

Abstract
Wild populations of Triatoma infestans, the main vector of Chagas disease in the Southern Cone countries, may be involved in reinfestation of human dwellings, limiting the success of vector-control campaigns in Bolivia. Knowledge of the distribution of these populations remains incomplete. We report here the detection of T. infestans wild populations in large areas in the department of La Paz, Bolivia. Among 18 sylvatic areas investigated, 17 were positive with T. infestans specimens. The infection rate of captured T. infestans with Trypanosoma cruzi was 85.7% in adult specimens. These results expand the geographical distribution of wild populations of T. infestans; it may be distributed throughout the Inter-Andean Dry Forest eco-region of Bolivia. The current information allows us to propose the hypothesis that a sylvatic origin of the reinfestation is located in the valleys of La Paz.
Introduction
Chagas disease is a major public health problem in Latin America. Trypanosoma cruzi (Kinetoplastida: Trypanosomatidae), the causative agent of the disease, is transmitted by contaminated feces of triatomine bugs. In Bolivia and other countries of the Southern Cone in South America, Triatoma infestans (Klug 1834) remains the main vector species.
In the Southern Cone, control of Chagas disease is being pursued by elimination of T. infestans (principally through insecticidal spraying). Control efforts have relied on the assumption that this species was almost exclusively domestic, although a few wild populations had been reported in the Cochabamba valley in Bolivia.1–3 More recently, other wild foci of T. infestans have been described, however, indicating that wild populations of T. infestans have a larger dispersion than previously assumed.4–6 As new species or populations tend to invade the ecological niche released by a first species, wild populations of T. infestans could infest habitats free of domestic triatomine populations and jeopardize vector-control efforts.7,8
We found wild populations of T. infestans in two valleys of La Paz, Bolivia, where transmission of Trypanosoma cruzi persists. New hypotheses about the geographical distribution of wild populations of T. infestans and the epidemiological risk that they represent are suggested.
Materials and methods
Study areas.
In conjunction with the National Program of Chagas disease (PNCH), two valleys under vector control since 2003, Rio La Paz and Rio Luribay, were chosen. Despite insecticidal spraying, locals were still suffering domiciliary reinfestation by T. infestans. In these valleys, 6 and 12 trapping areas between 2,159 and 2,864 m altitude were selected (Figure 1; Table 1). All areas were situated in wild environments, which will be referred as “sylvatic” in this work, at a distance varying from 20 to 500 m from human dwellings. Fields closest to rivers are dedicated to vegetable crops and maize in the two valleys. Surrounding mountains have scarce vegetation dominated by thorny plants and cacti. People also cultivate prickly pear (Opuntia ficus-indica) and the cochineal (Dactylopius coccus) insect used for food coloring and cosmetics. Both valleys are situated in the Inter-Andean Dry Forest eco-region.9 Climate in this area is semi-arid with an average annual temperature of 18°C (range = 8.3–30.8°C), and mean relative humidity is 5.8%. Rainfall occurs between December and April, and winter and spring are very dry (June to November).
Figure 1.
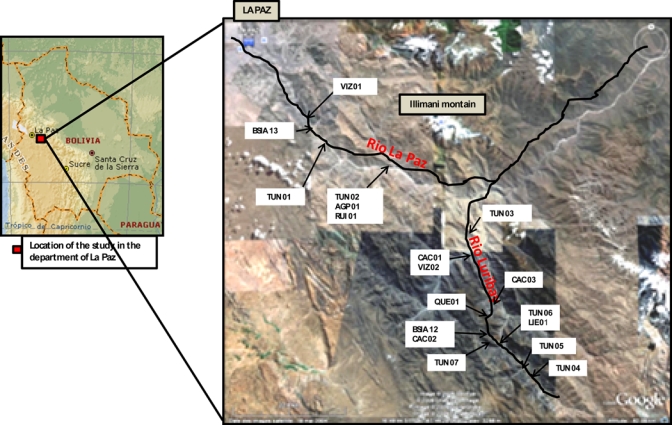
Map of Rio La Paz and Rio Luribay valleys (lines) obtained from Google Earth. All the studied areas are located between VIZ 01 (2,821 m) and RUI 01 (2,459 m) and between TUN 04 (2,767 m) and TUN 03 (2,095 m) in Rio La Paz and Rio Luribay, respectively. This figure appears in color at www.ajtmh.org.
Table 1.
Description, geographical localization, and infestation of the investigated sylvatic areas in two valleys of the department of La Paz, Bolivia
| Valley | Location code | Nearest village | Description of trapping areas (distance from nearest habitat) | Latitude | Longitude | Alt. (m) | Date of trapping | Number of traps placed | Number of positive traps | % of positive traps | Total number of triatomine bugs |
|---|---|---|---|---|---|---|---|---|---|---|---|
| Rio La Paz | VIZ 01 | Aucani | Deep cracks in a small highland that are shelters for vizcachas (Andean small rodent) (200 m). | 16°41'25.47" | 68°00'39.76" | 2,821 | 5/8/2008 | 13 | 2 | 15.4 | 4 |
| 8/20/2008 | 25 | 6 | 24.0 | 22 | |||||||
| BSIA 13 | El Palomar | Wilderness mountainside with sparse vegetation of mainly cacti and thorny vegetation (50–200 m). | 16°42'02.75" | 67°59'54.91" | 2,765 | 9/10/2008 | 48 | 4 | 8.3 | 22 | |
| TUN 01 | Huayhuasi | Field of prickly pears adjacent to some houses outside the village (20–150 m). | 16°42'25.47" | 67°59'37.64" | 2,757 | 5/8/2008 | 30 | 6 | 20.0 | 46 | |
| 7/16/2008 | 49 | 9 | 18.4 | 59 | |||||||
| 8/19/2008 | 30 | 8 | 26.7 | 50 | |||||||
| AGP 01 | Tahuapalca | Edges of footpaths between crop fields around the village (20–200 m). | 16°43'05.03" | 67°52'30.59" | 2,380 | 5/10/2008 | 23 | 1 | 4.3 | 2 | |
| TUN 02 | Tahuapalca | Field of prickly pears overhanging the village (50–150 m). | 16°43'11.53" | 67°52'25.70" | 2,427 | 5/11/2008 | 29 | 12 | 41.4 | 37 | |
| RUI 01 | Tahuapalca | Ruin of a 19th century barracks with mud walls next to a small cemetery on a highland overhanging the village of Tahuapalca (500 m). | 16°42'56.45" | 67°52'13.53" | 2,459 | 5/10/2008 | 40 | 15 | 37.5 | 34 | |
| Total Rio La Paz | 287 | 63 | 22.0 | 276 | |||||||
| Rio Luribay | TUN 04 | Carayapu | Wilderness mountainside with sparse vegetation of mainly cacti and thorny vegetation, which is adjacent to some house of the village (50–300 m). | 17°08'10.80" | 67°35'17.90" | 2,767 | 6/17/2008 | 30 | 6 | 20.0 | 10 |
| TUN 05 | Pucuma | Wilderness mountainside and cemetery outside the village (100–300 m). | 17°07'32.00" | 67°35'59.50" | 2,864 | 6/17/2008 | 20 | 6 | 30.0 | 14 | |
| LIE 01 | Cuty | Rock-pile boundary walls and wilderness field at the entrance of the village (50–100 m). | 17°04'44.60" | 67°37'57.00" | 2,602 | 6/19/2008 | 25 | 12 | 48.0 | 40 | |
| TUN 06 | Catavi | Field of prickly pears adjacent to the church of the village (50–100 m). | 17°04'25.20" | 67°37'59.70" | 2,493 | 6/18/2008 | 20 | 3 | 15.0 | 10 | |
| TUN 07 | Peña Colorada | Field of prickly pears outside the village (50–300 m). | 17°04'24.20" | 67°38'42.70" | 2,543 | 6/18/2008 | 30 | 1 | 3.3 | 2 | |
| CAC 02 | El Calvario | Field of prickly pears adjacent to the cemetery of the village (50–150 m). | 17°04'07.70" | 67°39'25.50" | 2,645 | 6/18/2008 | 25 | 5 | 20.0 | 7 | |
| BSIA 12 | Luribay | Wilderness mountainside with sparse vegetation of mainly cacti and thorny vegetation (50–200 m). | 17°03'34.80" | 67°39'58.40" | 2,583 | 10/31/2008 | 47 | 11 | 23.4 | 44 | |
| QUE 01 | Achocara Bajo | Cliff of red earth along the road, forming small caves, and isolated from any village (500 m of a solitary house). | 17°01'54.80" | 67°40'38.60" | 2,159 | 6/20/2008 | 40 | 5 | 12.5 | 7 | |
| CAC 03 | Palca | Field of prickly pears adjacent to some houses outside the village (20–100 m). | 17°00'30.10" | 67°39'25.20" | 2,356 | 6/20/2008 | 49 | 41 | 83.7 | 263 | |
| VIZ 02 | Lacayani | Large stones at the side of the road (500 m far from an isolated house). | 16°55'48.80" | 67°41'32.90" | 2,182 | 6/19/2008 | 39 | 31 | 79.5 | 284 | |
| CAC 01 | Lacayani | Wilderness valley with sparse vegetation composed mainly of cacti and thorny vegetation (500 m far from isolated houses). | 16°55'33.20" | 67°41'42.60" | 2,210 | 6/18/2008 | 9 | 0 | – | 0 | |
| TUN 03 | Khola | Wilderness site with sparse vegetation of mainly cacti and thorny vegetation (50–500 m). | 16°53'12.20" | 67°42'43.10" | 2,095 | 6/18/2008 | 25 | 7 | 28.0 | 21 | |
| Total Rio Luribay | 359 | 128 | 35.7 | 702 | |||||||
| Total | 646 | 191 | 29.6 | 978 |
Survey of human population.
During April and May 2008, 305 inhabitants from 31 different villages in these valleys were questioned about their knowledge of triatomine bugs (locally called vinchucas) in domestic/sylvatic environments and their localization. They were also asked if they have ever seen vinchucas flying.
Collection of wild triatomines and laboratory processing.
Collection was carried out between May and October 2008 using mice-baited adhesive traps.10 The traps were placed in small caves and cracks, in burrows, or under vegetation. They were set in the afternoon and inspected the next morning. More than 20 traps were placed in most of the trapping areas (Table 1). All insects caught by a trap were placed together in one tube, and they were transported alive to the laboratory. The identification of triatomines (adults and nymphs) was performed according to morphological taxonomic keys.11 Sex and stages were also determined. Feces from each bug were examined for the presence of trypanosomatides by direct microscopical observation at 400× magnification. Bugs were then dissected under a safety hood, and legs and digestive tracks were conserved for further studies.
Results
Knowledge of the triatomine bugs by the surveyed population.
More than 96% of interviewed locals recognized the vector known as vinchuca in the two valleys (Rio La Paz = 117 interviews; Rio Luribay = 188 interviews). Triatomines have been observed in 12% of interviewees' homes within the previous 12 months, and 7% have seen them during the prior month. Moreover, 9% have seen bugs during the year in their peridomicile. In addition, 39.1% of women and 63% of men surveyed claimed to have seen bugs in the sylvatic environment: in the hills, under rocks, in fields of prickly pear, in ruins, and in cemeteries. Finally, 30% of those surveyed reported that they have seen the bugs flying. “They fly from the hill towards the houses, they fly to the roof of the houses or to the window in the night, and we hear them arriving to the tinny roof” were expressions frequently used by the interviewees.
Areas of collection.
Triatomine bugs were found in all trapping areas except one (area CAC 01) in which only nine traps have been placed (Table 1). A total of 646 traps were used during the study, and 29.6% of them caught wild T. infestans specimens. The proportion of positive traps in an area was variable and ranged from 3.3% in TUN 07 to 83.7% in area CAC 03, but only two areas had fewer than 5% positive. The five areas with greatest infestation (> 37.5% of traps were positive; Figure 2A–E) were wilderness areas with thorny vegetation and prickly pear fields with rock-pile boundary walls (areas TUN 02, CAC 03, and LIE 01), a ruin with mud walls providing shelter for small rodents (area RUN 01), and a mountainside composed of large rocks where excrements of vizcachas (rodents belonging to the Chinchillidae family) were observed (area VIZ 02).
Figure 2.

Different sampling areas. (A) Field of prickly pears overhanging the village of Tahuacalpa. (B) Ruin of 19th century barracks with mud walls next to a small cemetery on a highland overhanging the village of Tahuapalca. (C) Rock-pile boundary walls and wilderness field at the entrance of the village of Cuty. (D) Field of prickly pears adjacent to houses outside of the village of Palca. (E) Large stones along the driveway at 500 m from an isolated house. (F) Deep cracks in the ground of a small highland where vizcachas (rodents belonging to the Chinchillidae family) have been observed. This figure appears in color at www.ajtmh.org.
Interestingly, in the positive area VIZ 01 (small highland), traps were placed in deep cracks formed in sediment by erosion (2–5 m) that serve as shelters for vizcachas (Figure 2F).
Triatomine bug collection and infection rate.
A total of 978 T. infestans specimens were collected in 17 positive areas. The number of triatomines by positive trap ranged from 1.4 triatomines per trap in areas QUE 01 and CAC 02 to 9.16 triatomines per trap in area VIZ 02. Adult insects as well as nymphal instars were caught in all areas, and a predominance of young nymphs was observed among the total population (1st + 2nd + 3rd nymphal instars = 70.1%; Figure 3). The proportion of positive traps was significantly higher in Rio Luribay than in Rio La Paz (χ2 P < 0.001), but triatomine density per positive trap was not significantly different (Mann–Whitney–Wilcoxon test, P > 0.05). A total of 448 T. infestans feces were examined, and positive samples for trypanosomatide were found in all the trapping areas (Table 2). The infection rates ranged from 3.8% in first nymphal instar to 85.7% in adult insects with no significant differences between sex (Figure 4; χ2 P > 0.05). Preliminary analysis of feces by polymerase chain reaction (PCR) confirmed infection of T. infestans by T. cruzi species (data not shown).12
Figure 3.
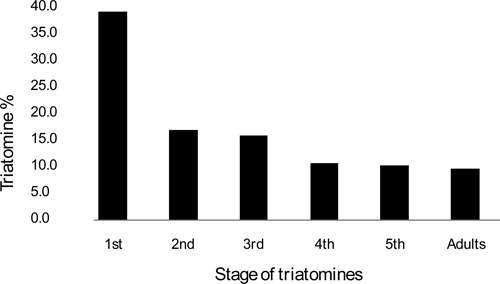
Age structure of wild T. infestans populations caught in the valleys of Rio La Paz and Rio Luribay.
Table 2.
Infection rate of sylvatic T. infestans captured in the 17 positive areas in the department of La Paz
| Area code | Number of adults | % of infected adults | Number of nymphs | % of infected nymphs | % of total infected | ||
|---|---|---|---|---|---|---|---|
| Infected | Not infected | Infected | Not infected | ||||
| VIZ 01 | 3 | 0 | 100.0 | 1 | 2 | 33.3 | 66.6 |
| BSIA 13 | 1 | 0 | 100.0 | 3 | 12 | 20.0 | 25.0 |
| TUN 01 | 7 | 1 | 87.5 | 40 | 36 | 52.6 | 55.9 |
| AGP 01 | 0 | 0 | – | 2 | 0 | 100.0 | 100.0 |
| TUN 02 | 7 | 0 | 100.0 | 11 | 6 | 64.7 | 75.0 |
| RUI 01 | 5 | 0 | 100.0 | 14 | 7 | 66.6 | 73.1 |
| Total Rio La Paz | 23 | 1 | 95.8 | 71 | 63 | 53.0 | 59.5 |
| TUN 04 | 2 | 2 | 50.0 | 0 | 5 | 0.0 | 22.2 |
| TUN 05 | 0 | 0 | – | 2 | 6 | 25.0 | 25.0 |
| LIE 01 | 2 | 1 | 66.6 | 12 | 4 | 75.0 | 73.7 |
| TUN 06 | 2 | 1 | 66.6 | 1 | 2 | 33.3 | 50.0 |
| TUN 07 | 0 | 0 | – | 0 | 2 | 0.0 | 0.0 |
| CAC 02 | 3 | 0 | 100.0 | 1 | 0 | 100.0 | 100.0 |
| BSIA 12 | 8 | 0 | 100.0 | 6 | 6 | 50.0 | 70.0 |
| QUE 01 | 0 | 0 | – | 1 | 1 | 50.0 | 50.0 |
| CAC 03 | 6 | 3 | 66.6 | 47 | 72 | 39.5 | 41.4 |
| VIZ 02 | 2 | 0 | 100.0 | 22 | 51 | 30.1 | 32.0 |
| TUN 03 | 0 | 0 | – | 11 | 6 | 64.7 | 64.7 |
| Total Rio Luribay | 25 | 7 | 78.1 | 103 | 155 | 39.9 | 44.1 |
| Total | 48 | 8 | 85.7 | 174 | 218 | 44.4 | 49.5 |
Figure 4.
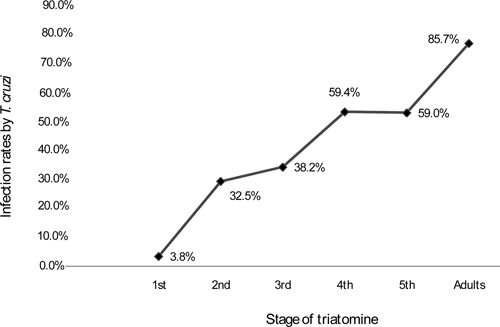
Infection rate of the different stages of wild T. infestans caught in the valleys of Rio La Paz and Rio Luribay.
Discussion
Since the first identification of a wild population of T. infestans in 1946, several reports have described the existence of other wild foci in the department of Cochabamba, Bolivia.1–5 Subsequent reports have suggested a wider distribution of wild T. infestans in Bolivia than just the Cochabamba valley, and findings of wild T. infestans in Argentina (LA Ceballos and others, unpublished data) and Chile have also been reported.8,13
This fieldwork in two La Paz valleys reported here showed a wide distribution of wild populations of T. infestans. The sequences of cytochrome b gene of some insects confirmed the species T. infestans. In both valleys, wild populations were easily captured in places very near the human habitat as well as remote places, and the percentage of positive traps was similar to that reported previously in Cochabamba (~30%).4
T. infestans was found in settings never described before as habitats of this species (thorny hills, fields of prickly pear, screes, rock-pile boundary walls, ruins, and deep cracks formed by erosion). Indeed, the typical ecotope of wild T. infestans has been previously thought to be a rocky outcrop, except for the T. infestans Dark Morph found in the Gran Chaco region.8 These new findings suggest the existence of a continuous distribution of wild populations in the valleys of La Paz. However, the distribution and abundance of these populations are little compatible with secondary wild colonization of the environment by domestic populations. However, the full distribution of T. infestans remains to be more accurately measured. One approach would be to investigate the different eco-regions of Bolivia where domestic T. infestans is endemic. Indeed, the valleys of La Paz as well as the valleys of Cochabamba, where wild populations exist, belong to the Inter-Andean Dry Forest eco-region, but the endemic area of domestic populations of T. infestans includes other eco regions where few data of sylvatic foci are available.
The Bolivian PNCH has expressed interest in whether or not occurrences of reinfestation by T. infestans have a sylvatic origin. In both valleys investigated here, the PNCH reports continuous reinfestation, and this was confirmed by local residents. Moreover, local residents reported seeing T. infestans in the sylvatic environment and flying to dwellings. These data suggested the capacity of the bug to fly from their natural habitats to human-developed areas. Moreover, our work points to the utility of adding interviews with locals to help determine the presence of sylvatic triatomines: information from our interviewees was strongly corroborated in the field. To effectively assess the epidemiological role of wild T. infestans populations, it is necessary to better understand the mechanisms of the reinfestation and to discriminate residual populations, which can develop after insecticide spraying, from reinfestation by sylvatic populations. Therefore, population genetic analyses by different genetic markers should be useful. Also, it is necessary to bring eco-epidemiological research to understand the anthropological, sociological, and biological determinants of reinfestation.14,15
Acknowledgments
We extend our gratitude to the authorities of the Ministerio de Salud y Deporte and Mirko Rojas, head of the National Program of Chagas Control (PNCH), for their help. This study received financial support from the Institut de Recherche pour le Développement (IRD) and the Agence Nationale de la Recherche (ANR) of France.
Footnotes
Authors' addresses: Rosio Buitrago, Etienne Waleckx, Marie-France Bosseno, Faustine Zoveda, Renata Salas, François Noireau, and Simone Frédérique Brenière, Institut de Recherche pour le Développement (IRD), Representación en Bolivia, UR 016 Caractérisation et Contrôle des Populations de Vecteurs, Avenida Hernando Siles N° 5290, Esq. Calle 7, Obrajes, C.P. 9214, La Paz, Bolivia. Pablo Vidaurre and Elio Mamani, Servicio Departamental de Salud La Paz (SEDES La Paz), Calle Capitán Ravelo No. 2180, Zona Central, La Paz, Bolivia.
References
- 1.Torrico RA. Hallazgo de Eratyrus mucronatus, infestación natural de vinchucas de cerro y Eutriatoma sordida en Cochabamba. An Lab Central Cochabamba. 1946;1:19–23. [Google Scholar]
- 2.Dujardin JP, Tibayrenc M, Venegas E, Maldonado L, Desjeux P, Ayala FJ. Isozyme evidence of lack of speciation between wild and domestic Triatoma infestans (Heteroptera, Reduviidae) in Bolivia. J Med Entomol. 1987;24:40–45. doi: 10.1093/jmedent/24.1.40. [DOI] [PubMed] [Google Scholar]
- 3.Bermudez H, Balderrama F, Torrico F.1993Identification and characterization of sylvatic foci of Triatoma infestans in Central Bolivia Am J Trop Med Hyg 49: (Suppl):371 [Google Scholar]
- 4.Noireau F, Flores R, Gutierrez T, Dujardin JP. Detection of sylvatic dark morphs of Triatoma infestans in the Bolivian Chaco. Mem Inst Oswaldo Cruz. 1997;92:583–584. doi: 10.1590/s0074-02761997000500003. [DOI] [PubMed] [Google Scholar]
- 5.Cortez MR, Pinho AP, Cuervo P, Alfaro F, Solano M, Xavier SCC, D'Andrea PS, Fernandes O, Torrico F, Noireau F, Jansen AM. Trypanosoma cruzi (Kinetoplastida Trypanosomatidae): ecology of the transmission cycle in the wild environment of the Andean valley of Cochabamba, Bolivia. Exp Parasitol. 2006;114:305–313. doi: 10.1016/j.exppara.2006.04.010. [DOI] [PubMed] [Google Scholar]
- 6.Cortez MR, Emperaire L, Piccinali RV, Gurtler RE, Torrico F, Jansen AM, Noireau F. Sylvatic Triatoma infestans (Reduviidae, Triatominae) in the Andean valleys of Bolivia. Acta Trop. 2007;102:47–54. doi: 10.1016/j.actatropica.2007.02.014. [DOI] [PubMed] [Google Scholar]
- 7.Krebs CJ.1985Ecology: The Experimental Analysis of Distribution and Abundance New York: Harper and Row, 3rd ed. [Google Scholar]
- 8.Noireau F, Cortez MGR, Monteiro FA, Jansen AM, Torrico F. Can wild Triatoma infestans foci in Bolivia jeopardize Chagas disease control efforts? Trends Parasitol. 2005;21:7–10. doi: 10.1016/j.pt.2004.10.007. [DOI] [PubMed] [Google Scholar]
- 9.Ibisch PL, Merida G. Biodiversidad: La riqueza de Bolivia. Estado de Conocimiento y Conservación. Santa Cruz de la Sierra: Editorial FAN; 2003. [Google Scholar]
- 10.Noireau F, Abad-Franch F, Valente SAS, Dias-Lima A, Lopes CM, Cunha V, Valente VC, Palomeque FS, de Carvalho-Pinto CJ, Sherlock I, Aguilar M, Steindel M, Grisard EC, Jurberg J. Trapping triatominae in silvatic habitats. Mem Inst Oswaldo Cruz. 2002;97:61–63. doi: 10.1590/s0074-02762002000100009. [DOI] [PubMed] [Google Scholar]
- 11.Lent H, Wygodzinsky P. Revision of the triatominae (Hemiptera, Reduviidae), and their significance as vectors of Chaga's disease. Bull Am Mus Nat Hist. 1979;163:123–520. [Google Scholar]
- 12.Bosseno MF, Garcia LS, Baunaure F, Gastelum EM, Gutierrez MS, Kasten FL, Dumonteil E, Brenière SF. Identification in triatomine vectors of feeding sources and Trypanosoma cruzi variants by heteroduplex assay and a multiplex miniexon polymerase chain reaction. Am J Trop Med Hyg. 2006;74:303–305. [PubMed] [Google Scholar]
- 13.Bacigalupo BA, Segura MJA, Garcia CA, Hidalgo CJ, Galuppo GS, Cattan PE. Primer hallazgo de vectores de la enfermedad de Chagas asociados a matorrales silvestres en la Región Metropolitana, Chile. Rev Med Chil. 2006;134:1230–1236. doi: 10.4067/s0034-98872006001000003. [DOI] [PubMed] [Google Scholar]
- 14.Dujardin JP, Cardozo L, Schofield CJ. Genetic analysis of Triatoma infestans following insecticidal controlinterventions in central Bolivia. Acta Trop. 1996;61:263–266. doi: 10.1016/0001-706x(96)00008-3. [DOI] [PubMed] [Google Scholar]
- 15.Cerere MC, Vazquez-Prockopec DGM, Gütler RE, Kitron U. Spatio-temporal analysis of reinfestation by Triatoma infestans (Hemiptera: Reduviidae) following insecticida spraying in a rural community in Northwestern Argentina. Am J Trop Med Hyg. 2004;71:803–810. [PMC free article] [PubMed] [Google Scholar]