

Abstract
Flexor tendon injuries are often encountered clinically and typically require surgical repair. Return of function after repair is limited due to adhesion formation, which leads to reduced tendon gliding, and due to a lack of repair site strength, which leads to repair site gap formation or rupture. The application of the growth factors basic fibroblastic growth factor (bFGF) and platelet derived growth factor BB (PDGF-BB) has been shown to have the potential to enhance tendon healing. The objectives of this study were to examine: (1) the conditions over which delivery of bFGF can be controlled from a heparin-binding delivery system (HBDS) and (2) the effect of bFGF and PDGF-BB released from this system on tendon fibroblast proliferation and matrix gene expression in vitro over a 10-day interval. Delivery of bFGF was controlled using a HBDS. Fibrin matrices containing the HBDS retained bFGF better than did matrices lacking the delivery system over the 10-day period studied. Delivery of bFGF and PDGF-BB using the HBDS stimulated tendon fibroblast proliferation and promoted changes in the expression of matrix genes related to tendon gliding, strength, and remodeling. Both growth factors may be effective in enhancing tendon healing in vivo.
Keywords: Growth factor, Tissue engineering, Fibrin
INTRODUCTION
One-third of all acute injuries in workers are to the upper extremity, many of which are open hand wounds requiring flexor tendon repair.8,15,17 While advances in treatment over the past three decades have resulted in steady improvements in clinical outcomes, a large percentage of tendon injuries are debilitating.13 Experimental studies have shown that return of function following flexor tendon repair relies on two factors: (1) smooth gliding of the tendon within its sheath5,39 and (2) the development of repair site strength.5,10 Effective gliding after operative repair depends on the prevention of adhesions between the tendon surface and its sheath and on the production of lubricating molecules at the tendon surface. Development of repair site strength depends on the production and remodeling of collagen at the healing tendon–tendon interface. While modifications in rehabilitation variables have led to improvements in tendon gliding, clinical, and experimental outcomes remain substantially below normal in terms of digital range of motion and repair site strength. As healing tendons are at greatest risk for rupture in the first few weeks after surgical repair, recent treatment approaches have focused on accelerating the early repair process by manipulating the biologic environment of the repair.11,14,19,33
Growth factors have been shown to be powerful regulators of biological function and their presence in tendons is highly regulated in both time and space.19 It is hypothesized that controlled delivery of growth factors may enhance tendon fibroblast proliferation and extracellular matrix synthesis. An accelerated repair process could improve gliding and strength and could substantially reduce the risk of tendon rupture. While numerous growth factors have shown promise for enhancing tendon and ligament healing (e.g., IGF-1, TGF-β), we chose to study platelet derived growth factor BB (PDGF-BB) and basic fibroblast growth factor (bFGF). It has been reported previously that controlled release of PDGF-BB from fibrin matrices using a heparin-binding delivery system (HBDS) promoted cell proliferation and extracellular matrix remodeling.23,33 Additionally, it has been demonstrated that PDGF-BB delivery after tendon repair in vivo promotes improved tendon gliding.11 However, PDGF-BB delivery did not improve tendon strength, and the mechanism for improved tendon gliding was unclear. In order to explain the observed in vivo effects of PDGF-BB, the current study examines the effect of PDGF-BB on fibroblast matrix gene expression in vitro. Furthermore, the potential of an additional growth factor, bFGF, on intrasynovial tendon repair was examined. This growth factor has been shown to promote fibroblast proliferation and extracellular matrix synthesis in vitro and in vivo.7,19,30,32
The objectives of this study were to examine: (1) the range of conditions (e.g., dosage, delivery system formulation, presence of cells) over which delivery of bFGF can be controlled from a HBDS and (2) the effect of bFGF and PDGF-BB released from this system on tendon fibroblast proliferation and matrix gene expression in vitro. It was hypothesized that the release of bFGF could be controlled using a HBDS and that controlled release of bFGF and PDGF-BB would promote increased cell proliferation and extracellular matrix synthesis.
METHODS
Overview
In order to examine the potential of PDGF-BB and bFGF for enhancing flexor tendon repair, the suitability of a HBDS for controlled growth factor release was evaluated in vitro. The effect of each growth factor on cell proliferation and the expression of extracellular matrix related genes by primary tendon fibroblasts was examined. The expression of extracellular matrix related genes important for tendon gliding (lubricin and hyaluronic acid synthase),27,28 tendon strength (collagen I, collagen III, and decorin),36 and tendon remodeling (matrix metalloproteinase 1 and 13)18,37 was then measured (Table 1).
TABLE 1.
Extracellular matrix genes examined with qPCR (canine primers for lubricin are indicated below the table; all other canine primers were obtained from Invitrogen, CA).
| Category | Gene |
|---|---|
| Tendon gliding | Lubricin, hyaluronan synthase 2 (HAS2) |
| Tendon strength | Collagen I (COL I), collagen III (COL III), decorin |
| Tendon remodeling | Matrix metalloproteinase 1 (MMP-1) and 13 (MMP-13) |
Growth Factor Delivery System
Sustained delivery of PDGF and bFGF was achieved using a growth factor delivery system as described previously.24 The delivery system can be designed to administer growth factors in a manner that is tailored specifically to the temporal progression of tissue regeneration. Key elements of the system include a bi-domain peptide with a Factor XIIIa substrate derived from the N-terminus of α2-plasmin inhibitor, and a C-terminal heparin-binding domain derived from anti-thrombin III. The bi-domain peptide is covalently crosslinked to a fibrin matrix during coagulation by the transglutaminase activity of Factor XIIIa; the peptide immobilizes heparin non-covalently to the matrix which, in turn, immobilizes heparinbinding growth factor, preventing its diffusion from the matrix. Release of growth factor from the matrix may occur via three mechanisms: (1) Passive release by dissociation of growth factor from matrix-bound heparin and subsequent diffusion; (2) Active release by proteolytic degradation of the fibrin matrix; or (3) Active release by enzymatic degradation of heparin. The first mechanism is passive and occurs in the presence or absence of cells. With the latter two methods, macrophages and fibroblasts infiltrate and remodel the matrix by releasing the enzymes heparinase and plasmin.
All reagents and chemicals were purchased from Sigma–Aldrich (Saint Louis, MO) unless otherwise indicated. In vitro experiments were performed with PDGF-BB and bFGF (R&D Systems, Minneapolis, MN); and varying concentrations of the delivery system component heparin. Fibrin matrices comprising of 10 mg/mL fibrinogen (EMD Chemicals Inc., San Diego, CA), 6.9 mM CaCl2, and 12.5 units/mL thrombin in Tris-buffered saline (TBS, 137 mM NaCl, 2.7 mM KCl, 33 mM Tris, pH 7.4) were made. In one group, heparin was not added to serve as the “no delivery system” group (growth factor to heparin ratio of 1:0). In other groups, the concentration of heparin was varied to yield HBDS groups with growth factor to heparin molar ratios of 1:100, 1:1000, or 1:10000 respectively. ATIII peptide (dLNQEQVSPK(βA)FAKLAARLYR KA-NH2, where dL denotes dansyl leucine) was used at concentrations corresponding to the different amounts of heparin to maintain a constant molar ratio of 1:10 for heparin to peptide. Passive release was studied for all HBDS groups. Cell-mediated release was studied for the 1:0 and 1:1000 groups. Passive release and cell-mediated release have been demonstrated for PDGF in previous studies.23,33 For cell proliferation and gene expression studies using bFGF and PDGF, only the 1:0 and 1:1000 groups were evaluated.
Cell Isolation
Canine tendon fibroblasts were used for all cell-based experiments to allow comparison with the most clinically relevant animal model of flexor tendon repair.11,22,33 Fibroblasts were isolated from flexor digitorum profundus (FDP) tendons as described previously.23,33 FDP tendons were isolated and bathed in wash media containing 10× penicillin/streptomycin and 10× Fungizone in DMEM. The tendons were cut into 1 mm square sections and a collagenase solution (4500 Units/mL collagenase in DMEM) was added. This solution was incubated at 37 °C for 1 h. Trypsin/EDTA was then added and the solution was incubated at 37 °C for an additional 15 min. Culture medium containing DMEM, 10% fetal bovine serum (FBS, Hyclone Laboratories, Logan, UT), 1× Penicillin/ Streptomycin, 1% non-essential amino acids, and 1% sodium pyruvate (Tissue Culture Support Center, Washington University, Saint Louis, MO) in DMEM was then added and the solution was spun for 10 min at 1200×g. The supernatant was removed, more media was added, and the solution was spun again. The isolated cells and the remaining tissue were incubated overnight (37 °C, 5% CO2, and 100% humidity) in DMEM containing 10% FBS with additional antibiotics (5× Pen/Strep, 1× Fungizone). The following day, the floating tissue was collected and spun down to liberate more cells. The cell and tissue pellet was plated in a 75-mm tissue culture flask in culture medium. The next day, the media was changed and the floating tissue was discarded. The media was changed three times a week until the cells became confluent. The cells were used between passages two and three for experiments.
bFGF Release Kinetics
Passive Release
Fibrin matrices were made as described previously.11,23,24,33 Only the release kinetics of bFGF were examined in the current study as PDGF-BB results are reported elsewhere.23 Briefly, fibrinogen was dissolved for 4–6 h in a 30 mm Petri dish and then dialyzed overnight against 4 L of Tris-buffered saline (TBS) using 6000–8000 Daltons molecular weight cut off dialysis tubing (Fisher Scientific, Pittsburg, PA). The fibrinogen solution was filtered and diluted with TBS to obtain a stock solution of 20 mg/mL. Fibrin matrices were made with 0.25 μg/mL bFGF in 400 μL volume in 24-well tissue culture plates. The wells were pre-coated with bovine serum albumin (BSA) to prevent non-specific protein adsorption. The matrices were allowed to polymerize for 1 h at 37 °C, 100% humidity. A 400 μL volume wash of Tris buffered saline plus 2% BSA (TBS-2% BSA) was then added to each fibrin matrix. The entire 400 μL wash was collected and replaced five times over the first 24 h, and then once per day for 10 days. The wash volumes were stored at −80 °C. At the end of 10 days, the matrices were cut into fragments and placed in 4 mL of extraction buffer containing 10 mg/mL heparin, 1% BSA, 2 M sodium chloride, and 0.01% Triton-X in phosphate buffered saline. The fragmented matrices were incubated in the extraction buffer for 72 h at 4 °C with constant shaking. After 72 h, the extraction buffer was collected and stored at −80 °C. A sandwich ELISA for bFGF (R&D systems Minneapolis, MN) was performed on the wash samples and endpoint extract volumes following the manufacturer’s instructions. Changes between groups and over time were tested using a repeated measures analysis of variance (ANOVA) followed by a Fisher’s Least Squares Differences post-hoc test (significance level set at p < 0.05). The total bFGF was determined to be sum of the bFGF in the wash solutions and that remaining in the fibrin after 10 days. Percent of cumulative release was the amount of bFGF in all the washes up to that time point added together and divided by the total bFGF measured.
Active Release
Fibrin matrices were made as described above. The matrices were washed five times with TBS-2% BSA in the first 24 h and wash volumes were collected. The next day, canine tendon fibroblasts were seeded on top of the fibrin matrices at a concentration of 50,000 cells/well in 400 μL culture medium containing DMEM, 1% FBS, 1× L-Glutamine, 1× Penicillin/Streptomycin, 1% non-essential amino acids, and 1% sodium pyruvate. Seeding cells on the surface of the gel recreates the scenario seen in vivo when the fibrin gel is applied to a repaired flexor tendon. Four cell isolations from four different animals were used. Each of the isolations was plated three times (i.e., “in triplicate”), resulting in 12 sets of data from 4 cell isolations (N = 4 cell isolations in triplicate). Media was changed and collected daily for 10 days. The media samples were collected and stored at 80 °C. After 10 days, the remaining fibrin matrices were fragmented and treated as described above to collect extract volumes. A sandwich ELISA for bFGF was performed on the TBS-BSA washes, media samples, and extract volumes to measure the bFGF released at each day from the HBDS and no HBDS groups by cell-mediated degradation of the fibrin matrix. The ratio of bFGF released from HBDS to no HBDS at each day was then calculated to characterize the cell-mediated release profile of the growth factor from the fibrin matrix. A one-way ANOVA with planned polynomial contrast was used to compare bFGF ratios over time. Paired t-tests were used to compare release of bFGF from HBDS to no HBDS at each time point.
Biologic Assays
Cell Proliferation
The biologic effects of PDGF and bFGF on tendon fibroblasts were investigated at two different time points (5 and 10 days) using a cell-proliferation assay. Three doses (0.125, 0.25, and 1.25 μg/mL) of bFGF or PDGF were used in each cell-seeded fibrin matrix without or with the HBDS. Fibrin matrices (400 μL volume) were made and seeded with tendon fibroblasts as described above. Four experiments were performed with different cell isolations (N = 4 cell isolations in triplicate). Media was changed daily and discarded. At the end of 5 and 10 days, media was removed and 200 μL of 1 U/mL plasminogen was added on top of each matrix to dissolve the fibrin. The matrices were placed back in the incubator. After 2 h, the dissolved matrix solution from each well was collected in a 1.5 mL tube. The wells were trypsinized with 200 μL of 0.05–0.02 M trypsin–EDTA to remove any adherent cells and the solution was added to the tubes as well. The tubes were centrifuged at 12,353 g for 5 min to spin down the cells. The supernatant was removed and the tubes were stored at −80 °C. Cell number was quantified in a 96-well plate by a fluorescence-based DNA assay (CyQUANT kit, Invitrogen Corporation, Carlsbad, CA) according to manufacturer’s instructions. The assay depends on binding of a fluorescent dye to DNA. The level of fluorescence is proportional to the number of cells in the sample and has a linear detection range of 50 to 200,000 cells in a 200 μL volume. The cell pellets were thawed at room temperature and 200 μL of cell lysis buffer containing CyQUANT GR dye was added to each tube. The samples were then transferred to a 96-well plate and fluorescence was measured on a SpectraMax M2 microplate reader at 480 nm excitation and 520 nm emission wavelengths. A standard curve was made ranging from 0 to 250,000 cells. Cell number was calculated from the fluorescence readings using the standard curve. Paired t-tests were used to compare cell proliferation of growth factor-treated groups to control and between release groups (i.e., no HBDS vs. HBDS). A multi-factor ANOVA followed by Fisher’s Least Squares Differences post-hoc test was used to compare effect of growth factor and dose on cell proliferation. Significance was set at p < 0.05.
Gene Expression
We determined changes in gene expression by canine tendon fibroblasts treated with either bFGF or PDGF at two doses (0.125 and 1.25 μg/mL) and with or without HBDS using quantitative real time polymerase chain reaction (qPCR). A separate group of cell-seeded matrices was not treated with growth factor and served as the ‘control’ group. Fibrin matrices were made in 400 μL volume and seeded with cells as described earlier. Four experiments were performed with different cell isolations (N = 4 cell isolation in triplicates). Media was changed daily and discarded. At the end of 5 and 10 days, media was removed and 200 μL of 1 U/mL plasminogen was added on top of each matrix to dissolve the fibrin. The matrices were placed back in the incubator. After 2 h, the dissolved matrix solution was collected in a 2 mL tube. The wells were trypsinized with 200 μL of 0.05–0.02 M trypsin–EDTA to remove any adherent cells and the solution was added to the tubes as well. The triplicates were pooled within a group for RNA extraction and qPCR. The tubes were centrifuged at 12,353 g for 5 min to spin down the cells. RNA was extracted immediately and mRNA expression was quantified by qPCR. Gene expression changes were measured for the matrix related genes lubricin, hyaluronic acid synthase 2 (HAS2), type I collagen (COL I), type III collagen (COL III), decorin, and matrix metalloproteinase 1 (MMP1) and 13 (MMP13) (Table 1; note that MMP1 and MMP13 were only examined for the 0.125 μg/mL dose). GAPDH was used as housekeeping gene. Total RNA was isolated using the RNeasy mini kit (Qiagen, CA) following the manufacturer’s protocol. Briefly, the cells were lysed using a lysis buffer and 70% ethanol was added to aid nucleic acid precipitation. The samples were loaded into RNeasy mini spin columns and centrifuged. The nucleic acid phase was bound to the RNeasy silica membrane. DNase treatment was performed using DNase I (Qiagen, CA) following manufacturer’s instructions. RNA was eluted using nuclease-free water and stored at −80 °C. RNA yield was quantified using a NanoDrop spectrophotometer (Thermo Scientific, DE) and quality analysis was performed using gel electrophoresis with the Agilient 2100 Bioanalyzer (Agilient Technologies, CA). 500 ng RNA was reverse transcribed to cDNA using the Thermoscript RT kit (Invitrogen Corporation, CA) following manufacturer’s instructions. All primers for real-time PCR except lubricin, were predesigned (Qiagen, CA). The forward and reverse primers for lubricin were custom designed. Real time PCR reactions were performed using SYBR Green chemistry on a 7300 sequence detection system (Applied Biosystems, CA). Results were expressed as fold change (Experimental/Control) and were calculated using the Delta Delta Ct method. To calculate this, the Ct values of the growth factor group and the control group were first normalized to the housekeeping gene GAPDH. These normalized Ct values were then compared between the growth factor group and the control group to calculate the fold change of one group relative to the other. Paired t-tests were used to compare gene expression of growth factor-treated groups to controls and between release groups (i.e., no HBDS vs. HBDS). A multifactor ANOVA followed by Fisher’s Least Squares Differences post-hoc test was used to compare the effect of growth factor and dose on gene expression. Significance was set at p < 0.05.
RESULTS
bFGF Release Kinetics
Passive release of bFGF from fibrin matrices depended on the molar ratio of bFGF to heparin (Fig. 1). The release rate was significantly slower with increasing concentrations of heparin. After 2 days, 90% of the growth factor was released from gels without heparin (i.e., 1:0 growth factor to heparin ratio). In contrast, 61, 37, and 20%, of the bFGF were released from the 1:100, 1:1000, and 1:10000 gels, respectively. After 10 days, 99, 71, 47, and 22% of the growth factor were released from 1:0, 1:100, 1:1000, and 1:10000 gels, respectively.
FIGURE 1.
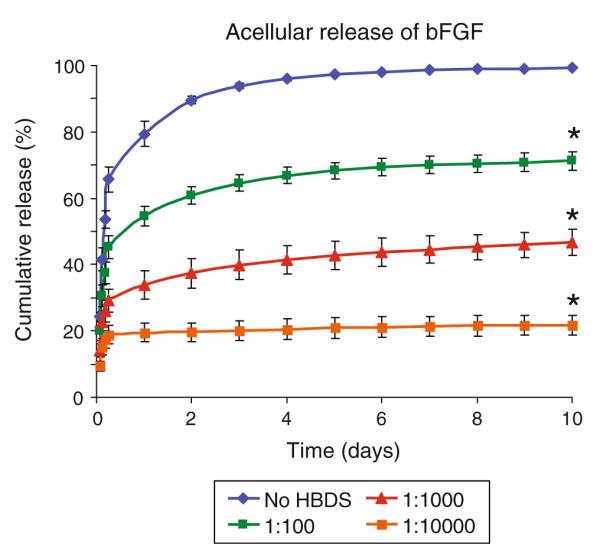
The passive release of bFGF from fibrin matrices depended on the molar ratio of bFGF to heparin. The release rate was significantly slower with increasing concentrations of heparin (*p < 0.05 at day 10 vs. no delivery system).
Active (i.e., cell mediated) release of bFGF depended on the presence of the delivery system (Fig. 2). The amount of growth factor released from the gels in the first day was significantly higher in the no-HBDS group. Approximately twice as much bFGF was released in the first 4.5 h in the no-HBDS group compared to the 1:1000 HBDS group. However, the bFGF fraction released for days 2–10 was significantly higher from the HBDS matrices than from the matrices without HBDS. A statistically significant increasing linear trend was observed over time in the bFGF ratio of HBDS to no HBDS. These results demonstrate that fibrin matrices containing the delivery system were able to retain bFGF better than matrices lacking a delivery system and allow active cell mediated release at later time points.
FIGURE 2.
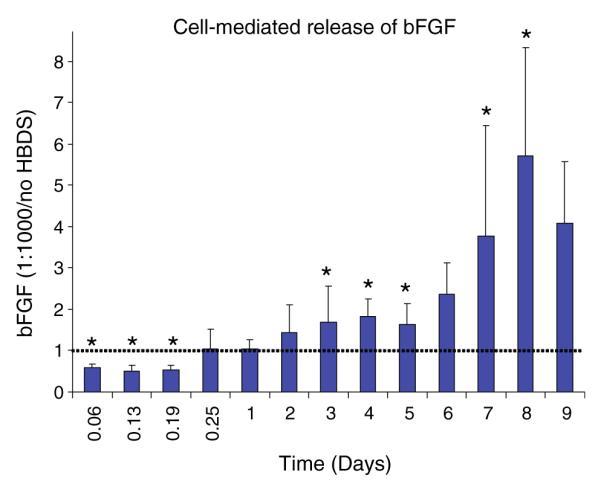
The bFGF fraction released in the first day was higher from the no-HBDS group (i.e., bolus release). However, the bFGF fraction released for days 2–10 was higher from the HBDS matrices than from the matrices without HBDS (*p < 0.05). A statistically significant increasing linear trend was observed over time in the bFGF ratio of HBDS to no HBDS.
Cell Proliferation
Both PDGF-BB and bFGF stimulated cell proliferation at most doses (i.e., compared to control), and cell number was significantly higher in HBDS groups compared to the no-HBDS groups (Fig. 3). PDGF-BB was more effective in stimulating cell proliferation than bFGF at 5 days for the 1.25 μg/mL dose and at 10 days for the 0.125 and 1.25 μg/mL doses. There was no significant effect of growth factor dose and results did not differ between days 5 and 10. These results demonstrate that sustained delivery of PDGF-BB or bFGF can enhance cell proliferation to a greater extent than in unmodified fibrin matrices.
FIGURE 3.

Both PDGF-BB and bFGF stimulated cell proliferation at most doses, and proliferation was significantly higher in HBDS groups vs. no-HBDS groups (symbols above the bars indicate comparisons between 1:1000 and no HBDS, symbols within bars indicate comparisons to control) (paired t-test, *p < 0.05, trends are indicated by #p < 0.1). PDGF-BB was more effective in stimulating cell proliferation than bFGF at 5 days for the 1.25 μg/mL dose and at 10 days for the 0.125 and 1.25 μg/mL doses. There was no effect of dose. Results at days 5 (panel a) and 10 (panel b) were similar.
Gene Expression
Expression of lubricin and HAS2 were upregulated by bFGF at both timepoints (Fig. 4, 0.125 μg/mL dose shown). COL I and COL III were downregulated at 5 days due to PDGF-BB and bFGF and at 10 days due to bFGF (Fig. 5, 0.125 μg/mL dose shown). Expression of decorin was upregulated at 5 days by bFGF (Fig. 5, 0.125 μg/mL dose shown). MMP-1 and MMP-13 were significantly upregulated by bFGF (Fig. 6, 0.125 μg/mL dose shown). There were few significant differences in gene expression when comparing 1:1000 HBDS to no-HBDS. Results at day 5 were similar to day 10. There was no significant effect of growth factor dose (i.e., results for the 0.125 μg/mL dose were similar to those for the 1.25 μg/mL dose, see Supplementary Material).
FIGURE 4.

At the 0.125 μg/mL dose, expression of lubricin and HAS2 were upregulated by bFGF (results are presented as fold changes of growth factor treated relative to control; symbols above the bars indicate comparisons between 1:1000 HBDS and no HBDS, symbols within bars indicate comparisons to control, i.e., a growth factor to control ratio of 1) (paired t-test, *p < 0.05, trends are indicated by #p < 0.1).
FIGURE 5.

At the 0.125 μg/mL dose, COL I and COL III were downregulated at 5 days by PDGF-BB and bFGF and at 10 days by bFGF. Decorin expression was increased by both growth factors, but these changes were only significant at 5 days for the bFGF groups. There were few significant differences in gene expression when comparing HBDS to no-HBDS (results are presented as fold changes of growth factor treated relative to control; symbols above the bars indicate comparisons between 1:1000 HBDS and no HBDS, symbols within bars indicate comparisons to control, i.e., a growth factor to control ratio of 1) (paired t-test, *p < 0.05, trends are indicated by #p < 0.1).
FIGURE 6.

At the 0.125 μg/mL dose, expression of MMP-1 and MMP-13 were upregulated by bFGF (results are presented as fold changes of growth factor treated relative to control; symbols above the bars indicate comparisons between 1:1000 HBDS and no HBDS, symbols within bars indicate comparisons to control, i.e., a growth factor to control ratio of 1) (paired t-test, *p < 0.05, trends are indicated by #p < 0.1).
DISCUSSION
During the first 3 weeks post-tendon repair, fibroblasts migrate into the wound, proliferate, and begin to synthesize collagen type I and III.5 In the months following this initial proliferative phase of healing, proliferation decreases and collagen synthesis and remodeling increase, leading to improvements in repair-site strength. Endogenous healing appears to be limited by the rate of cellular infiltration and the rates of collagen synthesis and remodeling. Therefore, the use of exogenous growth factor delivery in the first 10 days following repair may accelerate the rate of cellular proliferation, extracellular matrix synthesis, and extracellular matrix remodeling and may result in an enhanced repair process.
Growth factors delivered by bolus injection are likely cleared before local fibroblasts appear at a repair site in vivo. Therefore, our long term goal is to provide sustained growth-factor delivery to a surgically repaired tendon. In the current study we demonstrated that bFGF can be administered in a controlled manner in vitro using a heparin-binding delivery system (HBDS). The passive and cell-mediated growth factor release results demonstrate that fibrin matrices containing a HBDS have the capacity to retain bFGF in an improved manner compared to matrices lacking the delivery system. Previously, we showed that PDGF-BB can also be delivered in a controlled manner in vitro using a HBDS.22,33 Subsequent in vivo studies demonstrated a biologic and functional effect of PDGF-BB delivered with this system.11,33 The release of bFGF demonstrated in the current study was slower than the release of PDGF-BB reported previously using the same delivery system. This is likely due to a higher affinity of bFGF for heparin than that of PDGF-BB for heparin. The biologic outcomes in the current study indicate that both bFGF and PDGF-BB may be useful for improving tendon healing.
Operatively, repaired flexor tendon injuries are at greatest risk of rupture in the first few weeks after surgery.6 Acceleration of tendon healing may reduce rupture rates and improve clinical outcomes after repair. Increased fibroblast proliferation during the first few days of tendon healing may lead to increased production of extracellular matrix and remodeling of that matrix at later timepoints. In the current study, we demonstrated that both PDGF-BB and bFGF stimulate cell proliferation at five and 10 days of culture. This is consisted with our previous report showing increased cell proliferation due to PDGF-BB at 6 days.23 In addition, we noted that controlled delivery of the growth factors with a HBDS further enhanced proliferation. Use of these growth factors in vivo may therefore enhance proliferation which may accelerate tendon healing.
Gliding of intrasynovial tendon within its sheath is essential for digital function. The low friction contact that exists in uninjured tissue is primarily determined by the lubricants hyaluronic acid and lubricin. A number of studies have shown improvements in tendon gliding with exogenous application of hyaluronic acid.34 However, other studies have shown no benefit of hyaluronic acid treatment.12 It is likely that the form of hyaluronic acid and how it is attached to the surface of the tendon is of vital importance to its effectiveness. More recently, lubricin was identified on the surface of flexor tendons and at the interface of collagen fiber bundles within the tendon.26 This protein likely plays important roles in the lubrication of intrasynovial tendon and in the inhibition of adhesion formation. We demonstrated a significant upregulation in lubricin and hyaluronic acid synthase with bFGF treatment. It follows that bFGF treatment may have potential with regard to restoring the gliding surface of flexor tendon following repair through the production of lubricating molecules such as lubricin and hyaluronic acid.
PDGF-BB and bFGF treatment led to a significant downregulation of collagens I and II and a significant upregulation in the collagen degradation enzymes matrix metalloproteinase (MMP) 1 and 13. Decorin, a glycoprotein that limits collagen fibrillogenesis,4 was also upregulated due to growth factor treatment, indicating an overall catabolic state with regard to collagenous matrix production. These results are contrary to some reports showing increased extracellular matrix production after treatment with these growth factors.7,31,32,35,38 Our previous study showed increased collagen production at the protein level due to PDGF-BB.23 Therefore, downregulation of collagen gene expression due to PDGF-BB in the current study may not necessarily translate to decreased production of collagen at the protein level and the difference between these two results could be due to post-transcriptional events. Consistent with our bFGF results, however, are a number of recent in vivo and in vitro studies that have shown that bFGF downregulates collagen expression and increases MMP expression in a dose dependent manner.9,20,25,29 Conflicting results in the literature may be due to a lack of consideration for mechanical loading during treatment. Numerous studies have shown that collagen production and MMP expression are highly regulated by mechanical load.1–3 Therefore, bFGF may have a unique effect on tendon fibroblasts that are cyclically loaded compared to those that are not (e.g., during rehabilitation after flexor tendon repair). With regard to PDGF-BB, a recent study demonstrated that the growth factor may not stimulate matrix production directly. Pierce et al.21 showed that PDGF is a potent chemoattractant for wound macrophages and fibroblasts and may stimulate these cells to express endogenous growth factors (e.g., TGF-β) which, in turn, directly stimulate new collagen synthesis.
In order to apply the results of the current in vitro study to the in vivo situation, a number of issues must be considered. First, the active release kinetics of bFGF from the HBDS described in this study may differ from the active release kinetics in vivo. Release could be slower in vivo due to the lack of fibroblasts in the fibrin matrix at the time of implantation or active release could be faster due to additional cell types that enter the wound site along with fibroblasts within the first few days of the surgical repair. However, while the rate of bFGF released in absolute terms may be different in vivo than was seen in the current in vitro study, the nature of the release should be the same. Specifically, growth factor release will depend on passive diffusion of the growth factor from the matrix and on active release of the growth factor through matrix breakdown by local infiltrating cells.
Second, while the critical period for flexor tendon healing is the first 3 weeks after repair, our delivery system was only capable of delivering factors for 10 days, and we only looked at outcome measures at 5 and 10 days. However, enhancing cell proliferation and matrix synthesis in the first 10 days after repair may be sufficient to accelerate the accrual of strength in this early repair period and reduce the risk of rupture. Longer release of certain factors may further enhance remodeling and improve the long term success of the healing tendon.
Third, while the tendon repair environment includes a multitude of cell types, our study only examined the effects of bFGF and PDGF-BB on primary tendon fibroblasts. While these fibroblasts will be the major responding cells at the repaired tendon, other cell types will also be present. For example, bFGF may stimulate angiogenesis by acting on cells brought to the repair site during the inflammatory response.7,16 This increased vascularity may or may not be beneficial to healing; increased vasculature may promote adhesion formation at the tendon surface leading to decreased tendon gliding and/or it may bring additional factors to the repair site that accelerate healing.
Translation of the current study to in vivo use will require consideration of the three issues presented above. In our previous in vivo studies using PDGF-BB in a canine flexor tendon model, we found that biologic enhancement led to improved gliding but not to improved strength. Similarly, others have shown that successful in vitro improvements in gliding through tendon surface treatment do not always translate to improvements in gliding in vivo. In the current study, we demonstrated that the controlled delivery of bFGF could be achieved using an heparin-binding delivery system. Fibrin matrices containing a HBDS were able to retain bFGF better than matrices lacking the delivery system. The controlled delivery of these factors in vivo may allow for sustained biologic stimulation in the early period after repair. The biologic outcomes in the current study indicate that both bFGF and PDGF-BB show promise for improving tendon healing. Delivery of bFGF and PDGF-BB using a HBDS was effective in stimulating tendon fibroblast proliferation. While the HBDS did not have a dramatic effect on gene expression, the growth factors did promote changes in expression of extracellular matrix genes related to tendon gliding, strength, and remodeling. Stimulation of lubricating molecules in particular holds great promise for improving gliding properties in vivo.
Supplementary Material
ACKNOWLEDGMENTS
This study was funded by the National Institutes of Health (AR033097).
Footnotes
ELECTRONIC SUPPLEMENTARY MATERIAL The online version of this article (doi:10.1007/s10439-009-9844-5) contains supplementary material, which is available to authorized users.
REFERENCES
- 1.Arnoczky SP, Lavagnino M, Egerbacher M. The mechanobiological aetiopathogenesis of tendinopathy: is it the over-stimulation or the under-stimulation of tendon cells? Int. J. Exp. Pathol. 2007;88:217–226. doi: 10.1111/j.1365-2613.2007.00548.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Arnoczky SP, Tian T, Lavagnino M, Gardner K. Ex vivo static tensile loading inhibits MMP-1 expression in rat tail tendon cells through a cytoskeletally based mechanotransduction mechanism. J. Orthop. Res. 2004;22:328–333. doi: 10.1016/S0736-0266(03)00185-2. [DOI] [PubMed] [Google Scholar]
- 3.Berry CC, Shelton JC, Bader DL, Lee DA. Influence of external uniaxial cyclic strain on oriented fibroblast-seeded collagen gels. Tissue Eng. 2003;9:613–624. doi: 10.1089/107632703768247313. [DOI] [PubMed] [Google Scholar]
- 4.Birk DE, Nurminskaya MV, Zycband EI. Collagen fibrillogenesis in situ: fibril segments undergo post-depositional modifications resulting in linear and lateral growth during matrix development. Dev. Dyn. 1995;202:229–243. doi: 10.1002/aja.1002020303. [DOI] [PubMed] [Google Scholar]
- 5.Boyer MI, Goldfarb CA, Gelberman RH. Recent progress in flexor tendon healing. The modulation of tendon healing with rehabilitation variables. J. Hand Ther. 2005;18:80–85. doi: 10.1197/j.jht.2005.02.009. (quiz 86) [DOI] [PubMed] [Google Scholar]
- 6.Boyer MI, Strickland JW, Engles D, Sachar K, Leversedge FJ. Flexor tendon repair and rehabilitation: state of the art in 2002. Instr. Course Lect. 2003;52:137–161. [PubMed] [Google Scholar]
- 7.Chan BP, Fu S, Qin L, et al. Effects of basic fibroblast growth factor (bFGF) on early stages of tendon healing: a rat patellar tendon model. Acta Orthop. Scand. 2000;71:513–518. doi: 10.1080/000164700317381234. [DOI] [PubMed] [Google Scholar]
- 8.Feuerstein M, Miller VL, Burrell LM, Berger R. Occupational upper extremity disorders in the federal workforce. Prevalence, health care expenditures, and patterns of work disability. J. Occup. Environ. Med. 1998;40:546–555. doi: 10.1097/00043764-199806000-00007. [DOI] [PubMed] [Google Scholar]
- 9.Fukui N, Katsuragawa Y, Sakai H, Oda H, Nakamura K. Effect of local application of basic fibroblast growth factor on ligament healing in rabbits. Rev. Rhum. Engl. Ed. 1998;65:406–414. [PubMed] [Google Scholar]
- 10.Gelberman RH, Boyer MI, Brodt MD, Winters SC, Silva MJ. The effect of gap formation at the repair site on the strength and excursion of intrasynovial flexor tendons. An experimental study on the early stages of tendon-healing in dogs. J. Bone Joint Surg. Am. 1999;81:975–982. doi: 10.2106/00004623-199907000-00010. [DOI] [PubMed] [Google Scholar]
- 11.Gelberman RH, Thomopoulos S, Sakiyama-Elbert SE, Das R, Silva MJ. The early effects of sustained platelet-derived growth factor administration on the functional and structural properties of repaired intrasynovial flexor tendons: an in vivo biomechanic study at 3 weeks in canines. J. Hand Surg. Am. 2007;32:373–379. doi: 10.1016/j.jhsa.2006.12.009. [DOI] [PubMed] [Google Scholar]
- 12.Hagberg L. Exogenous hyaluronate as an adjunct in the prevention of adhesions after flexor tendon surgery: a controlled clinical trial. J. Hand Surg. Am. 1992;17:132–136. doi: 10.1016/0363-5023(92)90128-c. [DOI] [PubMed] [Google Scholar]
- 13.Kelsey JL. Upper Extremity Disorders: Frequency, Impact and Cost. Churchill Livingstone; New York, NY: 1997. [Google Scholar]
- 14.Khanna A, Friel M, Gougoulias N, Longo UG, Maffulli N. Prevention of adhesions in surgery of the flexor tendons of the hand: what is the evidence? Br. Med. Bull. 2009;90:85–109. doi: 10.1093/bmb/ldp013. [DOI] [PubMed] [Google Scholar]
- 15.Klein MB, Yalamanchi N, Pham H, Longaker MT, Chang J. Flexor tendon healing in vitro: effects of TGF-beta on tendon cell collagen production. J. Hand Surg. Am. 2002;27:615–620. doi: 10.1053/jhsu.2002.34004. [DOI] [PubMed] [Google Scholar]
- 16.Kobayashi D, Kurosaka M, Yoshiya S, Mizuno K. Effect of basic fibroblast growth factor on the healing of defects in the canine anterior cruciate ligament. Knee Surg. Sports Traumatol. Arthrosc. 1997;5:189–194. doi: 10.1007/s001670050049. [see comments] [DOI] [PubMed] [Google Scholar]
- 17.Kreiger N, Kelsey JL, Harris C, Pastides H. Injuries to the upper extremity: patterns of occurrence. Clin. Plast. Surg. 1981;8:13–19. [PubMed] [Google Scholar]
- 18.Lo IK, Marchuk LL, Hollinshead R, Hart DA, Frank CB. Matrix metalloproteinase and tissue inhibitor of matrix metalloproteinase mRNA levels are specifically altered in torn rotator cuff tendons. Am. J. Sports Med. 2004;32:1223–1229. doi: 10.1177/0363546503262200. [DOI] [PubMed] [Google Scholar]
- 19.Molloy T, Wang Y, Murrell G. The roles of growth factors in tendon and ligament healing. Sports Med. 2003;33:381–394. doi: 10.2165/00007256-200333050-00004. [DOI] [PubMed] [Google Scholar]
- 20.Palmon A, Roos H, Edel J, et al. Inverse dose- and time-dependent effect of basic fibroblast growth factor on the gene expression of collagen type I and matrix metalloproteinase-1 by periodontal ligament cells in culture. J. Periodontol. 2000;71:974–980. doi: 10.1902/jop.2000.71.6.974. [DOI] [PubMed] [Google Scholar]
- 21.Pierce GF, Mustoe TA, Lingelbach J, et al. Platelet-derived growth factor and transforming growth factor-beta enhance tissue repair activities by unique mechanisms. J. Cell Biol. 1989;109:429–440. doi: 10.1083/jcb.109.1.429. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Potenza AD. Detailed evaluation of healing processes in canine flexor digital tendons. Mil. Med. 1962;127:34–47. [PubMed] [Google Scholar]
- 23.Sakiyama-Elbert SE, Das R, Gelberman RH, et al. Controlled-release kinetics and biologic activity of platelet-derived growth factor-BB for use in flexor tendon repair. J. Hand. Surg. Am. 2008;33:1548–1557. doi: 10.1016/j.jhsa.2008.05.030. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Sakiyama-Elbert SE, Hubbell JA. Development of fibrin derivatives for controlled release of heparin-binding growth factors. J. Controlled Release. 2000;65:389–402. doi: 10.1016/s0168-3659(99)00221-7. [DOI] [PubMed] [Google Scholar]
- 25.Silverio-Ruiz KG, Martinez AE, Garlet GP, et al. Opposite effects of bFGF and TGF-beta on collagen metabolism by human periodontal ligament fibroblasts. Cytokine. 2007;39:130–137. doi: 10.1016/j.cyto.2007.06.009. [DOI] [PubMed] [Google Scholar]
- 26.Sun Y, Berger EJ, Zhao C, et al. Mapping lubricin in canine musculoskeletal tissues. Connect. Tissue Res. 2006;47:215–221. doi: 10.1080/03008200600846754. [DOI] [PubMed] [Google Scholar]
- 27.Sun Y, Chen MY, Zhao C, An KN, Amadio PC. The effect of hyaluronidase, phospholipase, lipid solvent and trypsin on the lubrication of canine flexor digitorum profundus tendon. J. Orthop. Res. 2008;26:1225–1229. doi: 10.1002/jor.20624. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Taguchi M, Sun YL, Zhao C, et al. Lubricin surface modification improves tendon gliding after tendon repair in a canine model in vitro. J. Orthop. Res. 2009;27:257–263. doi: 10.1002/jor.20731. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Takayama S, Murakami S, Miki Y, et al. Effects of basic fibroblast growth factor on human periodontal ligament cells. J. Periodontal Res. 1997;32:667–675. doi: 10.1111/j.1600-0765.1997.tb00577.x. [DOI] [PubMed] [Google Scholar]
- 30.Tang JB, Cao Y, Zhu B, et al. Adeno-associated virus-2-mediated bFGF gene transfer to digital flexor tendons significantly increases healing strength. An in vivo study. J. Bone Joint Surg. Am. 2008;90:1078–1089. doi: 10.2106/JBJS.F.01188. [DOI] [PubMed] [Google Scholar]
- 31.Tang JB, Xu Y, Ding F, Wang XT. Tendon healing in vitro: promotion of collagen gene expression by bFGF with NF-kappaB gene activation. J. Hand. Surg. Am. 2003;28:215–220. doi: 10.1053/jhsu.2003.50052. [DOI] [PubMed] [Google Scholar]
- 32.Thomopoulos S, Harwood FL, Silva MJ, Amiel D, Gelberman RH. Effect of several growth factors on canine flexor tendon fibroblast proliferation and collagen synthesis in vitro. J. Hand. Surg. Am. 2005;30:441–447. doi: 10.1016/j.jhsa.2004.12.006. [DOI] [PubMed] [Google Scholar]
- 33.Thomopoulos S, Zaegel M, Das R, et al. PDGF-BB released in tendon repair using a novel delivery system promotes cell proliferation and collagen remodeling. J. Orthop. Res. 2007;25:1358–1368. doi: 10.1002/jor.20444. [DOI] [PubMed] [Google Scholar]
- 34.Uchiyama S, Amadio PC, Ishikawa J, An KN. Boundary lubrication between the tendon and the pulley in the finger. J. Bone Joint Surg. Am. 1997;79:213–218. [PubMed] [Google Scholar]
- 35.Wang XT, Liu PY, Xin KQ, Tang JB. Tendon healing in vitro: bFGF gene transfer to tenocytes by adeno-associated viral vectors promotes expression of collagen genes. J. Hand. Surg. Am. 2005;30:1255–1261. doi: 10.1016/j.jhsa.2005.06.001. [DOI] [PubMed] [Google Scholar]
- 36.Woo SL, An K, Frank CB, et al. Anatomy, biology, and biomechanics of tendon and ligament. In: Buckwalter JA, Einhorn TA, Simon SR, editors. Orthopaedic Basic Science. AAOS; Rosemont, IL: 2000. pp. 581–616. [Google Scholar]
- 37.Xu Y, Murrell GA. The basic science of tendinopathy. Clin. Orthop. Relat. Res. 2008;466:1528–1538. doi: 10.1007/s11999-008-0286-4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Yoshikawa Y, Abrahamsson SO. Dose-related cellular effects of platelet-derived growth factor-BB differ in various types of rabbit tendons in vitro. Acta Orthop. Scand. 2001;72:287–292. doi: 10.1080/00016470152846646. [DOI] [PubMed] [Google Scholar]
- 39.Zhao C, Sun YL, Amadio PC, et al. Surface treatment of flexor tendon autografts with carbodiimide-derivatized hyaluronic Acid. An in vivo canine model. J. Bone Joint Surg. Am. 2006;88:2181–2191. doi: 10.2106/JBJS.E.00871. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.