

Abstract
Relationships were examined between spatial learning and hippocampal concentrations of the α, β2, and γ isoforms of protein kinase C (PKC), an enzyme implicated in neuronal plasticity and memory formation. Concentrations of PKC were determined for individual 6-month-old (n = 13) and 24-month-old (n = 27) male Long–Evans rats trained in the water maze on a standard place-learning task and a transfer task designed for rapid acquisition. The results showed significant relationships between spatial learning and the amount of PKC among individual subjects, and those relationships differed according to age, isoform, and subcellular fraction. Among 6-month-old rats, those with the best spatial memory were those with the highest concentrations of PKCγ in the particulate fraction and of PKCβ2 in the soluble fraction. Aged rats had increased hippocampal PKCγ concentrations in both subcellular fractions in comparison with young rats, and memory impairment was correlated with higher PKCγ concentrations in the soluble fraction. No age difference or correlations with behavior were found for concentrations of PKCγ in a comparison structure, the neostriatum, or for PKCα in the hippocampus. Relationships between spatial learning and hippocampal concentrations of calcium-dependent PKC are isoform-specific. Moreover, age-related spatial memory impairment is associated with altered subcellular concentrations of PKCγ and may be indicative of deficient signal transduction and neuronal plasticity in the hippocampal formation.
Protein kinase C (PKC) comprises a family of enzymes activated indirectly by G-protein-coupled receptor stimulation. Multiple lines of research implicate PKC in activity-dependent neuronal plasticity and memory formation. Among these, PKC activity is important for induction and maintenance phases of long-term potentiation, a putative cellular mechanism of memory formation (1–7). Administration of PKC inhibitors impairs memory formation (8–10), whereas agents that increase PKC activity enhance memory formation (9–12). Training is associated with translocation of PKC from the soluble to the particulate subcellular compartments and may mediate neuronal plasticity necessary for learning (refs. 13–17, but see ref. 18).
In addition to evidence that PKC is necessary for memory formation among young animals, other reports suggest that alterations in PKC contribute to age-related memory deficits. For example, translocation of PKC from the soluble to the particulate fraction is decreased (19) and phosphorylation of the protein F1/B-50/GAP-43, a substrate of PKC related to adult neuronal plasticity and memory formation, is reduced in the hippocampus of aged rats (20, 21). The measures of PKC activity used in those studies cannot distinguish among the various isoforms of PKC. In recent research that used antisera selective for individual PKC isoforms, PKCγ was the only calcium-dependent isoform reported to show age-related changes in hippocampal concentration, with a significantly higher amount of this isoform in the particulate, or membrane-bound, fraction than in young rats (19). The current study examined the subcellular concentrations of PKCγ in the hippocampus of individual 6- and 24-month-old rats after spatial memory training. Two additional calcium-dependent isoforms—PKCα and PKCβ2—were measured in the hippocampus, and PKCγ was measured in the neostriatum of the same subjects for comparison. The study was designed to assess if spatial learning on a task that depends on the integrity of the hippocampal formation is associated with the subcellular concentrations of these isoforms of PKC in the hippocampus.
MATERIALS AND METHODS
Male Long–Evans rats (Charles River Breeding Laboratories) were obtained either at 4 months of age and tested at 6 months of age (young group) or obtained as retired breeders at 9 months of age and tested at 24 months of age (aged group). All rats were provided water and food ad libitum and maintained in a pathogen-free vivarium at 25°C on a 12-hr light-dark cycle with lights on at 0700 hr.
Two white circular tanks (1.8 m × 0.6 m) were maintained in separate rooms. Each was filled to 35 cm with 27°C water and made opaque with white tempera paint (150 ml). In the tank used for the place-learning task, white cloth extramaze cues were attached to a black curtain surrounding the maze and remained in a fixed position throughout training. A white retractable platform (12-cm diameter) was situated near the center of one of four maze quadrants and 1 cm below the water surface. A black platform extending 2 cm above the surface was used during cue training. For the place-learning transfer task, black cloth extramaze cues were attached to a white curtain surrounding the second maze. All measures of performance in the apparatus were obtained with a computer tracking system (HVS Imaging, Hampton, UK).
In the initial phase of place training in the first maze, rats were placed in the pool at one of four randomly chosen start locations spaced equally around the perimeter and allowed 90 sec to swim to an escape platform located in a constant position and camouflaged 1 cm below the opaque water surface. If rats did not find the platform within 90 sec, then they were led to it by the experimenter and allowed to remain there for 15 sec. Rats received three trials per day for 8 consecutive days. On every sixth trial (probe trial), the platform was retracted to the bottom of the pool for 30 sec to measure spatial bias relative to the platform position. Rats received six additional cue-training trials in the same maze on the ninth day of the protocol, during which a visible platform was positioned in one of the four pool quadrants for each trial. Cumulative search error was computed by sampling 10 times per sec the rat’s distance from the escape platform and averaging these values into 1-sec blocks. The blocks were summed, and the proximity score that would result from optimal performance was subtracted to correct for different start locations. The learning index is the sum of weighted proximity scores measured during probe trials; low scores reflect search near the escape platform, whereas high scores reflect search farther away from the target. Pathlength is the total distance swum from the start location to the target, and latency is the total duration of the trial from when the rat was placed in the water until it located the escape platform (for further description of behavioral measures, see ref. 22).
Two weeks after training was completed on the place-learning task, rats were trained in the second water maze of identical dimensions but in a new spatial environment with a different configuration of spatial cues. The behavioral procedures for this task were designed to provide a temporally defined episode of spatial learning before assessment of PKC concentrations. Trials were conducted as described for the place-learning task, but each rat received one session of six trials followed 30 min later by a retention test that consisted of a single probe trial.
Rats were killed by decapitation immediately after the retention test of the transfer training protocol, and the entire hippocampus was dissected bilaterally on an ice-cold stage and homogenized as described below (23). The dorsal anterior neostriatum was dissected as described in detail elsewhere (24). Individual tissue samples were weighed and then homogenized in 5 vol of ice-cold buffer [20 mM Tris⋅HCl, pH 7.4/0.25 M sucrose/2 mM EDTA/10 mM EGTA/5 mM DTT/0.234 mM leupeptin/1 mM phenylmethylsulfonyl fluoride]. Sample homogenates were centrifuged at 100,000 × g for 60 min at 4°C. The supernatant was removed from each sample, and an aliquot was taken for determination of total protein concentration (25). The remaining supernatant was mixed (1:1) with 2× SDS sample buffer (0.2 M SDS/20% glycerol/10% β-mercaptoethanol/0.004% bromophenol blue/1.5% Tris) and heated in a boiling water bath for 7 min. The boiled samples, comprising the soluble fraction, were frozen at −80°C until Western blot analysis. The pellets were resuspended in homogenizing buffer containing 0.1% Triton X-100, incubated for 60 min at 4°C, and centrifuged at 100,000 × g for 60 min at 4°C. The supernatant was removed and vortexed, and an aliquot was taken for protein determination. The remaining supernatant was mixed (1:1) with 2× SDS sample buffer and heated as described above. The boiled samples, comprising the particulate fraction, were frozen at −80°C until Western blot analysis. The hippocampi and anterior dorsal neostriata were dissected from the brains of an additional nine naive, 6-month-old rats. Tissues were pooled for each brain region, subcellular fractions were obtained as described above, and these samples were loaded onto each gel to serve as control tissue standards during subsequent Western blot analyses.
Tissue extracts were subjected to 8% SDS/PAGE. Four gels were run simultaneously containing subcellular fractions from naive 6-month-old rats (control standards) and from rats used in the behavioral study. After separation, the samples were transferred electrophoretically overnight at 4°C to polyvinylidene fluoride immobilon membranes (Millipore). Membranes were washed at room temperature three times for 5 min each in PBS, then three times for 15 min each in a solution containing 5% nonfat dry milk, 0.03% Tween-20, PBS (NFDM-Tween-PBS). Membranes were then incubated with polyclonal antisera to PKCα (1:10,000), PKCβ2 (1:5,000), or PKCγ (1:5,000) for 2 hr at room temperature in NFDM-Tween-PBS (26). Preliminary experiments showed that hippocampal concentrations of PKCβ1 were very low, and this isoform was not tested further (data not shown). Membranes then were washed as described above with PBS and NFDM-Tween-PBS and incubated with peroxidase-labeled secondary antisera (Kierkegard & Perry Laboratories) at a 1:20,000 dilution in NFDM-Tween-PBS for 2 hr at room temperature. The membranes then were washed extensively with a 0.1% Tween-20 PBS solution, incubated for 5 min in chemiluminescent reagents (Pierce), exposed to hyperfilm for intervals ranging from seconds to minutes, and developed.
Films were digitized (Imaging Research, St. Catherines, ON, Canada), and integrated measures comprised of optical density were multiplied by the target area in pixels were recorded for each band (Fig. 1). Linear functions of increasing protein concentrations were produced by using this measure in the range of protein concentrations tested (see Fig. 1B). The integrated measures from a set of tissue standards run with each gel were plotted, and values for the individual samples were extrapolated from the standard curves. ANOVA was used to examine acquisition of place learning and concentrations of PKC isoforms in the soluble and particulate fractions. All samples were tested in two independent experiments for each PKC isoform. Significant correlations were found between duplicates for each treatment condition (mean r = 0.65 ± 0.07, all P <0.05), and values used in subsequent analyses represent the mean of duplicate samples. Correlations were tested between behavioral indices of learning and protein concentrations in subcellular fractions.
Figure 1.
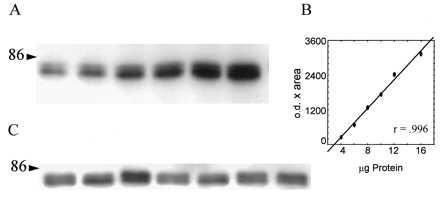
(A) Representative image of standards prepared from pooled tissue of naive, young rats, ranging from 4–16 μg of total protein and immunostained for PKCγ. Each gel was loaded with standards in this range for either soluble or particulate fractions from either the hippocampus or the neostriatum. (B) Plot and linear regression of tissue standards in A. The abscissa represents the concentration of total protein loaded in each lane, and the ordinate represents the integrated measure of band optical density multiplied by the area in number of pixels. (C) Representative image of sample lanes immunostained for PKCγ. Each lane was loaded with 10 μg of total protein from an individual rat from either the soluble or the particulate fraction of either the hippocampus or the neostriatum. Samples were pseudorandomly loaded on the gel such that each group (young, aged-unimpaired, and aged-impaired) was represented once in each group of three lanes. Ten samples were run simultaneously with six standards on each gel, and sample values were extrapolated from the corresponding standard curve.
RESULTS
Aged rats differed significantly from young rats on all measures of place learning; these included cumulative search error [F(1,38) = 25.7, P < 0.01] (see Fig. 2A), training trial pathlength [F(1,38) =7.1, P < 0.05], training trial latency [F(1,27) = 33.0, P < 0.01], and a learning index derived from multiple probe trials [F (1,38) = 21.9, P < 0.01] (see Fig. 2A Inset). In contrast, aged rats were not different from young rats during cue training on measures of cumulative search error [F(1,38) = 0.9, P > 0.05], pathlength [F(1,38) = 2.0, P > 0.05], or latency [F(1,38) = 0.4, P > 0.05], indicating that impaired place learning among aged rats was not because of sensorimotor deficits.
Figure 2.
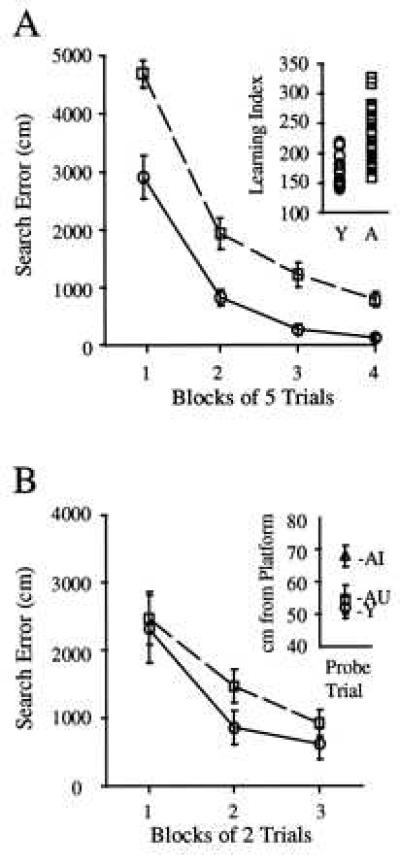
(A)Twenty-four-month-old rats (□) were significantly impaired in comparison with 6-month-old rats (○) during place learning. Aged rats (A, Inset) showed greater heterogeneity than young rats (Y). (B) There was no reliable difference between 24-month-old rats (□) and 6-month-old rats (○) during acquisition of the transfer task. During the probe trial (Inset), the subgroup of aged rats that performed outside the range of young rats on the original spatial task (AI, aged impaired; n = 15) also showed significantly less spatial bias than either aged rats that performed within the range of the young rats (AU, aged-unimpaired; n = 12) or young rats (Y, n = 13).
Aged and young rats did not differ reliably during massed training trials in the transfer task (Fig. 2B). There was an overall decrease in cumulative search error across the six training trials of the place-learning transfer task [F(5,190) = 7.39, P < 0.001]. There was neither an effect of age [F(1,38) = 1.01, P > 0.05] nor a trial x age interaction [F(5,190) = 1.26, P > 0.05] during acquisition of the transfer task. The results of the probe trial taken 30 min after the sixth training trial, however, indicated that the young rats swam significantly closer to the platform location than aged rats [F(1,38) = 4.20, P < 0.05]. Thus aged rats did not acquire and/or retain as strong a spatial bias as the young rats. Note that the individual differences in retention after transfer training coincided with subgroups of impaired and unimpaired aged rats identified in the original place-learning task (Fig. 2B, Inset).
Table 1 shows the mean (± SEM) hippocampal concentrations of the γ, α, and β2 isoforms of PKC among young and aged rats. The concentration of PKCγ was higher in aged rats in comparison with young rats [F(1,35) = 4.79, P < 0.05] whereas no significant effects of age were detected for either PKCα or PKCβ2, [F(1,30) = 0.22, F(1,38) = 0.24, respectively, P > 0.05]. An analysis of PKCγ conducted on samples from the neostriatum revealed no effect of age [F(1,28) = 0.53, P > 0.05] (data not shown).
Table 1.
Hippocampal concentrations of PKC isoforms in soluble and particulate subcellular fractions of young and aged rats
| PKC isoform | Subcellular fraction | Young rats | Aged rats |
|---|---|---|---|
| γ* | Soluble | 9.90 ± 0.37 | 10.70 ± 0.28 |
| Particulate | 8.08 ± 0.60 | 8.85 ± 0.37 | |
| α | Soluble | 7.22 ± 0.30 | 7.10 ± 0.27 |
| Particulate | 5.81 ± 0.44 | 5.44 ± 0.30 | |
| β2 | Soluble | 5.68 ± 0.39 | 6.13 ± 0.34 |
| Particulate | 4.66 ± 0.48 | 4.73 ± 0.38 |
Values represent the mean ± SEM of integrated optical density and area measures of individual samples immunostained for PKC isoforms and calibrated to standards in μg of total protein (see Fig. 1).
Aged rats had higher overall concentrations of PKCγ than young rats, but the age x subcellular fraction interaction was not significant.
The groups of young and aged rats were analyzed independently for correlations between PKC concentrations and spatial memory. For each isoform, correlations were tested between the concentrations in the soluble and particulate fractions and two behavioral measures: the graded learning index from the original place learning task and the distance from the training platform position during the transfer training probe trial. Among young rats, a significant correlation was found between the learning index and the concentrations of PKCγ in the particulate, but not the soluble, subcellular fraction (r = −0.65, P < 0.05; Fig. 3A). Young rats with the lowest learning index scores, indicating better performance, were those with the highest concentrations of PKCγ in the particulate fraction, which contains primarily membrane-bound proteins. In addition, a significant correlation was found between the learning index and PKCβ2 concentrations in the soluble, but not the particulate, fraction of young rats (r = −0.58, P < 0.05; see Fig. 3B): those with the lowest learning index scores had the highest concentrations in the soluble fraction. A similar relationship was observed between the learning index and PKCβ2 concentrations in the particulate fraction of young rats, but it was not reliable statistically (r = −0.44, P > 0.05).
Figure 3.

(A) There was a significant correlation between the learning index and the concentrations of PKCγ in the particulate fraction of young rats (r = −0.65, P < 0.05). Young rats with the lowest learning index scores, indicating the best spatial memory, had the highest concentrations of PKCγ in the particulate, or membrane-bound, fraction. (B) The learning index was correlated significantly with PKCβ2 concentrations in the soluble fraction among young rats (r = −0.58, P < 0.05). Young rats with the lowest learning index scores had the highest concentrations of PKCβ2 in the soluble fraction. (C) A significant correlation was found between the learning index and the concentrations of PKCγ in the soluble fraction of aged rats (r = 0.40, P < 0.05). Aged rats with the highest learning index scores, indicating the worst spatial memory, had the highest concentrations of PKCγ in the soluble, or cytosolic, fraction.
Among aged rats, a significant correlation was found between the learning index and the concentrations of PKCγ in the soluble, but not the particulate, fraction (r = 0.40, P < 0.05; see Fig. 3C). Aged rats with the highest learning index scores, indicating the most impaired performance, had the highest concentrations of PKCγ in the soluble fraction. The significant correlations we observed were all between the learning index derived from initial place-learning and PKC concentrations and not between performance on the single probe trial after transfer training and PKC.
DISCUSSION
Aged rats were impaired during place learning in comparison to young rats, but they also showed a broad range of spatial memory performance (see Fig. 2A, Inset). In subsequent training, young rats rapidly learned the transfer task and had robust retention on a probe trial given 30 min after training. Although aged rats did not differ significantly from young rats on transfer training trials, they had poorer retention on the probe trial. Moreover, subgroups of aged rats identified by spatial memory performance on the initial place-learning task also were identified 2 weeks later by spatial memory performance on the transfer task (see Fig. 2B, Inset). Thus individual differences in spatial memory among aged rats can be detected reliably and are stable for at least 2 weeks.
Examination of the relationship between neurobiological measures obtained in this study and behavioral performance indicated that proficiency in the hippocampal-dependent task was correlated with PKC concentrations in each age group. In all such instances, the learning index, but not the probe trial performance after transfer training, was associated with the concentrations of PKC isoforms in subcellular fractions. The sensitivity of the learning index measure may derive from the fact that it incorporates multiple assessments on probe trials that reflect both rate of learning and savings across sessions conducted over several days. By comparison, the transfer task yields a measure that comes from a single probe trial.
Among 6-month-old rats, those with the lowest learning indices, indicating the best spatial memory, were those with the highest concentrations of PKCγ and PKCβ2. The relationships, however, were restricted to different subcellular fractions; spatial memory was related to PKCγ concentrations in the particulate fraction and to PKCβ2 in the soluble fraction. The γ-isoform of PKC was implicated in spatial memory after reports that PKCγ immunoreactivity is increased in hippocampal pyramidal neurons, granule cells, and their associated dendrites in mice trained on a spatial memory task (16). PKCγ also is found in higher concentrations in the hippocampi of a strain of mice 60–90 days old known to exhibit better spatial learning than a strain that learns less readily (27). The current study, a quantitative demonstration that spatial memory among individual animals is related to concentrations of PKCγ in hippocampal subcellular fractions, provides further support for a role for the γ-isoform of PKC in memory formation. Although transgenic mice that do not express PKCγ show only mild spatial memory impairment (28), those results may be because of compensatory developmental processes in the transgenic model, e.g., recruitment of other isoforms to compensate for PKCγ deficiency (29) rather than to PKCγ serving a minor role during spatial memory formation.
Our finding that higher concentrations of PKCγ in the particulate fraction are related to better spatial memory is consistent with the view that translocation of PKC from the cytosol to neuronal membranes is an important step in its role in plasticity associated with learning. That model implies that relationships between spatial memory and subcellular PKC concentrations are because of learning-induced redistribution of PKC rather than to differences in subcellular concentrations a priori. If this were the case, then correlations between PKC concentrations and behavior would not be apparent at longer intervals between transfer training and death. At the same time, the current findings suggest that a simple translocation model may be inadequate to account for the training-related status of individual PKC isoforms; PKCβ2 concentrations in the soluble, rather than the particulate fraction, were correlated with spatial memory among young rats. This result is consistent with evidence of differences in the distribution of PKCγ and PKCβ2 after training (30).
The current study supports a role for PKC isoforms during memory formation in young rats, but it also implicates PKC in age-related memory impairment. Among aged rats, those with the highest learning index scores, indicating the most impaired performance, were those with the highest PKCγ concentrations in the soluble fraction. Aged rats also had higher concentrations of hippocampal PKCγ in the particulate fraction than young rats, but we did not find any relationship between that measure and spatial memory. We did not find increased PKCγ concentrations in a comparison structure, the neostriatum, in which other neurobiological markers were reported to show age- and memory-related changes (24, 31, 32). Reports that aging is associated with increased cytosolic calcium in hippocampal neurons (refs. 33 and 34, but see ref. 35) suggest a mechanism for the age-related increase in particulate PKCγ that we observed, as calcium is instrumental in translocation of PKCγ from cytosolic to membrane-bound states. It is unlikely, however, that translocation is the only mechanism of age-related PKC dys-regulation as we observed increases in both soluble and particulate subcellular fractions of aged rats with spatial memory impairment. It is also unlikely that increased subcellular PKC concentrations are caused by increased protein expression, because mRNA for PKC isoforms is not altered among aged rats (36, 37). Thus age-related dys-regulation of PKC most likely is because of impairment of posttranslational mechanisms. The current finding that PKCγ is elevated in the particulate fraction of aged rats, but not related to memory as it is in young rats, suggests a failure among some aged rats to uncouple inactive PKCγ from neuronal membranes. In addition, age-associated changes among phosphatases or other enzymes that dephosphorylate substrates of PKC also may be linked to PKC dys-regulation.
A recent study reported no loss of neurons in the hippocampus of aged rats from the same population used in the current investigation and including a subgroup of aged rats impaired on a hippocampal-dependent spatial memory task (38). Those findings suggested that age-related memory impairment may be because of changes in functional properties of hippocampal neurons that occur in the absence of neuron loss. The current results indicate one such mechanism by which hippocampal neuronal plasticity may be compromised with age; PKC not only is related to spatial memory among young rats, but dys-regulation of PKC is associated with spatial memory impairment among individual aged rats.
Acknowledgments
We are grateful to Robert McMahan for assistance with behavioral testing and to Jennifer Smith and Michael Ulmer for technical assistance. This research was supported by Public Health Service Grant F32-MH11337 to P.J.C., National Institute of Environmental Health Sciences intramural research program to W.C.W., and National Institute of Mental Health and National Institute on Aging Grants K05-MH01149 and PO1-AG00973 to M.G.
ABBREVIATION
- PKC
protein kinase C
References
- 1.Lovinger D M, Routtenberg A. J Physiol. 1988;400:321–333. doi: 10.1113/jphysiol.1988.sp017122. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Klann E, Chen S J, Sweatt J D. J Biol Chem. 1991;266:24253–24256. [PubMed] [Google Scholar]
- 3.Klann E, Chen S J, Sweatt J D. Proc Natl Acad Sci USA. 1993;90:8337–8341. doi: 10.1073/pnas.90.18.8337. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Wang J H, Feng D P. Proc Natl Acad Sci USA. 1992;89:2576–2580. doi: 10.1073/pnas.89.7.2576. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Colley P A, Routtenberg A. Brain Res Rev. 1993;18:115–122. doi: 10.1016/0165-0173(93)90009-o. [DOI] [PubMed] [Google Scholar]
- 6.Leahy J C, Luo Y, Kent C S, Meiri K F, Vallano M L. Neuroscience. 1993;52:563–574. doi: 10.1016/0306-4522(93)90406-6. [DOI] [PubMed] [Google Scholar]
- 7.Sacktor T C, Osten P, Valsamis H, Jiang X, Naik M U, Sublette E. Proc Natl Acad Sci USA. 1993;90:8342–8346. doi: 10.1073/pnas.90.18.8342. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Mathis C, Lehmann J, Ungerer A. Eur J Pharmacol. 1992;220:107–110. doi: 10.1016/0014-2999(92)90020-5. [DOI] [PubMed] [Google Scholar]
- 9.Serrano P A, Beniston D S, Oxonian M G, Rodriguez W A, Rosenzweig M R, Bennett E L. Behav Neural Biol. 1994;61:60–72. doi: 10.1016/s0163-1047(05)80045-7. [DOI] [PubMed] [Google Scholar]
- 10.Zhao W Q, Sedman G L, Gibbs M E, Ng K T. Behav Brain Res. 1994;60:151–160. doi: 10.1016/0166-4328(94)90142-2. [DOI] [PubMed] [Google Scholar]
- 11.Paylor R, Rudy J W, Wehner J M. Behav Brain Res. 1991;45:189–193. doi: 10.1016/s0166-4328(05)80085-3. [DOI] [PubMed] [Google Scholar]
- 12.Yang H C, Lee E H. Chin J Physiol. 1993;36:115–123. [PubMed] [Google Scholar]
- 13.Bank B, DeWeer A, Kuzirian A M, Rasmussen H, Alkon D L. Proc Natl Acad Sci USA. 1988;85:1988–1992. doi: 10.1073/pnas.85.6.1988. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Olds J L, Anderson M L, McPhie D L, Staten L D, Alkon D L. Science. 1989;245:866–869. doi: 10.1126/science.2772638. [DOI] [PubMed] [Google Scholar]
- 15.Olds J M, Golski S, McPhie D L, Olton D, Mishkin M, Alkon D L. J Neurosci. 1990;10:3707–3713. doi: 10.1523/JNEUROSCI.10-11-03707.1990. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Van der Zee E A, Compaan J C, de Boer M, Luiten P G. J Neurosci. 1992;12:4808–4815. doi: 10.1523/JNEUROSCI.12-12-04808.1992. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Van der Zee E A, Compaan J C, Bohus B, Luiten P G M. Hippocampus. 1995;5:349–362. doi: 10.1002/hipo.450050408. [DOI] [PubMed] [Google Scholar]
- 18.Spieler K, Schoch P, Martin J R, Haefely W. Pharmacol Biochem Behav. 1993;46:553–560. doi: 10.1016/0091-3057(93)90543-3. [DOI] [PubMed] [Google Scholar]
- 19.Battaini F, Elkabes S, Bergamaschi S, Ladisa V, Lucchi L, DeGraan P N E, Schuurman T, Wetsel W C, Trabucchi M, Govoni S. Neurobiol Aging. 1995;16:137–148. doi: 10.1016/0197-4580(94)00154-5. [DOI] [PubMed] [Google Scholar]
- 20.Barnes C A, Mizumori S J Y, Lovinger D M, Sheu F S, Murakami S Y, Chan S Y, Linden D J, Nelson R B, Routtenberg A. Neurobiol Aging. 1988;9:393–398. doi: 10.1016/s0197-4580(88)80086-1. [DOI] [PubMed] [Google Scholar]
- 21.Gianotti C, Porta A, De Grann P N, Oestreicher A B, Nunzi M G. Neurobiol Aging. 1993;14:401–406. doi: 10.1016/0197-4580(93)90098-v. [DOI] [PubMed] [Google Scholar]
- 22.Gallagher M, Burwell R, Burchinal M. Behav Neurosci. 1993;107:618–626. doi: 10.1037//0735-7044.107.4.618. [DOI] [PubMed] [Google Scholar]
- 23.Glowinski J, Iverson L L. J Neurochem. 1966;13:655–669. doi: 10.1111/j.1471-4159.1966.tb09873.x. [DOI] [PubMed] [Google Scholar]
- 24.Gallagher M, Burwell R D, Kodsi M H, McKinney M, Southerland S, Vella-Rountree L, Lewis M H. Neurobiol Aging. 1990;11:507–514. doi: 10.1016/0197-4580(90)90111-c. [DOI] [PubMed] [Google Scholar]
- 25.Bradford M M. Anal Biochem. 1976;72:248–254. doi: 10.1016/0003-2697(76)90527-3. [DOI] [PubMed] [Google Scholar]
- 26.Wetsel W C, Khan W A, Merchenthaler I, Rivera H, Halpern A E, Phung H M, Negro-Vilar A, Hannun Y A. J Cell Biol. 1992;117:121–133. doi: 10.1083/jcb.117.1.121. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Bowers B J, Christensen S C, Pauley J R, Paylor R, Yuva L, Dunbar S E, Wehner J M. J Neurochem. 1995;64:2737–2746. doi: 10.1046/j.1471-4159.1995.64062737.x. [DOI] [PubMed] [Google Scholar]
- 28.Abelovich A, Paylor R, Chen C, Kim J J, Wehner J M, Tonegawa S. Cell. 1993;75:1263–1271. doi: 10.1016/0092-8674(93)90614-v. [DOI] [PubMed] [Google Scholar]
- 29.Wehner J M, Bowers B J, Paylor R. Behav Genet. 1996;26:301–312. doi: 10.1007/BF02359386. [DOI] [PubMed] [Google Scholar]
- 30.Van der Zee E A, Luiten P G M, Disterhoft J F. Prog Neuropsychopharmacol Biol Psychiatry. 1996;21:531–572. doi: 10.1016/s0278-5846(97)00017-1. [DOI] [PubMed] [Google Scholar]
- 31.Fischer W, Gage F H, Bjorklund A. Eur J Neurosci. 1989;1:34–45. doi: 10.1111/j.1460-9568.1989.tb00772.x. [DOI] [PubMed] [Google Scholar]
- 32.Nicolle M M, Bizon J L, Gallagher M. Neuroscience. 1996;74:741–756. doi: 10.1016/0306-4522(96)00147-9. [DOI] [PubMed] [Google Scholar]
- 33.Campbell L W, Hao S Y, Thibault O, Blalock E M, Landfield P W. J Neurosci. 1996;16:6286–6295. doi: 10.1523/JNEUROSCI.16-19-06286.1996. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Kirischuk S, Verkhratsky A. Life Sci. 1996;59:451–459. doi: 10.1016/0024-3205(96)00324-4. [DOI] [PubMed] [Google Scholar]
- 35.Hartmann H, Velbinger K, Eckert A, Muller W E. Neurobiol Aging. 1996;17:557–563. doi: 10.1016/0197-4580(96)84400-9. [DOI] [PubMed] [Google Scholar]
- 36.Battaini F, Elkabes S, Bergamaschi V L, Lucchi L, de Grann P N E, Schuurman T, Wetsel W C, Trabucchi M, Govoni S. Neurobiol Aging. 1995;16:137–148. doi: 10.1016/0197-4580(94)00154-5. [DOI] [PubMed] [Google Scholar]
- 37.Narang N, Crews F T. Neurochem Res. 1995;10:1119–1126. doi: 10.1007/BF00995373. [DOI] [PubMed] [Google Scholar]
- 38.Rapp P R, Gallagher M. Proc Natl Acad Sci USA. 1996;93:9926–9930. doi: 10.1073/pnas.93.18.9926. [DOI] [PMC free article] [PubMed] [Google Scholar]