

Abstract
Axon regeneration in the mature mammalian central nervous system (CNS) is extremely limited after injury. Consequently, functional deficits persist after spinal cord injury (SCI), traumatic brain injury, stroke, and related conditions that involve axonal disconnection. This situation differs from that in the mammalian peripheral nervous system (PNS), where long- distance axon regeneration and substantial functional recovery can occur in the adult. Both extracellular molecules and the intrinsic growth capacity of the neuron influence regenerative success. This chapter discusses determinants of axon regeneration in the PNS and CNS.
1 Introduction
Central nervous system (CNS) axons do not spontaneously regenerate after injury in adult mammals. In contrast, peripheral nervous system (PNS) axons readily regenerate, allowing recovery of function after peripheral nerve damage. Aguayo and colleagues demonstrated that at least some mature CNS neurons retain the capacity to regenerate when provided with a permissive peripheral nerve graft (Richardson et al. 1980, 1984; David and Aguayo, 1981; Benfey and Aguayo, 1982). This work suggested that the PNS environment is stimulatory and/or that the CNS environment is inhibitory for axon growth. Subsequent studies identified both growth- promoting factors in the PNS and growth- inhibiting factors in the CNS. Inhibitors of regeneration include specific proteins in CNS myelin and molecules associated with the astroglial scar. In addition, slower debris clearance in the CNS relative to the PNS may impede axonal re-growth. The cell-autonomous failure of the cell of axotomized CNS neurons to induce those growth- promoting genes, which are highly upregulated by injured PNS neurons also limits brain and spinal cord repair. An understanding of factors which influence axon growth is critical for the development of therapeutics to promote CNS regeneration.
2 Axon Regeneration in the Peripheral Nervous System
2.1 Overview of Peripheral Nervous System Regeneration
After peripheral nerve injury, axons readily regenerate. The distal portion of the axon, which is disconnected from the cell body, undergoes Wallerian degeneration. This active process results in fragmentation and disintegration of the axon. Debris is removed by glial cells, predominantly macrophages. Proximal axons can then regenerate and re-innervate their targets, allowing recovery of function.
2.2 Regeneration-Associated Genes
Following axotomy, PNS neurons upregulate numerous regeneration-associated genes (RAGs). Some of these genes have a direct role in axon regeneration, while others do not. A number of RAGs have been shown to be important for neurite outgrowth and/or regeneration. These include c-Jun (Raivich et al. 2004), activating transcription factor-3 (ATF-3) (Seijffers et al. 2006), SRY-box containing gene 11 (Sox11) (Jankowski et al. 2009), small proline-repeat protein 1A (SPRR1A) (Bonilla et al. 2002), growth-associated protein-43 (GAP-43) and CAP-23 (Bomze et al. 2001).
One strategy to identify RAGs involves injuring a peripheral nerve, and then observing gene expression changes in the corresponding cell bodies (Bonilla et al. 2002; Tanabe et al. 2003; Costigan et al. 2002). A number of such studies have used gene profiling technology to examine gene expression changes in sensory neurons following axotomy. For example, Bonilla et al. (2002) demonstrated that SPRR1A is highly induced in dorsal root ganglion (DRG) neurons one week after sciatic nerve transection (protein increased more than 60-fold from whole DRGs). Immunohistochemistry demonstrated expression of SPRR1A in DRG neuronal cell bodies and regenerating peripheral axons. SPRR1A expression is also increased after sciatic nerve injury in the ventral horn motor neuron cell bodies and sensory fibers within the spinal cord (Fig. 1). Herpes simplex virus-mediated overexpression of SPRR1A in embryonic chick DRG neurons promotes neurite outgrowth. The association of SPRR1A expression with regeneration and its ability to promote neurite outgrowth suggest that it may have a role in axon regeneration.
Fig. 1.
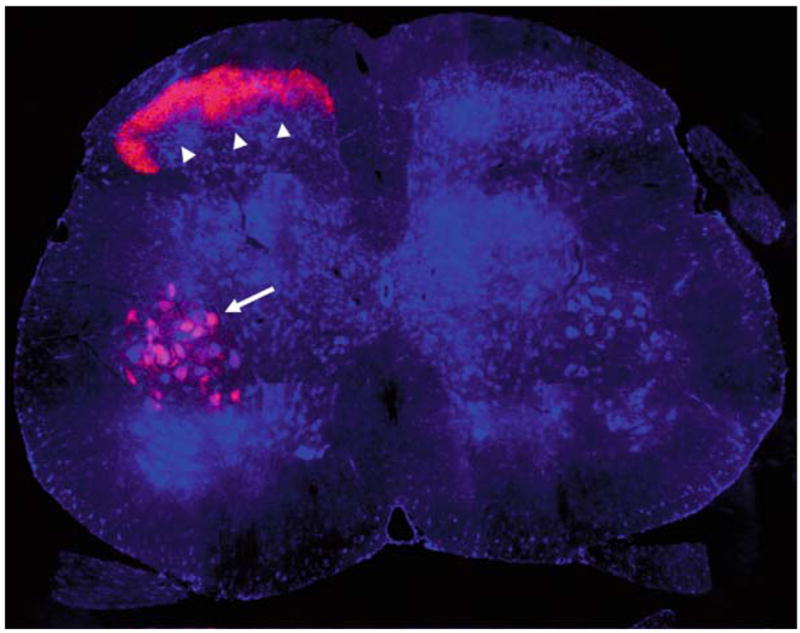
SPRR1A upregulation in the central process of primary afferent sensory neurons and in motoneurons after sciatic nerve injury. The sciatic nerve was crushed at the mid-thigh on one side of an adult mouse. Seven days later, the animal was sacrificed, and L5 spinal cord transverse sections were processed for anti-SPRR1A immunohistology (red) and for Nissl Stain (blue). Note the intense SPRR1A protein upregulation in afferent terminals in the dorsal horn (arrowheads) and in ventral horn motoneurons (arrow). Upregulation is confined to the injured side (left) with very low levels of SPRR1A on the intact side (right). Dorsal is up. Methods as in Bonilla et al. (2002). Image courtesy of Dr. William B. Cafferty
ATF-3 is a transcriptional factor induced in sensory neurons after injury (Tanabe et al. 2003; Boeshore et al. 2004). Over expression of ATF-3 promotes neurite outgrowth (Seijffers et al. 2006). Sox11 and c-Jun are injury-induced transcription factors and are required for efficient nerve regeneration (Jankowski et al. 2009; Raivich et al. 2004). Transcriptional factors associated with regeneration, such as c-Jun, appear to induce the expression of other RAGs and thereby may promote a growth state (Raivich et al. 2004).
3 Axon Regeneration in the Central Nervous System
3.1 Overview of Central Nervous System Regeneration
Pioneering work by Aguayo and colleagues demonstrated that adult mammalian CNS neurons, which normally do not regenerate, are able to grow for long distances into the permissive environment of a peripheral nerve graft (Richardson et al. 1980, 1984; David and Aguayo 1981; Benfey and Aguayo 1982). These studies demonstrated that the environment is a critical determinant of axon regeneration. Subsequently, numerous molecules were identified within the CNS that limit regeneration.
The two major classes of CNS regeneration inhibitors are the myelin-associated inhibitors (MAIs) and the chondroitin sulfate proteoglycans (CSPGs). These molecules limit axon regeneration, and, by interfering with their function, some degree of growth in the adult CNS is achieved.
Cell-autonomous factors are also important determinants of CNS regeneration failure. CNS neurons do not upregulate growth-associated genes to the same extent as do PNS neurons. Consequently, their ability to regenerate is limited even in the absence of inhibitors. Increasing the intrinsic growth capacity of neurons allows modest axon regeneration within the CNS (Bomze et al. 2001; Neumann and Woolf 1999).
Axon regeneration is one of many factors influencing recovery after CNS damage. Sprouting of uninjured axons can also contribute dramatically to functional improvements. Additionally, plasticity at the synaptic level may underlie a certain degree of recovery seen even in the absence of treatments (i.e., learning to use spared neuronal circuitry in new ways). CNS regeneration studies do not always distinguish between these different mechanisms, and, for the purpose of this discussion, they will be considered together. Replacement of lost neuronal cell bodies, a prominent component of many CNS disorders, is beyond the scope of this chapter.
3.2 Myelin-Associated Inhibitors
MAIs are proteins expressed by oligodendrocytes as components of CNS myelin. MAIs impair neurite outgrowth in vitro and are thought to limit axon growth in vivo after CNS damage. MAIs include Nogo-A (Chen et al. 2000; GrandPre et al., 2000), myelin-associated glycoprotein (MAG) (McKerracher et al., 1994), oligodendrocyte myelin glycoprotein (OMgp) (Kottis et al. 2002), ephrin-B3 (Benson et al. 2005) and Semaphorin 4D (Sema4D) (Moreau-Fauvarque et al. 2003). Three of these (Nogo-A, MAG and OMgp) interact with a neuronal Nogo-66 receptor 1 (NgR1) to limit axon growth. These three structurally unrelated ligands also show affinity for a second axon growth-inhibiting receptor, paired immunoglobulin-like receptor B (PirB) (Atwal et al. 2008). For the most part, MAIs are not found in PNS myelin, which is produced by Schwann cells rather than oligodendrocytes. An exception is MAG, which is present in PNS myelin but is cleared much more rapidly by glial cells in the periphery than in the brain and spinal cord.
One of the most well- characterized MAIs is Nogo-A. Genetic deletion of Nogo-A promotes corticospinal (Fig. 2) and raphespinal tract growth and enhances functional recovery after SCI, although this phenotype is modulated by strain background, age and axonal tract (Kim et al. 2003b; Simonen et al. 2003; Zheng et al. 2003; Dimou et al., 2006). Even after controlling for these factors, certain targeted mutations create a greater axon growth response than others (Cafferty et al. 2007b). However, even the least growth-promoting mutation of the Nogo gene has an enhanced growth phenotype after pyramidotomy (Cafferty and Strittmatter 2006). In addition, antibodies that target Nogo-A promote axonal growth and functional recovery after CNS injury (Z’Graggen et al., 2000; Wiessner et al. 2003; Seymour et al. 2005). An anti-Nogo-A antibody has advanced to clinical trials for SCI.
Fig. 2.
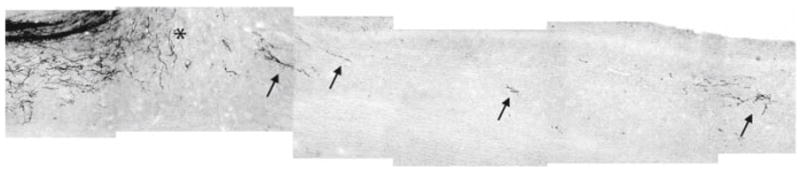
Corticospinal tract (CST) axonal tracing in mice lacking Nogo-A/B after mid-thoracic spinal cord dorsal hemisection. A parasagittal section of thoracic spinal cord from a mouse with a mutation in the Nogo gene that prevents Nogo-A and Nogo-B expression. The CST is traced from a cortical biotin dextran amine (BDA) injection after dorsal hemisection. Rostral is left; dorsal is up. The lesion is indicated by the asterisk. Note the evidence of CST fiber growth caudal to the lesion site (arrows). Significantly less BDA tracing is present caudal to the lesion in control animals (not shown). This photographic montage is a different image from the same mice described in Kim et al. (2003b)
Two inhibitory portions of Nogo-A have been identified. The first, termed Nogo-66, is a 66 amino acid fragment which interacts with NgR1 on the neuronal membrane (Fournier et al. 2001). An adjacent 24 amino acid sequence, although not inhibitory in itself, facilitates picomolar-affinity binding of the Nogo-66 to NgR1 (Hu et al. 2005). The Nogo-66 domain can also interact directly with a secondary receptor, PirB, on the surface of neurons (Atwal et al. 2008). The other inhibitory portion of Nogo-A Amino-Nogo, acts via an independent mechanism to disrupt neuronal integrin function (Hu and Strittmatter 2008).
Two other Nogo isoforms exist (Nogo-B and Nogo-C), which contain the inhibitory Nogo-66 loop found in Nogo-A but lack the Amino-Nogo sequence. These isoforms are not found naturally in myelin, but transgenic overexpression of Nogo-C in Schwann cells, which normally do not express any Nogo isoform, delays peripheral nerve regeneration. This demonstrates the ability of Nogo-66 to limit axon regeneration in vivo (Kim et al. 2003a).
MAG is another inhibitory protein present in CNS myelin (McKerracher et al., 1994). MAG interacts with several neuronal receptors to limit neurite outgrowth in vitro, including NgR1 (Liu et al. 2002; Domeniconi et al. 2002), gangliosides (Vyas et al. 2002; Mehta et al. 2007), Nogo receptor 2 (NgR2) (Venkatesh et al. 2005) and PirB (Atwal et al. 2008). The relative importance of each receptor varies with neuronal type. For example, in postnatal DRG neurons, NgR1 mediates the majority of inhibition by MAG, whereas in postnatal cerebellar granule neurons, GT1b appears to be more important (Mehta et al. 2007). Although MAG inhibits neurite outgrowth in vitro, no evidence of enhanced corticospinal tract (CST) regeneration or sprouting after SCI or optic nerve injury is observed in MAG knockout mice (Bartsch et al. 1995). Thus, MAG appears to have a less important role in limiting CNS axon growth than other inhibitors such as Nogo-A.
OMgp, another MAI that interacts with NgR1 and PirB, also limits neurite outgrowth in vitro (Kottis et al. 2002). CNS regeneration studies with OMgp knockout mice have not been described.
NgR1 is a glycosylphosphatidylinositol (GPI)-linked membrane receptor, which lacks a transmembrane or cytoplasmic domain. It interacts with coreceptors LINGO-1 (Mi et al. 2004) and p75 (Wang et al, 2002) or TAJ/TROY (Park et al. 2005; Shao et al. 2005), depending on neuronal type, to limit axon growth. Enhanced rubrospinal and raphespinal, but not corticospinal, axon regeneration is observed in NgR1 knockout mice after SCI (Kim et al. 2004). The enhanced axonal growth is correlated with improved functional recovery. Although CST regeneration is not observed after SCI in mice lacking NgR1, corticofugal axons do show enhanced growth in these mice after a stroke (Li et al. 2004) or pyramidotomy (Cafferty and Strittmatter 2006). Thus, NgR1 limits axonal growth and functional recovery after CNS damage.
NgR1 function can be blocked by a soluble form of extracellular NgR1 fused to human Fc (NgR(310)ecto-Fc). NgR(310)ecto-Fc promotes corticospinal and raphe-spinal growth and functional recovery after SCI in rats (Li et al. 2004). In addition, transgenic mice which secrete NgR(310)ecto under control of the GFAP promoter (causing reactive astrocytes to secrete high levels of the protein after CNS injury) show enhanced functional recovery after SCI (Li et al. 2005).
A competitive NgR1 antagonist, Nogo-extracellular peptide, residues 1–40 (NEP1-40), binds to, but does not activate, NgR1. NEP1-40 attenuates inhibition of neurite outgrowth by Nogo-A and CNS myelin. After SCI, NEP1-40 promotes corticospinal and raphespinal regeneration and functional recovery, even when the initiation of treatment is delayed for one week (GrandPre et al., 2002; Li and Strittmatter, 2003). Because NEP1-40 blocks Nogo-66, but not other NgR1 ligands, it is less effective than NgR(310)ecto-Fc.
The studies described above confirm the importance of NgR1 and its ligands in limiting CNS regeneration. In vivo functional studies of other MAI receptors have not yet been reported.
3.3 Chondroitin Sulfate Proteoglycans
The astroglial scar, which forms after CNS injury, is a physical barrier to regeneration, and also contains inhibitory molecules that impede axon growth. CSPGs are the main inhibitory molecules found in the glial scar (reviewed in Morgenstern et al. 2002). CSPGs are upregulated by reactive astrocytes after CNS damage and are both membrane bound and secreted into the extracellular space. Members of this class of inhibitors include neurocan (Asher et al. 2000), versican (Schmalfeldt et al., 2000), brevican (Yamada et al. 1997), phosphacan (Inatani et al. 2001), aggrecan (Carmen et al. 2007) and NG2 (Dou and Levine, 1994). A receptor for CSPGs has not been identified.
Interfering with CSPG function promotes axon regeneration in the CNS. CPSGs contain core proteins with attached glycosaminoglycan (GAG) side chains, which can be cleaved by the bacterial enzyme Chondroitinase ABC. This enzyme reduces the inhibitory activity of CSPGs in vitro (McKeon et al. 1995; Carmen et al. 2007). Moreover, when Chondroitinase ABC is administered after spinal contusion in rats, regeneration of both descending CST and ascending sensory fibers can be detected (Bradbury et al. 2002). This axonal growth is accompanied by enhanced recovery of associated locomotor and proprioceptive functions. Several other studies have confirmed that Chondrotinase ABC promotes axonal growth after CNS injury (Barritt et al. 2006; Massey et al. 2006; Cafferty et al. 2007a).
3.4 Other Axon Regeneration Inhibitors
Axon regeneration inhibitors (ARIs) found in the CNS that are not present in myelin or the glial scar include repulsive guidance molecule (RGM) and semaphorin 3A (Sema3A). Evidence that these molecules limit CNS regeneration include studies demonstrating that an anti-RGMa antibody (Hata et al. 2006) or a small molecule inhibitor of Sema3A (Kaneko et al. 2007) promote functional recovery after SCI in rats.
3.5 Inhibitory Signaling Pathways
Multiple ARIs have been shown to activate the small GTPase ras homolog gene family, member A (RhoA) (Niederost et al. 2002; Fournier et al. 2003; Shao et al. 2005). Activated RhoA, in turn, activates Rho-associated coiled-coil containing protein kinase 2 (ROCK2), a kinase that regulates actin cytoskeletal dynamics (reviewed in Schmandke et al. 2007). Activation of ROCK2 results in cessation of neurite growth. Interfering with RhoA or ROCK2 activity promotes CNS axon regeneration and functional recovery.
Ibuprofen, which inhibits RhoA, promotes corticospinal and raphespinal sprouting after spinal contusion (Fu et al. 2007; Wang et al. 2009), as well as long- distance raphespinal axon regeneration after a complete spinal cord transection (Wang et al. 2009). Tissue sparing at the lesion site is also enhanced by ibuprofen and thus may contribute to functional recovery (Wang et al. 2009).
The ROCK2 inhibitor Y27632 promotes CST sprouting and locomotor recovery after dorsal hemi section spinal injury in rats (Fournier et al. 2003). In addition, ROCK2 knockout mice show enhanced functional recovery after SCI (Duffy PJ, Schmandke A, and Strittmatter SM, unpublished observations). Thus, ROCK2 is an important mediator of CNS regeneration failure.
Some evidence suggests that epidermal growth factor receptor (EGFR) contributes to CNS regeneration failure. One study demonstrated enhanced optic nerve regeneration after treatment with the irreversible EGFR inhibitor PD168393 (Koprivica et al. 2005). This study provides evidence that trans-activation of EGFR mediates inhibition of neurite outgrowth by MAIs and CSPGs. Another study observed that PD168393 enhances sparing, and/or regeneration of 5-hydroxytryptophan-immunoreactive (serotonergic) fibers caudal to a spinal cord lesion (Erschbamer et al. 2007). Thus, EGFR activation appears to limit recovery after CNS trauma.
Other molecules that have been implicated in ARI- signaling include protein kinase C, (Sivasankaran et al. 2004), LIM kinase, Slingshot phosphatase and cofilin (Hsieh et al. 2006).
3.6 Intrinsic Growth State of the Neuron
In contrast to the PNS, the upregulation of peripheral RAGs (see Sect. 2.2) is relatively modest in the CNS after injury (Fernandes et al. 1999; Marklund et al. 2006). This paucity of RAG expression appears to be partially responsible for the limited ability of CNS neurons to regenerate. Increasing RAG expression in CNS neurons improves their regenerative ability. For example, Bomze et al. (2001) demonstrated that overexpressing GAP-43 and CAP-23 together promotes sensory axon regeneration after SCI.
DRG neurons have a peripheral process and a central process. Injury to the peripheral process results in robust upregulation of RAGs, as described above. However, injury to the central process by dorsal rhizotomy or spinal cord dorsal hemi section does not induce nearly as robust of a regenerative response, and central processes fail to regenerate in the CNS. Injury of peripheral axons one week prior to central injury (termed a conditioning lesion) allows some degree of sensory fiber regeneration within the spinal cord (Neumann and Woolf 1999). The conditioning lesion appears to enhance the growth state of the neuron such that its central process is able to regenerate in the CNS environment.
Cyclic adenosine monophosphate (cAMP) is a second messenger molecule which influences the growth state of the neuron. cAMP levels are increased by a peripheral conditioning lesion (Qiu et al. 2002). Elevation of cAMP levels by intra-ganglionic injection of a membrane-permeable cAMP analog, dibutyryl cAMP (db-cAMP), mimics the growth-promoting effects of a conditioning lesion, promoting regeneration of sensory axons within the spinal cord (Neumann et al. 2002; Qiu et al. 2002). In vivo injection of db-cAMP prior to DRG removal also improves the ability of dissociated DRG cultures to grow on MAG or CNS myelin in vitro, indicating that cAMP elevation can promote growth in the presence of MAIs (Qiu et al. 2002).
Rolipram, a phosphodiesterase 4 inhibitor, increases cAMP by interfering with its hydrolysis. When delivered 2 weeks after spinal cord hemisection, rolipram increases serotonergic axon regeneration into embryonic spinal tissue grafts implanted at the lesion site at the time of injury (Nikulina et al. 2004). Reactive gliosis is reduced by rolipram, and this might contribute to the functional recovery observed. Additionally, enhanced axonal sparing and myelination are induced by cAMP elevation in combination with Schwann cell grafts after SCI, compared to Schwann cell grafts alone (Pearse et al. 2004), suggesting additional mechanisms by which cAMP elevation could lead to functional improvements. Nonetheless, the demonstration of serotonergic axon growth into grafts at the lesion site in both of these studies indicates that cAMP elevation can induce CNS axon regeneration.
cAMP elevation activates protein kinase A (PKA) and induces CREB-mediated transcription of various growth-associated genes, including IL-6 and arginase I. Subsequent synthesis of polyamines by arginase I has been proposed as a possible mechanism by which cAMP increases neurite growth (Cai et al. 2002).
4 Conclusion
Regeneration of the injured mammalian CNS was once thought to be an unachievable goal. Recent advances in our understanding of factors which limit CNS regeneration and those which facilitate PNS regeneration have lead to therapies which allow some degree of recovery from brain and SCI in animal models. These findings open the possibility of promoting regeneration of the damaged human CNS.
References
- Asher RA, Morgenstern DA, Fidler PS, Adcock KH, Oohira A, Braistead JE, Levine JM, Margolis RU, Rogers JH, Fawcett JW. Neurocan is upregulated in injured brain and in cytokine-treated astrocytes. J Neurosci. 2000;20:2427–2438. doi: 10.1523/JNEUROSCI.20-07-02427.2000. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Atwal JK, Pinkston-Gosse J, Syken J, Stawicki S, Wu Y, Shatz C, Tessier-Lavigne M. PirB is a functional receptor for myelin inhibitors of axonal regeneration. Science. 2008;322:967–970. doi: 10.1126/science.1161151. [DOI] [PubMed] [Google Scholar]
- Barritt AW, Davies M, Marchand F, Hartley R, Grist J, Yip P, McMahon SB, Bradbury EJ. Chondroitinase ABC promotes sprouting of intact and injured spinal systems after spinal cord injury. J Neurosci. 2006;26:10856–10867. doi: 10.1523/JNEUROSCI.2980-06.2006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bartsch U, Bandtlow CE, Schnell L, Bartsch S, Spillmann AA, Rubin BP, Hillenbrand R, Montag D, Schwab ME, Schachner M. Lack of evidence that myelin-associated glycoprotein is a major inhibitor of axonal regeneration in the CNS. Neuron. 1995;15:1375–1381. doi: 10.1016/0896-6273(95)90015-2. [DOI] [PubMed] [Google Scholar]
- Benfey M, Aguayo AJ. Extensive elongation of axons from rat brain into peripheral nerve grafts. Nature. 1982;296:150–152. doi: 10.1038/296150a0. [DOI] [PubMed] [Google Scholar]
- Benson MD, Romero MI, Lush ME, Lu QR, Henkemeyer M, Parada LF. Ephrin-B3 is a myelin-based inhibitor of neurite outgrowth. Proc Nat Acad Sci USA. 2005;102:10694–10699. doi: 10.1073/pnas.0504021102. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bomze HM, Bulsara KR, Iskandar BJ, Caroni P, Pate Skene JH. Spinal axon regeneration evoked by replacing two growth cone proteins in adult neurons. Nat Neurosci. 2001;4:38–43. doi: 10.1038/82881. [DOI] [PubMed] [Google Scholar]
- Bonilla IE, Tanabe K, Strittmatter SM. Small proline-rich repeat protein 1A Is expressed by axotomized neurons and promotes axonal outgrowth. J Neurosci. 2002;22:1303–1315. doi: 10.1523/JNEUROSCI.22-04-01303.2002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bradbury EJ, Moon LDF, Popat RJ, King VR, Bennett GS, Patel PN, Fawcett JW, McMahon SB. Chondroitinase ABC promotes functional recovery after spinal cord injury. Nature. 2002;416:636–640. doi: 10.1038/416636a. [DOI] [PubMed] [Google Scholar]
- Brosamle C, Huber AB, Fiedler M, Skerra A, Schwab ME. Regeneration of lesioned corti-cospinal tract fibers in the adult rat induced by a recombinant, humanized in-1 antibody fragment. J Neurosci. 2000;20:8061–8068. doi: 10.1523/JNEUROSCI.20-21-08061.2000. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cafferty WB, Strittmatter SM. The Nogo-Nogo receptor pathway limits a spectrum of adult cns axonal growth. J Neurosci. 2006;26:12242–12250. doi: 10.1523/JNEUROSCI.3827-06.2006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cafferty WB, Yang S-H, Duffy PJ, Li S, Strittmatter SM. Functional axonal regeneration through astrocytic scar genetically modified to digest chondroitin sulfate proteoglycans. J Neurosci. 2007a;27:2176–2185. doi: 10.1523/JNEUROSCI.5176-06.2007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cafferty WB, Kim J-E, Lee J-K, Strittmatter SM. Response to correspondence: Kim et al. “Axon regeneration in young adult mice lacking Nogo-A/B.” Neuron 38, 187–199. Neuron. 2007b;54:195–199. doi: 10.1016/j.neuron.2007.04.005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cai D, Qiu J, Cao Z, McAtee M, Bregman BS, Filbin MT. Neuronal cyclic amp controls the developmental loss in ability of axons to regenerate. J Neurosci. 2001;21:4731–4739. doi: 10.1523/JNEUROSCI.21-13-04731.2001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cai D, Deng K, Mellado W, Lee J, Ratan RR, Filbin MT. Arginase I and polyamines act downstream from cyclic AMP in overcoming inhibition of axonal growth MAG and myelin in vitro. Neuron. 2002;35:711–719. doi: 10.1016/s0896-6273(02)00826-7. [DOI] [PubMed] [Google Scholar]
- Chan CC, Roberts CR, Steeves JD, Tetzlaff W. Aggrecan components differentially modulate nerve growth factor-responsive and neurotrophin-3-responsive dorsal root ganglion neurite growth. J Neurosci Res. 2008;86:581–592. doi: 10.1002/jnr.21522. [DOI] [PubMed] [Google Scholar]
- Chen MS, Huber AB, van der Haar ME, Frank M, Schnell L, Spillmann AA, Christ F, Schwab ME. Nogo-A is a myelin-associated neurite outgrowth inhibitor and an antigen for monoclonal antibody IN-1. Nature. 2000;403:434–439. doi: 10.1038/35000219. [DOI] [PubMed] [Google Scholar]
- Costigan M, Befort K, Karchewski L, Griffin R, D’Urso D, Allchorne A, Sitarski J, Mannion J, Pratt R, Woolf C. Replicate high-density rat genome oligonucleotide microarrays reveal hundreds of regulated genes in the dorsal root ganglion after peripheral nerve injury. BMC Neurosci. 2002;3:16. doi: 10.1186/1471-2202-3-16. [DOI] [PMC free article] [PubMed] [Google Scholar]
- David S, Aguayo AJ. Axonal elongation into peripheral nervous system “bridges” after central nervous system injury in adult rats. Science. 1981;214:931–933. doi: 10.1126/science.6171034. [DOI] [PubMed] [Google Scholar]
- Dimou L, Schnell L, Montani L, Duncan C, Simonen M, Schneider R, Liebscher T, Gullo M, Schwab ME. Nogo-A-deficient mice reveal strain-dependent differences in axonal regeneration. J Neurosci. 2006;26:5591–5603. doi: 10.1523/JNEUROSCI.1103-06.2006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Domeniconi M, Cao Z, Spencer T, Sivasankaran R, Wang KC, Nikulina E, Kimura N, Cai H, Deng K, Gao Y, He Z, Filbin MT. Myelin-associated glycoprotein interacts with the Nogo66 receptor to inhibit neurite outgrowth. Neuron. 2002;35:283–290. doi: 10.1016/s0896-6273(02)00770-5. [DOI] [PubMed] [Google Scholar]
- Dou CL, Levine JM. Inhibition of neurite growth by the NG2 chondroitin sulfate proteoglycan. J Neurosci. 1994;14:7616–7628. doi: 10.1523/JNEUROSCI.14-12-07616.1994. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Erschbamer M, Pernold K, Olson L. Inhibiting epidermal growth factor receptor improves structural, locomotor, sensory, and bladder recovery from experimental spinal cord injury. J Neurosci. 2007;27:6428–6435. doi: 10.1523/JNEUROSCI.1037-07.2007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fernandes KJ, Fan DP, Tsui BJ, Cassar SL, Tetzlaff W. Influence of the axotomy to cell body distance in rat rubrospinal and spinal motoneurons: differential regulation of GAP-43, tubulins, and neurofilament-M. J Comp Neurol. 1999;414:495–510. doi: 10.1002/(sici)1096-9861(19991129)414:4<495::aid-cne6>3.0.co;2-s. [DOI] [PubMed] [Google Scholar]
- Fournier AE, GrandPre T, Strittmatter SM. Identification of a receptor mediating Nogo-66 inhibition of axonal regeneration. Nature. 2001;409:341–346. doi: 10.1038/35053072. [DOI] [PubMed] [Google Scholar]
- Fournier AE, Takizawa BT, Strittmatter SM. Rho kinase inhibition enhances axonal regeneration in the injured CNS. J Neurosci. 2003;23:1416–1423. doi: 10.1523/JNEUROSCI.23-04-01416.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fu Q, Hue J, Li S. Nonsteroidal anti-inflammatory drugs promote axon regeneration via RhoA inhibition. J Neurosci. 2007;27:4154–4164. doi: 10.1523/JNEUROSCI.4353-06.2007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- GrandPre T, Nakamura F, Vartanian T, Strittmatter SM. Identification of the Nogo inhibitor of axon regeneration as a Reticulon protein. Nature. 2000;403:439–444. doi: 10.1038/35000226. [DOI] [PubMed] [Google Scholar]
- GrandPre T, Li S, Strittmatter SM. Nogo-66 receptor antagonist peptide promotes axonal regeneration. Nature. 2002;417:547–551. doi: 10.1038/417547a. [DOI] [PubMed] [Google Scholar]
- Hata K, Fujitani M, Yasuda Y, Doya H, Saito T, Yamagishi S, Mueller BK, Yamashita T. RGMa inhibition promotes axonal growth and recovery after spinal cord injury. J Cell Biol. 2006;173:47–58. doi: 10.1083/jcb.200508143. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hsieh SH, Ferraro GB, Fournier AE. Myelin-associated inhibitors regulate cofilin phospho-rylation and neuronal inhibition through LIM kinase and slingshot phosphatase. J Neurosci. 2006;26:1006–1015. doi: 10.1523/JNEUROSCI.2806-05.2006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hu F, Strittmatter SM. The N-terminal domain of Nogo-A inhibits cell adhesion and axonal outgrowth by an integrin-specific mechanism. J Neurosci. 2008;28:1262–1269. doi: 10.1523/JNEUROSCI.1068-07.2008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hu F, Liu BP, Budel S, Liao J, Chin J, Fournier A, Strittmatter SM. Nogo-A interacts with the Nogo-66 receptor through multiple sites to create an isoform-selective subnanomolar agonist. J Neurosci. 2005;25:5298–5304. doi: 10.1523/JNEUROSCI.5235-04.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Inatani M, Honjo M, Otori Y, Oohira A, Kido N, Tano Y, Honda Y, Tanihara H. Inhibitory effects of neurocan and phosphacan on neurite outgrowth from retinal ganglion cells in culture. Invest Ophthalmol Vis Sci. 2001;42:1930–1938. [PubMed] [Google Scholar]
- Jones LL, Margolis RU, Tuszynski MH. The chondroitin sulfate proteoglycans neurocan, brevican, phosphacan, and versican are differentially regulated following spinal cord injury. Exp Neurol. 2003;182:399–411. doi: 10.1016/s0014-4886(03)00087-6. [DOI] [PubMed] [Google Scholar]
- Kaneko S, Iwanami A, Nakamura M, Kishino A, Kikuchi K, Shibata S, Okano HJ, Ikegami T, Moriya A, Konishi O, Nakayama C, Kumagai K, Kimura T, Sato Y, Goshima Y, Taniguchi M, Ito M, He Z, Toyama Y, Okano H. A selective Sema3A inhibitor enhances regenerative responses and functional recovery of the injured spinal cord. Nat Med. 2007;12:1380–1389. doi: 10.1038/nm1505. [DOI] [PubMed] [Google Scholar]
- Kim J-E, Bonilla IE, Qiu D, Strittmatter SM. Nogo-C is sufficient to delay nerve regeneration. Mol Cell Neurosci. 2003a;23:451–459. doi: 10.1016/s1044-7431(03)00076-9. [DOI] [PubMed] [Google Scholar]
- Kim J-E, Li S, GrandPré T, Qiu D, Strittmatter SM. Axon regeneration in young adult mice lacking Nogo-A/B. Neuron. 2003b;38:187–199. doi: 10.1016/s0896-6273(03)00147-8. [DOI] [PubMed] [Google Scholar]
- Kim J-E, Liu BP, Park JH, Strittmatter SM. Nogo-66 receptor prevents raphespinal and rubrospinal axon regeneration and limits functional recovery from spinal cord injury. Neuron. 2004;44:439–451. doi: 10.1016/j.neuron.2004.10.015. [DOI] [PubMed] [Google Scholar]
- Kottis V, Thibault P, Mikol D, Xiao ZC, Zhang R, Dergham P, Braun PE. Oligodendrocyte-myelin glycoprotein (OMgp) is an inhibitor of neurite outgrowth. J Neurochem. 2002;82:1566–1569. doi: 10.1046/j.1471-4159.2002.01146.x. [DOI] [PubMed] [Google Scholar]
- Lauren J, Hu F, Chin J, Liao J, Airaksinen MS, Strittmatter SM. Characterization of myelin ligand complexes with neuronal Nogo-66 receptor family members. J Biol Chem. 2007;282:5715–5725. doi: 10.1074/jbc.M609797200. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lee J-K, Kim J-E, Sivula M, Strittmatter SM. Nogo receptor antagonism promotes stroke recovery by enhancing axonal plasticity. J Neurosci. 2004;24:6209–6217. doi: 10.1523/JNEUROSCI.1643-04.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Li S, Strittmatter SM. Delayed systemic Nogo-66 receptor antagonist promotes recovery from spinal cord injury. J Neurosci. 2003;23:4219–4227. doi: 10.1523/JNEUROSCI.23-10-04219.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Li S, Liu BP, Budel S, Li M, Ji B, Walus L, Li W, Jirik A, Rabacchi S, Choi E, Worley D, Sah DWY, Pepinsky B, Lee D, Relton J, Strittmatter SM. Blockade of Nogo-66, myelin-associated glycoprotein, and oligodendrocyte myelin glycoprotein by soluble Nogo-66 receptor promotes axonal sprouting and recovery after spinal injury. J Neurosci. 2004;24:10511–10520. doi: 10.1523/JNEUROSCI.2828-04.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Li S, Kim J-E, Budel S, Hampton TG, Strittmatter SM. Transgenic inhibition of Nogo-66 receptor function allows axonal sprouting and improved locomotion after spinal injury. Mol Cell Neurosci. 2005;29:26–39. doi: 10.1016/j.mcn.2004.12.008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Liu BP, Fournier A, GrandPre T, Strittmatter SM. Myelin-associated glycoprotein as a functional ligand for the Nogo-66 receptor. Science. 2002;297:1190–1193. doi: 10.1126/science.1073031. [DOI] [PubMed] [Google Scholar]
- Marklund N, Fulp CT, Shimizu S, Puri R, McMillan A, Strittmatter SM, McIntosh TK. Selective temporal and regional alterations of Nogo-A and small proline-rich repeat protein 1A (SPRR1A) but not Nogo-66 receptor (NgR) occur following traumatic brain injury in the rat. Exp Neurol. 2006;197:70–83. doi: 10.1016/j.expneurol.2005.08.029. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mason MR, Lieberman AR, Grenningloh G, Anderson PN. Transcriptional upregulation of SCG10 and CAP-23 is correlated with regeneration of the axons of peripheral and central neurons in vivo. Mol Cell Neurosci. 2002;20:595–615. doi: 10.1006/mcne.2002.1140. [DOI] [PubMed] [Google Scholar]
- Massey JM, Hubscher CH, Wagoner MR, Decker JA, Amps J, Silver J, Onifer SM. Chondroitinase ABC digestion of the perineuronal net promotes functional collateral sprouting in the Cuneate nucleus after cervical spinal cord injury. J Neurosci. 2006;26:4406–4414. doi: 10.1523/JNEUROSCI.5467-05.2006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- McKerracher L, David S, Jackson DL, Kottis V, Dunn RJ, Braun PE. Identification of myelin-associated glycoprotein as a major myelin-derived inhibitor of neurite growth. Neuron. 1994;13:805–811. doi: 10.1016/0896-6273(94)90247-x. [DOI] [PubMed] [Google Scholar]
- Mehta NR, Lopez PH, Vyas AA, Schnaar RL. Gangliosides and Nogo Receptors Independently Mediate Myelin-associated Glycoprotein Inhibition of Neurite Outgrowth in Different Nerve Cells. J Biol Chem. 2007;282:27875–27886. doi: 10.1074/jbc.M704055200. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Merkler D, Metz GAS, Raineteau O, Dietz V, Schwab ME, Fouad K. Locomotor recovery in spinal cord-injured rats treated with an antibody neutralizing the myelin-associated neurite growth inhibitor Nogo-A. J Neurosci. 2001;21:3665–3673. doi: 10.1523/JNEUROSCI.21-10-03665.2001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mi S, Lee X, Shao Z, Thill G, Ji B, Relton J, Levesque M, Allaire N, Perrin S, Sands B, Crowell T, Cate RL, McCoy JM, Pepinsky RB. LINGO-1 is a component of the Nogo-66 receptor/p75 signaling complex. Nat Neurosci. 2004;7:221–228. doi: 10.1038/nn1188. [DOI] [PubMed] [Google Scholar]
- Moreau-Fauvarque C, Kumanogoh A, Camand E, Jaillard C, Barbin G, Boquet I, Love C, Jones EY, Kikutani H, Lubetzki C, Dusart I, Chedotal A. The Transmembrane Semaphorin Sema4D/CD100, an inhibitor of axonal growth, is expressed on oligodendrocytes and upregulated after CNS lesion. J Neurosci. 2003;23:9229–9239. doi: 10.1523/JNEUROSCI.23-27-09229.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Morgenstern DA, Asher RA, Fawcett JW. Chondroitin sulphate proteoglycans in the CNS injury response. Prog Brain Res. 2002;137:313–332. doi: 10.1016/s0079-6123(02)37024-9. [DOI] [PubMed] [Google Scholar]
- Neumann S, Woolf CJ. Regeneration of dorsal column fibers into and beyond the lesion site following adult spinal cord injury. Neuron. 1999;23:83–91. doi: 10.1016/s0896-6273(00)80755-2. [DOI] [PubMed] [Google Scholar]
- Neumann S, Bradke F, Tessier-Lavigne M, Basbaum AI. Regeneration of sensory axons within the injured spinal cord induced by intraganglionic cAMP elevation. Neuron. 2002;34:885–893. doi: 10.1016/s0896-6273(02)00702-x. [DOI] [PubMed] [Google Scholar]
- Niederost B, Oertle T, Fritsche J, McKinney RA, Bandtlow CE. Nogo-A and myelin-associated glycoprotein mediate neurite growth inhibition by antagonistic regulation of RhoA and Rac1. J Neurosci. 2002;22:10368–10376. doi: 10.1523/JNEUROSCI.22-23-10368.2002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nikulina E, Tidwell JL, Dai HN, Bregman BS, Filbin MT. The phosphodiesterase inhibitor rolipram delivered after a spinal cord lesion promotes axonal regeneration and functional recovery. Proc Natl Acad Sci USA. 2004;101:8786–8790. doi: 10.1073/pnas.0402595101. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Park JB, Yiu G, Kaneko S, Wang J, Chang J, He Z. A TNF receptor family member, TROY, is a coreceptor with Nogo receptor in mediating the inhibitory activity of myelin inhibitors. Neuron. 2005;45:345–351. doi: 10.1016/j.neuron.2004.12.040. [DOI] [PubMed] [Google Scholar]
- Pearse DD, Pereira FC, Marcillo AE, Bates ML, Berrocal YA, Filbin MT, Bunge MB. cAMP and Schwann cells promote axonal growth and functional recovery after spinal cord injury. Nat Med. 2004;10:610–616. doi: 10.1038/nm1056. [DOI] [PubMed] [Google Scholar]
- Qiu J, Cai D, Dai H, McAtee M, Hoffman PN, Bregman BS, Filbin MT. Spinal axon regeneration induced by elevation of cyclic AMP. Neuron. 2002;34:895–903. doi: 10.1016/s0896-6273(02)00730-4. [DOI] [PubMed] [Google Scholar]
- Raivich G, Bohatschek M, Da Costa C, Iwata O, Galiano M, Hristova M, Nateri AS, Makwana M, Ls R-S, Wolfer DP, Lipp H-P, Aguzzi A, Wagner EF, Behrens A. The AP-1 transcription factor c-Jun is required for efficient axonal regeneration. Neuron. 2004;43:57–67. doi: 10.1016/j.neuron.2004.06.005. [DOI] [PubMed] [Google Scholar]
- Richardson PM, McGuinness UM, Aguayo AJ. Axons from CNS neurons regenerate into PNS grafts. Nature. 1980;284:264–265. doi: 10.1038/284264a0. [DOI] [PubMed] [Google Scholar]
- Richardson PM, Issa VM, Aguayo AJ. Regeneration of long spinal axons in the rat. J Neurocytol. 1984;13:165–182. doi: 10.1007/BF01148324. [DOI] [PubMed] [Google Scholar]
- Schmalfeldt M, Bandtlow CE, Dours-Zimmermann MT, Winterhalter KH, Zimmermann DR. Brain derived versican V2 is a potent inhibitor of axonal growth. J Cell Sci. 2000;113:807–816. doi: 10.1242/jcs.113.5.807. [DOI] [PubMed] [Google Scholar]
- Schmandke A, Schmandke A, Strittmatter SM. ROCK and Rho: biochemistry and neuronal functions of Rho-associated protein kinases. Neuroscientist. 2007;13:454–469. doi: 10.1177/1073858407303611. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Seijffers R, Allchorne AJ, Woolf CJ. The transcription factor ATF-3 promotes neurite outgrowth. Mol Cell Neurosci. 2006;32:143–154. doi: 10.1016/j.mcn.2006.03.005. [DOI] [PubMed] [Google Scholar]
- Seymour AB, Andrews EM, Tsai S-Y, Markus TM, Bollnow MR, Brenneman MM, O’Brien TE, Castro AJ, Schwab ME, Kartje GL. Delayed treatment with monoclonal antibody IN-1 1 week after stroke results in recovery of function and corticorubral plasticity in adult rats. J Cereb Blood Flow Metab. 2005;25:1366–1375. doi: 10.1038/sj.jcbfm.9600134. [DOI] [PubMed] [Google Scholar]
- Shao Z, Browning JL, Lee X, Scott ML, Shulga-Morskaya S, Allaire N, Thill G, Levesque M, Sah D, McCoy JM, Murray B, Jung V, Pepinsky RB, Mi S. TAJ/TROY, an orphan TNF receptor family member, binds Nogo-66 receptor 1 and regulates axonal regeneration. Neuron. 2005;45:353–359. doi: 10.1016/j.neuron.2004.12.050. [DOI] [PubMed] [Google Scholar]
- Simonen M, Pedersen V, Weinmann O, Schnell L, Buss A, Ledermann B, Christ F, Sansig G, van der Putten H, Schwab ME. Systemic deletion of the myelin-associated outgrowth inhibitor Nogo-A improves regenerative and plastic responses after spinal cord injury. Neuron. 2003;38:201–211. doi: 10.1016/s0896-6273(03)00226-5. [DOI] [PubMed] [Google Scholar]
- Sivasankaran R, Pei J, Wang KC, Zhang YP, Shields CB, Xu X-M, He Z. PKC mediates inhibitory effects of myelin and chondroitin sulfate proteoglycans on axonal regeneration. Nat Neurosci. 2004;7:261–268. doi: 10.1038/nn1193. [DOI] [PubMed] [Google Scholar]
- Steward O, Zheng B, Banos K, Yee KM. Response to: Kim et al. “Axon regeneration in young adult mice lacking Nogo-A/B.” Neuron 38, 187–199. Neuron. 2007;54:191–195. doi: 10.1016/j.neuron.2007.04.004. [DOI] [PubMed] [Google Scholar]
- Tanabe K, Bonilla I, Winkles JA, Strittmatter SM. Fibroblast growth factor-inducible-14 is induced in axotomized neurons and promotes neurite outgrowth. J Neurosci. 2003;23:9675–9686. doi: 10.1523/JNEUROSCI.23-29-09675.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Venkatesh K, Chivatakarn O, Lee H, Joshi PS, Kantor DB, Newman BA, Mage R, Rader C, Giger RJ. The Nogo-66 receptor homolog NgR2 is a sialic acid-dependent receptor selective for myelin-associated glycoprotein. J Neurosci. 2005;25:808–822. doi: 10.1523/JNEUROSCI.4464-04.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Vyas AA, Patel HV, Fromholt SE, Heffer-Lauc M, Vyas KA, Dang J, Schachner M, Schnaar RL. Gangliosides are functional nerve cell ligands for myelin-associated glycoprotein (MAG), an inhibitor of nerve regeneration. Proc Natl Acad Sci USA. 2002;99:8412–8417. doi: 10.1073/pnas.072211699. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wang KC, Kim JA, Sivasankaran R, Segal R, He Z. p75 interacts with the Nogo receptor as a co-receptor for Nogo, MAG and OMgp. Nature. 2002;420:74–78. doi: 10.1038/nature01176. [DOI] [PubMed] [Google Scholar]
- Wiessner C, Bareyre FM, Allegrini PR, Mir AK, Frentzel S, Zurini M, Schnell L, Oertle T, Schwab ME. Anti-Nogo-A antibody infusion 24 hours after experimental stroke improved behavioral outcome and corticospinal plasticity in normotensive and spontaneously hypertensive rats. J Cereb Blood Flow Metab. 2003;23:154–165. doi: 10.1097/01.WCB.0000040400.30600.AF. [DOI] [PubMed] [Google Scholar]
- Yamada H, Fredette B, Shitara K, Hagihara K, Miura R, Ranscht B, Stallcup WB, Yamaguchi Y. The brain chondroitin sulfate proteoglycan brevican associates with astrocytes ensheathing cerebellar glomeruli and inhibits neurite outgrowth from granule neurons. J Neurosci. 1997;17:7784–7795. doi: 10.1523/JNEUROSCI.17-20-07784.1997. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Z’Graggen WJ, Fouad K, Raineteau O, Metz GAS, Schwab ME, Kartje GL. Compensatory sprouting and impulse rerouting after unilateral pyramidal tract lesion in neonatal rats. J Neurosci. 2000;20:6561–6569. doi: 10.1523/JNEUROSCI.20-17-06561.2000. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zheng B, Ho C, Li S, Keirstead H, Steward O, Tessier-Lavigne M. Lack of enhanced spinal regeneration in nogo-deficient mice. Neuron. 2003;38:213–224. doi: 10.1016/s0896-6273(03)00225-3. [DOI] [PubMed] [Google Scholar]