

Abstract
Blindsight is the rare and paradoxical ability of some human subjects with occipital lobe brain damage to discriminate unseen stimuli in their clinically blind field defects when forced-choice procedures are used, implying that lesions of striate cortex produce a sharp dissociation between visual performance and visual awareness. Skeptics have argued that this is no different from the behavior of normal subjects at the lower limits of conscious vision, at which such dissociations could arise trivially by using different response criteria during clinical and forced-choice tests. We tested this claim explicitly by measuring the sensitivity of a hemianopic patient independently of his response criterion in yes-no and forced-choice detection tasks with the same stimulus and found that, unlike normal controls, his sensitivity was significantly higher during the forced-choice task. Thus, the dissociation by which blindsight is defined is not simply due to a difference in the patients’ response bias between the two paradigms. This result implies that blindsight is unlike normal, near-threshold vision and that information about the stimulus is processed in blindsighted patients in an unusual way.
The striate cortex is the initial cortical locus for visual processing in the brain. Destruction or denervation of part of this area produces a scotoma in the corresponding part of the visual field (1), but forced-choice tests have revealed that some patients retain an ability to detect and discriminate stimuli presented within their scotomata despite the fact that they deny seeing them (2–4). This phenomenon is called blindsight. Research on monkeys, as well as brain-damaged patients, suggests that visual sensitivity in the scotoma could be mediated by parallel projections involving the superior colliculus, the lateral geniculate nucleus, the pulvinar nucleus, and the extrastriate cortex (5–12). Blindsight is of special interest to cognitive neuroscientists and philosophers because it implies that conscious awareness of visual stimuli requires an intact striate cortex and that cortical activity in other visual areas is by itself incapable of sustaining a conscious representation, with the possible exception of moving stimuli (13).
By definition, blindsight is a dissociation between judgements about visual stimuli presented in two different tasks, namely the yes-no (yn) and forced-choice paradigms. In the yn paradigm, one of two possible stimuli is presented on each trial, and the patient must judge which was presented. In effect, this is the basis of perimetry because patients must indicate whether or not they see a stimulus presented at systematically sampled points in the visual field. (The fact that a visual stimulus is presented on every trial in clinical perimetry does not affect this distinction between the two kinds of task.) In the two alternative forced-choice (2afc) paradigm, each of two different stimuli is presented every trial, and the patient must judge which interval contained a specified stimulus. The dissociation can be dramatic, with performance in the same patient varying between ≈50% correct with the yn paradigm and ≈90% correct with the 2afc paradigm, despite emphatic denials of visual experience on the part of the patient.
Although these procedural criteria seem straightforward, skeptics have argued against a distinction between blindsight and normal, near-threshold vision because, in both cases, apparent dissociations could arise from using different response criteria in the two kinds of task (14). This claim is important because it implies that nothing can be learned about the mechanisms underlying visual awareness by studying patients with blindsight that could not be learned from studying normal subjects, an assumption that underlies recent investigations of “blindsight” in normal subjects (15–17).
According to signal detection theory (SDT) (18), subjects’ judgement depends not only on their sensitivity to differences between the stimuli but also on their response criterion (also called bias), which is the tendency to select one or other stimulus independently of sensitivity. The effect of response criterion on performance depends on the measure of sensitivity used. In SDT, sensitivity is represented by a statistic (d′) computed so as to be independent of response criterion. But the measure usually used is percentage correct, which reflects sensitivity reliably only in the absence of any response bias, to the extent that a subject with fixed sensitivity to a given stimulus could score anything from 50 to 95% correct, depending on the subject’s response criterion (Fig. 1). The problem is that responding in 2afc tasks tends always to be bias-free whereas responding in yn tasks does not, raising the possibility that the much studied dissociation between performance in perimetry and in forced-choice tests in patients with blindsight could be accounted for simply by the subjects’ use of consistently different response criteria in the two kinds of task (14). This has never been tested explicitly. We therefore compared directly the sensitivity of a blindsighted hemianopic subject, measured independently of response criterion, when detecting identical stimuli during yn tasks and 2afc tasks.
Figure 1.

Signal detection. (Top left) Two standard normal distributions represent a subject’s internal (neural) signals associated with absence of a stimulus (S−, left) and presence of a stimulus (S+, right). The subject must chose a criterion above which he should respond “S+” and below which he should respond “S− .” As the distributions overlap, judgements based on any criterion will result in some misidentifications, yielding some errors (false alarms and misses) as well as correct responses (hits and correct rejections). The subject’s sensitivity, d′, in units of SD, is given by d′ = z(H) − z(F) where hit rate, H, = number of hits/(number of hits + misses) and false alarm rate, F, = number of false alarms/(number of false alarms + correct rejections) and z is the inverse of the normal distribution function. When the subject cannot discriminate at all, H = F and d′ = 0. The subject’s criterion, c, is given by c = −0.5 [z(H) + z(F)] and is equal to 0 when false alarm and miss (M) rates are equal, negative when F > M, and positive when F < M. Thus, c is a measure of response bias, the tendency of the subject to say S+ irrespective of the actual number of S+s presented. (Top right) The relation between sensitivity and response bias, or ROC, represented as plots of hit rate vs. false alarm rate. Curves deviating from the major diagonal represent lines of isosensitivity, which describe the relation between hit and false alarm rates as bias changes at constant sensitivity. Curves deviating from the minor diagonal represent lines of isobias across the range of sensitivities. (Bottom) Variation of percentage correct with hit rate (left) and response criterion (right) at various fixed sensitivities calculated from SDT given that S+ and S− have equal probabilities of presentation. The latter graph is symmetric about the c = 0 line. The graphs show how the performance of a subject with constant sensitivity could vary between 50 and 95% correct depending on the subject’s response criterion.
METHODS
The subject was a 40-year-old man, G.Y., who had incurred, at the age of 8 years, a unilateral lesion in the left striate cortex that resulted in a right homonymous hemianopia with macular sparing extending 3.5° into the otherwise blind hemifield (19). Residual visual sensitivity within his scotoma enables him to detect, localize, and discriminate transient stimuli presented in his blind hemifield (11, 19–24), many of which he is unaware of, as in blindsight (25). In the present experiment, we required him simply to detect the presence or absence of visual targets presented in his scotoma during yn and 2afc tests.
The stimuli were static, vertical, black-and-white, square-wave gratings with a spatial frequency of 0.25 cycle per degree and a mean luminance of 5.0 cd/m2; the stimuli were 20° × 20° in size and were displayed for 200 ms against a background of 5.0 cd/m2 at four contrasts (0.65, 0.75, 0.85, and 0.95). They were generated on an SVGA graphics board with a spatial resolution of 1270 × 780 pixels and 256 gray levels per pixel, displayed with γ correction in the center of a 15-inch RGB color monitor (Chuntex Electronics model 1565, Taiwan), and presented monocularly to the upper quadrant of the subject’s right, blind hemifield, centered 30° to the right of the vertical meridian and 12° above the horizontal meridian at a distance of 28.5 cm. The sighted left visual field was filled with a white background that reflected ambient light at an intensity of 5.0 cd/m2.
Each temporal interval in which a stimulus was displayed consisted of an auditory signal (200 ms), followed after 100 ms by a target (grating; 200 ms) or blank, and lasted for 2000 ms. y/n trials consisted of just one such interval whereas 2afc trials consisted of two contiguous intervals. The end of a trial was indicated by an auditory signal (200 ms) 1 octave higher, after which the subject responded with “yes” or “no” to indicate whether or not a target had been presented in the yn task or “first” or “second” to indicate in which interval the target been presented in the 2afc task, followed by a rating to indicate his confidence in the correctness of his response on a scale of 1 (least) to 4 (most). Because each decision (“yes” or “no” or “first” or “second”) had four ratings associated with it, there were eight permissible responses altogether. In some conditions, no responses were made in the more extreme categories of response. The subject was not specifically required to rate his awareness (visual or otherwise) of the stimulus. On several occasions, during pauses, G.Y. responded to questions about his experience by saying that he did not see the targets.
For each condition, there were 1000 trials, consisting of 250 trials with each target contrast, randomly interleaved, in which the ratio of targets to blanks was exactly 1.0. Trials were presented in blocks of 200, with short breaks (1–2 minutes) within them after every 50 trials and longer breaks (20–60 minutes) between them.
Testing was carried out under mesopic conditions. The subject sat at a table with his head supported by a chin rest and forehead restraint and gazed at a fixation spot while stimuli were presented to his right eye, his left eye being occluded with a light-tight patch. Fixation was monitored with closed-circuit TV, and a trial was aborted if the subject moved his eye at all. In practice, this occurred on fewer than 0.5% of trials. Trials were initiated from, and responses were typed into, the keyboard of a microcomputer by an investigator who could see the subject’s eye on the closed-circuit monitor but could not see the stimulus being displayed.
To control for light scatter, two kinds of tests were performed in which an additional half-patch was used to occlude the blind hemiretina of the tested eye. The first measured the contrast threshold at which the stimulus could be detected with 2afc responding and an adaptive thresholding algorithm. The program consistently moved to maximum contrast and stayed there. Second, 50 2afc detection trials were given with stimulus contrast set to maximum. Again, no significant sensitivity was found (score = 27/50, d′ = 0.142, c = −0.05, χ2 = 0.311, df = 1, not significant). This result indicates that there was insufficient extraocular light scatter associated with the targets for detection by the intact hemiretina to account for the subject’s detection when the blind hemiretina was unoccluded in this experiment. This result is as expected if the mean luminance of the gratings matched the luminance of the background.
Three male control subjects (two naïve subjects, one author) also were tested in the yn and 2afc detection tasks. The apparatus and procedures were similar, except that the range of contrasts used was necessarily much lower (0.005–0.015). The stimuli were generated with a VSG2/3 stimulus generator (Cambridge Research Systems, Kent, U.K.) with a resolution of 1024 × 768 pixels and 4096 gray levels.
RESULTS
Responses were sorted by contrast, and condition and receiver operating characteristics (ROCs: graphs of hit rate vs. false alarm rate) were fitted to the data using a maximum-likelihood algorithm (26, 27). ROCs were fitted to 2afc responses by casting them as responses to alternative pairs of presentations, <Target, Blank> or <Blank, Target>, like a yn design (28).The ROC parameters are presented in Table 1, and the resultant graphs are shown in Fig. 2.
Table 1.
ROC parameters for yn and 2afc trials sorted by subject and contrast
| Subject | Contrast | Task | Trials | A | sA | B | sB | P(B) | da | sda | Z | P(Z) |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| G.Y. (hemianope) | 0.95 | yn | 250 | 1.570 | 0.203 | 1.300 | 0.176 | NS | 1.354 | 0.146 | 3.729 | *** |
| 2afc | 250 | 2.623 | 0.389 | 0.450 | 0.179 | NS | 2.392 | 0.237 | ||||
| 0.85 | yn | 250 | 1.247 | 0.158 | 0.927 | 0.117 | NS | 1.293 | 0.144 | 3.092 | *** | |
| 2afc | 250 | 2.570 | 0.324 | 0.896 | 0.191 | NS | 1.914 | 0.140 | ||||
| 0.75 | yn | 250 | 0.947 | 0.152 | 1.086 | 0.130 | NS | 0.908 | 0.137 | 4.707 | *** | |
| 2afc | 250 | 2.401 | 0.286 | 0.872 | 0.175 | NS | 1.810 | 0.134 | ||||
| 0.65 | yn | 250 | 0.855 | 0.146 | 1.052 | 0.124 | NS | 0.833 | 0.137 | 4.287 | *** | |
| 2afc | 250 | 2.203 | 0.254 | 0.909 | 0.165 | NS | 1.631 | 0.126 | ||||
| M.F. (control) | 0.014 | yn | 250 | 1.581 | 0.211 | 1.356 | 0.184 | NS | 1.327 | 0.147 | −1.024 | NS |
| 2afc | 250 | 1.568 | 0.185 | 0.947 | 0.130 | NS | 1.139 | 0.110 | ||||
| 0.011 | yn | 250 | 1.713 | 0.244 | 1.600 | 0.226 | *** | 1.285 | 0.148 | 0.154 | NS | |
| 2afc | 250 | 1.843 | 0.215 | 0.982 | 0.154 | NS | 1.314 | 0.117 | ||||
| 0.008 | yn | 250 | 1.916 | 0.271 | 1.701 | 0.256 | *** | 1.373 | 0.152 | 0.308 | NS | |
| 2afc | 250 | 1.541 | 0.185 | 0.994 | 0.134 | NS | 1.093 | 0.109 | ||||
| 0.005 | yn | 250 | 2.077 | 0.308 | 1.883 | 0.288 | *** | 1.378 | 0.153 | −1.260 | NS | |
| 2afc | 250 | 1.500 | 0.172 | 0.856 | 0.117 | NS | 1.140 | 0.110 | ||||
| S.R. (control) | 0.011 | yn | 250 | 2.493 | 0.327 | 1.157 | 0.218 | NS | 2.306 | 0.191 | 0.228 | NS |
| 2afc | 250 | 3.177 | 0.609 | 0.896 | 0.315 | NS | 2.367 | 0.187 | ||||
| 0.010 | yn | 250 | 3.208 | 0.481 | 1.818 | 0.349 | NS | 2.187 | 0.187 | −0.004 | NS | |
| 2afc | 250 | 2.991 | 0.491 | 0.934 | 0.277 | NS | 2.186 | 0.165 | ||||
| 0.009 | yn | 250 | −0.074 | 0.147 | 1.111 | 0.135 | NS | −0.069 | 0.138 | 0.308 | NS | |
| 2afc | 250 | 0.273 | 0.174 | 1.277 | 0.172 | NS | 0.168 | 0.106 | ||||
| 0.008 | yn | 250 | 0.247 | 0.143 | 1.072 | 0.124 | NS | 0.238 | 0.137 | −1.260 | NS | |
| 2afc | 250 | 0.423 | 0.156 | 1.024 | 0.126 | NS | 0.295 | 0.105 | ||||
| P.A. (control) | 0.020 | yn | 250 | 5.349 | – | 1.895 | – | – | 3.53 | – | – | – |
| 2afc | 250 | 2.350 | – | 0.065 | – | – | 1.82 | – | ||||
| 0.015 | yn | 250 | 7.861 | – | 1.100 | – | – | 7.479 | – | – | – | |
| 2afc | 250 | 3.313 | – | 0.895 | – | – | 1.868 | |||||
| 0.010 | yn | 250 | 0.909 | 0.155 | 1.155 | 0.139 | NS | 0.842 | 0.137 | 1.912 | * | |
| 2afc | 250 | 1.612 | 0.197 | 0.924 | 0.143 | NS | 1.184 | 0.115 | ||||
| 0.005 | yn | 250 | 1.154 | 0.177 | 1.292 | 0.168 | NS | 0.999 | 0.141 | 0.045 | NS | |
| 2afc | 250 | 1.288 | 0.159 | 0.797 | 0.111 | NS | 1.007 | 0.109 |
A, y intercept of ROC plotted in z coordinates, equal to d′ when variances of the two underlying distributions are equal; sA, SD of A; B, slope of ROC plotted in z coordinates, equivalent to the ratio of the variances of the two underlying distributions; sB, SD of B; P(B), probability of B deviating from a value of 1.0 by chance; da, sensitivity index, assuming unequal variance of underlying distributions; sda, SD of da; Z, difference between da(yn) and da(2afc) expressed as a normal deviate (see ref. 30); and P, probability of the difference between da(yn) and da(2afc) arising by chance (∗, P < 0.05; ∗∗, P < 0.01; ∗∗∗, P < 0.001). Parameters are presented only if ROCs could be fitted adequately at the P < 0.01 level.
Figure 2.

ROCs fitted to ratings assigned by the hemianopic subject to stimuli of various contrasts presented in his scotoma during yn (black circles) and 2afc (white circles) detection tasks.
In subject G.Y.’s case, all fits were adequate at the P < 0.05 level of significance, except for the curve for the yn condition at the highest contrast (0.95), which was adequate at the P < 0.01 level. A striking feature of these ROCs is that their slopes in z-transformed coordinates (parameter B in Table 1) were all close to, and not significantly different from, unity (with exception of the curve already mentioned), implying that the variances of the distributions underlying the detection of blanks and targets were equal.
Sensitivity was measured as da, a variant of d′ that takes into account nonunit slopes of z-transformed ROCs (equivalent to the difference between the distribution means divided by the root–mean–square average of the SDs) (29), and da(2afc) was adjusted by a factor of 1/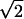 , as required by SDT, to take account of the fact that there are two opportunities to decide whether a target was presented in 2afc. Sensitivity is plotted as a function of target contrast in both kinds of task in Fig. 3. G.Y.’s sensitivity in the 2afc task was significantly greater than in the yn task (P ≪ 0.001) at every contrast tested. For responses pooled across all contrasts, the magnitude of the difference between da(yn) (1.08) and da(2afc) (1.91) measured in his scotoma was ≈2-fold. The sensitivity of the control subjects was not significantly different between the two kinds of task at any contrast.
, as required by SDT, to take account of the fact that there are two opportunities to decide whether a target was presented in 2afc. Sensitivity is plotted as a function of target contrast in both kinds of task in Fig. 3. G.Y.’s sensitivity in the 2afc task was significantly greater than in the yn task (P ≪ 0.001) at every contrast tested. For responses pooled across all contrasts, the magnitude of the difference between da(yn) (1.08) and da(2afc) (1.91) measured in his scotoma was ≈2-fold. The sensitivity of the control subjects was not significantly different between the two kinds of task at any contrast.
Figure 3.

Sensitivity as a function of stimulus contrast for yn (black circles) and 2afc (white circles) detection tasks in the hemianopic subject and three control subjects. Data were plotted only if ROCs could be fitted adequately to responses at the P < 0.01 level. Error bars indicate 95% confidence limits. Note the different contrast scale for G.Y. and the control subjects.
DISCUSSION
As emphasized by Campion et al. (14) and illustrated in Fig. 1, differences in response bias alone between the yn and 2afc paradigms could potentially account for the entire dissociation on which blindsight depends. It is therefore surprising that SDT, which provides criterion-free measures of sensitivity, has rarely been applied in this field. Previously, Stoerig et al. (31) used SDT to analyze residual vision in a hemianopic subject tested with a “forced guessing” task (essentially a yn task) in which the probability of target and blank trials was varied systematically and widely among sessions to alter the subject’s expectations and therefore his bias. They found that the slopes of the ROCs were significantly >1.0, and they concluded that the SDT could not appropriately be applied to the data and advocated the use of nonparametric analysis, which was adopted in related studies (e.g., ref. 32). The present results are strikingly different, in that every ROC fitted bar one had a slope that was close to, and not significantly greater than, 1.0 in accordance with the equal variance assumption of basic SDT. The difference is probably caused by the ways in which the ROC points were generated. The former ones were obtained from single, large blocks of trials per ROC point per stimulus, with response bias being manipulated by altering the presentation probabilities of targets and blanks and with testing spread over several days. In contrast, we used a relatively economical rating scale method to measure sensitivity to all stimuli at all response criteria in a randomized design to minimize the effects on sensitivity of fluctuations in motivation and fatigue.
This is the first time SDT has been used to measure, independently of response bias, visual sensitivity to stimuli presented in the scotoma with both yn and 2afc detection paradigms in the same hemianopic subject.§ The result was clear. His sensitivity was significantly higher during the 2afc task, and the difference was ≈2-fold (Fig. 3). Because, empirically, 2afc detection tasks yield identical values of sensitivity as do yn tasks in normal subjects, once scaled by a factor of 1/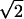 in accordance with SDT (as confirmed with control subjects under the present conditions), it follows that the hemianopic patient’s residual vision is unlike normal, near-threshold vision and that his brain processes information about the visual stimulus in his scotoma in an unusual way. This may explain why it is so difficult to reproduce blindsight-like dissociations in normal subjects (17).
in accordance with SDT (as confirmed with control subjects under the present conditions), it follows that the hemianopic patient’s residual vision is unlike normal, near-threshold vision and that his brain processes information about the visual stimulus in his scotoma in an unusual way. This may explain why it is so difficult to reproduce blindsight-like dissociations in normal subjects (17).
One possible explanation for the dissociation of sensitivity in blindsight is that the execution of yn and 2afc tasks might depend on different neural pathways, much as the quality of blindsight can depend on whether the subject’s response is verbal or reaching and grasping (33). For example, in the context of the distinction between “dorsal” occipito-parietal and “ventral” occipito-temporal cortical processing streams as specializations for spatial localization and object recognition (34) or perception and action (35), dorsal stream visual areas such as the middle temporal area are more strongly activated by visual stimuli in the absence of striate cortex than are ventral areas such as the inferotemporal cortex, and they may receive significantly more substantial subcortical input from the tectopulvinar pathway, on which their activation depends (7, 8). Thus, even a slight difference in the extent to which the two tasks activate the dorsal pathway could cause one to yield a greater estimate of visual sensitivity in the scotoma than the other. An alternative explanation could be related to perceptual memory. An observer’s ability to detect targets at low threshold in yn tasks may depend to some extent on comparing the present stimulus with an internal standard (a model of the target in visual memory). An abnormally depressed sensitivity in yn tasks, such as found here in the hemianopic subject, could indicate that a subject without striate cortex may be less able to visualize the target from memory and therefore less able to perform the discrimination.
Acknowledgments
We thank G.Y. for his cooperation and Penny Brown, Mazyar Fallah, Dr. Charles Gross, Dr. Sheila King, and Scott Regenbogen for their help. This work was supported by the Medical Research Council and a Network Grant from the Oxford McDonnell-Pew Centre for Cognitive Neuroscience.
ABBREVIATIONS
- yn
yes-no
- 2afc
two alternative forced-choice
- SDT
signal detection theory
- ROC
receiver operating characteristics
Footnotes
We planned also to test his sighted field in this way, but he was not available for the additional lengthy testing required.
References
- 1.Holmes G. Proc R Soc Lond B. 1945;132:348–361. [Google Scholar]
- 2.Pöppel E, Held R, Frost D. Nature (London) 1973;243:295–296. doi: 10.1038/243295a0. [DOI] [PubMed] [Google Scholar]
- 3.Weiskrantz L, Warrington E K, Sanders M D, Marshall J. Brain. 1974;97:709–728. doi: 10.1093/brain/97.1.709. [DOI] [PubMed] [Google Scholar]
- 4.Weiskrantz L. Blindsight: A Case Study and Its Implications. Oxford: Oxford Univ. Press; 1986. [Google Scholar]
- 5.Mohler C W, Wurtz R H. J Neurophsyiol. 1977;40:74–94. doi: 10.1152/jn.1977.40.1.74. [DOI] [PubMed] [Google Scholar]
- 6.Rodman H R, Gross C G, Albright T D. J Neurosci. 1989;9:2033–2050. doi: 10.1523/JNEUROSCI.09-06-02033.1989. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Rodman H R, Gross C G, Albright T D. J Neurosci. 1990;10:1154–1164. doi: 10.1523/JNEUROSCI.10-04-01154.1990. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Gross C G. Neuropsychologia. 1991;29:497–515. doi: 10.1016/0028-3932(91)90007-u. [DOI] [PubMed] [Google Scholar]
- 9.Cowey A, Stoerig P. Trends Neurosci. 1991;14:140–145. doi: 10.1016/0166-2236(91)90085-9. [DOI] [PubMed] [Google Scholar]
- 10.Stoerig P, Cowey A. Brain. 1997;20:535–559. doi: 10.1093/brain/120.3.535. [DOI] [PubMed] [Google Scholar]
- 11.King S M, Azzopardi P, Cowey A, Oxbury J, Oxbury S. Visual Neurosci. 1996;13:1–13. doi: 10.1017/s0952523800007082. [DOI] [PubMed] [Google Scholar]
- 12.Azzopardi P, King S M, Cowey A. Soc Neurosci Abstr. 1996;22:399. [Google Scholar]
- 13.Barbur J L, Watson J D G, Frackowiak R S J, Zeki S. Brain. 1993;116:1293–1302. doi: 10.1093/brain/116.6.1293. [DOI] [PubMed] [Google Scholar]
- 14.Campion J, Latto R M, Smith Y M. Behav Brain Sci. 1983;3:423–486. [Google Scholar]
- 15.Meeres S L, Graves R E. Neuropsychologia. 1990;28:1231–1237. doi: 10.1016/0028-3932(90)90039-q. [DOI] [PubMed] [Google Scholar]
- 16.Kolb F C, Braun J. Nature (London) 1995;377:293–294. doi: 10.1038/377336a0. [DOI] [PubMed] [Google Scholar]
- 17.Morgan M J, Mason A J S, Solomon J A. Nature (London) 1997;385:401–402. doi: 10.1038/385401b0. [DOI] [PubMed] [Google Scholar]
- 18.Green D M, Swets J A. Signal Detection Theory and Psychophysics. New York: Wiley; 1996. [Google Scholar]
- 19.Barbur J L, Ruddock K, Waterfield V A. Brain. 1980;102:905–928. doi: 10.1093/brain/103.4.905. [DOI] [PubMed] [Google Scholar]
- 20.Blythe I M, Bromley J M, Kennard C, Ruddock K H. Nature (London) 1986;320:619–621. doi: 10.1038/320619a0. [DOI] [PubMed] [Google Scholar]
- 21.Blythe I M, Kennard C, Ruddock K H. Brain. 1987;110:887–905. doi: 10.1093/brain/110.4.887. [DOI] [PubMed] [Google Scholar]
- 22.Weiskrantz L, Harlow A J, Barbur J L. Brain. 1991;114:2269–2282. doi: 10.1093/brain/114.5.2269. [DOI] [PubMed] [Google Scholar]
- 23.Barbur J L, Harlow A J, Weiskrantz L. Proc R Soc Lond B. 1994;343:157–166. doi: 10.1098/rstb.1994.0018. [DOI] [PubMed] [Google Scholar]
- 24.Brent P J, Kennard C, Ruddock K H. Proc R Soc Lond B. 1994;256:219–225. doi: 10.1098/rspb.1994.0073. [DOI] [PubMed] [Google Scholar]
- 25.Weiskrantz L, Barbur J L, Sahrie A. Proc Natl Acad Sci USA. 1995;92:6122–6126. doi: 10.1073/pnas.92.13.6122. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Dorfman D D, Alf E., Jr J Math Psychol. 1969;6:487–496. [Google Scholar]
- 27.Swets J A, Picket R M. Evaluation of Diagnostic Systems: Methods from Signal Detection Theory. New York: Academic; 1982. [Google Scholar]
- 28.Schulman A I, Mitchell R R. J Acoust Soc Am. 1966;40:473–477. doi: 10.1121/1.1910098. [DOI] [PubMed] [Google Scholar]
- 29.Simpson A J, Fitter M J. Psychol Bull. 1973;80:481–488. [Google Scholar]
- 30.Marascuilo L A. Psychometrika. 1970;35:237–243. [Google Scholar]
- 31.Stoerig P, Hübner M, Pöppel E. Neuropsychologia. 1985;23:589–599. doi: 10.1016/0028-3932(85)90061-2. [DOI] [PubMed] [Google Scholar]
- 32.Stoerig P, Pöppel E. Exp Brain Res. 1986;64:469–475. doi: 10.1007/BF00340483. [DOI] [PubMed] [Google Scholar]
- 33.Perenin M-T, Rossetti Y. Neuroreport. 1996;7:793–797. doi: 10.1097/00001756-199602290-00027. [DOI] [PubMed] [Google Scholar]
- 34.Ungerleider L G, Mishkin M. In: Analysis of Behavior. Ingle D J, Goodale M A, Mansfield R J W, editors. Cambridge, MA: MIT Press; 1982. pp. 549–586. [Google Scholar]
- 35.Milner A D, Goodale M A. The Visual Brain in Action. Oxford: Oxford Univ. Press; 1995. [Google Scholar]