

Abstract
Importin13 (IPO13), the newest member of importin-β family discovered recently, is a unique nucleus-cytoplasm bidirectional transport receptor protein. In this study, IPO13 expression in human corneal tissue, limbal epithelial primary explant and clonal culture was evaluated by immunostaining and reverse-transcription polymerase chain reasgon. IPO13 function was evaluated in the corneal epithelial culture treated with IPO13 inhibitor, or fetal bovine serum (FBS)-containing Dulbecco’s modified Eagle’s medium (DMEM) medium by colony-forming efficiency, clone growth capacity, MTT, immunostaining, and Western blotting assay. IPO13 protein was expressed mainly in nuclei of limbal epithelial basal cells, but not in the other cell layers of limbus and full thickness of corneal epithelia. IPO13 was expressed in the majority of epithelial cells in early-stage clones and in the margin of late-stage clones. IPO13 was positively expressed in mouse TKE2 progenitor cells cultured in keratinocyte serum-free defined medium, while it became negative in FBS-containing DMEM, which promoted TKE2 cell differentiation. In the presence of IPO13 inhibitor, IPO13 expression and the proliferative capacity decreased in human limbal epithelial clones and mouse TKE2 cells, which were accompanied with the cell differentiation. In conclusion, our findings demonstrate for the first time that IPO13 is uniquely expressed by human limbal basal epithelial cells, and plays an important role in maintaining the phenotype, high proliferative potential, and less differentiation of corneal epithelial progenitor cells, suggesting that IPO13 could serve as a novel potential marker for corneal epithelial progenitor cells.
Keywords: Importin13, Cornea, Epithelium, Progenitor cell, Limbal stem cell
Introduction
The tissue-specific stem cells residing in certain adult tissues have been recognized to possess the ability to regenerate tissues, and they offer great therapeutic potential for treating diseased and damaged tissues and serving as gene delivery vehicles [1–6]. The easily visualized well-defined tissues of the ocular surface (i.e., cornea and conjunctiva) make it an ideal region to study epithelial stem cell function. It has been known for more than two decades that the homeostasis of corneal epithelium is governed by a small subpopulation of corneal epithelial stem cells, which are located in the basal layer of the limbus in human [7–9]. Although Majo et al. recently reported that the limbus was not the only niche for corneal epithelial stem cells and the corneal epithelium of the mouse could be serially transplanted and contained oligopotent stem cells [10], these findings need to be further investigated by multiple groups of researchers. Limbal epithelial stem cells exhibit unique characteristics that satisfy the widely accepted criteria for defining adult stem cells, which include 1) slow-cycling or long cell-cycle time during homeostasis in vivo; 2) small size and poor differentiation with primitive cytoplasm; 3) high proliferative potential after wounding or placement in culture; 4) the ability for self-renewal and functional tissue regeneration; and 5) the limbal niche, which is believed to be important in maintaining the “stemness” of these stem cells [11].
A variety of stem cell markers have been proposed to identify the adult stem cells. The major markers proposed for epithelial stem cells in the past decade can be categorized into at least three groups: nuclear proteins, such as the transcription factor P63; cell membrane or transmembrane proteins, including integrins (integrin β1, α6, and α9), receptors (epidermal growth factor receptor and transferrin receptor CD71), and drug-resistance transporters (ABCG2); and cytoplasmic proteins such as cytokeratins (K15 and K19) and α-enolase. Limbal basal epithelial cells are not homogeneous but consist of diverse cell populations including stem cells, transit amplifying cells (TACs) and terminally differentiated cells (TDCs) [12, 13]. Corneal epithelial stem cell markers provide a good tool for identification and isolation of these stem cells. In recent years, many proteins were found to act as specific positive or negative markers to identify limbal stem cells (LSCs). Human LSCs positive markers include ABCG2, P63, and intergrin α9, and negative markers include K3/K12, and connexin (Cx) 43 [14–20]. However, these proposed markers have insufficiency in identifying stem cells. For examples, ABCG2 is also expressed in corneal limbal stroma cells [21]; P63 is expressed by not only LSCs but also TACs, even a few of TDCs; and K3/K12 are negative markers that cannot directly identify LSCs [22]. Although the concept that human corneal epithelial stem cells reside in the limbus is widely accepted, there is no direct method to identify the corneal epithelial stem cells to date because of the lack of unique, direct, and specific molecular markers [14, 18].
Nucleocytoplasmic transport is an essential activity in eukaryotic cells. For example, mRNAs, tRNAs, and ribosomes are produced in the nuclear compartment and need to be exported to cytoplasm where they function in translation. Conversely, all nuclear proteins are imported from cytoplasm. Almost all of these biological activities are carried out by the importin and exportin families. The nuclear transport pathways for many important large molecular nuclear proteins—although not all of them—are mediated by importin β-related transport receptors, which form a large superfamily [23].
Importin 13 (IPO13), a newest member of importin-β family discovered recently, has been verified as a unique bidirectional transport receptor protein related with cell development and physiologic function such as lung, brain, and heart embryonic development [24]. However, many other physiological functions of IPO13 still remain unknown. In this present study, we investigated the expression and function of this novel nucleocytoplasmic transport receptor protein IPO13 in human corneal and limbal epithelial cells with an intention to explore whether IPO13 could serve as a novel potential marker for corneal epithelial progenitor cells.
Materials and Methods
Materials and Reagents
Rabbit polyclonal antibody against IPO13 was purchased from Proteintech Group (clone M-5113; Chicago, IL, http://www.ptglab.com). Mouse monoclonal antibodies against P63 (clone 4A4) for Western blot and immunostaining were from Santa Cruz Biotechnology (Santa Cruz, CA, http://www.scbt.com) and Dako (Glostrup, Denmark, http://www.dako.com), respectively. Rat polyclonal antibody against ABCG2 (clone M-70) was from Abcam (Cambridge, U.K., http://www.abcam.com). Goat polyclonal antibodies against K12 (clone L-20) and Cx43 (clone C-20) were from Santa Cruz Biotechnology. Fluorescein isothio-cyanate (FITC)-conjugated secondary antibodies, swine anti-rabbit, goat anti-mouse, and rabbit anti-goat IgG were from Dako. Texas Red conjugated secondary antibodies, donkey anti-rabbit, mouse, and goat IgG were from Jackson Immunoresearch Laboratories (West Grove, PA, http://www.jacksonimmuno.com). Mouse monoclonal antibody against β-actin, HRP-conjugated secondary antibodies, goat anti-mouse, and rabbit IgG for Western blot were from Santa Cruz Biotechnology. DNA and protein markers were from Fermentas (Burlington, Ontario, Canada, http://www.fermentas.com). PVDF membrane was from Hybond (Piscataway, NJ, http://www.hybond.com). Enhanced chemiluminescence reagent kit was from Pierce Biotechnology Thermo (Rockford, IL, http://www.piercenet.com/). Film was from Kodak (XBT-1; Xiamen, China, http://www.kodak.com). Trizol and primers were from Invitrogen (Carlsbad, CA, http://www.invitrogen.com). Reverse transcription kit, Go Taq DNA polymerase, MgCl2 solution, 10 mM polymerase chain reaction (PCR) nucleotide mix and 5 × Go Taq Flexi buffer were from Promega (Madison, WI, http://www.promega.com). Immunohistochemistry ABC Kit, bovine serum albumin (BSA), DAB coloration liquid, and mounting medium for fluorescence with DAPI were from Vector Laboratories (Burlingame, CA, http://www.vectorlabs.com). Fetal bovine serum (FBS) was from Hyclone (Logan, UT, http://www.hyclone.com). Dulbecco’s modified Eagle’s medium (DMEM), Ham F-12, HEPES, amphotericin B, gentamicin, streptomycin, 0.05% trypsin/0.03% EDTA solution, keratinocyte serum-free defined medium (KSFM), EFG human recombinant and bovine pituitary extract (BPE) were from GIBCO (Grand Island, NY, http://www.invitrogen.com). Dispase II was from Roche Diagnostics (Basel, Switzerland, http://www.roche-applied-science.com). Cholera toxin subunit A, dimethyl sulfoxide (DMSO), hydrocortisone, insulin-transferrin-sodium selenite media supplement, human recombinant EGF, mitomycin C were from Sigma-Aldrich (St. Louis, http://www.sigmaaldrich.com). NIH 3T3 mouse fibroblasts cell line were from ATCC (Rockville, MD, http://www.atcc.org). All plastic ware was from Corning (Corning, NY, http://www.corning.com).
Human Corneal and Limbal Tissue
Fresh human corneoscleral tissues (less than 48 hours postmortem) from donors aged 18–50 years, were obtained from Xiamen Eye Center. They were cut through horizontal meridian, embedded in a mixture of 75% (volume) OCT compound and 25% (volume) Immu-Mount (Thermo Shandon Inc., Pittsburgh, http://www.thermo.com), and then frozen in liquid nitrogen. Sections (6-μm thick) were cut with a cryostat (Leica CM 1850UV, Leica, Heerbrugg, Switzerland, http://www.leica.com) and stored at −80°C until they were used for immunostaining.
Human Limbal Epithelial Cell Counting
The human limbal corneas were incubated with dispase II (5 mg/ml) at 4°C over night. The limbal epithelial sheets were then collected and treated with 0.25% trypsin/0.03% EDTA at 37°C for 10 minutes to isolate single cells. The isolated single limbal epithelial cells were centrifuged and made into cyto-centrifuge slides by a cytology centrifuge (StatSpin Cytofuge 2, StatSpin, Westwood, MA, http://www.statspin.com/) and then used for immunostaining. IPO13, p63, ABCG2, K12, and Cx43-positve or negative cells index was assessed by point counting through a Nikon Eclipse 50i microscope (Nikon, Melville, NY, http://nikonusa.com). A total of 500–900 nuclei were counted in six to eight representative fields because this number was considered a minimum requirement to obtain a representative sample [25]. The index was expressed as the number of positively (or negatively) labeled nuclei/the total number of nuclei × 100%.
Human Corneal Epithelial Primary Explant Culture and Clonal Culture
An 8-mm trephine was used to isolate the central cornea from the limbus. After removed the layers from stroma to endothelium, the detached epithelial sheets from both the central cornea and limbal rim were cut into small pieces about 0.5 cm2, which were undergone primary explant culture in supplemented hormonal epidermal medium (SHEM) containing equal amounts of DMEM and Ham’s F12 medium supplemented with 5% FBS, 5 ng/ml EGF, 5 μg/ml insulin,5 μg/ml transferrin, 5 ng/ml sodium selenite, 0.5 μg/ml hydrocortisone, 30 ng/ml cholera toxin A, 0.5% DMSO, 50 μg/ml gentamicin and streptomycin, and 1.25 μg/ml amphotericin B, at 37°C under 5% CO2 and 95% humidity. The media were firstly changed after tissue adhesion and epithelial cells outgrow from the explants. The media were changed every 2–3 days thereafter. The epithelial tissues and cells were harvested on the fourth day and used for experiments.
Corneal epithelial cell clonal culture was established from single cell suspensions, which were isolated from limbal and corneal tissues respectively and cocultured on a mitomycin C (MMC)–treated 3T3 fibroblast feeder layer using a previously reported method with modification [25, 26]. In brief, each corneoscleral rim was trimmed, and the endothelial layer and iris remnants were removed. An 8-mm trephine was used to isolate the cornea from the limbus, and both limbus and central cornea were incubated with dispase II (5 mg/ml) at 4°C over night. The limbal and corneal epithelial sheets were then collected and treated with 0.25% trypsin/0.03% EDTA at 37°C for 10 minutes to isolate single cells. Mouse NIH 3T3 fibroblasts, grown in DMEM containing 10% FBS to confluence, were treated with MMC (5 μg/ml) for 2 hours and then trypsinized and plated at a density of 2 × 104 cells/cm2 in 60-mm dishes. The isolated limbal and corneal epithelial single cells were seeded at a density of 1 × 102 cells/cm2 on a 3T3 feeder layer in SHEM at 37°C under 5% CO2 and 95% humidity, and the media were changed every 2–3 days.
Colony Forming Efficiency and Growth Capacity
To evaluate growth capacity changes of human limbal epithelial clonal culture treated with IPO13 inhibitor, a mitomycin C-treated 3T3 fibroblast feeder layer was used as previously described [26, 27]. Single limbal epithelial cell suspensions were seeded at 1 × 103 cells per cm2 into 12-well culture plates. IPO13 inhibitor at different concentrations (5 and 10 μg/ml) was added into clonal culture media at day 6 when early-stage clone formation with untreated clonal culture as controls. Clones were harvested at day 9. Colony forming efficiency (CFE) was calculated as the percentage of the number of clones generated divided by the number of epithelial cells plated in a well and clone growth capacity was evaluated when cultured cells were stained with crystal violet at day 9 when IPO13 inhibitor had treated for 3 days.
TKE2 Cell Line Culture
TKE2 is a murine limbal/corneal epithelium-derived progenitor cell line [28]. They were expanded in KSFM supplemented with 2.5 ng/ml human recombinant EGF, 25 μg/ml BPE, 1% penicillin, and streptomycin at the seeding density of 5,000 cells/cm2 and the media were changed every 3 days. In order to induce differentiation of TKE2 cells, the culture media were changed from KSFM into DMEM supplemented with 10% FBS, 1% penicillin, and streptomycin and cultured for further 3–4 days. TKE2 cells were also treated with Ran G19V/Q69L, the inhibitor of IPO13, at different concentrations (5 and 10 μg/ml) with untreated TKE2 cells as normal controls and cultured in KSFM for 6 days.
MTT Assay
To evaluate cells proliferative capacity of TKE2 cells treated with IPO13 inhibitor at different concentrations (5 and 10 μg/ml), TKE2 cells were seeded at 5 × 103 cells per cm2 into 96-well culture plates. IPO13 inhibitor at different concentrations (5 and 10 μg/ml) were added into culture media after TKE2 cells adhered to dish bottom, and untreated TKE2 cells were served as normal control. TKE2 cells were harvested at 24, 48, and 72 hours, respectively and subjected to MTT [3-(4, 5-dimethylthiazol-2-yl)-2, 5-diphenyltetrazolium bromide] assay. Briefly, 100 μl of 1 mg/ml MTT constituted in culture media was added into each well, followed by incubation for 4 hours at 37°C in the dark. After incubation, the MTT solution was removed and the stained cells were washed twice in PBS followed by air-drying. The MTT-formazan products were extracted with 100 μl of DMSO in the dark at room temperature. The absorbance was measured spectrophotometrically at 570 nm using a BioTek ELX800 microplate reader (BioTek Instruments, Winooski, VT, http://www.biotek.com).
Immunostaining
The corneal frozen sections were fixed in 4% paraformaldehyde in phosphate buffered saline (PBS) at room temperature for 15 minutes. For immunohistochemical staining, the endogenous peroxidases activity was quenched with 0.3% hydrogen peroxide in PBS for 10 minutes. Sections were incubated with 0.2% Triton X-100 in PBS for 15 minutes, and incubated with 2% BSA to block the nonspecific sites. The antibodies of IPO13, P63, ABCG2, K12, and Cx43 (1:50) were then applied and incubated at 4°C for 16–18 hours. After three washes with PBS for 15 minutes, the sections were incubated with biotinylated anti-mouse, rabbit, or goat IgG (1:50) using Vectastain Elite ABC kits according to the manufacturer’s protocol. The reaction product was then developed with diaminobenzidine (DAB), the peroxidase substrate for 2 minutes and counterstained with hematoxylin. The sections were mounted with mounting medium and examined with a Nikon Eclipse 50i microscope.
For immunofluorescent staining, corneal and limbal frozen sections, corneal and limbal epithelial clonal culture, as well as TKE2 cells were fixed with 4% paraformaldehyde at room temperature for 15 minutes and permeated with 0.2% Triton X-100 in PBS for 15 minutes. After three rinses with PBS for 5 minutes each and preincubation with 2% BSA in PBS for 30 minutes, sections were incubated with primary antibodies (1:50) at 4°C for 16–18 hours. After three times washes with PBS for 5 minutes each, FITC (excitation wavelength: 488 nm) or Texas Red (excitation wavelength: 597 nm) conjugated secondary antibodies were applied at a 1:50 dilution for 1 hour in a dark incubation chamber at 37°C. After three times washes with PBS for 5 minutes each, the sections were mounted with an antifade solution with DAPI and photographed with a Nikon Eclipse TE-2000U epifluorescent microscope with a DMX 1200 digital camera.
RNA Extraction and Semiquantitative Reverse-Transcription Polymerase Chain Reaction
Total RNA was extracted from corneal and limbal epithelial explant primary culture, using acid guanidinium thiocyanate-phenol-chloroform extraction. The total RNA was quantified by its absorption at 260 nm and stored at −80°C before use. Using a housekeeping gene, β-actin, as an internal control, the mRNA expression of different molecular markers was analyzed by semi-quantitative reverse-transcription polymerase chain reaction (RT-PCR). Briefly, first-strand cDNAs were synthesized from 0.6 μg of total RNA with reverse transcriptase. PCR amplification was then performed with specific primer pairs designed from published human gene sequences and PRIMER5 designing software (Table 1) for different markers. Semiquantitative RT-PCR was established by terminating reactions at intervals of 20, 24, 28, 32, 36, and 40 cycles for each primer pair to ensure that the PCR products formed were within the linear portion of the amplification curve. The fidelity of the RT-PCR products was verified by comparing their size with the expected cDNA bands and by sequencing the PCR products.
Table 1.
Human primer sequences used for semiquantitative reverse-transcription polymerase chain reaction
| Gene | Accession | Sense primer | Antisense primer | PCR product (bp) |
|---|---|---|---|---|
| IPO13 | NM_014652 | 5′ CCCTGAGGCACCTACTGT 3′ | 5′ AGGGTCCAGGTTTACTCTTT 3′ | 478 |
| ΔN P63 | XM_036421 | 5′ CAGACTCAATTTAGTGAG 3′ | 5′ AGCTCATGGTTGGGGCAC 3′ | 440 |
| ABCG2 | AY_017168 | 5′ AGTTCCATGGCACTGGCCATA 3′ | 5′ TCAGGTAGGCAATTGTGAGG 3′ | 379 |
| K12 | NM_000223 | 5′ ACATGAAGAAGAACCACGAGGATG 3′ | 5′ TCTGCTCAGCGATGGTTTCA 3′ | 150 |
| Cx43 | M_65188 | 5′ CCTTCTTGCTGATCCAGTGGTAC 3′ | 5′ ACCAAGGACACCACCAGCAT 3′ | 154 |
| β-actin | NM_001101 | 5′ CATCTCTTGCTCGAAGTCCA 3′ | 5′ ATCATGTTTGAGACCTTCAACA 3′ | 310 |
Western Blotting Assay
Proteins of TKE2 cells cultured in different media or treated with IPO13 inhibitor were extracted with cold RIPA buffer. Equal amounts of proteins extracted from lysates were subjected to electrophoresis on 8% Tricine gels (Invitrogen) and then electrophoretically transferred to PVDF membranes. After 1 hour of blocking in 3% BSA, the blots were incubated with primary antibodies to IPO13 and P63 (1:100), and β-actin (1:5000) as a loading control. After three times washes with Tris-buffered saline with 0.05% Tween 20 for 10 minutes each, the membranes were incubated with HRP conjugated goat anti-rabbit and mouse IgG (1:10,000) for 1 hour at room temperature. The specific binds were visualized by enhanced chemiluminescence reagents and recorded on film.
Statistical Analysis
One-way analysis of variance test was used to make comparisons among experimental groups and control groups, and post-hoc the Dunnett test was followed to compare each experimental group with the control group. A p value of ≤0.05 was considered significant.
Results
IPO13 Expression in Human Corneal and Limbal Tissue
The immunohistostaining on frozen sections of human corneoscleral tissues showed that IPO13 was intensively expressed in nuclei and weakly expressed in cytoplasm of certain basal cells of limbal epithelium, but not in limbal superficial cells and the entire corneal epithelia (Fig. 1A). P63, a proposed limbal stem cell positive marker, was also expressed intensively expressed in nuclei of certain basal cells and a few suprabasal cells of limbal epithelium, but not in the entire corneal epithelia (Fig. 1A). ABCG2, another proposed limbal stem cell positive marker, was expressed in the cytoplasm of certain basal cells and a few suprabasal cells of limbal epithelium, but not in the entire corneal epithelia (Fig. 1A). K12 and Cx43, the proposed limbal stem cell negative markers, were expressed in the cytoplasm (K12) and cell junction (Cx43) in the full thickness of central corneal epithelia and all the cell layers besides basal cells of limbal epithelia (Fig. 1A).
Figure 1.

IPO13 expression on corneal tissue level compared with other proposal markers. (A): Immunohistochemical or immunofluorescent staining for IPO13, P63, ABCG2, K12, and Cx43 in human corneal and limbal tissues. Limbus shows brighter staining on certain basal cells and no staining on cornea for IPO13, P63, and ABCG2. IPO13 was intensively expressed in nucleus and weakly expressed in cytoplasm of certain basal cells of limbal epithelia. K12 and Cx43 had the similar expression pattern of limbal basal cells showing no staining but the other epithelial cell layers showing bright staining. Brown indicates positive expression by immunohistochemical staining. Red fluorescence indicates positive expression and blue fluorescence indicates nuclear staining with DAPI by immunofluorescent staining. Original magnification: ×200. (B): Double immunofluorescent staining for IPO13 (green) with p63 (red), ABCG2 (red), K12 (red), or Cx43 (red) in the human limbal epithelial cyto-centrifuge slides with DAPI nuclear counter-staining (blue). Coexpression for IPO13 with p63 or ABCG2 is shown by white arrows (B1 and B2). Opposite expression patterns were observed between IPO13 and K12, with Cx43 shown by green arrows (B3 and B4). Original magnification: ×300. (C): Limbal and corneal epithelial explant primary culture at different stages shows that the cell size of cultured limbal epithelia was much smaller than that of cultured corneal epithelial. The density of cultured limbal epithelial cells was increased as the culture time increased and that of cultured corneal epithelial cells decreased; some epithelial cells were detached and some fibroblasts emerged at D14. Original magnification: ×100. (D): Semiquantiative reverse transcription polymerase chain reaction profiles show mRNA expression in limbal and corneal epithelial explant primary culture for IPO13 (478 bp), P63 (440 bp), ABCG2 (379 bp), K12 (150 bp), Cx43 (154 bp), with β-actin (310 bp) as internal control. A 100-bp DNA ladder is shown in the left lane. Original magnification: ×200. Abbreviations: M, DNA markers; L1 and L2, limbal epithelial explant primary cultures; C1 and C2, corneal epithelial explant primary cultures.
Co-Expression of IPO13 with Stem Cell-Associated and Differentiation Markers in the Human Limbal Epithelial Cells
To verify the aforementioned difference and IPO13 may be more specially identify human limbal epithelial stem cells, we used limbal epithelial cytocentrifuge slides to perform double immunostaining between IPO13 and p63, ABCG2, K12, Cx43, the proposed limbal stem cell-associated or differentiation markers. The results showed that IPO13 was exclusively expressed by certain limbal epithelial cells. In the human limbal epithelial cells (Fig. 1B), 6.8% of cells were IPO13-positive, 10.1% were p63-positive, 11.5% were ABCG2-positive, 87.3% were K12-positive, and 77.4% were Cx43-positive (Table 2). IPO13 was coexpressed by 79.1% p63-positive and 70.6% ABCG2-positive cells (Fig. 1B, Table 2). A few cells were observed to express only IPO13 without ABCG2 (Fig. 1B). IPO13 was also positively expressed by 73.5% K12-negative and 81.1% Cx43-negative cells (Fig. 1B, Table 2). Such differences of expression patterns and percentages between IPO13 and proposed limbal stem cell-associated or differentiation markers suggested that IPO13 might be more specific than these proposed markers in marking LSC.
Table 2.
Percentage of IPO13+ cells in the entire population and the subpopulations of human limbal epithelial cells
| IPO13+ | P63+ | ABCG2+ | K12+ | Cx43+ | K12− | Cx43− | |
|---|---|---|---|---|---|---|---|
| Positive cells in entire population (%) | 6.8 (4.5) | 10.1 (6.7)a | 11.5 (5.3)a | 87.3 (10.5)b | 77.4 (12.7)b | ||
| IPO13+ cells in subpopulation (%) | 79.1 (10.4)b | 70.6 (21.3)b | 73.5 (20.7)b | 81.1 (12.3)b |
Data are mean (SD).
p < .05.
p <.01 (n = 5, compared with the percentage of IPO13-positive cells in the entire cell population).
IPO13 Expression in Human Corneal and Limbal Epithelial Primary Explant and Clonal Culture
As a first step to determine whether IPO13 was restricted to the limbal epithelial progenitor cells, we performed human corneal and limbal epithelial primary explant culture. We noted that epithelial cells began to grow out from both limbal and corneal explants at days 2 to 4 after culturing. However, the size of cultured limbal epithelial cells was much smaller than that of cultured corneal epithelial cells (Fig. 1C). Furthermore, the density of limbal epithelial cells increased as the culture time was prolonged. In contrast, the density of cultured corneal epithelial cells decreased while some cells became detached at day 14 (Fig. 1C). Using the mRNA level of β-actin as an internal control, semiquantitative RT-PCR analysis revealed that IPO13 mRNA was expressed by limbal epithelial but not by corneal epithelial in primary explant culture (Fig. 1D). A similar pattern was observed for the expression of P63 mRNA. In contrast, ABCG2 mRNA was also expressed at a very low level by corneal epithelial cells. As expected, K12 and Cx43 mRNAs were expressed by corneal epithelial cells at much higher levels than limbal epithelial cells.
We then studied the IPO13 expression by human corneal and limbal epithelial clones derived from coculturing on 3T3 fibroblast feeder layers to verify its unique expression by limbal epithelial progenitor cells. We noted that clones first appeared on day 4 in limbal epithelial culture, and their number and size increased as the culture time was prolonged (Fig. 2A, upper panel). In contrast, no clone was observed in corneal epithelial culture throughout the entire period (Fig. 2A, lower panel). By day 10, the cell size in the center of the limbal clone was larger than that in the periphery where limbal epithelial cells were in contact with 3T3 fibroblasts (Fig. 2B), suggesting that cells in the periphery were less differentiated than those in the center. Double immunostaining revealed that IPO13 was expressed in most cells in the clone on day 5. Interestingly, IPO13 was positive in the nucleus of the cell in the periphery but in the cytoplasm of the cell in the center of the clone on day 5 and day 7 (Fig. 2C). The positive IPO13 nuclear staining cells decreased as the clonal culture time prolonged, and there were only a few positive cells in the margin of late stage clone (Fig. 2C). The P63 staining exhibited the similar pattern besides P63 was expressed in the nuclei of all cells in the earlier stage clone (Fig. 2C).
Figure 2.
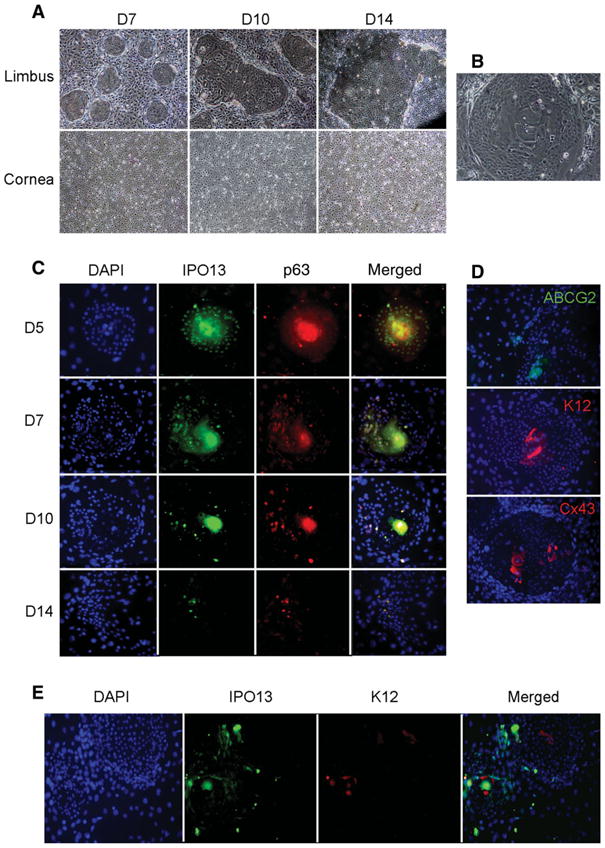
IPO13 expression in corneal epithelial clonal culture compared with other proposal markers. (A): Limbal and corneal epithelial clonal culture at different stages show that the number and size of clones in limbal epithelial clonal culture increased as the culture time prolonged. However, no clones were formed from the beginning to the end in corneal epithelial clonal cultures. (B): The cell size in the center of clone was much larger than that in the margin. (C): Immunofluorescent double staining for IPO13 and P63 in different stages clones showed that both IPO13 and P63 were expressed by almost all of the cells in early-stage clones. IPO13 was expressed in nuclei in the margin but in cytoplasm in the center of early-stage clones (D5, D7). IPO13-positive expression decreased as the clonal culture time prolonged, and there were only a few positive cells in the margin of late stage clones (D10, D14). P63 staining exhibited similar pattern. (D): Immunofluorescent staining showed that ABCG2 was only expressed in cytoplasm in the margin; K12 and Cx43 were only expressed in cytoplasm (K12) and cell junction (Cx43) in the center of clones. (E): Immunofluorescent double staining of IPO13 and K12 showed that IPO13 was expressed in nuclei in the margin of clones; however K12 was only expressed in cytoplasm in the center of clones. The two different expression patterns between IPO13 and K12 were almost opposite. Original magnification: ×200.
IPO13 and K12 double staining revealed that IPO13 was expressed in the nuclei in the margin of clone; however, K12 was only expressed in the cytoplasm in the center of clone. IPO13 and K12 expression were mutually exclusive in all clones (Fig. 2E). Immunofluorescent staining analysis revealed that ABCG2, the LSC positive marker, was only expressed in cytoplasm in the margin of clone; K12 and Cx43, the LSC negative markers, were only expressed in cytoplasm (K12) and cell junction (Cx43) in the center of late clone (Fig. 2D). These results collectively suggest that the cells in the clone margin were younger and more approached to progenitor cells than those in the center.
IPO13 Expression by Human Limbal Epithelial Clone Treated with IPO13 Inhibitor
To investigate the regulation of IPO13 on maintaining the phenotype and function of human limbal epithelial stem cells, we evaluated the changes of cell morphology, IPO13 and p63 expression, CFE and clone growth capacity of human limbal epithelial clones treated with IPO13 inhibitor. When treated with IPO13 inhibitor, the cell size became much bigger with smaller nucleus/cytoplasm ratio, which looked like differentiated cells even in early-stage clones (Fig. 3A). IPO13 and p63 expression in treated clones was down-regulated significantly depending on concentration of IPO13 inhibitor (Fig. 3B). Treated clones grew more slowly; and the size and number of forming clones decreased significantly depending on the concentration of IPO13 inhibitor, when compared with those of untreated control clones (Fig. 3C, 3D). These results suggest that IPO13 expression is important for maintaining the phenotype and proliferative property of limbal stem cells.
Figure 3.

Human limbal epithelial clonal culture treated with IPO13 inhibitor at different concentrations. (A): Clones treated for 3 days grew more slowly and clone size much smaller and cell size changed into much bigger even in early-stage clones than those of control group. Original magnification: × 100. (B): IPO13 and p63 positive expression. Original magnification: ×200. Clone growth capacity (C) and colony forming efficiency (D) decreased significantly depending on concentration of IPO13 inhibitor compared with normal controls. *p < .05 (compared with control).
IPO13 Expression by TKE2 Cells Treated with Changing Medium or IPO13 Inhibitor
In order to further verify that IPO13 was uniquely expressed by limbal epithelial progenitor cells and investigate how IPO13 regulates the differentiation of corneal epithelial progenitor cells, we evaluated the changes of cell phenotype and IPO13 expression in TKE2 cells treated with two different conditions, shifting medium, and IPO13 inhibitor. TKE2 cells cultured in KSFM could maintain progenitor cell characteristic, such as wide cell junction and small-cell size with big nucleus/cytoplasm ratio (Fig. 4A). Previous reports showed that TKE2 cells could be induced into differentiation by adding FBS or increasing calcium concentration in KSFM [27]. In this study, we modified the previous method, and found that the cell size and shape of TKE2 changed very quickly and pronouncedly to large squamous differentiated cells when medium was switched from KSFM into DMEM supplemented with 10% FBS for 1 day (Fig. 4). Immunofluorescent double staining of IPO13 and P63 showed that IPO13 was expressed intensively in the nuclei of TKE2 cells in KSFM. Nevertheless, IPO13 expression was significantly down-regulated when switched to DMEM supplemented with FBS (Fig. 4B). P63 expression also showed the same pattern (Fig. 4B). Western blotting assay result further confirmed the higher protein levels of IPO13 and P63 expression in TKE2 cells cultured in KSFM, but the decreased (IPO13) or no expression (P63) after the medium shift (Fig. 4C).
Figure 4.

TKE2 cells cultured in different media. (A): TKE2 cells cultured in keratinocyte serum-free defined medium (KSFM) had small-cell size with big nucleus/cytoplasm ratio (A1) and TKE2 cells rendered into large squamous cells when medium was switched from KSFM into Dulbecco’s modified Eagle’s medium (DMEM) supplemented with 10% fetal bovine serum (FBS) for 2 days (A2). (B): Immunofluorescent double staining for IPO13 and P63 by TKE2 cells cultured in different media. TKE2 cells in KSFM show intensive staining for IPO13 and P63 in nuclei and both of them have similar expression patterns. However, TKE2 in DMEM supplemented with FBS shows significant decreasing staining for IPO13 and P63. (C): Western blotting assay. TKE2 cells in KSFM show intensive expression for IPO13 and P63, but TKE2 cells in DMEM supplemented with FBS show downregulated expression of IPO13 and no expression of P63. Original magnification: ×200
The same phenomenon was also observed in TKE2 cells treated with IPO13 inhibitor, Ran G19V/Q69L. The cell size of TKE2 cells treated with IPO13 inhibitor changed to much bigger than that of control cells from day 3. The cells became spindle shape at low concentration (5 μg/ml) and squamous shape at high concentration (10 μg/ml; Fig. 5A). Immunofluorescent double staining of IPO13 and P63 revealed that both IPO13 and P63 were expressed intensively in the nuclei of TKE2 cells cultured in KSFM without IPO13 inhibitor. However, IPO13 expression was significantly downregulated depending on the concentration of IPO13 inhibitor added. P63 expression also showed the same pattern (Fig. 5B). Western blot result confirmed that both IPO13 and P63 protein expressions were significantly down-regulated depending on the concentrations of IPO13 inhibitor, P63 was no expression at high concentration of IPO13 inhibitor (Fig. 5C).
Figure 5.

TKE2 cells treated with IPO13 inhibitor. (A): TKE2 cells treated with IPO13 inhibitor at different concentrations. The cell size and shape among all groups were no any difference at D1. The cell size of TKE2 cells treated with IPO13 inhibitor changed into much bigger than that of control cells from D3. The cells became spindle shape at low concentration (5 μg/ml) and squamous shape at high concentration (10 μg/ml). (B): Immunofluorescent double staining of untreated TKE2 cells shows intensive staining for IPO13 and P63 in nuclei; both have similar expression patterns. However, TKE2 cells treated with IPO13 inhibitor show downregulated expression of IPO13 and P63 depending on concentration. (C): Western blotting assay. The expression of IPO13 and P63 were downregulated in TKE2 cells treated with IPO13 inhibitors depending on concentration. There were low levels of expression of IPO13 and no expression of P63 at high concentration. (D): MTT assay of TKE2 cells treated with an IPO13 inhibitor. The proliferative capacity of TKE2 cells treated with IPO13 inhibitor decreased significantly depending on concentration and treating time compared with normal controls. *, p < .05. **, p < .01 (compared with controls). Original magnification: ×200; high power: ×300.
In order to investigate IPO13 regulation on TKE2 cells proliferative capacity, MTT assay was performed. Figure 5D showed that TKE2 cells proliferative capacity decreased significantly when treated with IPO13 inhibitor at 24–72 hours of different time points compared with normal control (Fig. 5D). The results suggest that IPO13 may play a very important role in regulating corneal epithelial progenitor cell proliferation.
Discussion
As a novel member of importin-β family, IPO13 has been verified to play roles in importing and exporting some important proteins related with cell development and physiologic function. The substrates transported by IPO13 include pax-6, pax-3, glucocorticoid receptor, myopodin, UBC9, MGN, RBM8, and elF1A. IPO13 could interact with these cargoes and play intimate correlation with lung, brain, heart embryonic development. IPO13 also is the unique member of importin-β family that is regulated by double aspects, glucocorticoid and organism development stage [24, 29–38]. As a newfound nucleocytoplasmic transport receptor protein, many other physiologic functions of IPO13 in human different tissues and organs have been unknown yet. This present study for the first time explored the exclusive expression pattern of IPO13 in the basal cells of human limbal epithelia, the regulation of IPO13 in corneal epithelial progenitor cell differentiation and proliferation, and its potential role in identifying human LSCs.
The results of immunostaining and double staining of human corneal tissues, limbal epithelial cells as well as RT-PCR of their explant culture indicated that IPO13 was uniquely expressed by certain cells of basal layers in the limbal epithelia, whereas it is absent in the central corneal epithelia. IPO13 expression was intensively in nuclei and weakly in cytoplasm of these limbal basal cells. Whereas P63 and ABCG2 were expressed not only by limbal basal cells but also by some limbal suprabasal cells, even by a few of superficial cells (P63). The expression pattern of IPO13 was completely opposite to K12 and Cx43, the proposed negative markers for LSCs. In entire limbal epithelial cells, the percentage of IPO13-positive cells was the lowest. IPO13 was coexpressed only by a part of p63-positve cells and ABCG2-positve cells and also positively expressed by a part of K12-negative cells and Cx43-negative cells. These results suggest that the IPO13 expression may represent a characteristic of limbal progenitor cells and may be more specific than these proposed markers in marking LSCs.
We used the NIH mouse 3T3 cells line served as a feeder layer to promote progenitor cells to form the clone. The limbal epithelial clonal culture showed that the small cells in early-stage clone displayed epithelial stem cell-like phenotype. IPO13 was expressed by small cells in early-stage clone and in the margin of late stage clone; the expression pattern was similar to a proposed stem cell positive marker P63, but completely opposite to a differentiation marker K12. Interestingly, IPO13 was mainly positive in nuclei of the cell at the periphery while in cytoplasm of the cell in the center of the early-stage clone. This phenomenon may be correlated with IPO13 nucleocytoplasmic transport function. It has been demonstrated that IPO13 could be activated to import some nuclear protein cargoes, such as pax3, pax6, and glucocorticoid receptor that can bind to IPO13, into nucleus and then IPO13 releases these cargoes and lets them play biological functions in nucleus. Then IPO13 would export to cytoplasm where IPO13 waits for next binding and nuclear importing [29–31, 33]. Our results showed that IPO13 was mainly positive in nucleus of the cells at the periphery of clone, suggesting that it may be activated and play a functional role in maintaining progenitor cell properties. However, when only expressed in cytoplasm, IPO13 could be inactive so that the cells in the center of clone may be subjected to differentiation. We also found that the human central corneal epithelial cells did not form any clone in clonal culture (Fig. 2A) and there was no expression of IPO13 and p63 in central corneal epithelial cells of corneal tissue section (Fig. 1A). These results demonstrate again that no stem cell exists in human central cornea, which is different from the viewpoint of Majo [10].
We used human limbal epithelial cell clones and TKE2 cells, a murine limbal/corneal epithelium-derived progenitor cell line, to investigate the function of IPO13 in regulating corneal epithelial progenitor cell proliferation and differentiation. IPO13 was positively expressed by TKE2 cells cultured in KSFM. However, It was negatively expressed by TKE2 cells cultured in DMEM supplemented with 10% FBS, which promoted TKE2 cells to differentiate into mature epithelial cells and not maintain progenitor cell characteristics. In other words, IPO13 was only expressed by corneal epithelial progenitor cells, but not expressed by differentiated and mature corneal epithelial cells. Ran G19V/Q69L is an IPO13 inhibitor. The mutation Q69L abolishes GTP hydrolysis, and the mutation G19V results in a gain-of-function mutant in nucleus that is not, however, sensitive to the exchange factor RCC1, which is a guanine nucleotide exchanging factor acting on nuclear G protein Ran. Both of the two ways can block IPO13 transport function including nuclear importing and exporting [39]. When treated with IPO13 inhibitor, CFE and clonal growth capacity of human limbal epithelial progenitor cells and proliferative capacity of TKE2 cells all decreased significantly compared with normal control group. The IPO13 inhibitor-treated human limbal clones and TKE2 cells differentiated into spindle or squamous cells with the weak or negative expression of IPO13 and p63, as evaluated by immunostaining and Western blot analysis. In addition, western blotting results displayed that P63 protein had no expression when IPO13 protein still had low levels of expression (Figs. 4C, 5C). The reason may be that the positive expressional protein of IPO13 only existed in cytoplasm where IPO13 had not been activated; consequently, it cannot bind to nuclear protein to play its biological function. These results further suggest that IPO13 plays an important role in maintaining the undifferentiated phenotype and high proliferative potential of corneal epithelial progenitor cells.
In conclusion, our findings demonstrate for the first time that IPO13 is exclusively expressed by the basal cells of human limbal epithelia and primary cultured limbal epithelial progenitor cells. IPO13 plays an important role in regulating the proliferation and differentiation of corneal epithelial progenitor cells and in maintaining these progenitor cells from differentiation. Taken together, this study suggests that IPO13 could serve as a novel potential marker to identify the limbal epithelial progenitor cells.
Acknowledgments
This study was supported by grants from National Natural Science Fund of China (30700926), National High Technology Research and Development Program of China (2006AA02A131), Postdoctorate Fellowship of China (20070410807), Natural Science Fund of Fujian Province (2008J0113), Medical Innovation Project of Fujian Province (2007-CX-19), and Science and Technology Project of Xiamen (3502Z20074032).
Footnotes
Disclosure of Potential Conflicts of Interest
The authors indicate no potential conflicts of interest.
Author contributions: H.W., Y.X.Z., Z.G.L.: conception and design; H.W., J.X.Z., Z.G.L.: financial support; H.W., T.T., G.T., J.P., T.E.K., Z.G.L.: provision of study material or patients; H.W., J.T., Y.H.M.: collection and/or assembly of data; H.W., J.T., Y.H.M., W.L.: data analysis and interpretation; H.W.: manuscript writing; J.X.Z., Y.X.Z., Z.G.L.: administrative support; H.W., T.T., W.L., Y.P.Z., S.C.G.T., T.E.K., Z.G.L.: final approval of manuscript.
References
- 1.Tavazoie M, Van der Veken L, Silva-Vargas V, et al. A specialized vascular niche for adult neural stem cells. Cell Stem Cell. 2008;3:279–288. doi: 10.1016/j.stem.2008.07.025. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Kim K, Lee KM, Han DJ, et al. Adult stem cell-like tubular cells reside in the corticomedullary junction of the kidney. Int J Clin Exp Pathol. 2008;1:232–241. [PMC free article] [PubMed] [Google Scholar]
- 3.Roh JK, Jung KH, Chu K. Adult stem cell transplantation in stroke: its limitations and prospects. Curr Stem Cell Res Ther. 2008;3:185–196. doi: 10.2174/157488808785740352. [DOI] [PubMed] [Google Scholar]
- 4.Sotiropoulou PA, Candi A, Blanpain C. The majority of multipotent epidermal stem cells do not protect their genome by asymmetrical chromosome segregation. Stem Cells. 2008;26:2964–2973. doi: 10.1634/stemcells.2008-0634. [DOI] [PubMed] [Google Scholar]
- 5.Müller FJ, Laurent LC, Kostka D, et al. Regulatory networks define phenotypic classes of human stem cell lines. Nature. 2008;455:401–405. doi: 10.1038/nature07213. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Loebinger MR, Sage EK, Janes SM. Mesenchymal stem cells as vectors for lung disease. Proc Am Thorac Soc. 2008;5:711–716. doi: 10.1513/pats.200801-009AW. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Schermer A, Galvin S, Sun TT. Differentiation-related expression of a major 64K corneal keratin in vivo and in culture suggests limbal location of corneal epithelial stem cells. J Cell Biol. 1986;103:49–62. doi: 10.1083/jcb.103.1.49. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Tseng SC. Concept and application of limbal stem cells. Eye. 1989;3:141–157. doi: 10.1038/eye.1989.22. [DOI] [PubMed] [Google Scholar]
- 9.Thoft RA, Wiley LA, Sundarraj N. The multipotential cells of the limbus. Eye. 1989;3:109–113. doi: 10.1038/eye.1989.17. [DOI] [PubMed] [Google Scholar]
- 10.Majo F, Rochat A, Nicolas M, et al. Oligopotent stem cells are distributed throughout the mammalian ocular surface. Nature. 2008;456:250–255. doi: 10.1038/nature07406. [DOI] [PubMed] [Google Scholar]
- 11.Blau HM, Brazelton TR, Weimann JM. The evolving concept of a stem cell: entity or function? Cell. 2001;105:829–841. doi: 10.1016/s0092-8674(01)00409-3. [DOI] [PubMed] [Google Scholar]
- 12.Lavker RM, Sun TT. Epidermal stem cells: properties, markers, and location. Proc Natl Acad Sci USA. 2000;97:13473–13475. doi: 10.1073/pnas.250380097. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Schlötzer-Schrehardt U, Kruse FE. Identification and characterization of limbal stem cells. Exp Eye Res. 2005;81:247–264. doi: 10.1016/j.exer.2005.02.016. [DOI] [PubMed] [Google Scholar]
- 14.Chee KY, Kicic A, Wiffen SJ. Limbal stem cells: the search for a marker. Clin Experiment Ophthalmol. 2006;34:64–73. doi: 10.1111/j.1442-9071.2006.01147.x. [DOI] [PubMed] [Google Scholar]
- 15.dePaiva CS, Chen Z, Corrales RM, et al. ABCG2 transporter identifies a population of clonogenic human limbal epithelial cells. Stem Cells. 2005;23:63–73. doi: 10.1634/stemcells.2004-0093. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Chen Z, Howard EW, Stephen C, et al. Gap Junction protein connexin 43 serves as a negative marker for a stem cell-containing population of human limbal epithelial cells. Stem Cells. 2006;24:1265–1273. doi: 10.1634/stemcells.2005-0363. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Pellegrini G, Dellambra E, Golisano O, et al. P63 identifies keratinocyte stem cells. Proc Natl Acad Sci USA. 2001;98:3156–3161. doi: 10.1073/pnas.061032098. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Sun TT, Lavker RM. Corneal epithelial stem cells: Past, present, and future. J Investig Dermatol Symp Proc. 2004;9:202–207. doi: 10.1111/j.1087-0024.2004.09311.x. [DOI] [PubMed] [Google Scholar]
- 19.Qi H, Li DQ, Shine HD, et al. Nerve growth factor and its receptor TrkA serve as potential markers for human corneal epithelial progenitor cells. Exp Eye Res. 2008;86:34–40. doi: 10.1016/j.exer.2007.09.003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Qi Hong, Chuang Eliseu Yung, Yoon Kyung-Chul, et al. Patterned expression of neurotrophic factors and receptors in human limbal and corneal regions. Mol Vis. 2007;13:1934–1941. [PMC free article] [PubMed] [Google Scholar]
- 21.Du Y, Funderburgh ML, Mann MM, et al. Multipotent stem cells in human corneal stroma. Stem Cells. 2005;23:1266–1275. doi: 10.1634/stemcells.2004-0256. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Chen Z, Cintia SP, Luo L, et al. Characterization of putative stem cell phenotype in human limbal epithelia. Stem Cells. 2004;22:355–366. doi: 10.1634/stemcells.22-3-355. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Yoneda Y. Nucleocytoplasmic protein traffic and its significance to cell function. Genes Cells. 2000;5:777–787. doi: 10.1046/j.1365-2443.2000.00366.x. [DOI] [PubMed] [Google Scholar]
- 24.Mingot JM, Kostka S, Kraft R, et al. Importin 13: a novel mediator of nuclear import and export. EMBO J. 2001;20:3685–3694. doi: 10.1093/emboj/20.14.3685. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Kim HS, Jun SX, de Paiva CS, et al. Phenotypic characterization of human corneal epithelial cells expanded ex vivo from limbal explant and single cell culture. Exp Eye Res. 2004;79:41–49. doi: 10.1016/j.exer.2004.02.015. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Tseng SC, Kruse FE, Merritt J, et al. Comparison between serum-free and fibroblast-cocultured single-cell clonal culture systems: evidence showing that epithelial antiapoptotic activity is present in 3T3 fibroblast-conditioned media. Curr Eye Res. 1996;15:973–984. doi: 10.3109/02713689609017643. [DOI] [PubMed] [Google Scholar]
- 27.Li DQ, Chen Z, Song XJ, et al. Partial enrichment of a population of human limbal epithelial cells with putative stem cell properties based on collagen type IV adhesiveness. Exp Eye Res. 2005;80:581–590. doi: 10.1016/j.exer.2004.11.011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Kawakita T, Shimmura S, Hornia A, et al. Stratified epithelial sheets engineered from a single adult murine corneal/limbal progenitor cell. J Cell Mol Med. 2008;12:1303–1316. doi: 10.1111/j.1582-4934.2008.00297.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Ploski JE, Shamsher MK, Radu A. Paired-type homeodomain transcription factors are imported into the nucleus by karyopherin 13. Mol Cell Biol. 2004;24:4824–4834. doi: 10.1128/MCB.24.11.4824-4834.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Tao T, Lan J, Lukacs GL, et al. Importin 13 regulates nuclear import of the glucocorticoid receptor in airway epithelial cells. Am J Respir Cell Mol Biol. 2006;35:668–680. doi: 10.1165/rcmb.2006-0073OC. [DOI] [PubMed] [Google Scholar]
- 31.Tao T, Lan J, John F, Presley, et al. Nucleocytoplasmic shuttling of Igl2 is developmentally regulated in fetal lung. Am J Respir Cell Mol Biol. 2004;30:350–359. doi: 10.1165/rcmb.2003-0126OC. [DOI] [PubMed] [Google Scholar]
- 32.Yamaguchi YL, Tanaka SS, Yasuda K, et al. Stage-specific importin 13 activity influences meiosis of germ cells in the mouse. Dev Biol. 2006;297:350–360. doi: 10.1016/j.ydbio.2006.04.465. [DOI] [PubMed] [Google Scholar]
- 33.Rondanino C, Bousser MT, Monsigny M, et al. Sugar-dependent nuclear import of glycosylated proteins in living cells. Glycobiology. 2003;13:509–519. doi: 10.1093/glycob/cwg064. [DOI] [PubMed] [Google Scholar]
- 34.Liang J, Ke G, You W, et al. Interaction between importin 13 and myopodin suggests a nuclear import pathway for myopodin. Mol Cell Biochem. 2008;307:93–100. doi: 10.1007/s11010-007-9588-1. [DOI] [PubMed] [Google Scholar]
- 35.Yamasaki H, Sekimoto T, Ohkubo T, et al. Zinc finger domain of snail functions as a nuclear localization signal for importin-mediated nuclear import pathway. Genes Cells. 2005;10:455–464. doi: 10.1111/j.1365-2443.2005.00850.x. [DOI] [PubMed] [Google Scholar]
- 36.Shoubridge C, Cloosterman D, Parkinson-Lawerence E, et al. Molecular pathology of expanded polyalanine tract mutations in the Aristaless-related homeobox gene. Genomics. 2007;90:59–71. doi: 10.1016/j.ygeno.2007.03.005. [DOI] [PubMed] [Google Scholar]
- 37.Quan Yu, Ji Z, Wang X, et al. Evolutionary and transcriptional analysis of karyopherin beta superfamily proteins. Mol Cell Proteomics. 2008;7:1254–126. doi: 10.1074/mcp.M700511-MCP200. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Whitehurst AW, Robinson FL, Moore MS, et al. The death effector domain protein PEA-15 prevents nuclear entry of ERK2 by inhibiting required interactions. J Bio Chem. 2004;279:12840–12847. doi: 10.1074/jbc.M310031200. [DOI] [PubMed] [Google Scholar]
- 39.Ohba T, Seki T, Azuma Y, et al. Premature chromatin condensation induced by loss of RCC1 is inhibited by GTP- and GTPγS-ran, but not GDP-ran. J Bio Chem. 1996;271:14665–14667. doi: 10.1074/jbc.271.25.14665. [DOI] [PubMed] [Google Scholar]