

Abstract
Limitation of water loss and control of gas exchange is accomplished in plant leaves via stomatal guard cells. Stomata open in response to light when an increase in guard cell turgor is triggered by ions and water influx across the plasma membrane. Recent evidence demonstrating the existence of ATP-binding cassette proteins in plants led us to analyze the effect of compounds known for their ability to modulate ATP-sensitive potassium channels (K-ATP) in animal cells. By using epidermal strip bioassays and whole-cell patch-clamp experiments with Vicia faba guard cell protoplasts, we describe a pharmacological profile that is specific for the outward K+ channel and very similar to the one described for ATP-sensitive potassium channels in mammalian cells. Tolbutamide and glibenclamide induced stomatal opening in bioassays and in patch-clamp experiments, a specific inhibition of the outward K+ channel by these compounds was observed. Conversely, application of potassium channel openers such as cromakalim or RP49356 triggered stomatal closure. An apparent competition between sulfonylureas and potassium channel openers occurred in bioassays, and outward potassium currents, previously inhibited by glibenclamide, were partially recovered after application of cromakalim. By using an expressed sequence tag clone from an Arabidopsis thaliana homologue of the sulfonylurea receptor, a 7-kb transcript was detected by Northern blot analysis in guard cells and other tissues. Beside the molecular evidence recently obtained for the expression of ATP-binding cassette protein transcripts in plants, these results give pharmacological support to the presence of a sulfonylurea-receptor-like protein in the guard-cell plasma membrane tightly involved in the outward potassium channel regulation during stomatal movements.
Keywords: guard cell, ATP-sensitive potassium channels, outward potassium channels, ATP-binding cassette, stomata
The ATP-binding cassette (ABC) superfamily is probably the largest and most diverse family of proteins that mediate ATP-dependent transfer of solutes. More than 100 ABC transporters have been identified in species ranging from Escherichia coli to human (1). The presence of ABC proteins in plants has been recently established by the cloning of three genes coding for members of this group in Arabidopsis thaliana (2–4). There are biochemical evidences that at least one of these genes, AtMRP1 (3), codes for a glutathione S-conjugated transporter, involved in detoxification processes of xenobiotics through their transfer in the vacuole after conjugation with glutathione (3, 5). In animals, two ABC proteins are directly involved in ion-channel regulation at the plasma membrane level: the sulfonylurea receptor, a complex of an inward rectifier K+ channel and the sulfonylurea receptor (6), and the cystic fibrosis transmembrane conductance regulator, expressed in a variety of secreting epithelia, regulated by cAMP-dependent phosphorylation and by ATP. The ATP-sensitive potassium channel (K-ATP), first identified in heart (7), is highly specific for K+ and inhibited by micromolar concentration of intracellular ATP. Afterward, K-ATP channels have been characterized in numerous tissues where they serve as metabolic sensors (8). The clinical importance of mutations affecting these proteins (9), cystic fibrosis and non-insulin-dependent diabetes, has contributed to the development of powerful pharmacological tools. Sulfonylureas, a class of sulfonamides, are potent blockers of K-ATP channels in numerous tissues (8, 10). These hypoglycemic compounds have become the major therapeutic agents used to treat non-insulin-dependent diabetes since 1955 (11). Sheppard and Welsh (12) have also reported inhibition by sulfonylureas of the whole-cell cystic fibrosis transmembrane conductance regulator. Usually, in animal cells, the inhibitory effect of sulfonylureas can be reversed by drugs able to increase the K+ permeability of K-ATP channels. These drugs, called K+ channel openers (KCOs), are powerful vasodilator agents that have been developed for clinical use in patients with hypertension, asthma, epilepsy, and ischemic heart disease (13).
To our knowledge, effects of sulfonylureas and KCOs on ion-channel regulation have not been reported in plant cells. However, sulfonylureas such as chlorsulfuron are commonly used for their herbicidal activity. An early response to treatment with chlorsulfuron is an inhibition of cell division in plant tissue (14). Sulfonylurea herbicides act by inhibition of the acetolactate synthase, required for the biosynthesis of branched-chain amino acids (15). Until now, the only report of an ATP-sensitive ion channel in plant cells was from Katsuhara et al. (16) who observed a decrease in the channel open probability upon addition of ATP to the cytoplasmic side, in the plasma membrane of a characeae alga. However, they did not report experimental results concerning a modulation of these currents by sulfonylureas or KCOs.
The aim of the present study was to take advantage of these pharmacological tools to characterize the role of an ABC protein in the regulation of ion channels in plants. Guard cells were chosen because they are the electrophysiologically best investigated cells in higher plants (17). In leaf epidermis, guard cells control the aperture of the stomatal pore that they surround, thus limiting water loss and gas exchange to the atmosphere. In response to environmental factors such as light, the pore is open by swelling of guard cells that take up potassium via an inward potassium channel as observed by Schroeder et al. (18). In contrast, closure of the pore triggered by darkness, CO2, or abscisic acid is related to an anion efflux. The resulting membrane depolarization drives K+ through outward-rectifying potassium channels (19). Thus, K+ channels at the plasma membrane of guard cell appear to play a key role in the regulation of stomatal movements. In the absence of any report concerning the effects of K-ATP channel modulators, we investigated the effects of sulfonylureas and KCOs on epidermal strip bioassays and on potassium currents, measured by using the whole-cell patch-clamp technique in guard cell protoplasts. Additionally, a molecular probe homologue of the sulfonylurea receptor was used to search for a transcript expressed in guard cells.
MATERIALS AND METHODS
Plant Material.
Plants of Commelina communis L. were grown under controlled environmental conditions as described (20). Plants of Vicia faba cv. Long pod were grown in pots (3 parts compost/1 part perlite) in growth chambers with a 8-h photoperiod (21°C, relative humidity 70%, 300 μmol per m2 per s). Guard-cell protoplasts and epidermal strips were isolated from young fully expanded leaves of 3- to 5-week-old plants.
Epidermal Strip Experiments.
Leaves of C. communis L. or V. faba were prepared as described (20, 21). Incubation solutions were 60 or 10 mM KCl for C. communis L. or V. faba respectively, in 10 mM Mes, pH 6.2. After 2.5 h of incubation, stomatal apertures were measured with an optical microscope fitted with a camera lucida and a digitizing table linked to a personal computer. For each treatment, 100 stomatal apertures (20 from five different epidermal pieces) were recorded in less than 3 min. Each data point is the mean of these 100 stomatal apertures. Error bars correspond to a 95% confidence interval to the mean. Plot and curve fitting (nonlinear regression by the Levenberg–Marquardt method) were done by using origin 4.0 (MicroCal Software, Northampton, MA). The concentration producing half-maximum opening or closing of stomata is given as K½. Total number of trials is given by n.
Patch-Clamp Experiments.
Guard cell protoplasts were isolated as described by Miedema and Assmann (22). After isolation, they were kept in the dark at 0–2°C for at least 1 h in a solution containing 500 μM MgCl2, 500 μM CaCl2, 5 mM Mes (adjusted to pH 5.5 with KOH), and 450 mM sorbitol. Guard-cell protoplast diameters measured with a calibrated eyepiece micrometer during patch-clamp experiments typically ranged from 16 to 19 μm.
By using the patch-clamp technique (23) in the whole-cell mode, currents through K+ channels were recorded from the plasma membrane of V. faba guard-cell protoplasts. Patch-clamp pipettes were pulled from borosilicate capillaries (Kimax-51 34500, Touzart et Matignon, Vitry sur Seine, France) on a PC-10 Narishige vertical puller (Nikon), fire-polished on a MF-90 Narishige microforge, and coated with beeswax (Aldrich) before use. Patch pipette solution contained 80 mM potassium glutamate, 20 mM KCl, 10 mM Hepes, 2 mM MgCl2, 2 mM ATP, and sorbitol to adjust an osmolality of 480 mmol/kg (adjusted to pH 7.8 with KOH). Guard cells were extracellularly perfused with a solution containing 5 mM MgCl2, 1 mM CaCl2, 10 mM Hepes, 100 mM potassium glutamate, and sorbitol to adjust an osmolality of 460 mmol/kg (adjusted to pH 7.2 with KOH). Final concentrations of potassium were 119 mM in the electrode solution and 104 mM in the bath. In these conditions, pipettes had resistances of 15–20 MΩ. The potential across the membrane (Vm) was calculated by correcting the potential read directly from the amplifier (Vp) for the liquid junction potential (EL) between the 80 mM potassium glutamate internal solution and the 100 mM potassium glutamate external solution, such that Vm = Vp − EL (24). Because we obtained EL = +0.7 mV, this correction was finally omitted. Experiments were performed at room temperature (22 ± 2°C).
Whole-cell recordings were obtained using an Axon Instruments Axopatch 200B amplifier (DIPSI Industrie, Chatillon, France) connected to a personal computer via a Digidata 1200 interface (Axon Instruments, Foster City, CA). pclamp 6.0.2 software (Axon Instruments) was used for voltage pulse stimulation, on-line data acquisition, and data analysis. After obtention of the whole-cell configuration, the membrane potential was held to −50 mV and stimulations were applied only 5 min later. The voltage protocol consisted in stepping the membrane potential to voltages from +80 mV to −160 mV for 2 s with a −20-mV increment. The membrane potential was held to −50 mV for 6 s between each voltage step. To ensure the time stability of the whole-cell K+ currents, stimulations were applied at least during 15 min before application of any modulator. Ion-channel modulators were added to the bath solution perfusion and the solution was exchanged in less than 2 min. Leak currents were subtracted as described (25). Whole-cell steady-state K+ current values, used to plot current–voltage curves or evolution of the current vs. time, were determined as the mean value over a period of 200 ms (900 samples), beginning 1.5 s after initiation of the test pulse. Because of variation in surface area from one protoplast to another, currents were normalized by the whole-cell capacitance. Cell capacitance (5–10 pF) was measured before each stimulation throughout the whole experiment by using the built-in capacity compensation device of the amplifier. Thus, normalized currents (pA/pF) were calculated by dividing the current by the whole-cell capacitance. Whole-cell currents were low-pass-filtered at 1 kHz by the four-pole Bessel filter of the amplifier, digitized at 3 kHz before storing on computer disk and Gaussian low-pass-filtered at 300 Hz for figures.
Subsequently, the inhibition percentage by sulfonylureas and KCOs will be called Inh and calculated at −160 mV or +80 mV, for inward and outward K+ current, respectively, according to the following relation: Inh = {1 − [IK(t)/IK(ctr)]} × 100, where IK(t) and IK(ctr) are, respectively, the normalized current after and before application of the drug. Error bars correspond to a 95% confidence interval to the mean.
RNA Extraction and Northern Blot Analysis.
Total RNA from whole plants, mature leaves, cotyledons, and purified guard cells from 4- to 5-week-old A. thaliana were extracted as described (26). Expression of ABC protein-like transcripts was determined by Northern blot analyses under stringent conditions using an A. thaliana expressed sequence tag clone homologue of the sulfonylurea receptor (206F4T7). Detection of the signals from the hybridized filters was performed with a PhosphorImager (Molecular Dynamics).
Chemicals.
Common chemicals were purchased from Sigma. RP49356 and RP52891 (aprikalim) were gifts from Rhône-Poulenc Rorer (Vitry-sur-Seine, France), SR47063 was from Sanofi Recherche (Montpellier, France), and BRL38227 (levcromakalim) was from SmithKline Beecham Pharmaceuticals (Worthing, UK). Glibenclamide and tolbutamide were dissolved in dimethyl sulfoxide (DMSO). Final concentration of DMSO in the incubation solution never exceeded 0.1% in epidermal strip experiments and was found to have no effect on stomatal opening. Note that glibenclamide was insoluble at concentrations higher than 25 μM in 1% DMSO.
RESULTS
Sulfonylureas Induce a Dose-Dependent Increase in Stomatal Aperture.
The effects of glibenclamide and tolbutamide on stomatal movements were investigated by using epidermal strips incubated for 3 h in darkness or white light. These bioassays were essentially conducted on C. communis epidermal strips but similar results were obtained with V. faba (data not shown).
Incubation of epidermal strips in the presence of various concentrations of sulfonylureas and then measurement of stomatal apertures allowed us to obtain typical dose–response curves (Fig. 1A). After 3 h, mean stomatal apertures for controls were 2.4 ± 0.3 μm (n = 53 bioassays) in darkness and 6.9 ± 0.6 μm (n = 46) in white light. Whatever the light treatment, the higher the sulfonylurea concentration was, the larger the stomatal apertures were. The increase in stomatal aperture induced by sulfonylureas was greater in darkness than in light. On the basis of independent dose–response experiments, K½ was 2.7 ± 2.1 μM (n = 6) for glibenclamide and 50 ± 23 μM (n = 4) for tolbutamide. With tolbutamide, a plateau was never obtained until a toxic effect was observed as a clustering of the chloroplasts usually followed by disruption of their envelopes. A total of 27 experiments were done, always leading to a significant increase in stomatal aperture. As frequently observed in animal tissues, the apparent efficiency of tolbutamide was lesser than that of glibenclamide. For this reason glibenclamide was used for further studies.
Figure 1.
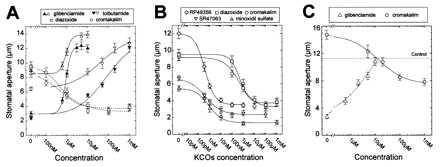
Typical bioassays illustrating the effects of sulfonylureas and KCOs on stomatal movements in C. communis. (A) A 2.5-h application of sulfonylureas (solid lines) induced a dose-dependent increase in stomatal aperture either in the dark (closed symbols) or under light (open symbols), whereas application of KCOs inhibited stomatal opening under light (dashed lines). (B) KCOs induced stomatal closure under light. Stomata were first illuminated for 3 h and then KCOs were added for a further 2.5 h. Differences in stomatal apertures, observed in absence of KCOs, are not attributable to experimental design but rather to an independent factor such as the annual cycle reported for the pump current by Lohse and Hedrich (27). (C) Stimulation of stomatal opening under illumination by 10 μM glibenclamide (solid line) was prevented by a concomitant application of cromakalim in a dose-dependent manner (○). Conversely, stomatal closure induced by 20 μM cromakalim under illumination (dashed line) was reversed by a concomitant addition of glibenclamide in a dose-dependent manner.
Potassium Channel Openers Inhibit Stomatal Opening and Induce Stomatal Closure.
In animal cells, many activators of potassium channels are also known for their ability to interact with the sulfonylurea receptor and to reverse the effects of glibenclamide and tolbutamide (28, 29). Because we demonstrated that sulfonylureas are able to promote an increase in stomatal aperture, we investigated the effects of KCOs on the regulation of stomatal movements. Among these KCOs, the effects of benzopyrans (cromakalim, SR47063, and BRL38227), benzothiadiazine (diazoxide), minoxidil (minoxidil sulfate), and thioformamides (RP49356 and RP52891) were tested in epidermal strips experiments. Stomata were first allowed to open in white light for 2.5 h, and then KCOs were added to the incubation medium for 2.5 h before measuring the final stomatal aperture. KCOs were able to prevent stomatal opening triggered by light in C. communis epidermal strips, as shown Fig. 1A for cromakalim and diazoxide and to induce stomatal closure in the light (Fig. 1B, n = 39). Similar results were obtained with V. faba (data not shown). Although cromakalim and diazoxide were active in the micromolar range (K½ = 1.3 μM and 0.5 μM; n = 5 and 4 respectively), RP49356, RP52891, minoxidil sulfate, and SR47063 very efficiently decreased stomatal aperture in light at subnanomolar concentrations (K½ = 0.5 nM, 0.9 nM, 0.6 nM, 0.4 nM, respectively; n = 11). Thus, maximum stomatal closure was obtained at about 10 nM RP49356, 10 nM RP52891, 10 nM minoxidil sulfate, or 10 nM SR47063 or with 100 μM cromakalim or diazoxide.
KCOs Reverse the Effect of Glibenclamide in a Dose-Dependent Manner.
As sulfonylureas and KCOs produce antagonistic effects on stomatal movements, the result of a concomitant application of these compounds was investigated in bioassays. As the typical experiment depicted in Fig. 1C (solid line) shows, addition of cromakalim was able to prevent the stomatal opening triggered by 10 μM glibenclamide (n = 5). Conversely, the stomatal closure induced by 20 μM cromakalim was reversed by a subsequent application of glibenclamide (dashed line; n = 4). The reversal effect obtained in the presence of both compounds was larger as the dose of cromakalim or glibenclamide increased, suggesting an apparent competition between these compounds. In two additional experiments, 0.4 nM RP49356 was found as efficient as 10 μM cromakalim to reverse the effect of 10 μM glibenclamide.
Glibenclamide Inhibits the Outward Potassium Current.
In control experiments, whole-cell current density at steady state 10 min after breaking the patch was 85 ± 6 pA/pF and −61 ± 5 pA/pF, for outward and inward potassium channels, respectively (n = 47 cells). After perfusion with 5 μM glibenclamide (0.1% DMSO), the inhibition developed slowly and it was sometimes difficult to distinguish the sulfonylurea effect from a possible run-down (n = 6). To test higher concentrations of glibenclamide, up to 25 μM, we were at the same time constrained to increase the DMSO percentage, up to 1%. Thus, in two groups of experiments, K+ channel activity in guard-cell protoplasts after application of DMSO with/without 25 μM glibenclamide were separately analyzed. A typical experiment among 34 replicates is presented in Fig. 2 A–D. Control recordings of inward and outward potassium currents are presented in Fig. 2A. Thirty minutes after perfusion with 25 μM glibenclamide (Fig. 2B), a partial but specific inhibition of the outward K+ current was observed (Inh = 52.1% and −11.7%, for outward and inward currents, respectively). For this experiment, the whole time course of the outward potassium current at 80 mV is presented in Fig. 2D and shows a maximal inhibition of 65% after a 50-min incubation period with glibenclamide. Current–voltage relationships (Fig. 2C) confirm the specific inhibition of the outward potassium current. Because a high concentration of DMSO was required to solubilize glibenclamide, additional experiments were carried out to evaluate the effect of 1% DMSO in the bath solution on K+ channel activity. One typical result is illustrated in Fig. 2E and shows that the activity of the normalized outward current after perfusion with 1% DMSO was almost constant. Moreover, addition of glibenclamide still led to an inhibition of outward currents. In eight experiments based on this protocol, slight inhibitions by 1% DMSO were observed (6.4 ± 25% and 8.1 ± 24%, respectively for outward and inward K+ currents). These data demonstrate that the decrease of outward potassium currents was mediated by glibenclamide but not by DMSO. Fig. 2F describes one trial of seven trials to reverse the inhibitory effect of glibenclamide, by washing of the bath solution with a sulfonylurea-free solution. The current restoration was not obtained, the reason being probably the large solubility of glibenclamide in phospholipids (30).
Figure 2.
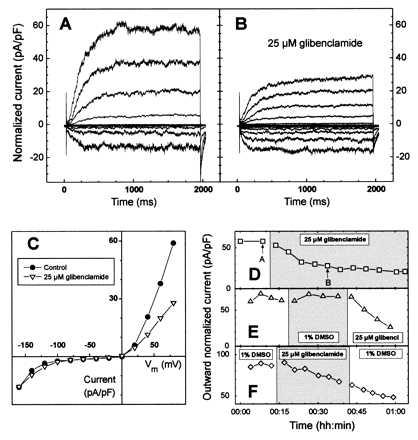
Partial inhibition by glibenclamide of outward potassium channels in V. faba guard cell. (A) Recordings of normalized inward (seen as downward deflections) and outward (upward deflections) potassium currents in whole-cell configuration. (B) Thirty minutes after perfusion with 25 μM glibenclamide, outward but not inward potassium currents were partially inhibited. (C) Superposition of current–voltage curves of the cell in A before (•) and after bath perfusion (B) with 25 μM glibenclamide (▿). (D) Reduction in the steady-state normalized outward potassium current vs. time after glibenclamide perfusion. Values correspond to the experiment described in A–C. Upward pointing arrows refer to current recordings illustrated in A and B. (E) Typical experiment illustrating the time course of the normalized outward potassium current. Channel activity was unaffected by DMSO perfusion but still sensitive to glibenclamide inhibition. (F) Even after a 20-min glibenclamide wash-out, the normalized outward potassium current was not recovered. (A–D) Whole-cell capacitance was 10 pF. Seal resistance was 2.5 GΩ. (D–F) Time course of the outward K+ current is given for a membrane potential of +80 mV. Changes of the bath solution are represented by alternating open and shaded areas.
On the basis of 34 experiments, a mean inhibition of 35 ± 8% of the outward potassium current was observed 20 min after perfusion with 25 μM glibenclamide. In 9 of these 34 experiments, outward K+ channel activity was decreased by 50–85%. In contrast, decrease in the inward K+ current in the presence of 25 μM glibenclamide was 18.2 ± 9% (n = 34), i.e., consistent with the run-down observed in the absence of glibenclamide (11.4 ± 8% each 10-min period, n = 44).
Outward Potassium Currents Inhibited by Glibenclamide Are Restored by Cromakalim Application.
To explain the results obtained in bioassays, the effect of a cromakalim application after glibenclamide inhibition of the whole-cell K+ channels was studied. A typical experiment is presented in Fig. 3. Application of 25 μM glibenclamide resulted in a 68% inhibition of the outward potassium current after 39 min (Fig. 3B), compared with control recordings (Fig. 3A). At the same time, the inward potassium current showed little or no evolution. When 25 μM glibenclamide in the bath solution was replaced with 100 μM cromakalim (Fig. 3C), a 75% increase in the outward K+ current was observed. Nevertheless, despite the continuous increase in the outward K+ current after cromakalim application (Fig. 3D), a complete restoration of the control current was not observed (Fig. 3, compare C to A). This phenomenon could result from a slow internal run-down of the current superposed on the competition between glibenclamide and cromakalim. Analysis of the normalized time course currents (Fig. 3D) showed that inward K+ currents slightly decrease during the whole experiment, without being affected by glibenclamide or cromakalim. Current–voltage curves in Fig. 3E confirm the specific effect of sulfonylureas and KCOs on outward potassium channels. In 11 cells, after a 39 ± 15% inhibition of the outward K+ current by 25 μM glibenclamide, current activity was enhanced by 40 ± 27% with KCOs (100 μM cromakalim or 100 nM RP49356, n = 8 and 3, respectively), and the inward current was inhibited by 12 ± 24%.
Figure 3.
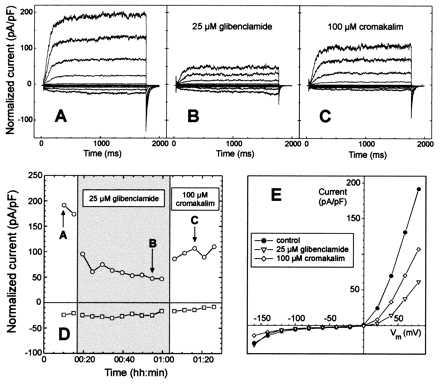
Inhibition of the outward potassium current by glibenclamide is partially reversed by cromakalim. (A) Recordings of normalized inward and outward potassium currents in whole-cell configuration. (B) Forty minutes after perfusion with 25 μM glibenclamide, outward potassium currents were partially inhibited without any noticeable effect on inward K+ currents. (C) Recovering of the outward potassium current 15 min after perfusion with 100 μM cromakalim. (D) Time course of the normalized inward and outward potassium current at +80 mV. Arrows indicate current recordings illustrated in A–C. Changes of the bath solution are represented by alternating open and shaded areas. (E) Superposition of current–voltage curves of the cell in A (•), after bath perfusion (B) with 25 μM glibenclamide (▿) and finally after perfusion (C) with 100 μM cromakalim (◊). (A–E) Whole-cell capacitance was 5 pF. Seal resistance was 1.5 GΩ.
Expression of an ABC Transcript in Guard Cells.
To investigate the presence of a transcript encoding for a sulfonylurea-receptor-like protein in guard cells, total RNAs from mature leaves, cotyledons, and purified guard cells were hybridized with a probe corresponding to an expressed sequence tag (206F4T7) identified as a coding sequence of a sulfonylurea-receptor-like protein. For all tissues analyzed, the probe hybridized with a 7-kb transcript (Fig. 4A), well within the range observed for other ABC transcripts. This band was the only 32P-labeled band detected. In accordance with the low level of rubisco in guard cells (20), when the small rubisco subunit of A. thaliana was used as a control probe, no band was detected in the lane corresponding to total RNA from guard cells (Fig. 4B) as described in epidermal fragments of potato leaves (31). This confirms that the guard cell preparation was devoid of mesophyll cell contaminants and that a transcript encoding a sulfonylurea-receptor-like protein is present in guard cells.
Figure 4.
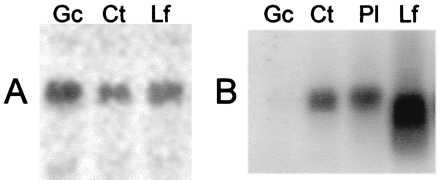
Northern blot analyses of expression of a sulfonylurea receptor homologue transcript (A) and of rubisco small subunit transcript (B) in guard cells (Gc), cotyledons (Ct), leaves (Lf), and whole plant (Pl). Ten micrograms of total RNA was loaded per lane.
DISCUSSION
In this study, the presence of a sulfonylurea-receptor-like protein in guard cells was investigated, and by using a pharmacological approach, its intervention in the control of stomatal movements and ion-channel regulation was analyzed. Sulfonylureas, tolbutamide and glibenclamide, two well-known ATP-sensitive potassium channels blockers in animal cells (28) induced stomatal opening in the dark and under light (Fig. 1A). Glibenclamide was more efficient than tolbutamide. In animal cells also, sulfonylureas potency appears very dependent on the compound and the tissue (8, 10, 32). It has been proposed that differences in lipophilicity of these compounds may be the major reason for the differences in apparent affinity for the receptor (11). In plant cells, very early studies have demonstrated that stomatal opening depends on K+ and anions repartition between the inside and outside of the cell (33). To explain the fact that sulfonylureas can induce stomatal opening, it was necessary to test, (i) a possible activation by these drugs of the inward K+ channel and (ii) an inhibition of the anionic or outward K+ channel. Fig. 2 A–D shows evidence that glibenclamide inhibits the outward potassium channel in guard-cell protoplasts, without any noticeable reduction in the activity of the inward K+ channel. In this study, outward potassium channel inhibition was obtained after perfusion with glibenclamide from the bath side of the membrane. Likewise, in animal cells, sulfonylureas are able to block K-ATP channels from both sides of the membrane. This is probably due to the fact that glibenclamide is a lipophilic molecule that may reach its target site by dissolving in the lipid phase, the un-ionized form being able to cross the membrane (30). Thus, glibenclamide may induce stomatal opening by a mechanism involving a yet unidentified receptor to finally result in the specific inhibition of the outward K+ channel.
Usually, to verify the specificity and to ensure that channel integrity is unaffected by an activator or inhibitor after its application, removal and washing of the drug from the bath solution is carried out. However, the relief of K-ATP channel inhibition by glibenclamide is frequently difficult to obtain, even after a long washing of the drug. After a 10-s exposure of isolated ventricular myocytes to 1 μM glibenclamide, Findlay (30) described (i) that the development of the inhibition continued for more than 170 s despite the removal of the drug and (ii) that the recovery of the current required an additional 400 s. He also demonstrated that the recovery from inhibition was largely dose-independent. In guard-cell protoplasts, as demonstrated for cystic fibrosis transmembrane conductance regulator (12), glibenclamide inhibition of the outward K+ channel was irreversible, even after prolonged washing of the sulfonylurea (Fig. 2F). Similar results of irreversible inhibition were obtained with inward K+ channels in guard cell by using charybdotoxin (34).
C. communis or V. faba bioassays showed that KCOs induce stomatal closure under light and prevent stomatal opening with high efficiency (subnanomolar K½ for RP49356, RP52891, SR47063, and minoxidil sulfate). This suggests that the K+ permeability through the outward K+ channel could be increased. KCOs concentrations efficient in promoting stomatal closure were generally lower than those needed for a physiological response in animal cells, typically 30–500 μM in heart tissue, 0.1–10 μM in smooth muscle, and 10–100 μM in β cells (8, 29). This result underlines the interest for the use of KCOs as a tool to study the physiology of stomatal movements. In whole-cell experiments, external perfusion of the bath with KCOs alone never resulted in a significant and reproductible activation of the outward K+ channel. However, to optimize the recording of outward K+ currents, the intracellular pipette pH was buffered at 7.8, a value at which outward K+ channels are already activated (34, 35). Moreover, as guard-cell protoplasts were basically isolated from closed stomata and kept in darkness until patch clamping, one could propose that, at the beginning of the experiment, protoplasts were already expressing the maximum efflux permeability to K+, reflecting their “closed state.” This argument may explain why noticeable activation of outward K+ channels was scarcely observed despite the evident efficiency of KCOs in epidermal strip experiments. Nevertheless, KCOs were able to activate outward K+ currents, because outward K+ channels previously inhibited by glibenclamide were reactivated by a cromakalim application (Fig. 3). Epidermal strip experiments demonstrate that KCOs and sulfonylureas triggered antagonistic effects upon stomatal movements. This result is in agreement with numerous studies in animal tissues in which glibenclamide reversed the effect of cromakalim or conversely (13, 29, 36). In this study, an apparent competition between these molecules was revealed when sulfonylureas and KCOs were simultaneously applied (Fig. 1C). This result reinforces the putative presence of a protein able to bind sulfonylureas and KCOs and involved in guard-cell osmoregulation.
Whether K-ATP channels are present and of functional significance in guard cells remains an open question. In this study, we demonstrate that stomatal movements are finely controlled by sulfonylureas and KCOs but this does not necessarily indicate the presence of K-ATP channels in our model system, as the relevant K+ channel and sulfonylurea receptors are separate molecules in animal cells (6). Moreover, the outward potassium channel from guard cells is certainly not an ATP-sensitive potassium channel because Wu and Assmann (25) described that whole-cell K+ currents were unaffected by the presence or absence of ATP in the pipette solution. Moreover, on the basis of comparisons with animal K-ATP channels, our experiments were performed in the presence of an inhibitory concentration of protons (known to activate K-ATP channels at a pH below 7) and with ATP in the pipette (37). However, the presence in guard cells of a protein functionally close to the sulfonylurea receptor may be suspected. Detection of a transcript in Northern blot analyses with a sulfonylurea receptor homologue probe (Fig. 4) supports the hypothesis that there is a sulfonylurea-receptor-like protein in guard cells. Such a sulfonylurea-receptor-like protein could be involved in the control of the outward potassium channel in guard cells similar to Kir 6.2 in pancreatic β cells (6) or Kir 1.1 in epithelial cells (38), two inward-rectifier potassium channels. However, we cannot excluded that the sulfonylurea-receptor-like protein that we described may be insensitive to intracellular ATP as reported for the sulfonylurea receptor 2B complexed to Kir 6.1 (39).
Thus, these results demonstrate the effects of K-ATP channels modulators on stomatal movements and guard cell plasma membrane K+ channel activity. They suggest the presence of a sulfonylurea-receptor-like protein, with a pharmacological profile similar to that of K-ATP channels in animal tissue and able to control, directly or not, the activity of the plasma membrane outward potassium channel in guard cells.
Acknowledgments
We thank Pr. S. M. Assmann for the generous gift of V. faba seeds and technical advice for the obtention of guard cell protoplasts. We gratefully acknowledge S. M. Assmann, H. Barbier-Brygoo, L. Romano, and M. B. Vivaudou for critically reading the manuscript.
ABBREVIATIONS
- ABC protein
ATP-binding cassette protein
- DMSO
dimethyl sulfoxide
- K-ATP channel
ATP-sensitive potassium channel
- KCO
potassium channel opener
References
- 1.Higgins C F. Cell. 1995;82:693–696. doi: 10.1016/0092-8674(95)90465-4. [DOI] [PubMed] [Google Scholar]
- 2.Dudler R, Hertig C. J Biol Chem. 1992;267:5882–5888. [PubMed] [Google Scholar]
- 3.Lu Y-P, Li Z-S, Rea P A. Proc Natl Acad Sci USA. 1997;94:8243–8248. doi: 10.1073/pnas.94.15.8243. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Marin, E., Leonhardt, N., Vavasseur, A. & Forestier, C. (1997) Biochim. Biophys. Acta, in press. [DOI] [PubMed]
- 5.Kreuz K, Tommasini R, Martinoia E. Plant Physiol. 1996;111:349–353. doi: 10.1104/pp.111.2.349. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Inagaki N, Gonoi T, Clement J P, Namba N, Inazawa J, Gonzalez G, Aguilar-Bryan L, Seino S, Bryan J. Science. 1995;270:1166–1170. doi: 10.1126/science.270.5239.1166. [DOI] [PubMed] [Google Scholar]
- 7.Noma A. Nature (London) 1983;305:147–148. doi: 10.1038/305147a0. [DOI] [PubMed] [Google Scholar]
- 8.Gopalakrishnan M, Janis R A, Triggle D J. Drug Dev Res. 1993;28:95–127. [Google Scholar]
- 9.Demolombe S, Escande D. Trends Pharmacol Sci. 1996;17:273–275. doi: 10.1016/0165-6147(96)10037-7. [DOI] [PubMed] [Google Scholar]
- 10.Ashcroft S J, Ashcroft F M. Cell Signalling. 1990;2:197–214. doi: 10.1016/0898-6568(90)90048-f. [DOI] [PubMed] [Google Scholar]
- 11.Panten U, Schwanstecher M, Schwanstecher C. Exp Clin Endocrinol. 1996;104:1–9. doi: 10.1055/s-0029-1211414. [DOI] [PubMed] [Google Scholar]
- 12.Sheppard D N, Welsh M J. J Gen Physiol. 1992;100:573–591. doi: 10.1085/jgp.100.4.573. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Lawson K. Pharmacol Ther. 1996;70:39–63. doi: 10.1016/0163-7258(96)00003-4. [DOI] [PubMed] [Google Scholar]
- 14.Ray T B. Pest Biochem Physiol. 1982;17:10–17. [Google Scholar]
- 15.Chaleff R S, Mauvais C J. Science. 1984;224:1143–1145. doi: 10.1126/science.224.4656.1443. [DOI] [PubMed] [Google Scholar]
- 16.Katsuhara M, Mimura T, Tazawa M. Plant Physiol. 1990;93:343–346. doi: 10.1104/pp.93.1.343. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Assmann S M. Annu Rev Cell Biol. 1993;9:345–375. doi: 10.1146/annurev.cb.09.110193.002021. [DOI] [PubMed] [Google Scholar]
- 18.Schroeder J I, Hedrich R, Fernandez J M. Nature (London) 1984;312:361–362. [Google Scholar]
- 19.Schroeder J I, Hagiwara S. Nature (London) 1989;338:427–430. [Google Scholar]
- 20.Cotelle V, Forestier C, Vavasseur A. Physiol Plant. 1996;98:619–628. [Google Scholar]
- 21.Weyers J, Meidner H. Methods in Stomatal Research. New York: Longman; 1990. pp. 135–137. [Google Scholar]
- 22.Miedema H, Assmann S M. J Membr Biol. 1996;154:227–237. doi: 10.1007/s002329900147. [DOI] [PubMed] [Google Scholar]
- 23.Hamill O P, Marty A, Neher E, Sakmann B, Sigworth F J. Pflügers Arch. 1981;391:85–100. doi: 10.1007/BF00656997. [DOI] [PubMed] [Google Scholar]
- 24.Barry P H, Lynch J W. J Membr Biol. 1991;121:101–117. doi: 10.1007/BF01870526. [DOI] [PubMed] [Google Scholar]
- 25.Wu W H, Assmann S M. Plant Physiol. 1995;107:101–109. doi: 10.1104/pp.107.1.101. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Verwoerd T C, Dekker B M M, Hoekema A. Nucleic Acids Res. 1989;17:2362. doi: 10.1093/nar/17.6.2362. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Löhse G, Hedrich R. Planta. 1992;188:206–214. doi: 10.1007/BF00216815. [DOI] [PubMed] [Google Scholar]
- 28.Ashcroft S J, Ashcroft F M. Biochim Biophys Acta. 1992;1175:45–59. doi: 10.1016/0167-4889(92)90008-y. [DOI] [PubMed] [Google Scholar]
- 29.Cook N S, Quast U. In: Potassium Channels: Structure, Classification, Function and Therapeutic Potential. Cook N S, editor. Chichester, U.K.: Ellis Horwood; 1990. pp. 181–255. [Google Scholar]
- 30.Findlay I. J Pharmacol Exp Ther. 1992;261:540–545. [PubMed] [Google Scholar]
- 31.Kopka J, Provart N J, Müller-Röber B. Plant J. 1997;11:871–882. doi: 10.1046/j.1365-313x.1997.11040871.x. [DOI] [PubMed] [Google Scholar]
- 32.Castle N A, Haylett D G. J Physiol. 1987;383:31–43. doi: 10.1113/jphysiol.1987.sp016394. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Willmer C, Fricker M. Stomata. 2nd Ed. London: Chapman & Hall; 1996. pp. 228–309. [Google Scholar]
- 34.Blatt M R. J Gen Physiol. 1992;99:615–644. doi: 10.1085/jgp.99.4.615. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Lemtiri-Chlieh F, MacRobbie E A C. J Membr Biol. 1994;137:99–107. doi: 10.1007/BF00233479. [DOI] [PubMed] [Google Scholar]
- 36.Sanguinetti M C, Scott A L, Zingaro G J, Siegl P K S. Proc Natl Acad Sci USA. 1988;85:8360–8364. doi: 10.1073/pnas.85.21.8360. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Vivaudou M, Forestier C. J Physiol. 1995;486:629–645. doi: 10.1113/jphysiol.1995.sp020840. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.McNicholas C M, Guggino W B, Schwiebert E M, Hebert S C, Giebisch G, Egan M E. Proc Natl Acad Sci USA. 1996;93:8083–8088. doi: 10.1073/pnas.93.15.8083. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Yamada M, Isomoto S, Matsumoto S, Kondo C, Shindo T, Horio Y, Kurachi Y. J Physiol. 1997;499:715–720. doi: 10.1113/jphysiol.1997.sp021963. [DOI] [PMC free article] [PubMed] [Google Scholar]