

Abstract
The menstrual cycle has been reported to alter mean arterial pressure (MAP), but not muscle sympathetic nerve activity (MSNA), during vestibular activation. Specifically, MAP responses to head-down rotation (HDR) are augmented during the midluteal (ML) phase compared to the early follicular (EF) phase in young, eumenorrheic women. The purpose of the present study was to determine if the menstrual cycle influences vestibular-mediated changes in limb blood flow. MSNA, MAP, heart rate, and limb blood flow responses to HDR were measured in 12 healthy women. Resting MSNA, MAP, heart rate, forearm blood flow and calf blood flow were not altered by the menstrual cycle. HDR elicited similar increases in MSNA during the EF (Δ3 ± 1 bursts/min; P < 0.05) and ML (Δ2 ± 1 bursts/min; P < 0.05) phase, but only increased MAP during the ML phase (Δ4 ± 2 mmHg; P < 0.05). HDR did not change heart rate during either the EF or ML phase. HDR elicited similar increases in calf vascular resistance during the EF (Δ6 ± 2 mmHg/mL/100mL/min; P < 0.05) and ML (Δ7 ± 2 mmHg/mL/100mL/min; P < 0.05) phases of the menstrual cycle. In contrast, HDR increased forearm vascular resistance during the ML phase (Δ4 ± 2 mmHg/mL/100mL/min; P<0.05), but not the EF phase (Δ0 ± 2 mmHg/mL/100mL/min). These findings suggest an increased transduction of sympathetic nerve activity into forearm vascular resistance during the ML phase, and reveal the first recorded divergent vascular response to vestibular excitation in human limbs.
Keywords: Muscle sympathetic nerve activity, autonomic activity, arterial blood pressure, head-down rotation, estrogen
INTRODUCTION
Orthostatic intolerance is the inability to maintain arterial blood pressure and cerebral perfusion while standing. Symptoms include lightheadedness, dizziness, nausea, and fatigue, and this condition can ultimately result in syncope. The baroreflex is recognized as the primary modulator of beat-to-beat arterial blood pressure during an orthostatic challenge, but evidence suggests the vestibulosympathetic reflex also contributes to postural blood pressure regulation (Monahan et al., 2002b; Ray, 2000b). In fact, the vestibulosympathetic reflex may actually be the first autonomic reflex to respond to orthostatic stress as the latency of the vestibulosympathetic reflex is shorter than the baroreflex (Kaufmann et al., 2002). Therefore, examining the vestibulosympathetic reflex in populations susceptible to orthostatic intolerance is clinically relevant.
It is well established that women are less likely to maintain postural blood pressure than men (Convertino, 1998; Fu et al., 2004; Fu et al., 2005). We recently examined the vestibulosympathetic reflex during the early follicular (EF) and midluteal (ML) phases of the menstrual cycle in young, healthy women (Lawrence et al., 2008). Although we reported no differences in vestibular-mediated sympathoexcitation between menstrual phases, blood pressure responses to vestibular activation were augmented during the ML phase (Lawrence et al., 2008). We credited these results to an increase in the transduction of sympathetic nerve activity into vascular resistance, a phenomenon referred to as ‘vascular transduction’, but could not conclude this with certainty as limb blood flow was not examined. Therefore, the primary purpose of the present study was to evaluate the influence of the menstrual cycle on vestibular-mediated changes in limb blood flow. Based on our previous study (Lawrence et al., 2008), we hypothesized an augmented vascular transduction during the ML phase of the menstrual cycle.
METHODS
Subjects
Twelve healthy women (age 21 ± 1 yr, height 166 ± 2 cm, weight 68 ± 4 kg) participated. Subjects were nonsmokers, nondiabetics, not taking oral contraceptives and had regular menstrual cycles (i.e., 26–30 days in length). All subjects were instructed to abstain from exercise, caffeine, and alcohol for 12 hours prior to laboratory testing. All participants provided written informed consent and the Michigan Technological University Institutional Review Board approved the experimental protocols.
Experimental design
Subjects were tested twice (at the same time of day) and selection of the initial menstrual cycle phase was randomized. Of the 12 subjects, 6 began the study during the EF phase and 6 began the study during the ML phase. Subjects were tested once during the EF phase (3 ± 0 days after start of menstruation) and once during the ML phase (23 ± 0 days after start of menstruation). The experiment was conducted with subjects lying prone. All subjects were adjusted so that the neck could maximally flex and perform head-down rotation (HDR) without interference from the table. The neck was then fully extended with a face rest supporting the head to approximate a position of normal gravitational orientation experienced during upright posture. Baseline values were recorded in this position for 3 minutes. Following baseline, the face rest was removed and HDR was performed for 3 minutes. During HDR, the neck was passively flexed (the head moved toward the floor and the chin moved toward the chest). Muscle sympathetic nerve activity (MSNA), heart rate, beat-to-beat arterial blood pressure, forearm blood flow and calf blood flow were acquired.
Measurements
Multifiber recordings of MSNA were made by inserting a tungsten microelectrode into the peroneal nerve in the popliteal region. A reference electrode was inserted subcutaneously 2–3 cm from the recording electrode. Both electrodes were connected to a differential preamplifier and then to an amplifier (total gain of 80,000), where the nerve signal was band-pass filtered (700–2,000 Hz) and integrated (time constant 0.1 seconds) to obtain a mean voltage display of the nerve activity. Satisfactory recordings of MSNA were defined by spontaneous pulse synchronous bursts that increased during end-expiratory apnea and did not change during stroking of the skin or auditory stimulation. MSNA recordings were not reproduced during both EF and ML phases in two subjects; thus, we report 10 subjects for MSNA.
Arterial blood pressure was measured using two techniques. Resting arterial blood pressure was measured three consecutive times (separated by 1 minute intervals) with an automated sphygmomanometer and reported as a mean value. Beat-to-beat arterial blood pressure was recorded continuously via Finometer (Finapres Medical Systems, Amsterdam, The Netherlands). The Finometer allowed for detection of precise changes in blood pressure during the interventions, while the sphygmomanometer was used for comparison of baseline arterial pressure during the EF and ML phases. Continuous heart rate was recorded using a three-lead electrocardiogram.
Blood flow recordings were collected using venous occlusion plethysmography (EC6, D.E. Hokanson Inc., Bellevue, WA, USA). Pressure cuffs were placed around the ankle and wrist and inflated to 220 mmHg to occlude the flow to the hand and foot. Mercury-in-silastic strain gauges were placed around the maximal circumference of the calf and forearm for detection of volume changes. Thigh and brachial cuffs were inflated during a time interval to 60 mmHg to block venous, but not arterial, flow of the limb. Inflow recordings were 8 seconds with reading intervals of 15 seconds. Vascular resistance was calculated as the mean arterial pressure (MAP) divided by limb blood flow. Only 10 subjects are reported for calf blood flow due to signal artifact in 2 subjects.
Statistical analysis
Heart rate, arterial blood pressure and MSNA data were imported and analyzed in WinCPRS on a Windows-formatted computer. Nerve activity was expressed as MSNA (burst frequency) and total MSNA. Total activity equals the total number of bursts multiplied by the averaged normalized burst area. Blood flow was collected and analyzed using NIVP3 software (D.E. Hokanson Inc., Bellevue, WA, USA). Flow slopes were determined by the software and confirmed by an investigator. Slope recordings with artifact were removed for statistical analysis.
All data were analyzed statistically using commercial software (SPSS 15.0; SPSS, Chicago, IL). A 2×2 repeated-measures ANOVA was calculated for changes in heart rate, MAP, MSNA and blood flow parameters during each condition (baseline and HDR) and across trials (menstrual cycle phases). Paired t-test statistics were calculated for post hoc analyses. Resting variables were compared using paired t-tests. Means were considered significantly different when P < 0.05 and results are expressed as mean ± SE. All delta (Δ) values are reported as changes from the baseline.
RESULTS
Baseline values during the EF and ML phases are presented in Table 1. Resting heart rates were ~10 beats/min lower in the prone position compared to the seated values reported in Table 1 (EF: 71 ± 3 beats/min; ML: 73 ± 3 beats/min). Resting MSNA, MAP, heart rate, forearm blood flow, and calf blood flow were not different between EF and ML phases.
Table 1.
Baseline Recordings
| Variable | n | EF | ML | P value |
|---|---|---|---|---|
| SAP (mmHg) | 12 | 107 ± 3 | 110 ± 3 | 0.29 |
| DAP (mmHg) | 12 | 68 ± 3 | 69 ± 2 | 0.78 |
| MAP (mmHg) | 12 | 81 ± 3 | 83 ± 2 | 0.52 |
| HR (beats/min) | 12 | 83 ± 5 | 84 ± 3 | 0.87 |
| MSNA (bursts/min) | 10 | 9 ± 2 | 6 ± 1 | 0.39 |
| CBF (mL/min/100mL) | 10 | 3.0 ± 0.3 | 2.4 ± 0.3 | 0.18 |
| FBF (mL/min/100mL) | 12 | 2.8 ± 0.3 | 2.7 ± 0.3 | 0.77 |
SAP, systolic arterial pressure; DAP, diastolic arterial pressure; MAP, mean arterial pressure; HR, heart rate; MSNA, muscle sympathetic nerve activity; CBF, calf blood flow; FBF, forearm blood flow; EF, early follicular phase; ML, mid-luteal phase; values are mean ± SE
Figure 1 demonstrates that HDR increased MSNA burst frequency during both the EF (Δ3 ± 1 bursts/min; P < 0.05) and ML (Δ2 ± 1 bursts/min; P < 0.05) phases, but only increased MAP (Δ4 ± 2 mmHg; P < 0.05) during the ML phase. The increases in MSNA burst frequency were not different between phases. HDR increased total MSNA during the ML phase (Δ172 ± 89 a.u.; P = 0.04) and tended to increase during the EF phase (Δ254 ± 154 a.u., P < 0.08). HDR did not alter heart rate during either phase.
Figure 1.
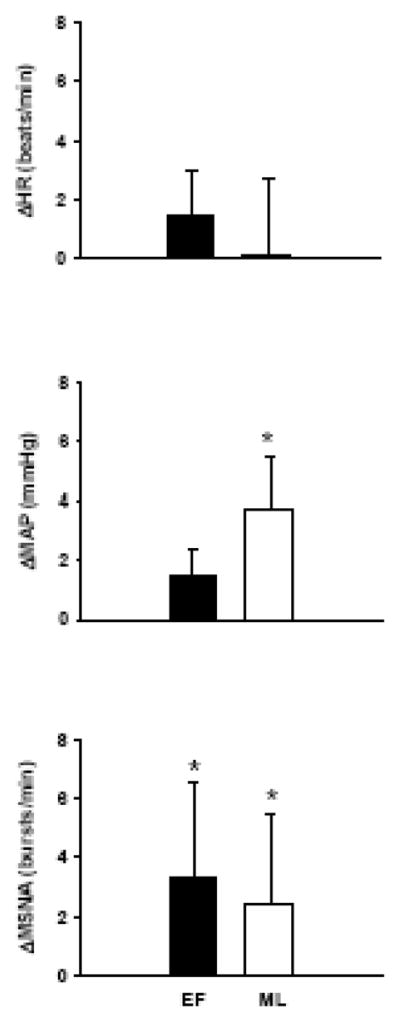
Changes in heart rate (HR), mean arterial pressure (MAP), and muscle sympathetic nerve activity (MSNA) during HDR. Subjects were examined during the early follicular (EF) and midluteal (ML) phases of the menstrual cycle. HDR increased MSNA during both phases, but only increased MAP during the ML phase. Values are reported as mean ± SE. *P < 0.05 vs. baseline.
HDR elicited similar decreases in calf blood flow during the EF (Δ−0.5 ± 0.2 ml/100mL/min; P < 0.05) and ML (Δ−0.2 ± 0.1 ml/100mL/min; P < 0.01) phases, but did not alter forearm blood flow during either phase. Figure 2 demonstrates that HDR elicited similar increases in calf vascular resistance during the EF (Δ6 ± 2 mmHg/mL/100mL/min; P < 0.05) and ML (Δ7 ± 2 mmHg/mL/100mL/min; P < 0.05) phases. In contrast, HDR increased forearm vascular resistance during the ML (Δ4 ± 2 mmHg/mL/100mL/min; P < 0.05), but not the EF (Δ0 ± 2 mmHg/mL/100mL/min), phase of the menstrual cycle.
Figure 2.

Changes in vascular resistance of the forearm (FVR) and calf (CVR) during head-down rotation (HDR). Subjects were examined during the early follicular (EF) and midluteal (ML) phases of the menstrual cycle. HDR increased CVR similarly during the EF and ML phases, but FVR increased only during the ML phase. Values are reported as mean ± SE. *P < 0.05 vs. baseline; †P < 0.05 vs. EF phase.
DISCUSSION
The present study reveals two novel findings. First, vestibular activation elicits a divergent forearm vascular response in women. Specifically, HDR induces forearm vasoconstriction during the ML, but not the EF, phase of the menstrual cycle. Second, forearm and calf vascular responses to HDR are similar during the ML, but not the EF, phase of the menstrual cycle. To our knowledge, this is the first report of divergent vascular responses during vestibular activation in humans. The consistent increase of calf vascular resistance during both the EF and ML phases supports the concept that vestibular activation contributes to postural blood pressure regulation. The divergent forearm vascular response suggests an augmentation of sympathetic neural transduction to the vascular smooth muscle of the forearm during the ML phase.
Vasoconstriction of the limbs during vestibular activation is well documented (Carter et al., 2005a; Hume et al., 1999; Monahan et al., 2002a; Ray, 2000a; Ray et al., 1997; Shortt et al., 1997). However, recent studies from both humans (Monahan et al., 2002a) and animals (Kerman et al., 2000; Wilson et al., 2006) suggest that the vestibular system may elicit patterned alterations in limb blood flow. Specifically, Monahan and Ray (2002a) reported that the HDR model of vestibular activation increased both calf and forearm vascular resistance, but that the increases in calf vascular resistance were 60% greater than the increases observed in the forearm. The present study supports the findings of Monahan and Ray (2002a), as we also demonstrate a greater vasoconstriction of the calf compared to the forearm. Unlike Monahan and Ray (2002a), who reported a >10% change in forearm blood flow, HDR did not alter forearm blood flow in the present study. One potential explanation for this difference is the subject pool. The study by Monahan and Ray (2002a) included men, while the present study was restricted to women. However, if sex differences exists regarding blood flow responses to vestibular activation, they appear to be isolated to the forearm as calf blood flow and calf vascular resistance responses to HDR appear to similar between sexes (Ray, 2000a).
More recently in animals, Wilson et al. (2006) reported that bilateral vestibular neurectomy did not alter brachial blood flow and vascular resistance during head-up tilt in conscious cats, whereas removal of vestibular inputs significantly blunted femoral vascular resistance. Kerman et al. (2000) also reported dissimilar blood flow to the forelimbs and hind limbs of cats during electrical vestibular stimulation. These studies (Kerman et al., 2000; Monahan et al., 2002a; Wilson et al., 2006) suggest that the vestibular system contributes importantly to vasoconstriction of body regions where blood pooling can occur during postural stress (i.e., legs), supporting the concept that vestibular-autonomic control may help to protect against the occurrence of orthostatic intolerance.
Until now, divergent vascular responses in the arms and legs of humans have been primarily limited to mental stress and isometric handgrip exercise (Carter et al., 2005b; Rusch et al., 1981). Rusch et al. (1981) reported that mental stress vasodilates the forearm, but not the calf, and that isometric handgrip exercise vasodilates the non-exercising forearm and vasoconstricts the calf. The results of the present study indicate that vestibular stimulation also elicits divergent limb vascular responses. However, this vestibular-mediated divergent vascular response was only present during the EF phase (i.e., low estrogen, low progesterone) in young, healthy women. The mechanisms responsible for this divergence are not entirely clear. A previous study indicates that HDR elicits similar increases of MSNA in the arm and leg (Monahan et al., 2002a). Furthermore, simultaneous recordings of MSNA during mental stress reveal divergent vascular responses to the forearm and calf despite similar levels of MSNA to both limbs (Carter et al., 2005b). Consistent with our previous study (Lawrence et al., 2008), the present study reports that the menstrual cycle did not alter vestibular-mediated MSNA in humans. Thus, it seems unlikely that MSNA is contributing to the divergent limb vascular response during the EF phase, but forearm MSNA was not measured in the present study.
Although the menstrual cycle does not appear to influence the vestibulosympathetic reflex, recent studies suggest the menstrual cycle may alter other sympathoexcitatory reflexes. In particular, MSNA responses to lower body negative pressure (Carter et al., 2009) and head-up tilt (Fu et al., 2009) are augmented during the ML phase of the menstrual cycle. Furthermore, other studies have reported an augmentation of sympathetic activity during the ML phase (Minson et al., 2000; Park et al., 2009). Overall, evidence is accumulating that the ML phase is often associated with some degree of augmented sympathetic neural sensitivity. In a recent study (Lawrence et al., 2008), we reported that the menstrual cycle did not alter MSNA responses to HDR, but that HDR increased MAP only during the ML phase. We suggested that the augmented MAP response was due to an increased sympathetic transduction to the vascular smooth muscle during the ML phase, thus fitting with the concept that sensitivity of sympathetic neural activity is heightened during high levels of estrogen and progesterone. Unfortunately, our previous study (Lawrence et al., 2008) did not have measures of limb blood flow to confirm.
The present study once again revealed that HDR elicits similar increases in MSNA during the EF and ML phases of the menstrual cycle, and increased MAP only during the ML phase. More importantly, we observed forearm vasoconstriction during the ML, but not EF, phase. Although vascular transduction is directly evaluated as the slope of the relationship between MSNA and vascular resistance during a sympatho-excitatory maneuver such as ischemic handgrip (Minson et al., 2000), the present findings indirectly support the concept of an augmented sympathetic transduction to the vasculature during the ML phase. However, this augmentation appears to be isolated to the upper limb extremities. It is unclear why this augmentation was only present in the forearm. The divergent vascular response observed in the present study, coupled with divergent vascular responses to mental stress (Carter et al., 2005b; Rusch et al., 1981), suggest a need to better understand the regulation of limb blood flow responses during sympathoexcitatory maneuvers. Moreover, the relations between female sex steroids and neurovascular control deserve more attention as women are more prone to orthostatic intolerance. The potential augmentation of sympathetic transduction to the vasculature during the ML phase reported in the present study, along with recent reports of augmented MSNA responses to orthostatic stress during the ML phase (Carter et al., 2009; Fu et al., 2009), suggest that female reproductive hormones may play a pivotal role in autonomic reflexes activated during orthostasis..
Increased sympathetic transduction to the forearm could reflect a larger quantity of norepinephrine (NE) released for a given level of MSNA. Two recent studies (Carter et al., 2009; Fu et al., 2009) have reported that orthostatic stress increases total MSNA, but not MSNA burst frequency, across the menstrual cycle. It is possible that increased sensitivity to sympathoexcitatory maneuvers during the ML phase may result in augmented signal strength, not signal rate, of sympathetic fibers. These findings may translate to individual sympathetic bursts providing increased NE to the adrenergic receptors. NE responses to HDR were not measured in the present study.
Another possible explanation for the divergent forearm vascular resistance could be a greater sensitivity of the forearm vasculature to a given amount of neurotransmitter released. Compared to the EF phase, sensitivity of α1-adrenergic receptors appears to be greater during the luteal phase (Freedman et al., 2000) and at mid-cycle (Chan et al., 2001). Infusion of equal amounts of NE during both the EF and ovulatory phases resulted in greater constriction during mid-cycle (Chan et al., 2001). Thus, the sensitivity to NE, not the amount of the NE, in the vasculature may be increased with elevated estrogen levels. Although the effects of endogenous gonadal hormones on α2-adrenergic receptors are not well established, evidence suggests that α2C-adrenergic receptors exist in human arterial smooth muscle cells and are involved with vasoconstriction (Chotani et al., 2005). Furthermore, treatment of 17β-estradiol on cultured human vascular smooth muscle cells interacted with cell surface receptors to cause an increase in α2C-adrenergic receptor transcription (Eid et al., 2007). If the expression of α2C-adrenergeic receptors is greater in the forearm during the ML phase, the response to HDR might result in greater vasoconstriction. This could explain the augmented vestibular-mediated vascular response reported during the ML phase. It is not fully understood what mechanisms are responsible for the increase in sympathetic sensitivity.
The data above suggests that the increase in sympathetic sensitivity is related to elevated estrogen. However, it is well recognized that estrogen has a vasodilator effect on smooth muscle. This response occurs through the excitation of β-adrenergic receptors. Kneale et al. (2000) blocked β-adrenergic receptors in both men and women using propanolol, and greater vasoconstriction to norepinephrine was observed in the women (Kneale et al., 2000). Kneale et al. (2000) conducted the study when estrogen was elevated and progesterone was low in their subjects (i.e., mid-cycle). Therefore, these data suggest that progesterone, not estrogen, may be contributing to the augmented forearm vasoconstriction observed in the present study.
In conclusion, vestibular activation elicits a divergent forearm vascular response in women. HDR induces forearm vasoconstriction during the ML, but not the EF, phase of the menstrual cycle. Furthermore, forearm and calf vascular responses to HDR are similar during the ML, but not the EF, phase of the menstrual cycle. The parallel and robust increase of calf vascular resistance during both the EF and ML phases supports the concept that vestibular activation contributes importantly to postural blood pressure regulation, whereas the divergent forearm vascular response suggests an augmentation of sympathetic neural transduction to the vascular smooth muscle of the forearm during the ML phase.
Acknowledgments
The authors thank Christopher Schwartz for technical assistance. This project was supported by a grant from the National Institutes of Health (HL-088689).
Footnotes
Publisher's Disclaimer: This is a PDF file of an unedited manuscript that has been accepted for publication. As a service to our customers we are providing this early version of the manuscript. The manuscript will undergo copyediting, typesetting, and review of the resulting proof before it is published in its final citable form. Please note that during the production process errors may be discovered which could affect the content, and all legal disclaimers that apply to the journal pertain.
References
- Carter JR, Cooke WH, Ray CA. Forearm neurovascular responses during mental stress and vestibular activation. Am J Physiol Heart Circ Physiol. 2005a;288:H904–907. doi: 10.1152/ajpheart.00569.2004. [DOI] [PubMed] [Google Scholar]
- Carter JR, Kupiers NT, Ray CA. Neurovascular responses to mental stress. J Physiol. 2005b;564:321–327. doi: 10.1113/jphysiol.2004.079665. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Carter JR, Lawrence JE, Klein JC. Menstrual cycle alters sympathetic neural responses to orthostatic stress in young, eumenorrheic women. Am J Physiol Endocrinol Metab. 2009;297:E85–91. doi: 10.1152/ajpendo.00019.2009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chan NN, MacAllister RJ, Colhoun HM, Vallance P, Hingorani AD. Changes in endothelium-dependent vasodilatation and alpha-adrenergic responses in resistance vessels during the menstrual cycle in healthy women. J Clin Endocrinol Metab. 2001;86:2499–2504. doi: 10.1210/jcem.86.6.7581. [DOI] [PubMed] [Google Scholar]
- Chotani MA, Mitra S, Eid AH, Han SA, Flavahan NA. Distinct cAMP signaling pathways differentially regulate alpha2C-adrenoceptor expression: role in serum induction in human arteriolar smooth muscle cells. Am J Physiol Heart Circ Physiol. 2005;288:H69–76. doi: 10.1152/ajpheart.01223.2003. [DOI] [PubMed] [Google Scholar]
- Convertino VA. Gender differences in autonomic functions associated with blood pressure regulation. Am J Physiol. 1998;275:R1909–1920. doi: 10.1152/ajpregu.1998.275.6.R1909. [DOI] [PubMed] [Google Scholar]
- Eid AH, Maiti K, Mitra S, Chotani MA, Flavahan S, Bailey SR, Thompson-Torgerson CS, Flavahan NA. Estrogen increases smooth muscle expression of alpha2C-adrenoceptors and cold-induced constriction of cutaneous arteries. Am J Physiol Heart Circ Physiol. 2007;293:H1955–1961. doi: 10.1152/ajpheart.00306.2007. [DOI] [PubMed] [Google Scholar]
- Freedman RR, Girgis R. Effects of menstrual cycle and race on peripheral vascular alpha-adrenergic responsiveness. Hypertension. 2000;35:795–799. doi: 10.1161/01.hyp.35.3.795. [DOI] [PubMed] [Google Scholar]
- Fu Q, Arbab-Zadeh A, Perhonen MA, Zhang R, Zuckerman JH, Levine BD. Hemodynamics of orthostatic intolerance: implications for gender differences. Am J Physiol Heart Circ Physiol. 2004;286:H449–457. doi: 10.1152/ajpheart.00735.2002. [DOI] [PubMed] [Google Scholar]
- Fu Q, Okazaki K, Shibata S, Shook RP, Vangunday TB, Galbreath MM, Reelick MF, Levine BD. Menstrual cycle effects on sympathetic neural responses to upright tilt. J Physiol. 2009 doi: 10.1113/jphysiol.2008.168468. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fu Q, Witkowski S, Okazaki K, Levine BD. Effects of gender and hypovolemia on sympathetic neural responses to orthostatic stress. Am J Physiol Regul Integr Comp Physiol. 2005;289:R109–116. doi: 10.1152/ajpregu.00013.2005. [DOI] [PubMed] [Google Scholar]
- Hume KM, Ray CA. Sympathetic responses to head-down rotations in humans. J Appl Physiol. 1999;86:1971–1976. doi: 10.1152/jappl.1999.86.6.1971. [DOI] [PubMed] [Google Scholar]
- Kaufmann H, Biaggioni I, Voustianiouk A, Diedrich A, Costa F, Clarke R, Gizzi M, Raphan T, Cohen B. Vestibular control of sympathetic activity. An otolith-sympathetic reflex in humans. Exp Brain Res. 2002;143:463–469. doi: 10.1007/s00221-002-1002-3. [DOI] [PubMed] [Google Scholar]
- Kerman IA, Emanuel BA, Yates BJ. Vestibular stimulation leads to distinct hemodynamic patterning. Am J Physiol Regul Integr Comp Physiol. 2000;279:R118–125. doi: 10.1152/ajpregu.2000.279.1.R118. [DOI] [PubMed] [Google Scholar]
- Kneale BJ, Chowienczyk PJ, Brett SE, Coltart DJ, Ritter JM. Gender differences in sensitivity to adrenergic agonists of forearm resistance vasculature. J Am Coll Cardiol. 2000;36:1233–1238. doi: 10.1016/s0735-1097(00)00849-4. [DOI] [PubMed] [Google Scholar]
- Lawrence JE, Ray CA, Carter JR. Vestibulosympathetic reflex during the early follicular and midluteal phases of the menstrual cycle. Am J Physiol Endocrinol Metab. 2008;294:E1046–1050. doi: 10.1152/ajpendo.00056.2008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Minson CT, Halliwill JR, Young TM, Joyner MJ. Influence of the menstrual cycle on sympathetic activity, baroreflex sensitivity, and vascular transduction in young women. Circulation. 2000;101:862–868. doi: 10.1161/01.cir.101.8.862. [DOI] [PubMed] [Google Scholar]
- Monahan KD, Ray CA. Limb neurovascular control during altered otolithic input in humans. J Physiol. 2002a;538:303–308. doi: 10.1113/jphysiol.2001.013131. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Monahan KD, Ray CA. Vestibulosympathetic reflex during orthostatic challenge in aging humans. Am J Physiol Regul Integr Comp Physiol. 2002b;283:R1027–1032. doi: 10.1152/ajpregu.00298.2002. [DOI] [PubMed] [Google Scholar]
- Park J, Middlekauff HR. Altered pattern of sympathetic activity with the ovarian cycle in female smokers. Am J Physiol Heart Circ Physiol. 2009;297:H564–568. doi: 10.1152/ajpheart.01197.2008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ray CA. Effect of gender on vestibular sympathoexcitation. Am J Physiol Regul Integr Comp Physiol. 2000a;279:R1330–1333. doi: 10.1152/ajpregu.2000.279.4.R1330. [DOI] [PubMed] [Google Scholar]
- Ray CA. Interaction of the vestibular system and baroreflexes on sympathetic nerve activity in humans. Am J Physiol Heart Circ Physiol. 2000b;279:H2399–2404. doi: 10.1152/ajpheart.2000.279.5.H2399. [DOI] [PubMed] [Google Scholar]
- Ray CA, Hume KM, Shortt TL. Skin sympathetic outflow during head-down neck flexion in humans. Am J Physiol. 1997;273:R1142–1146. doi: 10.1152/ajpregu.1997.273.3.R1142. [DOI] [PubMed] [Google Scholar]
- Rusch NJ, Shepherd JT, Webb RC, Vanhoutte PM. Different behavior of the resistance vessels of the human calf and forearm during contralateral isometric exercise, mental stress, and abnormal respiratory movements. Circ Res. 1981;48:I118–130. [PubMed] [Google Scholar]
- Shortt TL, Ray CA. Sympathetic and vascular responses to head-down neck flexion in humans. Am J Physiol. 1997;272:H1780–1784. doi: 10.1152/ajpheart.1997.272.4.H1780. [DOI] [PubMed] [Google Scholar]
- Wilson TD, Cotter LA, Draper JA, Misra SP, Rice CD, Cass SP, Yates BJ. Vestibular inputs elicit patterned changes in limb blood flow in conscious cats. J Physiol. 2006;575:671–684. doi: 10.1113/jphysiol.2006.112904. [DOI] [PMC free article] [PubMed] [Google Scholar]