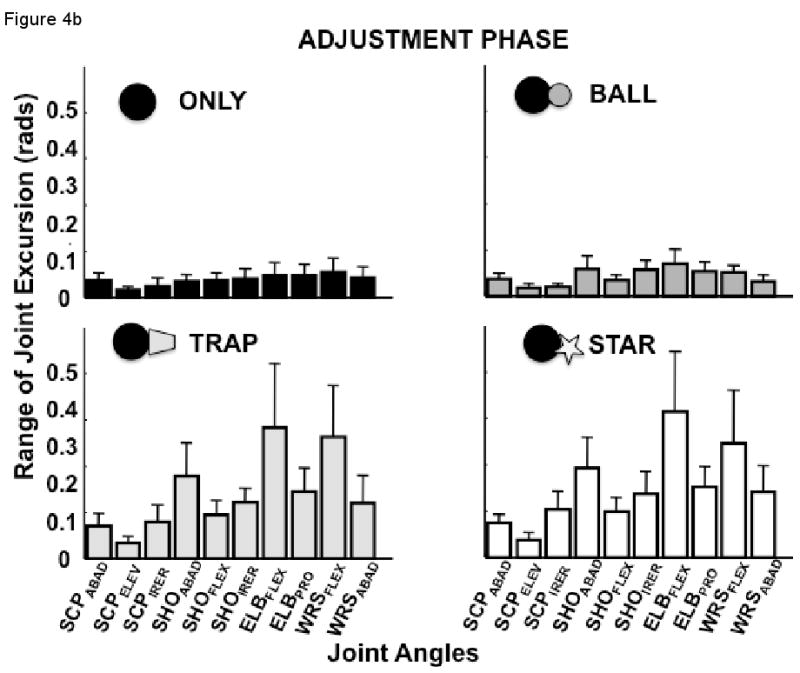
(a) Mean excursion (±SEM) of each joint across subjects and trials during the transport phase for the four experimental conditions; (b) Mean excursion (±SEM) of each joint across subjects and trials during the adjustment phase for the four experimental conditions.
For the transport phase, MANOVA revealed a significant condition effect for the scapula (F3, 27 = 4.2, p<0.05), shoulder (F3, 27 = 19.5, p<0.000001) distal (F3, 27 = 25.8, p<0.0000001) joint angle complexes. Post-hoc tests revealed that the excursion of the scapula angle complex was significantly less for the ONLY condition than for any of the other three conditions (all comparisons, p<0.05). However, there was no difference in this angular excursion among the BALL, TRAP or STAR conditions (all comparisons p>0.30). For the shoulder complex, the excursion was smallest for the ONLY condition compared to the BALL (p<0.0001) and TRAP (p<0.05) conditions, and approached being significantly smaller than the STAR condition (p=0.056). Excursion of the shoulder joint complex for the TRAP and STAR conditions did not differ (p>0.7) and were intermediate between the ONLY condition and the BALL condition (both p<0.01). Finally, for the distal joint complex (i.e. combined elbow and wrist joint angles), the condition differences were qualitatively identical to those for the shoulder joint complex. Shoulder joint excursion during performance of the ONLY condition was smaller than for the BALL (p<0.001), TRAP (p<0.01) and STAR (p<0.05) conditions, while the BALL condition had larger excursion, on average, than the TRAP (p<0.001) or STAR (p<0.0001) conditions. Once again, there was no difference in excursion between the STAR and TRAP conditions (p>0.08). Thus, even though all conditions required reaching to the same target, the amount of excursion for each condition during the transport phase was affected by the nature of the final action.
The MANOVA performed on the adjustment phase revealed a significant effect of experimental condition on excursions of the scapula joint complex (F3, 27 = 41.1, p<0.000001), the shoulder joint complex (F3, 27 = 53.6, p<0.00001), and the combined distal joints (F3, 27 = 38.0, p<0.0001) (Figure 4b). Interestingly, the pattern of differences were identical for all three joint complexes for the adjustment phase, with no difference between the ONLY and BALL conditions (scapula: p>0.67; shoulder: p>0.13; and distal joints: p>0.68) or between the TRAP and STAR conditions (scapula: p>0.08; shoulder: p>0.14; and distal joints: p>0.49), as revealed by post-hoc analyses. The excursions of all joint complexes were larger for the TRAP and STAR conditions than for the ONLY and BALL conditions (all comparisons, p<0.001). These results are indicative of the greater adjustments of joint positions required for appropriate object insertion in the more highly constrained conditions (STAR and TRAP).
Hand Path Variability
The average standard deviations of the resultant 3D hand position for each movement phase and condition are presented in Table 2. Differences in variability among the conditions depended on the phase of movement (F12, 108 = 3.3, p<0.001), as revealed by the two-way interaction in the repeated measures ANOVA. Further analyses revealed that there were no significant differences in hand path variability among conditions for the early or late transport phases or for the early or late adjustment phases as revealed by one-way ANOVA's (all p>0.1). Only for the mid-transport phase was there a difference in hand path variability among the conditions (F3, 27 = 4.22, p<0.05). Interestingly, hand path variability for the BALL condition was higher than any of the other conditions, but this difference reached significance only when compared to the ONLY condition (p<0.05).
Table 2.
Average standard deviation (m) of the resultant 3D hand position, computed across trials at each percentage of the movement, then averaged across each phase and all subjects (± standard error of the mean). ** = Significant differences, p<0.05.
| Phase | ONLY | BALL | TRAP | STAR |
|---|---|---|---|---|
| Early Transport | 0.0152 ± 0.0011 | 0.0166 ± 0.0016 | 0.0143 ± 0.0010 | 0.0134 ± 0.0009 |
| Mid Transport | **0.0287 ± 0.0026 | **0.0399 ± 0.0058 | 0.0313 ± 0.0035 | 0.0322 ± 0.0038 |
| Late Transport | 0.0235 ± 0.0020 | 0.0331 ± 0.0061 | 0.0297 ± 0.0034 | 0.0269 ± 0.0029 |
| Early Adjustment | 0.0220 ± 0.0018 | 0.0217 ± 0.0027 | 0.0223 ± 0.0020 | 0.0212 ± 0.0015 |
| Late Adjustment | 0.0212 ± 0.0021 | 0.0179 ± 0.0017 | 0.0182 ± 0.0017 | 0.0186 ± 0.0014 |
Mean hand-target orientation
Figure 5a depicts the yaw, pitch and roll angles (± standard error of the mean) of the hand with respect to the target coordinates, averaged across subjects. For all conditions, each angle begins to change relatively early in the transport phase, typically by at least 25% of the hand's trajectory. Additional small changes in orientation angle occurred during the adjustment phase, most notably for the TRAP and STAR conditions (Figure 5a), and less so for the BALL condition. In contrast, there appeared to be little additional change of the three orientation angles for the ONLY condition.
Figure 5.
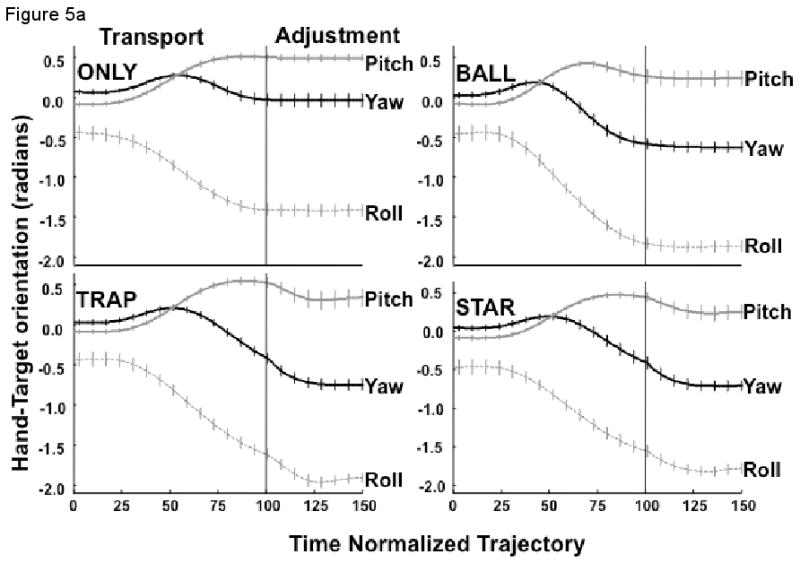
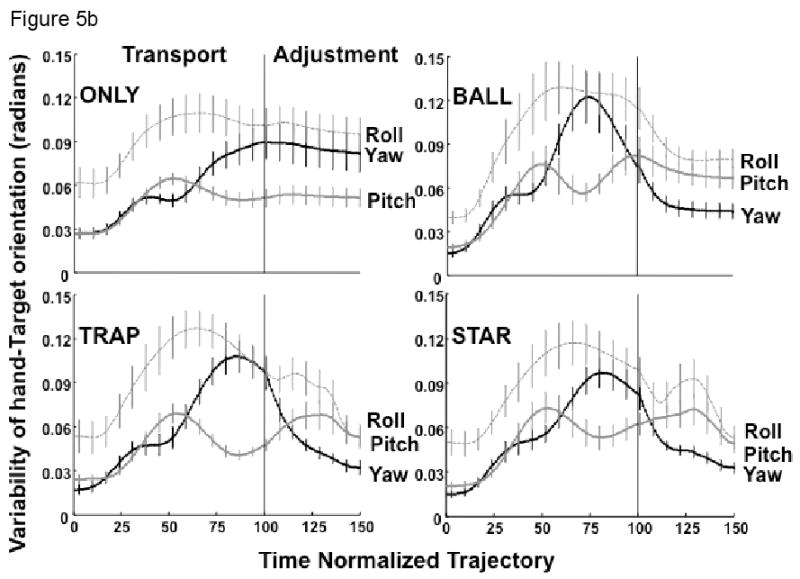
(a) The mean hand's orientation relative to the target for pitch (Z), roll (Y) and yaw (X) angles averaged across subjects (± SEM); (b) The average standard deviation across subjects (±SEM) of the hand's orientation angle to the target for pitch (Z), roll (Y) and yaw (X) angles. Control of the yaw angle was critical only for the TRAP and STAR conditions.
The amount of change of the yaw, pitch and roll angles differed between movement phases (F3, 7 =1.24, p<0.0001) and among the experimental conditions (F9, 81 = 7.30, p<0.0001), as revealed by the overall repeated measures MANOVA. The effect of experimental condition also depended on the movement phase (F9, 81 = 4.7, p<0.001), however. The univariate tests indicated that the main effects and the interaction were significant for all orientation angles. Post-hoc tests revealed that the change of all three orientation angles during the transport phase was greater for the BALL than the ONLY condition (all angles, p<0.01), greater for the TRAP than for the BALL condition (all angles, p<0.01), but was not different between the TRAP and STAR conditions (all p>0.11).
In contrast, the change of orientation angle did not differ between the BALL and ONLY conditions for the yaw, roll or pitch angles during the adjustment phase (all angles, p>0.09), as revealed by post-hoc comparisons. The change in orientation angle during the adjustment phase was again significantly larger for the TRAP compared to the BALL condition, the strongest effect being for the yaw angle (p<0.00001) compared to the roll (p<0.01) or pitch (p<0.05) angles. As was the case for the transport phase, there was no difference in the orientation angle change during the adjustment phase between the TRAP and STAR conditions (all p>0.14).
Hand-target orientation variability
Figure 5b presents the mean value across subjects of orientation angle variability (± standard error of the mean), computed as the standard deviation across trials. When the hand was at the target with the palm facing downward, pitch corresponded to rotation about an axis passing from the thumb to the little finger, roll corresponded to rotation about the long axis of the hand, and yaw corresponded to rotation about a vertical axis passing through the palmar to the dorsal surface of the hand (ulnar-radial deviation). To properly insert the trapezoid and star shaped objects into their corresponding target lid cutouts, yaw motion had to be well controlled, while control of the yaw angle was not crucial for either the BALL or ONLY conditions.
The overall MANOVA revealed a significant effect of experimental condition (F9, 81 = 4.4, p<0.0001) on the change of orientation angles. Although there was no main effect of phase (F3, 7 = 2.2, p>0.17), the condition by phase interaction was significant (F9, 81 = 2.7, p<0.01). Considering the transport phase, post-hoc comparisons indicated a larger change in the yaw angle for the BALL condition compared to the ONLY, TRAP or STAR conditions (all comparisons, p<0.01). The change in the yaw angle during transport was larger for the TRAP and STAR conditions than for the ONLY condition (p<0.05). There was no significant difference in the decrease of roll angle variability towards the end of transport among the BALL, TRAP and STAR conditions (all comparisons, p>0.2). However, only the decrease in variability for the TRAP condition was significantly larger than that for the ONLY condition (p<0.5). There was no significant difference between the TRAP and STAR conditions (all angles, p>0.25). No difference in the decrease in pitch angle variability during transport was found between the experimental conditions (all comparisons, p>0.3).
During the adjustment phase, the amount of change of the yaw angle did not differ between the ONLY and BALL conditions (p>0.1). The change in yaw angle was significantly smaller for the ONLY condition than for either the TRAP (p<0.001) or STAR (p<0.01) conditions, as revealed by post-hoc comparisons. This pattern of results was similar for the roll and pitch angles, with no differences between the ONLY and BALL (Roll: p>0.14; Pitch: p>0.17) conditions and significantly smaller changes in these orientation angles for the ONLY condition compared to the TRAP (Roll: p<0.0001; Pitch: p<0.05) or STAR conditions (Roll: p<0.001; Pitch: p<0.01). No difference was found between the TRAP and STAR conditions (all angles, p>0.08).
Joint configuration variance
Three-dimensional hand path
Figure 6a presents the results of the UCM variance analysis related to stability of the 3D hand path. For illustration, the variance components (VUCM, VORT) are presented after averaging across each percentage of the early, middle and terminal sub-phases of the transport phase and the early and late sub-phases of the adjustment phase.
Figure 6.
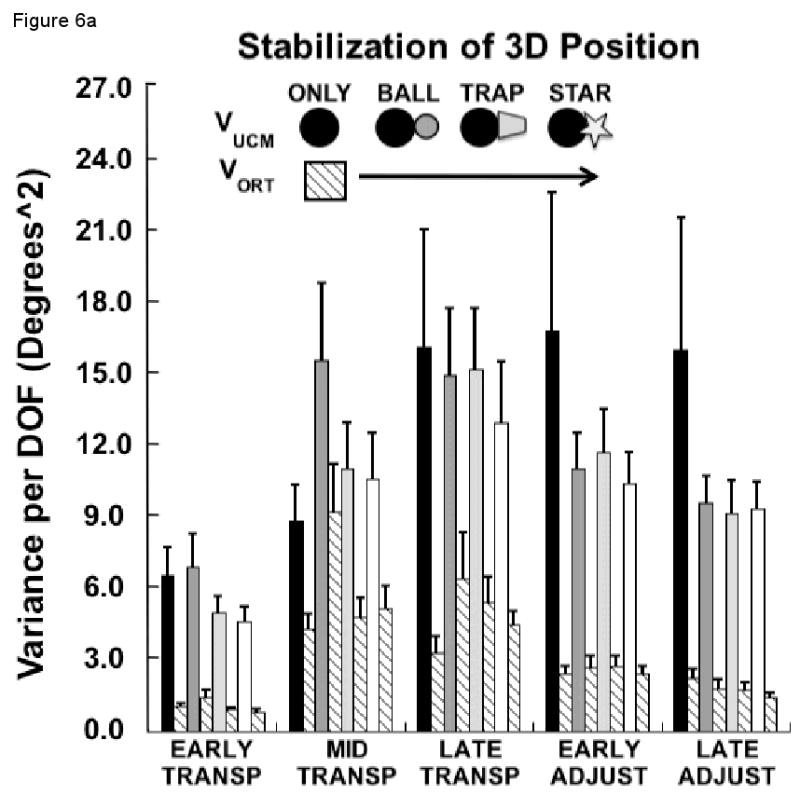
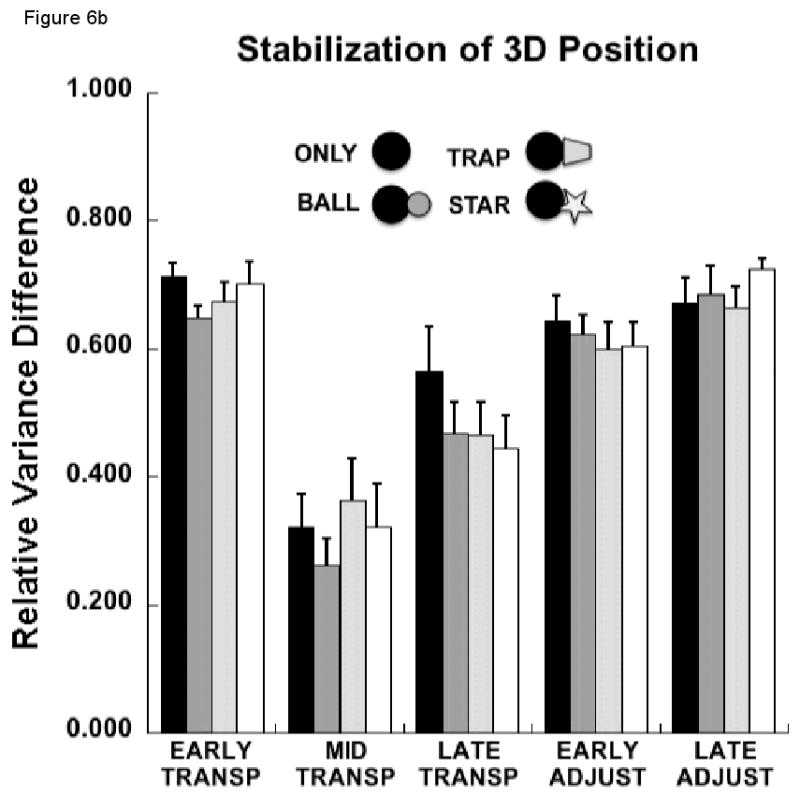
(a) Mean, across each sub-phase, of the components of joint configuration variance (VUCM and VORT) in degrees squared, averaged across subjects (±SEM) related to control of the 3D hand position for each experimental condition; (b) Mean, across each sub-phase, of the relative difference of joint variance components (VUCM - VORT / VUCM + VORT) averaged across subjects (±SEM) related to control of the 3D hand position for each experimental condition.
The three-way repeated measures ANOVA revealed that the joint variance component reflecting the use of motor abundance (VUCM) was greater than VORT (F1, 9 = 34.2, p<0.001). There was also a significant interaction between the experimental condition and phase (F12, 108 = 2.5, p<0.01), indicating that the total amount of joint variance differed between the conditions depending on the phase of the movement. To explore this further, we performed ANOVAs on each phase separately. Only during early (F3, 27 = 4.1, p<0.05) and middle (F3, 27 = 5.2, p<0.01) portions of the transport phase was there a difference in total variance between the conditions. During the early transport phase, this was due to significantly smaller total joint configuration variance in the STAR compared to the ONLY or BALL conditions (both p<0.05). In the middle phase of the transport phase, the BALL condition had significantly greater joint configuration variance than either the ONLY (p<0.05) or TRAP (p<0.05) conditions. The higher joint variance of the BALL condition compared to the STAR condition approached significance (p=0.06).
The ANOVA effects of greatest interest and most consistent with our experimental hypotheses, however, were the interactions between the factor variance component and factors experimental condition and movement phase. Of these interactions, neither the condition by variance component (p>0.37) nor the three-way (p>0.21) interactions were significant. The two components of variance changed differently across the phase of the movement, however (F4, 36 = 10.0, p<0.0001). Post-hoc analyses revealed that VUCM, reflecting the use of motor abundance, increased from early transport to the mid-transport sub-phase (p<0.01). It but did not differ significantly between the mid- and late transport sub-phases (p>0.86) or between the late transport and early adjustment sub-phases (p>0.26). There was a significant decrease of VUCM from the early adjustment to the late adjustment sub-phase (p<0.01, however.
The component VORT, which leads to variability of the 3D hand path, was smaller for the early transport sub-phase than for any of the other sub-phases (p<0.01). The increase of VORT from the early to mid-transport sub-phase, around the time of peak hand path velocity, is consistent with results of previous studies of pointing tasks (Tseng et al. 2002; Tseng and Scholz 2005a, 2005b). This component of variance did not differ between the mid- and late transport sub-phases (p=1.0), but it decreased significantly between the late transport and the early adjustment sub-phases (p<0.01) and from early to late adjustment sub-phases (p<0.01).
Figure 6b displays the relative variance difference (i.e., RVDIFF = [VUCM − VORT]/[VUCM − VORT]) across both condition and phase. This measure differed depending on the phase of the movement (F4, 36 = 33.5, p<0.0000001). There was no effect of condition (p>0.5), however, nor a condition by phase interaction (p>0.19). RVDIFF was smallest for the mid transport phase, around the time of peak velocity, due to a proportionally larger increase in VORT than VUCM during this phase. Post-hoc tests indicated that RVDIFF was significantly smaller during this sub-phase than during the early transport (p<0.0001) or the early (p<0.01) and late adjustment (p<0.001) phases. The difference in RVDIFF between mid- and late transport phases did not reach significance (p=0.075). The magnitude of RVDIFF gradually increased between late transport and early adjustment phases (p<0.05) and between early and late adjustment phases (p<0.01), reflecting the smaller decrease in VUCM than VORT during these sub-phases (Figure 6a).
Three-dimensional hand-target orientation
Figure 7a presents results related to the control of the hand's orientation with respect to the three axes defining the target's orientation, yaw, pitch and roll. On average, VUCM was larger than VORT (F1, 9 = 63.2, p<0.0001), although the strength of this difference depended on the combination of condition and movement phase (F12, 108 = 2.9, p<0.01). For example, during the late transport phase, the variance components did not differ significantly (p>0.06), and this did not depend on the experimental condition (p>0.39). Thus, there was no evidence for a synergy stabilizing hand-target orientation during this phase of the task.
Figure 7.
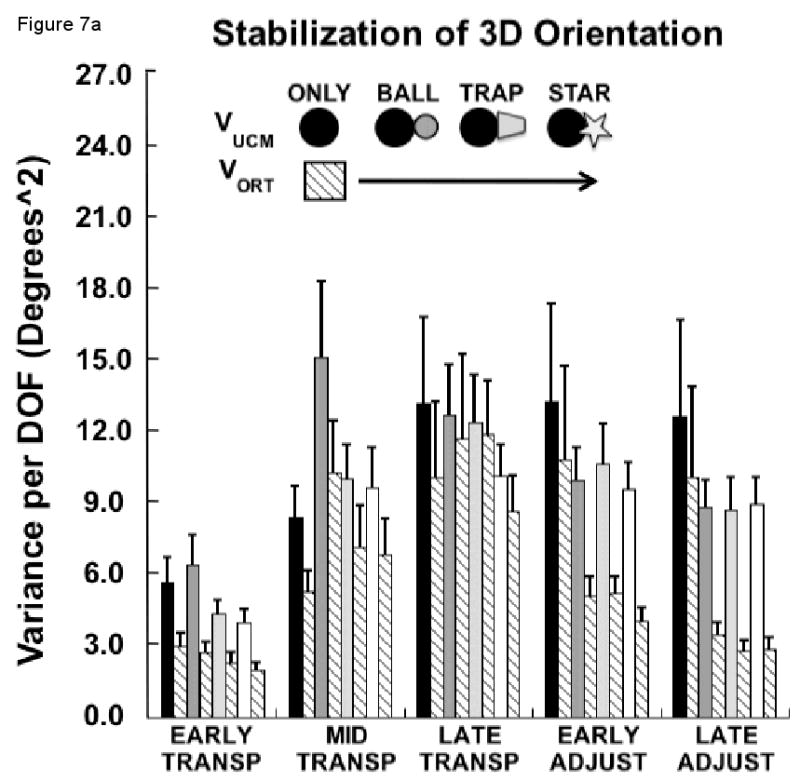
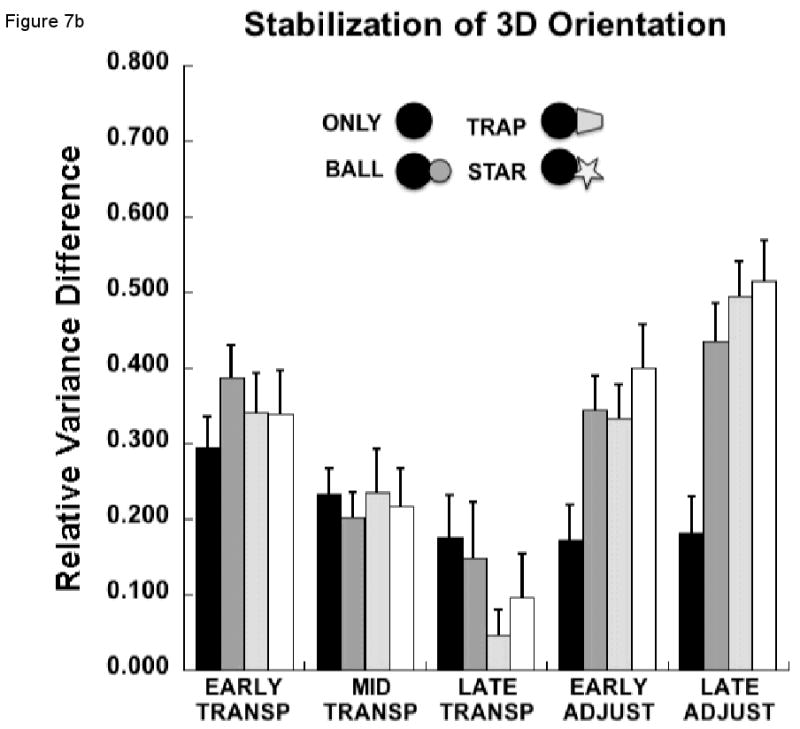
(a) Mean, across each sub-phase, of the components of joint configuration variance (VUCM and VORT) in degrees squared, averaged across subjects (±SEM) related to control of the 3D orientation of the hand to the target for each experimental condition; (b) Mean, across each sub-phase, of the relative difference of joint variance components (VUCM - VORT / VUCM + VORT) averaged across subjects (±SEM) related to control of the 3D orientation of the hand to the target for each experimental condition.
In contrast, there was a significant condition by variance component interaction for both the early (F3, 27 = 4.9, p<0.05) and late (F3, 27 = 5.3, p<0.01) sub-phases of the adjustment phase. Simple interaction effects between condition and the variance component were examined to further explore these significant interactions. During the early adjustment phase, the difference between the variance components differed depending on the experimental condition (F3, 27 = 5.3, p<0.01). Post-hoc tests revealed that the difference between VUCM and VORT was minimal for the ONLY condition compared to a large difference between the variance components for the BALL condition (p<0.01). However, the difference between VUCM and VORT was not different between the BALL and TRAP conditions (p>0.58) or between the TRAP and STAR conditions (p>0.93).
The same result was true for the late adjustment phase, with the difference between VUCM and VORT being significantly smaller for the ONLY condition compared to the BALL condition (p<0.05). The difference between the variance components did not differ between the BALL and TRAP (p>0.62) or between the TRAP and STAR (p>0.77) conditions. Thus, the synergy stabilizing hand-target orientation was only present for the three conditions that required controlled orientation during the adjustment phase.
These results were confirmed by examination of RVDIFF (Figure 7b). Note that higher RVDIFF indicates a stronger synergy stabilizing hand-target orientation. The two-way ANOVA indicated that the magnitude of RVDIFF depended on the phase of the movement (F4, 36 = 11.4, p<0.00001). Overall, RVDIFF related to stabilizing the hand-target orientation decreased from the early through the late transport phase and increased again thereafter (Figure 7b). However, the change in the magnitude of RVDIFF with phase also depended on the experimental condition (F12, 108 = 6.6, p<0.0000001). However, post-hoc contrasts revealed that this was true only for the early adjustment (F3, 27 = 5.99, p<0.01) and late adjustment (F3, 27 = 13.56, p<0.0001) phases, while RVDIFF did not differ among the experimental conditions during the early (p>0.20), mid (p>0.85) or late (p>0.33) transport phases. If anything, the synergy stabilizing hand orientation during the late transport phase was, on average, stronger for the two conditions requiring least orientation stability. During early adjustment to the target, RVDIFF for the ONLY condition was significantly smaller compared to the BALL condition (p<0.05), but RVDIFF did not differ between the BALL and TRAP (p>0.56) or between the TRAP and STAR (p>0.14) conditions, as revealed by post-hoc contrasts. For the late adjustment phase, RVDIFF for the ONLY condition was again smaller than for the BALL condition (p<0.01), while this measure did not differ between the BALL and TRAP (p>0.27) or between the TRAP and STAR (p>0.61) conditions.
Discussion
The present study was designed to investigate the role of motor abundance in facilitating the simultaneous resolution of position and orientation constraints by presenting experimental conditions that differed in their hand-target orientation requirement. The STAR and TRAP conditions produced the strongest orientation constraint by requiring subjects to control the hand's orientation to all three of the target axes. In contrast, the ONLY condition provided no explicit constraint on orientation because although the same sized, hand-held ball had to be transported an identical distance to the same final target location, it could be dropped into the open coffee can with a variety of orientations of the hand to the target axes. The orientation constraint for the BALL condition fell in between these extremes given that stabilization of the yaw angle was not essential for inserting the small ball that was attached to the hand-held ball into the similarly sized circular cutout of the target. Of course, subjects could have stabilized object orientation by default regardless of the explicit orientation constraint. However, the orientation requirements imposed by the different shapes, in particular, the trapezoid and star shapes, apparently were effective in constraining the hand's orientation. This was revealed by differences among the conditions in the angular excursions, the change in hand-target orientation during the transport and adjustment phases, as well the amount of variability of hand-target orientation about the yaw, pitch and roll axes.
The joint excursions differed less among the conditions during the transport phase compared to the adjustment phase. However, there were smaller excursions of the angles of the scapula complex, the shoulder complex, and the combined distal joint angles during transport to the target for the ONLY condition when compared to the other three conditions. This result suggests that there was some preparation during hand transport related to the hand-target orientation requirements.
During the adjustment phase, excursions of all joints were significantly larger in for the TRAP and STAR conditions compared to the ONLY and BALL conditions, whose excursions did not differ from one another. This difference is not surprising for the ONLY condition, which required minimal orienting of the hand. Although the adjustment time for the ONLY condition was shorter than that of the STAR condition, adjustment time for the ONLY condition did not differ from the TRAP condition. Therefore, this difference in excursion is unlikely due to differences in adjustment time. The longer than expected adjustment phase for the ONLY condition was due to a tendency to pause at the target before dropping the ball, likely the result of an attempt to ensure that the large ball would not hit the side of the can when dropped and miss the target. The joint excursion result is more surprising for the BALL condition, which did require orientation along the roll and pitch axes.
The striking difference among conditions between the movement phases appears to justify this separation for the other analyses. Given the larger joint excursions for the TRAP and STAR conditions, the presence of signal-dependent noise would lead to the expectation of higher joint variance affecting the hand's position or its orientation when compared to the BALL and ONLY conditions (Hamilton and Wolpert 2002; Harris and Wolpert 1998). The fact that joint variance leading to hand position or orientation variability was actually smaller in the former conditions, at least compared with the ONLY condition, attests to the fine control of orientation being exerted that was unrelated to overall joint excursion.
In addition to differences in joint excursions, the conditions differed in the amount of change of the orientation angles during the transport and adjustment phases. Orientation of the hand to the target began to change relatively systematically during the early phase of hand/object transport for all orientation angles (Figure 5a). The amount of change was smallest for the ONLY condition compared to the BALL and TRAP conditions, but not consistently so compared to the STAR condition. During adjustment to the target, the average change in the yaw, pitch and roll angles was not different between the ONLY and BALL conditions but was greater for the TRAP and STAR conditions, particularly for the yaw angle.
As with the change in average orientation angles, changes in the across-trials variability of yaw, pitch and roll angles differed among the experiment conditions even during the transport phase (Figure 5b). All orientation angles increased their variability between the early and mid sub-phases of hand/object transport. In most cases, this variability decreased again toward the end of the transport phase, most consistently for the yaw and roll angles. This was not true, however, for the ONLY condition, which exhibited a continued increase in across-trials variability up to the end of the transport phase. Stability of the yaw angle was, of course, not important for this condition. For the adjustment phase, variability continued to drop for the yaw and pitch angles and less consistently again for the pitch angle. This was true, however, only for the BALL, TRAP and STAR conditions, with the largest decrease for the latter two conditions. Variability of all orientation angles for the ONLY condition did not change during the adjustment phase from its value at the end of the transport phase. These findings confirm that participants did not control orientation as strongly when performing conditions with weaker orientation constraints.
Role of motor abundance in resolving the task constraints
To test the hypothesis that motor abundance plays an important role in resolving task constraints simultaneously, we used the UCM approach, which partitions variance of the motor elements into two components with respect to different movement variables hypothesized to be important for task success. One component of variance (VUCM) has no affect on the variable under consideration. Its magnitude reflects the use of motor abundance. The other variance component (VORT) leads to (presumably unwanted) variability of the performance variable. We predicted that if motor abundance makes easier the resolution of the two task constraints investigated here, i.e., hand position and orientation, then the UCM analysis related to stabilization of the hand's position at each point in its path to the target largely should remain unaffected by the degree of orientation constraint provided by the task. This may seem trivial given that both hand position and orientation need to be well controlled to put the object into the cutout. However, although VORT needs to be kept minimal to stabilize hand position, VUCM does not. Moreover, we predicted that the UCM results related to the stabilization of the 3D hand-target orientation should be stronger for the tasks requiring greater orientation control. That is, the proportion of total variance contributed by VUCM should be greatest for the TRAP and STAR conditions and somewhat lower for the BALL condition.
The results of the UCM analyses confirm these hypotheses, except for the BALL condition. The relative amount of VUCM and VORT computed with respect to the hand position differed across the transport and adjustment phases for all conditions. These results are consistent with previous reports of pointing tasks (Tseng et al. 2002; Tseng and Scholz 2005). Thus, the results of the UCM analysis related to stabilization of the 3D hand position support the contention that adding an orientation constraint does not affect position control. There was a tendency for VORT and particularly VUCM to be smaller in relation to position control during the adjustment phase for the three conditions requiring some hand-target orientation compared to the ONLY condition. This suggestion may reflect the need to resolve the additional constraint by restricting some joint combinations within the UCM for hand position control. However, these differences were not significant, due in large part to large individual variability for the ONLY condition. The fact that VUCM was still significantly higher than VORT with respect to position control during the adjustment phase, however, suggests that joint variations related to the control of hand orientation were constrained to limit their impact on position control. This result is consistent, in part, with the finding of Wang (1999) that the variability of the reach trajectory was little affected by different hand orientations except during the deceleration phase. The fact that they used different target orientations likely accounts for the difference during movement deceleration.
The transient increase in VORT in the middle of the movement resembles similar results in studies of multi-finger quick force production (Latash et al. 2002; Goodman et al. 2005). According to a model developed by Goodman and colleagues (Gutman et al. 1993; Goodman et al. 2005), this increase in variance may be related to variability of a timing parameter at the level of planning the trajectory.
Examination of the UCM results related to 3D hand orientation further confirmed our main hypothesis, and was consistent with the trends in hand-target orientation variability (Figure 5). When control of hand-target orientation was less critical (i.e., the ONLY condition), both VORT and VUCM were of similar magnitude during the late transport and both adjustment sub-phase. This was confirmed by the RVDIFF analysis, which can be used as an index of the strength of the synergy stabilizing a given performance variable (Latash et al. 2007). This index was substantially and significantly smaller for the ONLY condition compared to the other conditions during the adjustment phase. This finding is important because despite higher VORT associated with hand-target orientation for the ONLY condition and to a lesser extent for the BALL condition compared to the TRAP and STAR conditions (Figure 7a), this joint variability had little effect on indices of stabilization of the hand's 3D position (Figure 6b).
These results are consistent with recent findings of Zhang et al. (2008) who studied the coordination of a set of fingers during performance of a force production task with and without an additional constraint on the control of the moment of force. Their results showed that participants were able to successfully resolve the moment control constraint without adversely affecting the control of total force output.
The general idea that several patterns of co-variation among elemental variables related to the stabilization of different performance variables (multi-element synergies) can co-exist without interference is closely related to the principle of superposition introduced in robotics for the control of artificial grippers (Arimoto et al. 2000, 2001). According to this principle, complex motor tasks are reduced to a set of sub-tasks that are controlled by independent controllers. The output signals of the controller converge onto the same set of actuators where they are summed up. This method of control has been shown to lead to a decrease in the computation time as compared to control of the action as a whole. The principle of superposition has been confirmed in experimental studies of human prehension (Shim et al. 2003, 2005; Zatsiorsky et al. 2004). The current results, as well as the mentioned results of Zhang and co-authors (2008), may also be seen as examples of the principle of superposition afforded by the motor abundance of the multi-joint and multi-digit systems.
Is hand orientation controlled throughout the reach?
A number of recent studies have investigated the relationship between the control of hand position and hand orientation during the performance of skilled upper extremity tasks, with mixed conclusions (Cuijpers et al. 2004; Desmurget et al. 1996; Desmurget and Prablanc 1997; Fan et al. 2006; Fan and He 2006; Gosselin-Kessiby et al. 2008; Ma-Wyatt and McKee 2007; Marotta et al. 2003; Mitra and Turvey 2004). Soechting and Flanders (1993), for example, investigated the relationship between errors in orienting the hand to a cylindrical target when at the same or different spatial locations relative to the hand's location. The authors found that errors in hand-to-target orientation depended the target's spatial location when the target was visible. They nonetheless concluded that separate channels exist for estimating target orientation and spatial location, based in part on the finding that both variable and constant errors of hand-to-target orientation to a remembered target were independent of the target's location. This finding suggests the possibility of separate channels for control of these two features as well. Wang (1999) suggested that hand path and hand orientation were independently controlled based on the finding that several features of reaching, including path curvature and the timing of peak movement events, were unrelated to the target's orientation. Similar conclusions were drawn by Fan and colleagues based on the study of reaching to predictably and unpredictably perturbed target orientations in humans (Fan et al. 2006) as well as the results of cortical recordings during primate reaching (Fan and He 2006). The latter study revealed separate cortical cells firing in relation to hand transport and hand orientation. Nonetheless, other cells were reported to fire in response to both movement parameters, making questionable this interpretation (see also Stark et al. 2007).
In contrast, the results of several studies have led to the conclusion that hand transport and orientation are planned and controlled together. Desmurget et al. (1996) came to this conclusion based on the fact that the movement trajectory of a marker on the wrist was strongly affected by a cylinder's orientation and by the occurrence of perturbations of that orientation after reach onset (see also Mamassian 1997). Gosselin-Kessiby et al. (2008) reported a strong relationship between the peak velocity of hand orientation and hand transport during reaches requiring the insertion of a cylinder into a slot, leading them to conclude that hand orientation and position are “functionally coupled”. However, participants were instructed in that experiment to orient and transport the hand simultaneously. Similarly, in contrast to Wang (1999), Mitra and Turvey (2004) found that previously described reaching invariants such as bell-shaped velocity profiles and quasi-linear hand paths were violated when variations of hand orientation were required, suggesting that orientation and transport are not controlled independently. The differences in these studies were likely due to differences in the experimental design as well as statistical methods used to investigate the relationship between hand orientation and position.
On the one hand, our finding that the three orientation angles began to change their values early in the transport phase and continued to do so throughout that phase could suggest that the control of orientation and position are coupled (Figure 5a). However, we also found that hand-target orientation variance increased up until the late transport sub-phase. More importantly, the finding of similar indices of stabilization of the hand's 3D movement trajectory (i.e. RVDIFF) across conditions that had substantially different orientation constraints is compatible with the idea of independent control of trajectory and orientation.
Although movement parameters related to the hand's path might be correlated to changes in hand orientation, such findings do not themselves address the question of independent control of position and orientation. Moreover, unless independent DOFs are involved in the production of hand transport and hand orientation, the question of independence of control would be difficult to answer with correlation or multiple regression techniques typically used in the prior investigations. An extension of the ‘visuomotor channels’ hypothesis suggests that hand transport and orientation are separate processes carried out by different control mechanisms within the ‘dorsal stream’, and implemented by different effectors (Arbib 1981; Jeannerod 1981, 1999). However, despite evidence in favor of this hypothesis (Jeannerod 1999), several studies have revealed a contribution of proximal joints to hand-target orientation as well as to hand transport (Desmurget et al. 1996; Marotta et al. 2003; Gosselin-Kessiby et al. 2008; Wang 1999). Consistent with those results, we found substantially greater excursions of some of the clavicular/scapular and shoulder joints for the conditions requiring the strongest orientation to the target compared to the other conditions.
Related to this question is the identification of when orientation of the hand to the target actually begins during hand transport. Koshland and Hasan (1994) examined two-dimensional, planar reaching involving three joints and reported initial muscle activation that was independent of the required final hand orientation. Their result suggested that orientation is not controlled until the final part of reaching. If proximal joints play a role both in hand orientation and transport, then one could expect hand transport and orientation to evolve together during the reach (e.g., Cuijpers et al. 2004; Desmurget et al. 1996; Fan et al. 2006). In an earlier study of pistol shooting (Scholz et al. 2000), we found that stabilization of the gun's orientation to the target was present from the onset of movement until the time of shooting, while other variables such as the hand's absolute spatial position or the arm's center of mass coordinates were stabilized only early in the movement. Similar stabilization of these variables early in the movement was related, perhaps, to the fact that the position and orientation of the arm and gun both were well controlled initially, not necessarily that they involved dependent control processes. The evidence for stabilization of the hand-object orientation (Figure 7a) as well hand position (Figure 6a) during the early sub-phase of hand/object transport in the current experiment may have a similar origin.
Most studies to date investigating this issue have not considered explicitly the role of motor abundance in resolving various constraints on task performance. Without motor abundance, the control of hand transport and orientation would likely interfere with one another. It might be preferable in that case to resolve the two task constraints sequentially, orienting the hand only after reaching the target. Given motor abundance, however, simultaneous solution of these constraints is theoretically possible without mutual interference.
Cuijpers et al. (2004) examined, for example, the relationship between hand transport, hand orientation and grip aperture in relation to reaching to and grasping cylinders having different orientations. They reported that hand orientation changed gradually during the reach, explaining about 49% of the variance of final hand orientation by the time that the reach had achieved 35% of the target distance and explaining 90% of the final variance before the reach was 80% complete (see Figure 9 of Cuijpers et al. 2004). Those findings led the authors to conclude that orientation is specified gradually as the reach progresses. One might conclude from this that position and orientation are controlled together (see for example Desmurget et al. 1996). Cuijpers et al. emphasized that the correlation between early and late orientation does not mean that systematic errors in orienting the hand to the target do not occur. Indeed, their participants were reported to make such errors. However, we could expect that any variables expected to change values over time would show such a correlation, unless the orienting occurred only at reach termination. Although our task was different, we computed a similar measure to contrast differences in conclusions that might result from correlation approaches compared to the UCM analysis. Table 3 presents the results of this analysis from our current data. As with the results of Cuijpers et al. (2004), one could conclude that hand orientation was gradually specified as the reach progressed. This is, of course, not the conclusion we would arrive at from the UCM results. Results of the UCM analysis suggest that hand orientation was not well stabilized until the hand reached the target (Figure 7). Indeed, the stability of hand orientation was significantly diminished during the late transport phase, with VUCM minimally different from VORT and less so for the TRAP and STAR conditions compared to the ONLY and BALL conditions, only to reemerge during the adjustment phase. That is, during late transport, the variability of joint motions was as likely to lead to variability of the hand's orientation to the target as it did to stability of that orientation across repetitions. It is also of interest that the two conditions requiring the most precise orientation to insert the object had the smallest amount of terminal variability of hand orientation explained at 75% of hand transport (Table 3).
Table 3.
Variance of the final hand-target orientation explained by the hand's spatial orientation at three different percentages of hand transport to the target.
| Condition | 25% of Transport | 50% of Transport | 75% of Transport |
|---|---|---|---|
| ONLY | 0.548±0.078 | 0.867±0.002 | 0.983±0.00005 |
| BALL | 0.310±0.175 | 0.829±0.004 | 0.971±0.00006 |
| TRAP | 0.299±0.121 | 0.693±0.007 | 0.927±0.00025 |
| STAR | 0.342±0.088 | 0.697±0.006 | 0.917±0.00090 |
It is possible that this result was due to the nature of our target setup. That is, although the height of the coffee can was well below eye level, participants' line of sight to the cutout for object insertion in the can's lid was not ideal (Figure 1). Positioning the can on its side and facing the participants might have resulted in the earlier and more continuous control of hand orientation. Nonetheless, the results indicate that the control of hand position and orientation are not necessarily dependent and that the extent of their mutual control likely is related to the task requirements (e.g. Asuini et al. 2008).
On the use of abundance in reaching tasks
The results of the UCM analysis lead to a different conclusion about the control of reaching and orienting tasks than do several other studies of such tasks. For example, Desmurget and colleagues (1996) presented results suggesting that for a given initial hand position and a fixed location and orientation of the target, participants used a unique combination of joints to acquire the target (see also Greá et al. 2000). Consistent with this conclusion, Rosenbaum et al. (1999) argued that goal-directed movements are planned in joint space as sets of angles representing a desired or a reference posture. Fan et al. (2006) concluded that although motor abundance allows the attainment of a final hand position and orientation, their experimental results suggested reproducible arm and hand trajectories from trial to trial for the same target orientation and starting position (see also Flanders et al. 1992).
The conclusions of these studies have typically been based on analyses that focused on mean performance or that used multiple regression methods that depended on supporting the null hypothesis (e.g., Desmurget et al. 1996). In addition, the number of DOFs available to participants has often been limited. For example, Fan et al. (2006) analyzed only five of the seven possible joint angles of the arm, excluding the contribution of scapular motion. Moreover, no data on joint variability were presented in their report. In contrast, the current study measured and analyzed the motion of all seven joint angles of the arm as well as three clavicular/scapular motions. In contrast to those other results, participants in the current study exhibited significantly higher joint variability that was consistent with the use of multiple joint combinations to achieve an identical final hand position and hand-target orientation than joint variability leading to variability of those performance variables. This was the case despite our control of the initial hand position and hand-object orientation as well as the final target location and orientation.
Certainly, participants limit, more or less, the range of all possible joint combinations that are actually used to accomplish such a task. Some limitation results, of course, from joint excursion limits (Kamper and Rymer 1999). Individuals have been shown to typically use a particular sharing pattern among the motor elements for a given task (Li et al. 1998). For kinematic tasks, this may result in part from subjects avoiding otherwise possible combinations of joints (in terms of accomplishing the goal) for reasons of comfort. Nonetheless, those limits are relative, the sharing pattern representing only the average performance. The results of the current study are consistent with those of many previous studies of upper extremity tasks, showing that the use of motor abundance is a common feature of their control (Scholz et al. 2000; Tseng et al. 2002; Tseng and Scholz 2005a, 2005b; Tseng et al. 2006). The present results extend those findings by providing support for the hypothesis that an important advantage of motor abundance is the ability to resolve multiple task constraints simultaneously without significant interference.
Why are certain patterns of co-variation among elemental variables preferred when performing tasks involving abundant motor systems? Earlier studies emphasized a drop in variability of potentially important performance variables that could be achieved by such co-varied adjustments of elemental variables (Scholz & Schöner 1999; reviewed in Latash et al. 2002b). However, several recent studies have shown that indices of performance variability for abundant systems do not always differ substantially from such indices of non-abundant systems in comparable tasks. In particular, accurate, rapid force production tasks were performed with comparable indices of accuracy during one-finger (non-abundant) and multi-finger (abundant) tasks (Gorniak et al. 2008; Shapkova et al. 2008; but see Latash et al. 2001 for conflicting evidence). These findings, as well as the findings of the current study, have suggested that the main purpose of co-variation at the level of elemental variables may not be to reduce variability per se but to allow the system to stabilize several important performance variables simultaneously without sacrificing accuracy of performance of any of them. In other words, such multi-element synergies allow us to open the door by pressing on the handle with the elbow while carrying a cup of hot coffee in the hand.
Figure 2.
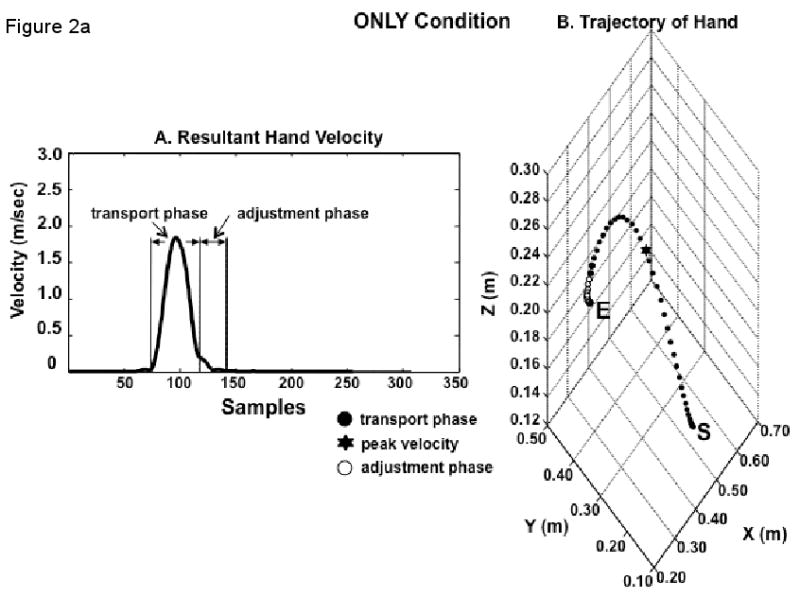
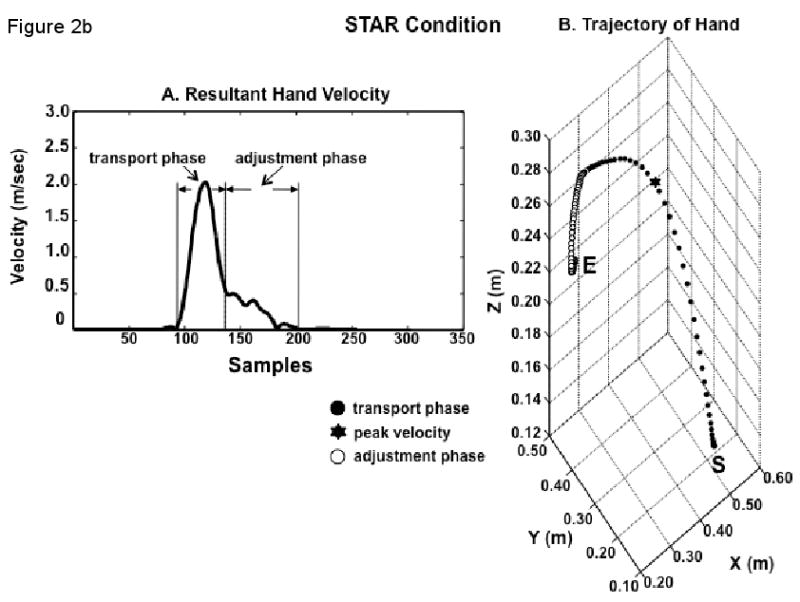
(a) Illustration of determination of the transport and adjustment phases for the ONLY condition using the resultant velocity (A) and changes in the smoothness of the three-dimensional hand path trajectory based on a marker placed on the proximal interphalangeal joint of the index finger (B). S = starting position; E = end of movement; (b) Illustration of estimates of the transport and adjustment phases for the STAR condition using the resultant velocity (A) and changes in the smoothness of the three-dimensional hand path trajectory based on a marker placed on the proximal interphalangeal joint of the index finger (B). S = starting position; E = end of movement.
Acknowledgments
Support for this work was provided by grant NS-035032, awarded to Mark Latash and grant NS050880, awarded to John Scholz.
References
- Ansuini C, Giosa L, Turella L, Altoe G, Castiello U. An object for an action, the same object for other actions: effects on hand shaping. Experimental Brain Research. 2008;185(1):111–119. doi: 10.1007/s00221-007-1136-4. [DOI] [PubMed] [Google Scholar]
- Arbib MA. Perceptual structures and distributed motor control. In: Brooks VB, editor. Handbook of Physiology, Section I: The nervous system. 2: Motor Control. Baltimore: Williams & Wilkins; 1981. pp. 1449–1480. [Google Scholar]
- Arimoto S, Nguyen PTA, Han HY, Doulgeri Z. Dynamics and control of a set of dual fingers with soft tips. Robotica. 2000;18:71–80. [Google Scholar]
- Arimoto S, Tahara K, Yamaguchi M, Nguyen PTA, Han HY. Principles of superposition for controlling pinch motions by means of robot fingers with soft tips. Robotica. 2001;19:21–28. [Google Scholar]
- Cuijpers RH, Smeets JBJ, Brenner E. Journal of Neurophysiology. 2004;91:2598–2606. doi: 10.1152/jn.00644.2003. [DOI] [PubMed] [Google Scholar]
- Danna-Dos-Santos A, Slomka K, Zatsiorsky VM, Latash ML. Muscle modes and synergies during voluntary body sway. Experimental Brain Research. 2007;179(4):533–550. doi: 10.1007/s00221-006-0812-0. [DOI] [PubMed] [Google Scholar]
- de Freitas SM, Scholz JP, Stehman AJ. Effect of motor planning on use of motor abundance. Neuroscience Letters. 2007;417(1):66–71. doi: 10.1016/j.neulet.2007.02.037. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Desmurget M, Prablanc C. Postural control of three-dimensional prehension movements. Journal of Neurophysiology. 1997;77(1):452–464. doi: 10.1152/jn.1997.77.1.452. [DOI] [PubMed] [Google Scholar]
- Desmurget M, Prablanc C, Arzi M, Rossetti Y, Paulignan Y, Urquizar C. Integrated control of hand transport and orientation during prehension movements. Experimental Brain Research. 1996;110(2):265–278. doi: 10.1007/BF00228557. [DOI] [PubMed] [Google Scholar]
- Domkin D, Laczko J, Djupsjobacka M, Jaric S, Latash ML. Joint angle variability in 3D bimanual pointing: uncontrolled manifold analysis. Experimental Brain Research. 2005;163(1):44–57. doi: 10.1007/s00221-004-2137-1. [DOI] [PubMed] [Google Scholar]
- Fan J, He J. Motor cortical encoding of hand orientation in a 3-D reach-to-grasp task. Conference Proceedings: 28th Annual International Conference of the IEEE Engineering in Medicine and Biology Society. 2006;1:5472–5475. doi: 10.1109/IEMBS.2006.259860. [DOI] [PubMed] [Google Scholar]
- Fan J, He J, Tillery SI. Control of hand orientation and arm movement during reach and grasp. Experimental Brain Research. 2006;171(3):283–296. doi: 10.1007/s00221-005-0277-6. [DOI] [PubMed] [Google Scholar]
- Gelfand IM, Latash ML. On the problem of adequate language in motor control. Motor Control. 1998;2(4):306–313. doi: 10.1123/mcj.2.4.306. [DOI] [PubMed] [Google Scholar]
- Goodman SR, Shim JK, Zatsiorsky VM, Latash ML. Motor variability within a multi-effector system: experimental and analytical studies of multi-finger production of quick force pulses. Experimental Brain Research. 2005;163(1):75–85. doi: 10.1007/s00221-004-2147-z. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gorniak SL, Duarte M, Latash ML. Do synergies improve accuracy? A study of speed-accuracy trade-offs during finger force production. Motor Control. 2008;12(2):151–172. doi: 10.1123/mcj.12.2.151. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gosselin-Kessiby N, Messier J, Kalaska JF. Evidence for automatic on-line adjustments of hand orientation during natural reaching movements to stationary targets. Journal of Neurophysiology. 2008;99(4):1653–1671. doi: 10.1152/jn.00980.2007. [DOI] [PubMed] [Google Scholar]
- Grea H, Desmurget M, Prablanc C. Postural invariance in three-dimensional reaching and grasping movements. Experimental Brain Research. 2000;134(2):155–162. doi: 10.1007/s002210000427. [DOI] [PubMed] [Google Scholar]
- Gutman SR, Latash ML, Almeida GL, Gottlieb GL. Kinematic Description of Variability of Fast Movements - Analytical and Experimental Approaches. Biological Cybernetics. 1993;69(56):485–492. [PubMed] [Google Scholar]
- Hamilton AF, Wolpert DM. Controlling the statistics of action: obstacle avoidance. J Neurophysiol. 2002;87(5):2434–2440. doi: 10.1152/jn.2002.87.5.2434. [DOI] [PubMed] [Google Scholar]
- Harris CM, Wolpert DM. Signal-dependent noise determines motor planning. Nature. 1998;394(6695):780–784. doi: 10.1038/29528. [DOI] [PubMed] [Google Scholar]
- Hsu WL, Scholz JP, Schoner G, Jeka JJ, Kiemel T. Control and estimation of posture during quiet stance depends on multijoint coordination. Journal of Neurophysiology. 2007;97(4):3024–3035. doi: 10.1152/jn.01142.2006. [DOI] [PubMed] [Google Scholar]
- Jeannerod M. Intersegmental coordination during reaching at natural visual objects. In: Long J, Baddely A, editors. Attention and Performance IX. Hillsdale: Lawrence Erlbaum; 1981. pp. 153–168. [Google Scholar]
- Jeannerod M. Visuomotor channels: Their integration in goal-directed prehension. Human Movement Science. 1999;18(23):201–218. [Google Scholar]
- Kamper DG, Zev Rymer W. Effects of geometric joint constraints on the selection of final arm posture during reaching: a simulation study. Experimental Brain Research. 1999;126(1):134–138. doi: 10.1007/s002210050723. [DOI] [PubMed] [Google Scholar]
- Koshland GF, Hasan Z. Selection of muscles for initiation of planar, three-joint arm movements with different final orientations of the hand. Experimental Brain Research. 1994;98(1):157–162. doi: 10.1007/BF00229121. [DOI] [PubMed] [Google Scholar]
- Krishnamoorthy V, Latash ML, Scholz JP, Zatsiorsky VM. Muscle synergies during shifts of the center of pressure by standing persons. Experimental Brain Research. 2003;152(3):281–292. doi: 10.1007/s00221-003-1574-6. [DOI] [PubMed] [Google Scholar]
- Krishnamoorthy V, Scholz JP, Latash ML. The use of flexible arm muscle synergies to perform an isometric stabilization task. Journal of Clinical Neurophysiology. 2007;118(3):525–537. doi: 10.1016/j.clinph.2006.11.014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Krishnamoorthy V, Yang JF, Scholz JP. Joint coordination during quiet stance: effects of vision. Experimental Brain Research. 2005;164(1):1–17. doi: 10.1007/s00221-004-2205-6. [DOI] [PubMed] [Google Scholar]
- Latash ML, Li S, Danion F, Zatsiorsky VM. Central mechanisms of finger interaction during one- and two-hand force production at distal and proximal phalanges. Brain Research. 2002;924(2):198–208. doi: 10.1016/s0006-8993(01)03234-6. [DOI] [PubMed] [Google Scholar]
- Latash ML, Scholz JP, Danion F, Schoner G. Structure of motor variability in marginally redundant multifinger force production tasks. Experimental Brain Research. 2001;141(2):153–165. doi: 10.1007/s002210100861. [DOI] [PubMed] [Google Scholar]
- Latash ML, Scholz JP, Danion F, Schoner G. Finger coordination during discrete and oscillatory force production tasks. Experimental Brain Research. 2002;146(4):419–432. doi: 10.1007/s00221-002-1196-4. [DOI] [PubMed] [Google Scholar]
- Latash ML, Scholz JP, Schoner G. Motor control strategies revealed in the structure of motor variability. Exercise and Sport Science Reviews. 2002;30(1):26–31. doi: 10.1097/00003677-200201000-00006. [DOI] [PubMed] [Google Scholar]
- Latash ML, Scholz JP, Schoner G. Toward a new theory of motor synergies. Motor Control. 2007;11(3):276–308. doi: 10.1123/mcj.11.3.276. [DOI] [PubMed] [Google Scholar]
- Li ZM, Latash ML, Zatsiorsky VM. Force sharing among fingers as a model of the redundancy problem. Experimental Brain Research. 1998;119(3):276–286. doi: 10.1007/s002210050343. [DOI] [PubMed] [Google Scholar]
- Ma-Wyatt A, McKee SP. Visual Information throughout a reach determines endpoint precision. Experimental Brain Research. 2007;179:55–56. doi: 10.1007/s00221-006-0767-1. [DOI] [PubMed] [Google Scholar]
- Mamassian P. Prehension of objects oriented in three-dimensional space. Experimental Brain Research. 1997;114(2):235–245. doi: 10.1007/pl00005632. [DOI] [PubMed] [Google Scholar]
- Marotta JJ, Medendorp WP, Crawford JD. Kinematic rules for upper and lower arm contributions to grasp orientation. J Neurophysiol. 2003;90(6):3816–3827. doi: 10.1152/jn.00418.2003. [DOI] [PubMed] [Google Scholar]
- Mitra S, Turvey MT. A rotation invariant in 3-D reaching. J Exp Psychol Hum Percept Perform. 2004;30(1):163–179. doi: 10.1037/0096-1523.30.1.163. [DOI] [PubMed] [Google Scholar]
- Murray RM, LI Z, Sastry SS. A mathematical introduction to robotic manipulation. CRC Press; Boca Raton, FL: 1994. [Google Scholar]
- Olafsdottir H, Zhang W, Zatsiorsky VM, Latash ML. Age-related changes in multifinger synergies in accurate moment of force production tasks. Journal of Applied Physiology. 2007;102(4):1490–1501. doi: 10.1152/japplphysiol.00966.2006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Oldfield RC. The assessment and analysis of handedness: the Edinburgh inventory. Neuropsychologia. 1971;9(1):97–113. doi: 10.1016/0028-3932(71)90067-4. [DOI] [PubMed] [Google Scholar]
- Rosenbaum DA, Meulenbroek RG, Vaughan J, Jansen C. Coordination of reaching and grasping by capitalizing on obstacle avoidance and other constraints. Experimental Brain Research. 1999;128(12):92–100. doi: 10.1007/s002210050823. [DOI] [PubMed] [Google Scholar]
- Scholz JP, Danion F, Latash ML, Schoner G. Understanding finger coordination through analysis of the structure of force variability. Biological Cybernetics. 2002;86(1):29–39. doi: 10.1007/s004220100279. [DOI] [PubMed] [Google Scholar]
- Scholz JP, Kang N, Patterson D, Latash ML. Uncontrolled manifold analysis of single trials during multi-finger force production by persons with and without Down syndrome. Experimental Brain Research. 2003;153(1):45–58. doi: 10.1007/s00221-003-1580-8. [DOI] [PubMed] [Google Scholar]
- Scholz JP, Kubo M. Implications of research on motor redundancy for rehabilitation of neurological patients. Journal of the Japanese Physical Therapy Association 2008 [Google Scholar]
- Scholz JP, Reisman D, Schoner G. Effects of varying task constraints on solutions to joint coordination in a sit-to-stand task. Experimental Brain Research. 2001;141(4):485–500. doi: 10.1007/s002210100878. [DOI] [PubMed] [Google Scholar]
- Scholz JP, Schoner G. The uncontrolled manifold concept: identifying control variables for a functional task. Experimental Brain Research. 1999;126(3):289–306. doi: 10.1007/s002210050738. [DOI] [PubMed] [Google Scholar]
- Scholz JP, Schoner G, Latash ML. Identifying the control structure of multijoint coordination during pistol shooting. Experimental Brain Research. 2000;135(3):382–404. doi: 10.1007/s002210000540. [DOI] [PubMed] [Google Scholar]
- Schöner G. Recent developments and problems in human movement science and their conceptual implications. Ecological Psychology. 1995;7:291–214. [Google Scholar]
- Schoner G, Scholz JP. Analyzing variance in multi-degree-of-freedom movements: uncovering structure versus extracting correlations. Motor Control. 2007;11(3):259–275. doi: 10.1123/mcj.11.3.259. [DOI] [PubMed] [Google Scholar]
- Shapkova EY, Shapkova AL, Goodman SR, Zatsiorsky VM, Latash ML. Do synergies decrease force variability? A study of single-finger and multi-finger force production. Experimental Brain Research. 2008;188(3):411–425. doi: 10.1007/s00221-008-1371-3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Shim JK, Latash ML, Zatsiorsky VM. Prehension synergies: trial-to-trial variability and hierarchical organization of stable performance. Experimental Brain Research. 2003;152(2):173–184. doi: 10.1007/s00221-003-1527-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Shim JK, Latash ML, Zatsiorsky VM. Prehension synergies in three dimensions. Journal of Neurophysiology. 2005;93(2):766–776. doi: 10.1152/jn.00764.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Shinohara M, Scholz JP, Zatsiorsky VM, Latash ML. Finger interaction during accurate multi-finger force production tasks in young and elderly persons. Experimental Brain Research. 2004;156(3):282–292. doi: 10.1007/s00221-003-1786-9. [DOI] [PubMed] [Google Scholar]
- Soderkvist I, Wedin PA. Determining the movements of the skeleton using well-configured markers. Journal of Biomechanics. 1993;26(12):1473–1477. doi: 10.1016/0021-9290(93)90098-y. [DOI] [PubMed] [Google Scholar]
- Soechting JF, Flanders M. Parallel, interdependent channels for location and orientation in sensorimotor transformations for reaching and grasping. Journal of Neurophysiology. 1993;70(3):1137–1150. doi: 10.1152/jn.1993.70.3.1137. [DOI] [PubMed] [Google Scholar]
- Stark E, Asher I, Abeles M. Encoding of reach and grasp by single neurons in premotor cortex is independent of recording site. Journal of Neurophysiology. 2007;97(5):3351–3364. doi: 10.1152/jn.01328.2006. [DOI] [PubMed] [Google Scholar]
- Tseng Y, Scholz JP, Schoner G. Goal-equivalent joint coordination in pointing: affect of vision and arm dominance. Motor Control. 2002;6(2):183–207. doi: 10.1123/mcj.6.2.183. [DOI] [PubMed] [Google Scholar]
- Tseng YW, Scholz JP. The effect of workspace on the use of motor abundance. Motor Control. 2005a;9(1):75–100. doi: 10.1123/mcj.9.1.75. [DOI] [PubMed] [Google Scholar]
- Tseng YW, Scholz JP. Unilateral vs. bilateral coordination of circle-drawing tasks. Acta Psychologica (Amst) 2005b;120(2):172–198. doi: 10.1016/j.actpsy.2005.04.001. [DOI] [PubMed] [Google Scholar]
- Tseng YW, Scholz JP, Schoner G, Hotchkiss L. Effect of accuracy constraint on joint coordination during pointing movements. Experimental Brain Research. 2003;149(3):276–288. doi: 10.1007/s00221-002-1357-5. [DOI] [PubMed] [Google Scholar]
- Tseng YW, Scholz JP, Valere M. Effects of movement frequency and joint kinetics on the joint coordination underlying bimanual circle drawing. Journal of Motor Behavior. 2006;38(5):383–404. doi: 10.3200/JMBR.38.5.383-404. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wang X. Three-dimensional kinematic analysis of influence of hand orientation and joint limits on the control of arm postures and movements. Biological Cybernetics. 1999;80:449–463. doi: 10.1007/s004220050538. [DOI] [PubMed] [Google Scholar]
- Zatsiorsky VM, Latash ML, Gao F, Shim JK. The principle of superposition in human prehension. Robotica. 2004;22:231–234. doi: 10.1017/S0263574703005344. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhang W, Scholz JP, Zatsiorsky VM, Latash ML. What do synergies do? Effects of secondary constraints on multidigit synergies in accurate force-production tasks. Journal of Neurophysiology. 2008;99(2):500–513. doi: 10.1152/jn.01029.2007. [DOI] [PMC free article] [PubMed] [Google Scholar]