

Abstract
1-Trichloromethyl-1,2,3,4-tetrahydro-β-carboline (TaClo) is neurotoxic when administered to the brain and alters motor behaviour following intraperitoneal administration. We have assessed the long-term effects of oral TaClo administration on nocturnal motor behaviour in rats. Two groups of rats received TaClo orally at a dose of either 0.2 or 0.4 mg/kg twice daily for 7 weeks. The control group was given saline. No change in locomotor activity was observed 4–9 days after the end of the 7-week administration of TaClo. In addition, the spontaneous motor activity was altered dose-dependently 9 months after oral TaClo administration, with an increase in the low-dose TaClo group and a decrease in the high-dose group. Oral administration of TaClo in rats may be useful in investigating the hypothesis that in Parkinson’s disease, an unknown pathogenic factor crossing the intestinal mucosa barrier can induce neurodegenerative processes eventually affecting the entire brain.
Keywords: 1-Trichloromethyl-1,2,3,4-tetrahydro-β-carboline; TaClo; Oral administration; Motor behaviour; Nocturnal activity; Rat; Parkinson’s disease
Introduction
Since the discovery that the neurotoxin 1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine (MPTP) can induce parkinsonian symptoms in primates including man (Burns et al. 1983; Davis et al. 1979) it has been suggested that environmental toxins may play a role in the aetiology and pathogenesis of Parkinson’s disease. The search for structural analogues of MPTP in the environment has identified several compounds such as herbicides (e.g. paraquat), alkaloids (e.g. 1,2,3,4-tetrahydroisoquinolines) and β-carbolines. The neurotoxin 1-trichloromethyl-1,2,3,4-tetrahydro-β-carboline (TaClo) belongs to the group of β-carbolines and bears striking similarities to MPTP, a neurotoxin that is widely being used to produce an animal model of Parkinson’s disease. TaClo is readily formed under physiological conditions from the biogenic amine tryptamine (Ta) and trichloroacetaldehyde (chloral, Clo; Bringmann and Hille 1990). Chloral can be formed in vivo following exposure to the industrial solvent trichloroethylene (TRI). An association between the exposure to trichloroethylene and Parkinson’s disease in humans has been observed (Kochen et al. 2003).
Behavioural consequences of TaClo administration have been demonstrated (Sontag et al. 1995). Chronic treatment of rats with TaClo induced an acute behavioural supersensitivity to the dopaminergic agonist apomorphine. This supersensitivity is not permanent and is later replaced by reduced motor activity when TaClo is completely metabolized (Heim and Sontag 1997). These findings suggest a TaClo-induced progressive degeneration of the dopaminergic system which is probably mediated via the formation of free radicals or the inhibition of complex I.
The neurodegenerative process underlying Parkinson’s disease involves not only neurons in the central nervous system but also in the enteric and peripheral nervous systems. It has been suggested that an unidentified pathogenic factor enters the enteric nervous system and is then retrogradely transferred to the central nervous system where it induces the characteristic patterns of neurodegeneration in Parkinson’s disease (Braak et al. 2003). This factor might cross the mucosal barrier of the intestine and could then be incorporated into the axon terminal of the vagus nerve and transported in a retrograde manner to the vagus nucleus. The question arises whether TaClo could be such an unknown pathogenetic factor.
TaClo is neurotoxic when administered to the brain and alters motor behaviour following intraperitoneal administration. The hypothesis by Braak et al. (2003) suggests that TaClo may exert its neurotoxic effects by crossing the mucosal barrier. We have therefore assessed the effects of oral TaClo administration on motor behaviour in rats.
Materials and methods
Twenty-seven male Wistar rats aged three to 4 months were used. The animals were kept on a 12:12 h light/dark circle (room temperature 21°C, humidity 55%). The rats were divided in three groups. The two TaClo groups received TaClo diluted in saline at a dose of 0.2 mg/kg (N = 9) or 0.4 mg/kg (N = 8) twice daily for 7 weeks. The control animals (N = 10) were given saline twice daily for 7 weeks. Both TaClo and saline were administered orally at about 11 a.m. and 5 p.m.
In order to assess the acute effects of TaClo administration, the rats’ spontaneous motor activity was measured for 1 h between 8 and 12 a.m. Nine months after TaClo or saline administration, the spontaneous motor activity of all rats was recorded during the whole night. Motor activity was automatically measured using an Autotrack System (Columbus Instruments, Columbus OH, USA). The following parameters were measured using the Autotrack System: (1) distance travelled, i.e. the total distance (in cm) a rat walked during the recording; (2) stereotype time, i.e. the time (in sec) a rat spent with movements covering less than 4.8 cm; and (3) burst of stereotype movements, i.e. the total number of movements covering less than 4.8 cm. Data were recorded over 1-h intervals.
The rats were placed in the Autotrack System at 1 p.m. in order to allow them to adapt to the experimental environment. The recording of motor activity started at 8 p.m. and lasted for 10 h. The light was turned off at 6 p.m. and turned on at 6 a.m. During the testing the rats had free access to food and water.
The comparison of the rats’ activity between the groups was performed both for the total of 10 h and for each hour separately. Statistical analysis was performed using analysis of variance and the Student–Newman–Keuls test. For statistical analysis an alpha level of 0.05 was applied. All statistical analyses were carried out using the Statistical Package for Social Sciences 12.0 for Windows.
All experiments were performed in accordance with national laws (German law on Protection of Animals) and the principles of laboratory animal care (NIH publication No. 86-23, revised 1985). None of the animals died during the experiment.
Results
Spontaneous motor activity 4–9 days after the end of TaClo administration
Distance travelled (Data not shown)
With regard to the distance travelled no statistically significant differences between the groups were found (F2,29 = 1.2; p > 0.05).
Spontaneous nocturnal motor activity 9 months after the end of TaClo administration
Separate analysis for each 1-h interval
Distance travelled (Fig. 1a)
Fig. 1.
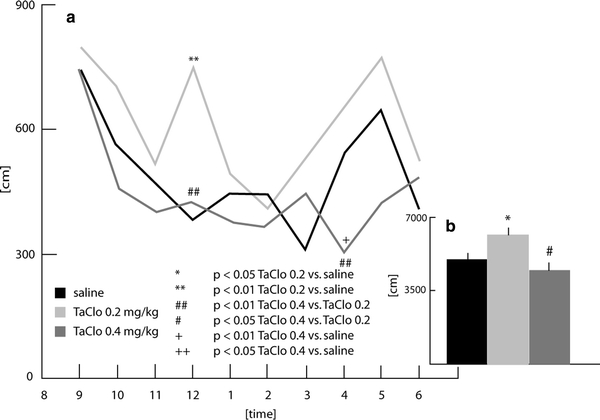
Distance travelled by the rats 9 months after the end of oral TaClo administration
In the 1-h interval ending at midnight, the three groups differed with regard to the distance travelled (F2,26 = 8.89; p < 0.01). Post-hoc analysis showed that the group treated with 0.2 mg/kg TaClo displayed a significant increase in activity compared to both the other TaClo group (0.4 mg/kg) and the saline group (p < 0.01). In the interval ending at 4 a.m., the groups differed in their activity (F2,26 = 5.45; p < 0.05). The TaClo-0.4 group showed significantly less activity than the TaClo-0.2 (Student–Newman–Keuls test, p < 0.01) and the saline groups (p < 0.01). The remaining comparisons failed to reach significance.
Stereotype time (Fig. 2a)
Fig. 2.
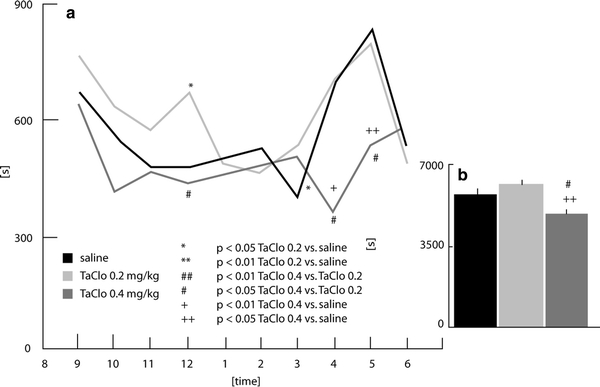
Time spent with stereotype movements in rats 9 months after the end of oral TaClo administration
The three groups differed in the interval ending at midnight (F2,26 = 5.98; p < 0.01). Post-hoc analysis revealed that the TaClo-0.2 group spent more time with stereotype movements than the control group (p < 0.05) and the TaClo-0.4 group (p < 0.05). Differences between groups were also found in the 1-h intervals ending at 4 a.m. (F2,26 = 6.17, p < 0.05) and 5 a.m. (F2,26 = 5.27, p < 0.05). The TaClo-0.4 group showed a significantly reduced stereotype time compared to the other groups. (Student–Newman–Keuls test; at 4 a.m.: TaClo-0.4 vs. saline p < 0.01, TaClo-0.4 vs. TaClo-0.2 p < 0.05; at 5 a.m.: TaClo-0.4 vs. saline p < 0.05, TaClo-0.4 vs. TaClo-0.2 p < 0.05). The other comparisons failed to reach significance.
Burst of stereotype movement (Fig. 3a)
Fig. 3.
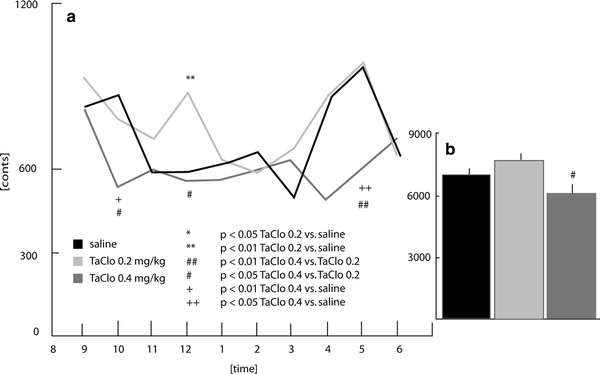
Burst of stereotype movements in rats 9 months after the end of oral TaClo administration
A difference between groups was observed in the interval ending at 10 p.m. (F2,26 = 6.3, p < 0.01), the TaClo-0.4 group showed significantly fewer bursts than the saline group (p < 0.01) and the TaClo-0.2 group (p < 0.05). A group difference at midnight (F2,26 = 5.75, p < 0.01) was characterised by a significant increase in bursts in the TaClo-0.2 group compared to the saline (p < 0.01) and TaClo-0.4 groups (p < 0.05).
Analysis for a total of 10 h
Distance travelled (Fig. 1b)
A significant difference between the groups was observed (F2,26 = 4.71, p < 0.05). Post-hoc analysis using Student–Newman–Keuls test showed that the TaClo-0.2 rats walked more than those of the saline (p < 0.05) and TaClo-0.4 groups (p < 0.05).
Stereotype time (Fig. 2b)
The time spent with stereotype movements differed between the groups (F2,26 = 6.38, p < 0.01). Post-hoc analysis revealed that the TaClo-0.4 group spent less time with stereotype movements than both the control group (p > 0.05) and the TaClo-0.2 group (p < 0.01).
Burst of stereotype movements (Fig. 3b)
The number of stereotype movements differed between the groups (F2,26 = 5.43, p < 0.01) with the TaClo-0.4 group displaying significantly fewer movements than the TaClo-0.2 group (p < 0.01).
Discussion
The aim of the present study was to investigate the long-term effects of oral administration of the neurotoxin TaClo on the spontaneous nocturnal activity of rats. Previous studies have shown that TaClo is neurotoxic to the dopaminergic system (for review see Riederer et al. 2002) and induces changes of the motor behaviour of rats (Heim and Sontag 1997; Sontag et al. 1995). In these studies, however, TaClo was injected intraperitoneally, while the neurotoxin was administered orally in the present study. This approach was used against the background of previous studies suggesting that the pathogenesis of Parkinson’s disease begins in the enteric nervous system and is induced by an unknown pathogenic factor which passes the gastrointestinal mucosal barrier (Braak et al. 2003). TaClo might be such a factor causing neurodegeneration following oral administration.
The present study has demonstrated for the first time that oral TaClo administration affects the spontaneous motor activity of rats during the night. The rats treated with 0.2 mg/kg of TaClo walked longer distances than both the control animals and the rats treated with 0.4 mg/kg of TaClo (Fig. 1b). In addition, only the TaClo-0.2 group showed an activity peak around midnight (Fig. 1a). This data shows that the treatment of rats with TaClo-0.2 over 7 weeks increases the distance travelled and changes the nocturnal activity pattern in rats. These results refer to locomotor activity. Non-locomotor activities such as stereotype movements were also shown to be altered following TaClo administration (Figs. 2a, 3a). The TaClo-0.4 group spent significantly less time with stereotype movements and the burst of stereotype movements was reduced, indicating a general reduction of motor activity.
These results show an increase in motor activity in the low-dose TaClo group and a decrease in the high-dose group at 9 months after the end of the treatment. However, a previous study (Sontag et al. 1995) showed a reduced activity in female rats 9 weeks after the end of a 7-week treatment with TaClo (0.2 mg/kg intraperitoneally). One explanation for the different results could be the gender of the rats used since sex-related differences in the metabolism of trichlorethylene (a precursor of TaClo) in the liver have been found (Elfarra et al. 1998). However, it is more likely that the way of TaClo administration contributes to these differences. Previous data indicate a slow neurodegenerative effect in rats following the intraperitoneal treatment with TaClo. In a previous study (Heim and Sontag 1997), the spontaneous locomotor activity as measured for 1 h in the morning was increased 4–9 days after a daily injection of TaClo-0.2 over 7 weeks. However, 9 months after TaClo administration this difference could not be observed. It has been suggested that dopamine receptors are upregulated as an acute response to the degenerative process induced by TaClo. In the present study, an increase in locomotor activity was not observed 4–9 days after the end of treatment. This suggests that orally administered TaClo does not induce neurodegenerative effects at this point in time. However, 9 months later the total nocturnal activity is decreased in the high-dose TaClo group indicating reduced dopaminergic functioning. Both oral and intraperitoneal administration appears to have similar behavioural effects on the motor system of rats, with a later onset following oral administration. The TaClo-0.4 group showed no marked differences to the control group. However, on the basis of the data of the TaClo-0.2 group one can conclude that oral administration was effective and caused neurodegenerative processes producing reduced motor activity later on (Heim and Sontag 1997). It is possible that the TaClo-0.4 group had developed reduced motor activity, i.e. the TaClo-induced degeneration developed faster in the TaClo-0.4 group than in the TaClo-0.2 group. The difference between the two TaClo groups suggests therefore a dose-dependent effect of TaClo.
Furthermore, not only the amount of motor activity is affected but also the pattern of activity (Figs. 1a, 2a, 3a). This might hint to a disturbed sleeping pattern which can also be observed Parkinson’s disease (Partinen 1997). Stereotactically administered TaClo has been shown to induce the release of both dopamine and serotonin (Gerlach et al. 1998). In addition, it has been shown that TaClo reduces serotonin uptake in the human serotonergic cell line JAR (Bringmann et al. 2000). Since serotonin is known to be involved in the regulation of sleep, the present findings could indicate that TaClo affects serotonin-related behaviours in rats.
In summary, the present study has demonstrated that TaClo administered orally induces alterations in spontaneous motor behaviour which are similar to those observed following intraperitoneal administration. These findings suggest that TaClo can induce neurodegeneration following both intraperitoneal and oral administration. Oral administration of TaClo in rats may be useful in investigating the hypothesis by Braak et al. (2002) that an unknown pathogenic factor crossing the mucosa barrier can induce neurodegenerative processes which eventually affect the entire brain.
References
- Braak H, Del TK, Bratzke H, Hamm-Clement J, Sandmann-Keil D, Rub U. Staging of the intracerebral inclusion body pathology associated with idiopathic Parkinson’s disease (preclinical and clinical stages) J Neurol. 2002;249(Suppl 3):III/1–III/5. doi: 10.1007/s00415-002-1301-4. [DOI] [PubMed] [Google Scholar]
- Braak H, Del TK, Rub U, Vos RA, Jansen Steur EN, Braak E. Staging of brain pathology related to sporadic Parkinson’s disease. Neurobiol Aging. 2003;24:197–211. doi: 10.1016/S0197-4580(02)00065-9. [DOI] [PubMed] [Google Scholar]
- Bringmann G, Hille A. Endogenous alkaloids in man, VII: 1-trichloromethyl-1, 2, 3, 4-tetrahydro-beta-carboline—a potential chloral-derived indol alkaloid in man. Arch Pharm (Weinheim) 1990;323:567–569. doi: 10.1002/ardp.19903230903. [DOI] [PubMed] [Google Scholar]
- Bringmann G, Bruckner R, Mossner R, Feineis D, Heils A, Lesch KP. Effect of 1-trichloromethyl-1, 2, 3, 4-tetrahydro-beta-carboline (TaClo) on human serotonergic cells. Neurochem Res. 2000;25:837–843. doi: 10.1023/A:1007521625088. [DOI] [PubMed] [Google Scholar]
- Burns RS, Chiueh CC, Markey SP, Ebert MH, Jacobowitz DM, Kopin IJ. A primate model of parkinsonism: selective destruction of dopaminergic neurons in the pars compacta of the substantia nigra by N-methyl-4-phenyl-1,2,3,6-tetrahydropyridine. Proc Natl Acad Sci USA. 1983;80:4546–4550. doi: 10.1073/pnas.80.14.4546. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Davis GC, Williams AC, Markey SP, Ebert MH, Caine ED, Reichert CM, Kopin IJ. Chronic Parkinsonism secondary to intravenous injection of meperidine analogues. Psychiatry Res. 1979;1:249–254. doi: 10.1016/0165-1781(79)90006-4. [DOI] [PubMed] [Google Scholar]
- Elfarra AA, Krause RJ, Last AR, Lash LH, Parker JC. Species- and sex-related differences in metabolism of trichloroethylene to yield chloral and trichloroethanol in mouse, rat, and human liver microsomes. Drug Metab Dispos. 1998;26:779–785. [PubMed] [Google Scholar]
- Gerlach M, Xiao AY, Heim C, Lan J, God R, Feineis D, Bringmann G, Riederer P, Sontag KH. 1-Trichloromethyl-1,2,3,4-tetrahydro-beta-carboline increases extracellular serotonin and stimulates hydroxyl radical production in rats. Neurosci Lett. 1998;257:17–20. doi: 10.1016/S0304-3940(98)00791-5. [DOI] [PubMed] [Google Scholar]
- Heim C, Sontag KH. The halogenated tetrahydro-beta-carboline “TaClo”: a progressively-acting neurotoxin. J Neural Transm Suppl. 1997;50:107–111. [PubMed] [Google Scholar]
- Kochen W, Kohlmuller D, Biasi P, Ramsay R. The endogeneous formation of highly chlorinated tetrahydro-beta-carbolines as a possible causative mechanism in idiopathic Parkinson’s disease. Adv Exp Med Biol. 2003;527:253–263. doi: 10.1007/978-1-4615-0135-0_29. [DOI] [PubMed] [Google Scholar]
- Partinen M. Sleep disorder related to Parkinson’s disease. J Neurol. 1997;244:S3–S6. doi: 10.1007/BF03160564. [DOI] [PubMed] [Google Scholar]
- Riederer P, Foley P, Bringmann G, Feineis D, Bruckner R, Gerlach M. Biochemical and pharmacological characterization of 1-trichloromethyl-1,2,3,4-tetrahydro-beta-carboline: a biologically relevant neurotoxin? Eur J Pharmacol. 2002;442:1–16. doi: 10.1016/S0014-2999(02)01308-0. [DOI] [PubMed] [Google Scholar]
- Sontag KH, Heim C, Sontag TA, God R, Reichmann H, Wesemann W, Rausch WD, Riederer P, Bringmann G. Long-term behavioural effects of TaClo (1-trichloromethyl-1,2,3,4-tetrahydro-beta-carboline) after subchronic treatment in rats. J Neural Transm. 1995;46(Suppl):283–289. [PubMed] [Google Scholar]