

Abstract
Cellular signaling via epidermal growth factor (EGF) and EGF-like ligands can determine cell fate and behavior. Osteoblasts, which are responsible for forming and mineralizing osteoid, express EGF receptors and alter rates of proliferation and differentiation in response to EGF receptor activation. Transgenic mice over-expressing the EGF-like ligand betacellulin (BTC) exhibit increased cortical bone deposition; however, because the transgene is ubiquitously expressed in these mice, the identity of cells affected by BTC and responsible for increased cortical bone thickness remains unknown. We have therefore examined the influence of BTC upon mesenchymal stem cell (MSC) and pre-osteoblast differentiation and proliferation. BTC decreases the expression of osteogenic markers in both MSCs and pre-osteoblasts; interestingly, increases in proliferation require hypoxia-inducible factor-alpha (HIF-α), as an HIF antagonist prevents BTC-driven proliferation. Both MSCs and pre-osteoblasts express EGF receptors ErbB1, ErbB2, and ErbB3, with no change in expression under osteogenic differentiation. These are the first data that demonstrate an influence of BTC upon MSCs and the first to implicate HIF-α in BTC-mediated proliferation.
Keywords: Stem cell, Osteoblast, Differentiation, Proliferation, Betacellulin, Cell culture, Human
Introduction
The skeleton continually undergoes alterations in size and shape in response to humoral, mechanical, and paracrine stimuli. Degradation of pre-existing bone by osteoclasts and ensuing replacement with osteoid by osteoblasts require concerted activity in order to maintain skeletal homeostasis and to prevent pathologic conditions such as osteoporosis or osteopetrosis. Osteoblast number is partially governed by the proliferation and osteogenic differentiation of mesenchymal stem cell (MSC) precursors; hence, stimuli that enhance MSC number or osteogenic differentiation emerge as ideal methods to combat bone loss resulting from pharmacology or aging.
Epidermal growth factor (EGF) mediates diverse processes of proliferation, differentiation, adhesion, and survival (Citri and Yarden 2006; Yarden and Sliwkowski 2001). The EGF family of ligands consists of EGF, amphiregulin, betacellulin (BTC), epiregulin, epigen, heparin-binding EGF-like growth factor (HB-EGF), and transforming growth factor-alpha (TGF-α; Harris et al. 2003; Xian 2007). EGF signaling occurs through ErbB receptor tyrosine kinases, of which ErbB1/EGF-R is bound by all seven family members. Three members (EGF, amphiregulin, and TGF-α) can only bind to ErbB1, whereas BTC, epiregulin, and HB-EGF can also bind to ErbB4 to form ErbB1/ErbB4 heterodimers. ErbB2, although lacking any known cognate ligand, is the preferred co-receptor for ErbB1, and the heterodimerization of ErbB2 and ErbB1 generates more potent signaling than ErbB1 homodimers (Qin et al. 2005; Yarden 2001; Yarden and Sliwkowski 2001); ErbB3 is a pseudokinase and therefore lacks any traditional catalytic kinase domain. Once bound by EGF or EGF-like ligand, homo- or heterodimerized ErbB receptors initiate multiple signaling pathways that affect proliferation, survival, and differentiation and that are mediated Ras-Raf-mitogen-activated protein kinase (Ras-Raf-MAPK; Yarden and Sliwkowski 2001), the phosphatidyl-inositol-3-kinase/Akt pathway (Xian 2007), and phospholipase C-γ/protein kinase C (Yarden and Sliwkowski 2001).
EGF and related ligands exert disparate effects upon osteoblast differentiation and proliferation. In vitro studies have revealed that EGF inhibits bone nodule formation (Antosz et al. 1987) and decreases alkaline phosphatase (ALP) activity (Nicolas et al. 1990). Similar findings have been reported for osteoblastic cells treated with amphiregulin (Qin et al. 2005). Further, over-expression of ErbB1 in ROS rat osteosarcoma cells decreases the expression of osteoblastic markers (Chien et al. 2000). Comparable results are observed with in vivo models. EGF transgenic mice, under the control of a β-actin promoter, demonstrate a 20%–50% reduction in body weight (Wong et al. 2000) and decreased cortical bone thickness compared with wild-type controls (Chan and Wong 2000). Similar observations, including impaired longitudinal growth and overall disproportionate skeletal growth, are detected in mice transgenic for BTC (Schneider et al. 2005). Whereas EGF suppresses the osteoblastic phenotype, it demonstrates mitogenic effects upon osteoblastic cells (Loza et al. 1995; Nicolas et al. 1990; VandePol et al. 1989), as do amphiregulin and TGF-α (Qin et al. 2005). Thus, a general theme regarding EGF signaling in bone seems to be increased proliferation and decreased osteogenic differentiation. Schneider et al. (2009) have recently demonstrated that BTC transgenic mice have enhanced cortical, but reduced trabecular, bone thickness. To date, however, no study has examined the influence of BTC upon osteoprogenitor proliferation or differentiation. We report that BTC attenuates the osteogenic differentiation of MSC and committed pre-osteoblastic cells, whilst simultaneously increasing their proliferation through a mechanism involving the stabilization of hypoxia-inducible factor (HIF). These are the first data to demonstrate an influence of BTC upon MSC and the first to implicate HIF in BTC-mediated proliferation.
Materials and methods
Cell culture
Human MSCs (hMSCs) were purchased from Lonza; cells were isolated from iliac crest bone biopsies from a 19-year-old male African-American, a 20-year-old male African-American, and an 18-year-old male Caucasian with no known pathologic conditions. hMSCs (passages 2–6) were cultured in α-MEM supplemented with 10% fetal bovine serum (FBS; Invitrogen) and 1% penicillin/streptomycin (P/S; Invitrogen) and maintained in a humidified incubator at 37°C with 5% CO2. MC3T3-E1, clone 14, pre-osteoblastic cells were purchased from American Type Culture Collection and cultured in α-MEM supplemented with 10% FBS and 1% P/S and maintained in a humidified incubator at 37°C with 5% CO2. Cells were sub-cultured with 0.05% trypsin/EDTA when 80% confluent.
Osteogenic differentiation
hMSCs or MC3T3-E1 cells were seeded into 6-well plates at a density of 3000 cells/cm2. When 70% confluent, media was replaced with osteogenic differentiation media consisting of 10% FBS, 1% P/S, 5 mM β-glycerophosphate acid (VWR International) and 50 µg/ml L-ascorbic acid 2-phosphate (Waco) in α-MEM (Invitrogen).
RNA extraction and quantitative polymerase chain reaction
Total RNA was collected with RNeasy Mini kit (Qiagen) according to the manufacturer’s instructions and including on-column digestion of genomic DNA. RNA quality was assessed by the ratio of absorbance at 260 nm and 280 nm. Total RNA (0.2–1 μg) was reverse-transcribed with a Superscript First-Strand Synthesis System (Invitrogen). Quantitative reverse transcription with the polymerase chain reaction (RT-PCR) was performed by using the TaqMan Universal PCR Master Mix (Applied Biosystems) on a Mastercycler realplex2 (Eppendorf); proprietary primers and probes, whose sequences are not disclosed, were purchased from Applied Biosystems. Amplification conditions were 50°C for 2 min, 95°C for 10 min, followed by 40 cycles at 95°C for 15 s and 60°C for 30 s. Quantitative PCR results were first normalized to ribosomal protein 13 (Rpl13) transcript level to yield ΔCt. Results are expressed as 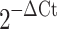 or
or 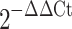 , after Livak and Schmittgen (2001).
, after Livak and Schmittgen (2001).
ALP activity assay
Whole cell protein lysates were collected on days 3, 7, or 21 in 0.1% Triton X-100, 10 mM TRIS pH 8, 1 mM EDTA, and protease inhibitor cocktail (Calbiochem). Lysates (10 μl) were incubated with p-nitrophenol phosphate (Sigma) and incubated at 37°C for 30 min. The reaction was terminated with the addition of 0.02 N NaOH, and activity was recorded by measuring absorbance (450 nm) on a microplate reader.
Proliferation assays
Cells were seeded into 60-mm dishes at a density of 9500/cm2 and cultured for 2 days, after which the medium was removed and replaced with BTC (0.5–500 ng/ml) in reduced-serum (2% FBS) medium. As a positive control for proliferation, a subset of cells was cultured in high-serum (20% FBS) medium in the absence of BTC. Cells were collected 4 days later by trypsinization and counted by using a Countess automated cell counter (Invitrogen). To determine the influence of HIF-α isoforms upon BTC-mediated proliferation, cells were treated with 10–25 μM YC-1, a small molecule post-transcriptional antagonist of HIF-1α accumulation (Chun et al. 2004). Proliferation under these conditions was assessed as above.
Western immunoblotting
Cells were briefly washed with phosphate-buffered saline, and whole-cell protein lysates were collected in 0.1% Triton X-100, 10 mM TRIS pH 8, 1 mM EDTA, supplemented with a protease and phosphatase inhibitor cocktail (Calbiochem). Samples were electrophoresed onto 7.5% TRIS-HCl gels (Bio-Rad), resolved onto 0.2-μm nitrocellulose membranes, and blocked in non-fat milk in TRIS-buffered saline supplemented with 0.1% Tween-20. Immunoreactivity against HIF-1α (1:200; Cayman Chemical) or β-actin (1:1000; Abcam) was visualized with appropriate horseradish-peroxidase-conjugated antibodies (Jackson Immunoresearch) and ECL Plus Western Blotting Detection Reagents (GE Healthcare). Densitometric analysis was performed with Adobe Photoshop.
Statistical analysis
The experimental design involved repeated measures analysis or dose-response comparisons with negative or positive controls. The data were assessed for normality with the Shapiro Wilk test, and analysis of variance was performed by using Proc GLM (SAS 9.1.2, SAS Institute). Either Tukey’s or Dunnet’s tests were used for post-hoc comparisons. Student’s t-test was used for pair-wise comparisons of the least square means to examine interaction effects. Statistical analyses and the type I error was maintained at <0.05 for all comparisons. Data are reported as arithmetic means ± SE or fold-change to BTC-free controls ± SE.
Results
BTC inhibits osteogenic differentiation of mesenchymal and pre-osteoblastic cells
We first examined the influence of BTC upon MSC differentiation toward the osteogenic lineage. hMSCs were cultured under osteogenic conditions and 0.5–500 ng/ml BTC for 3, 7, 14, or 21 days. ALP activity increased with time in culture in BTC-free controls, indicating progression of hMSCs toward an osteoblastic phenotype (Fig. 1a). We observed a trend for attenuation of ALP activity compared with BTC-free controls at all time points and in all concentrations of BTC. The difference in ALP activity between BTC-free controls and cells under treatment were significant on day 7 (5, 50, or 500 ng/ml) and day 21 (5 and 500 ng/ml; Fig. 1a). BTC-treated cells revealed significant increases in ALP activity at day 21 compared with cells treated with the same dose of BTC and collected at day 3, indicating that BTC slowed, but did not arrest, the osteogenic differentiation of MSCs. In further support of an inhibitory influence of BTC upon osteogenic differentiation, BTC at 50 or 500 ng/ml significantly attenuated transcription of the osteogenic transcription factor RUNX2 and osteoblast marker COL1A1 relative to BTC-free cultures (Fig. 1b). Similar trends were observed for both genes in cultures collected after 14 and 21 days of BTC treatment, but this was without statistical significance; after day 28, no significant effect of BTC was deteted on RUNX2 or COL1A1 expression (data not shown). Interestingly, the expression of osteocalcin, a marker of mature osteoblasts, was not significantly affected by BTC treatment (not shown). We examined the possibility that a stimulatory influence of BTC upon osteogenesis was mediated by cells further committed to the osteoblastic phenotype by using MC3T3-E1 pre-osteoblasts. Similar to hMSC under osteogenic culture, Col1a2 expression was reduced by BTC, although this did not achieve statistical significance (Fig. 1c). Thus, these data indicated that BTC, similar to EGF and EGF-like ligands TGF-α and amphiregulin, inhibited the differentiation of osteoprogenitor and osteoblastic cells.
Fig. 1.
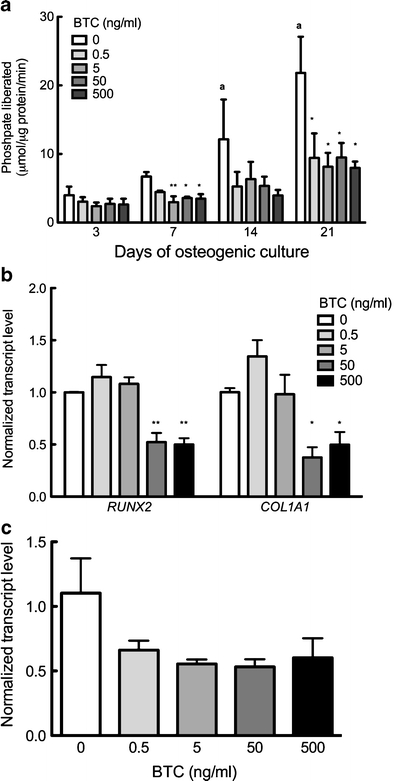
Effect of BTC on osteogenesis. hMSC or MC3T3-E1, under osteogenic conditions, were cultured in the presence of 0–500 ng/ml BTC. a Alkaline phosphatase activity assays of hMSCs after 3, 7, 14, or 21 days. b Quantitative polymerase chain reaction (qtPCR) analysis of RUNX2 and COL1A1 expression in hMSCs after 7 days of BTC treatment. c qtPCR analysis of Col1a2 expression in MC3T3-E1 cells after 7 days of BTC treatment. Bars Means ± SE. *P<0.05 compared with BTC-free culture within same timepoint; **P<0.01 compared with BTC-free culture within same timepoint; a P<0.05 compared with BTC-free culture on day 3; n=3–5 per experiment per time point
BTC increases MSC and pre-osteoblast proliferation
Based on the reported evidence that EGF and EGF-like ligands amphiregulin and TGF-α increase osteoblast proliferation (Loza et al. 1995; Nicolas et al. 1990; Qin et al. 2005; VandePol et al. 1989), we sought whether decreased osteogenic differentiation in response to BTC was paralleled with changes in proliferation. hMSCs and MC3T3-E1 pre-osteoblasts were cultured in the presence of 0.5–500 ng/ml BTC for 4 days. Addition of BTC at 5 ng/ml significantly increased hMSC cell proliferation approximately 2.2-fold over spontaneous proliferation observed in BTC-free cultures (Fig. 2a); BTC doses other than 5 ng/ml had no significant effect on cell proliferation. Similarly, MC3T3-E1 cultures with BTC demonstrated a significant increase in proliferation compared with BTC-free cultures (Fig. 2b), revealing a two-fold increase in cell number in response to 5 ng/ml BTC. These data confirmed a stimulatory influence of BTC upon cell proliferation and further revealed that this process was independent of mesenchymal or osteoblastic phenotype.
Fig. 2.
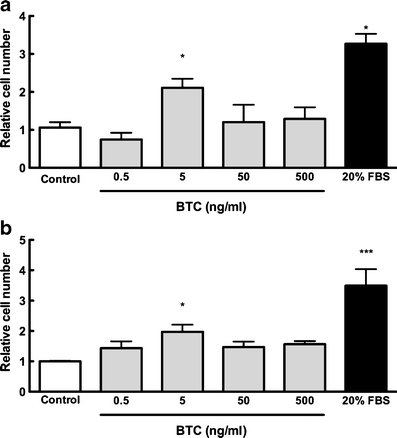
BTC stimulates cell proliferation. hMSCs (a) or MC3T3-E1 pre-osteoblasts (b) were cultured in the presence of 0.5–500 ng/ml BTC for 4 days, after which time cells were trypsinized and counted. Addition of BTC at 5 ng/ml significantly enhanced proliferation compared with control cultures (white bar). Cell proliferation in response to 20% serum is shown as a positive control. Because of variation between experiments, data are presented as the fold-change compared with BTC-free control. Bars Means ± SE. *P<0.05 compared with BTC-free culture; **P<0.001 compared with all other cultures; n=3–6 per experiment per time point
BTC induction of proliferation is mediated through HIF-α stabilization
EGF and EGF-like ligands stimulate a host of intracellular signaling cascades, including MAPK and Akt. An intriguing cascade involves activation of the transcription factor HIF-α (Peng et al. 2006; Schultz et al. 2006; Tanaka et al. 2006), the genetic stabilization of which mediates increased bone mass (Wan et al. 2008; Wang et al. 2007). Culture of hMSC with 5 ng/ml BTC for 24 h revealed a trend for HIF-1α accumulation compared with BTC-free controls, although to a lesser degree than that observed in cells cultured with the hypoxia mimetic, 100 μM cobalt chloride (Fig. 3a, b). This effect was transient and was not observed in cultures treated with BTC for 1, 2, 4, or 48 h (data not shown), indicating a transient influence of BTC upon HIF-1α stabilization. Because stable HIF-α is associated with proliferation (as observed in tumorigenic culture; Gardner and Corn 2008; Rankin and Giaccia 2008; Wouters and Koritzinsky 2008), we investigated whether BTC-mediated increases in osteoprogenitor proliferation were mediated via HIF-1α stabilization. We confirmed that the HIF-α antagonist YC-1 (10 μM) prevented HIF-1α stabilization in response to both hypoxia (1% O2) or hypoxia mimetics (100 μM CoCl2; Fig. 3c). Co-culture of hMSC cells with the HIF-α antagonist YC-1 revealed a dose-dependent attenuation of 5 ng/ml BTC-mediated proliferation (Fig. 3d). The inhibitory influence of the same doses of YC-1 on HIF-1α levels was confirmed by Western immunoblotting (data not shown). These findings indicated that the stimulatory effect of BTC upon cell proliferation was mediated by stabilized HIF-α.
Fig. 3.
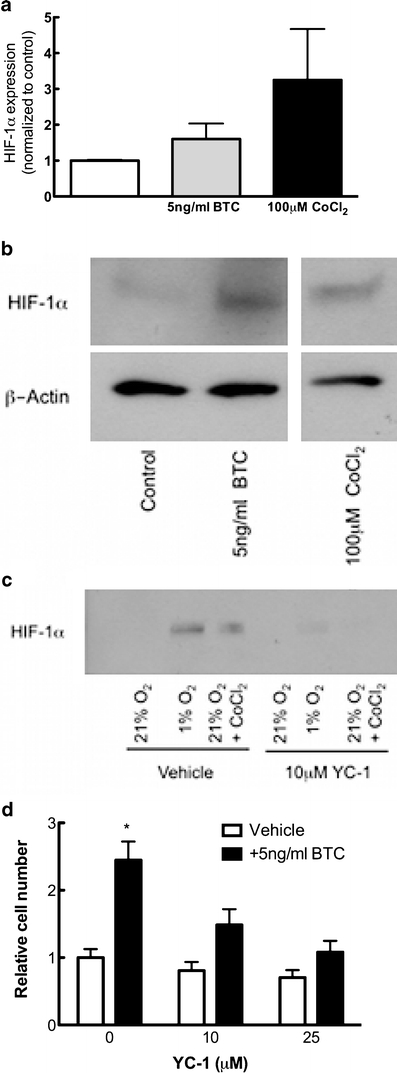
Influence of HIF-1α on BTC-driven proliferation. a hMSCs were cultured in the presence of 5 ng/ml BTC or 100 μM CoCl2 for 24 h, after which time whole-cell protein lysates were analyzed for HIF-1α, as normalized to β-actin. Treatment with BTC stabilized HIF-1α levels compared with BTC-free controls. b Western immunoblotting showed increased HIF-1α levels in BTC- and CoCl2-treated cells compared with control cultures. c Western immunoblotting demonstrated the induction of HIF-1α expression in response to hypoxia (1% O2) or hypoxia mimetics (100 μM CoCl2), whereas the addition of 10 μM YC-1 prevented HIF-1α stabilization under such conditions. d hMSCs were cultured in the presence of 0–25 μM YC-1 in the absence (white bars) or presence (black bars) of 5 ng/ml BTC. The addition of 10 or 25 μM YC-1 attenuated BTC-driven proliferation. Because of variation between experiments, data are presented as fold change compared with BTC-free control. Bars Means ± SE. *P<0.05 compared with BTC-free culture; n=3–4 experimental replicates
ErbB receptor expression in stem cells and pre-osteoblasts
The mRNA expression of ErbB receptor subtypes in hMSC and MC3T3-E1 was examined by quantitative PCR. We found that both naive (i.e., prior to further osteogenic differentiation) MSC and pre-osteoblasts express EGFR/ErbB1 (Fig. 4a), ErbB2 (Fig. 4b), and ErbB3 (Fig. 4c); no transcript for ErbB4 was observed (data not shown). All ErbB isoform receptors were expressed at higher levels in MSC than in MC3T3-E1, suggesting that osteogenic differentiation decreased ErbB expression, or that the levels changed as a function of immortalization. However, hMSCs cultured for 21 days under osteogenic or maintenance conditions revealed no significant differences in the transcript expression of ErbB1, ErbB2, or ErbB3 (not shown), suggesting that the relative differences in individual ErbB isoform expression was a species, rather than phenotypic, difference.
Fig. 4.
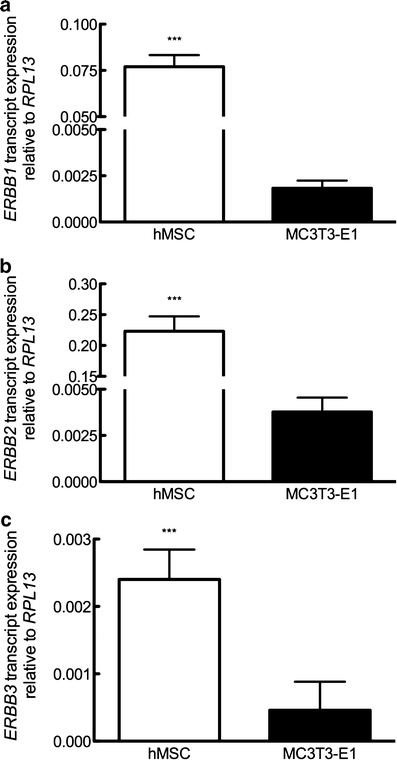
Profile of ERBB transcript expression. Expression of ERBB1 (a), ERBB2 (b), and ERBB3 (c) isoform transcript in hMSC and MC3T3-E1 cells. Bars Means ± SE. ***P<0.001 compared with MC3T3-E1; n=3–4 per experiment per condition
BTC does not influence osteoclast-regulating factors interleukin-6, osteoprotegerin, or RANKL
A potential explanation for the site-specific effects of BTC upon cortical and trabecular remodeling involves the regulation of osteoclast formation by osteoblasts. Thus, we examined the influence of BTC upon the expression of interleukin-6 (IL-6), osteoprotegerin (OPG), and receptor activator of NF-κB ligand (RANKL) in MSCs under osteogenic conditions. BTC had no effect upon IL-6 expression (Fig. 5a) and elicited a trend, albeit insignificant, for increased RANKL expression (Fig. 5b). In contrast, BTC at 50 or 500 ng/ml significantly decreased OPG transcript after 7 days in culture (Fig. 5c); the inhibitory influence of BTC upon the OPG transcript was transitory and was not observed after 14 or 21 days in culture. The significant reduction in OPG transcript and the trend for the increased RANKL expression promoted a trend for increased RANKL/OPG ratios, thereby favoring osteoclastogenesis, although the trend did not achieve statistical significance (Fig. 5d).
Fig. 5.
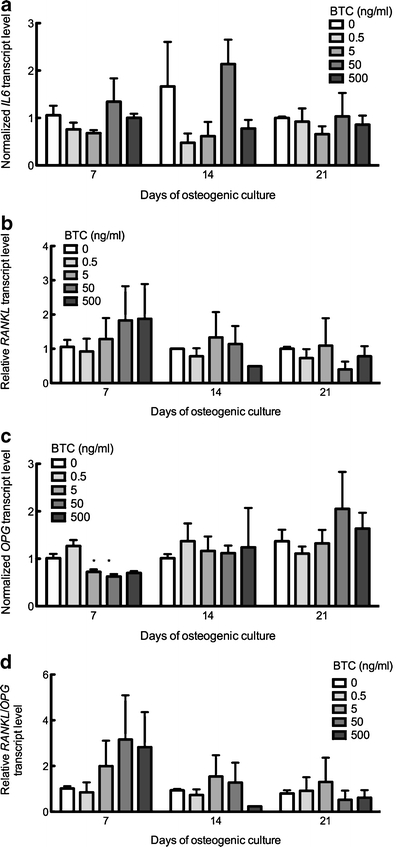
Influence of BTC upon osteoclast regulatory factors. hMSC were cultured under osteogenic conditions in the presence of 0.5–500 ng/ml BTC for 7–21 days, from which total RNA lysates were examined by qtPCR analysis for (a) IL-6, (b) RANKL, (c) OPG, or (d) RANKL/OPG. Bars Means ± SE. *P<0.05 compared with BTC-free culture within same timepoint. n=3–4 per experiment per time point
Discussion
The debate regarding the influence of EGF and EGF-like ligands upon osteoblast function and differentiation is ongoing. Whereas these growth factors have been shown to reduce osteogenic differentiation (Antosz et al. 1987; Nicolas et al. 1990; Qin et al. 2005), a recent report by Schneider et al. (2009) has demonstrated that BTC transgenic mice exhibit increased endocortical bone apposition and femoral bone mineral density but also present with decreased trabecular bone mineral density. In order to begin to delineate the mechanisms producing these site-specific effects of BTC, we have examined the influence of recombinant BTC upon the osteogenic differentiation of MSCs and committed pre-osteoblasts. We have found that BTC attenuates differentiation toward an osteogenic phenotype, as demonstrated by reductions in ALP activity and transcription of RUNX2 and COL1A1 (Fig. 1). Interestingly, we have observed no significant influence of BTC, regardless of dose, upon the expression of the mature osteoblast marker osteocalcin. Thus, BTC functions similar to EGF (Nicolas et al. 1990) and amphiregulin (Qin et al. 2005) to reduce osteogenic differentiation. Interestingly, the ability of EGF, and presumably of BTC, to attenuate cellular differentiation is not specific to the osteogenic phenotype, as EGF also decreases the chondrogenic and adipogenic differentiation of MSCs and, in doing so, reversibly maintains their multilineage potential (Krampera et al. 2005).
We have observed a mitogenic influence of BTC upon MSCs (Fig. 2a) and on committed pre-osteoblasts (Fig. 2b); co-culture with a pharmacologic antagonist, YC-1, which promotes HIF-α degradation and inhibits its de novo synthesis (Chun et al. 2004), dose-dependently attenuates the proliferative response of MSCs to BTC (Fig. 3d). HIF-α is a transcription factor that, under conditions of sufficient molecular oxygen (i.e., under normoxia), is targeted for proteasomal degradation; in contrast, under conditions of reduced molecular oxygen (hypoxia), HIF-α is stabilized, complexes with HIF-β, and translocates to the nucleus to induce gene expression. We have recently demonstrated that HIF-1α is responsible for increases in EP1 prostanoid receptor under hypoxia (Genetos et al. 2009). Surprisingly, growth factors, similar to EGF and BTC, and hormones are able to stabilize HIF-α under non-hypoxic conditions. Fetal calf serum, thrombin, and angiotensin II all stabilize HIF-1α and promote its binding to DNA to induce gene transcription in vascular smooth muscle cells (Richard et al. 2000). Similarly, Schultz et al. (2006) have demonstrated that HIF-1α contributes to the proliferative response of smooth muscle cells to fibroblast growth factor-2 and platelet-derived growth factor. Hypoxia exerts pleiotropic effects upon MSCs and has been implicated in the commitment toward chondrogenic (Kanichai et al. 2008) or neuronal (Pacary et al. 2006, 2007, 2008) lineages; in contrast, we have recently shown that hypoxia attenuates the osteogenic differentiation of MSCs (Raheja et al. 2009). Taken together, these data indicate that growth factor stabilization of HIF-α under nonhypoxic conditions promotes cell proliferation.
EGF and EGF-like ligands also influence skeletal remodeling by controlling bone resorption. Rats administered high doses of EGF reveal increased osteoclast number on trabecular bone surfaces (Marie et al. 1990), and EGF transgenic mice produce thinner bones than wild-type controls (Chan and Wong 2000; Xian 2007). Interestingly, considerable controversy exists as to whether osteoclasts express receptors for EGF or EGF-like ligands. Martineau-Doize et al. (1988) have demonstrated that osteoclasts fail to bind [125I]-iodo-EGF, suggesting that osteoclasts lack EGF receptors; these findings have subsequently been confirmed by Zhu et al. (2007). In contrast, Yi et al. (2008) report that EGF-receptor increases during osteoclastogenesis in response to RANKL, and that EGF-receptor signaling is required for RANKL-mediated osteoclastogenesis. The most likely cell type influenced by EGF to affect osteoclasts or their precursors is the osteoblast, primarily because of the close microenvironment in which these cells act as neighbors and modulate osteoclast formation and activity. Osteoblasts regulate osteoclast formation via RANKL, OPG, and IL-6, among other cytokines. Zhu et al. (2007) have demonstrated that EGF indirectly stimulates osteoclast formation via the decreased OPG expression by osteoblasts; similarly, we have observed a transient inhibitory influence of BTC upon OPG expression, but no influence upon RANKL transcription. Consequently, the ratio of the RANKL/OPG transcript demonstrates a trend above unity, indicating a stimulatory effect upon osteoclastogenesis.
EGF ligand binding induces manifold cellular responses depending upon the ErbB receptors mobilized. We have found that both MSCs and committed pre-osteoblasts express three of the four EGF receptors, lacking any expression of ErbB4. Among the three ErbB isoforms found within these cells, ErbB2 is maximally expressed, followed by ErbB1; ErbB3 is also expressed, albeit at approximately 25-fold lower levels than ErbB2 (Fig. 4a, c). Osteogenic differentiation has revealed no significant change in ErbB isoform expression (Fig. 4a–c). Based upon a Scatchard analysis revealing the KD of ErbB1 and BTC as 0.5 nM (4.5 ng/ml) in 3T3 cells (Watanabe et al. 1994), our chosen doses of 0.5, 5, 50, and 500 ng/ml correspond to, approximately, 10%, 50%, 90%, and 99% ErbB1 receptor occupancy, respectively. The lack of expression of ErbB4, a binding partner for BTC, in MSC or pre-osteoblasts indicates that the stimulatory influence of BTC upon proliferation and its inhibitory effect upon differentiation is mediated by ErbB1 homodimers or ErbB1/ErbB2 heterodimer or, potentially, an ErbB2/ErbB3 heterodimer (Pinkas-Kramarski et al. 1998). Indeed, each of these processes (increased proliferation and attenuated differentiation) are probably regulated by a different ErbB receptor complex, because of the variable affinity of ErbB1 and ErbB2 for ligands.
Our findings do not however reveal the mechanisms that explain site-specific differences in cortical formation and trabecular resorption in BTC transgenic mice. The most likely scenario results from the different microenvironments in which osteoblasts reside in cortical versus trabecular bone: osteoblasts within a remodeling compartment in cancellous bone are intimately exposed to a cytokine-rich red-marrow milieu, whereas osteoblasts within cortical bone are controlled more by mechanical or systemic factors such as parathyroid hormone (PTH) and 1α,25-(OH)2 vitamin D3 (Mundy 1999). Indeed, vitamin D3 has been shown to enhance osteoblast maturation in response to EGF (Yarram et al. 2004), and PTH induces EGF and EGF-like ligand expression (Qin and Partridge 2005; Qin et al. 2005; Swarthout et al. 2001; Xian 2007). In contrast, localized factors present within the bone marrow stroma, such as insulin-like growth factor (IGF), decrease osteoblast differentiation and instead promote proliferation (VandePol et al. 1989). IGF is a powerful regulator of post-natal bone growth, and EGF regulates IGF and IGF-binding protein (IGFBP) levels (Xian 2007). In further support of this possibility, reductions in body weight that occur in EGF transgenic mice are associated with serum IGFBP-3 levels (Chan and Wong 2000). Thus, an unexamined scenario is whether the stunted growth observed in EGF and BTC transgenics is indirectly mediated through altered IGF bioavailability.
This study has revealed that BTC stimulates the proliferation and inhibits the osteogenic differentiation of both MSCs and committed pre-osteoblasts. This stimulatory influence of BTC upon mesenchymal proliferation is mediated via the stabilization of HIF-1α. We have found that MSCs and committed osteoprogenitors express ErbB1, ErbB2, and ErbB3 but fail to express ErbB4; we have observed no change in the expression of these receptors under osteogenic conditions. Whereas previous work has demonstrated the influence of EGF, TGF-α, and amphiregulin upon osteoblast proliferation, our data are the first to reveal a similar role for BTC and are the first to provide a mechanistic explanation involving HIF-1α.
Acknowledgements
The authors are grateful to Chrisoula A. Toupadakis for critical review of the manuscript, to Prof. David P. Fyhrie for helpful comments on cortical and cancellous bone growth, and to Prof. Jeffrey M. Gimble for general discussion.
Open Access
This article is distributed under the terms of the Creative Commons Attribution Noncommercial License which permits any noncommercial use, distribution, and reproduction in any medium, provided the original author(s) and source are credited.
Footnotes
The authors are grateful to the Center for Equine Health (UC Davis) for funding to support this study.
Contributor Information
Damian C. Genetos, Phone: +1-530-7546865, FAX: +1-530-7540150, Email: dgenetos@ucdavis.edu
Rameshwar R. Rao, Email: ram.rao@gmail.com
Martin A. Vidal, Email: mavidal@ucdavis.edu
References
- Antosz ME, Bellows CG, Aubin JE. Biphasic effects of epidermal growth factor on bone nodule formation by isolated rat calvaria cells in vitro. J Bone Miner Res. 1987;2:385–393. doi: 10.1002/jbmr.5650020505. [DOI] [PubMed] [Google Scholar]
- Chan SY, Wong RW. Expression of epidermal growth factor in transgenic mice causes growth retardation. J Biol Chem. 2000;275:38693–38698. doi: 10.1074/jbc.M004189200. [DOI] [PubMed] [Google Scholar]
- Chien HH, Lin WL, Cho MI. Down-regulation of osteoblastic cell differentiation by epidermal growth factor receptor. Calcif Tissue Int. 2000;67:141–150. doi: 10.1007/s00223001128. [DOI] [PubMed] [Google Scholar]
- Chun YS, Yeo EJ, Park JW. Versatile pharmacological actions of YC-1: anti-platelet to anticancer. Cancer Lett. 2004;207:1–7. doi: 10.1016/j.canlet.2004.01.005. [DOI] [PubMed] [Google Scholar]
- Citri A, Yarden Y. EGF-ERBB signalling: towards the systems level. Nat Rev Mol Cell Biol. 2006;7:505–516. doi: 10.1038/nrm1962. [DOI] [PubMed] [Google Scholar]
- Gardner LB, Corn PG. Hypoxic regulation of mRNA expression. Cell Cycle. 2008;7:1916–1924. doi: 10.4161/cc.7.13.6203. [DOI] [PubMed] [Google Scholar]
- Genetos DC, Lee CM, Wong A, Yellowley CE. HIF-1alpha regulates hypoxia-induced EP1 expression in osteoblastic cells. J Cell Biochem. 2009;107:233–239. doi: 10.1002/jcb.22119. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Harris RC, Chung E, Coffey RJ. EGF receptor ligands. Exp Cell Res. 2003;284:2–13. doi: 10.1016/S0014-4827(02)00105-2. [DOI] [PubMed] [Google Scholar]
- Kanichai M, Ferguson D, Prendergast PJ, Campbell VA. Hypoxia promotes chondrogenesis in rat mesenchymal stem cells: a role for AKT and hypoxia-inducible factor (HIF)-1alpha. J Cell Physiol. 2008;216:708–715. doi: 10.1002/jcp.21446. [DOI] [PubMed] [Google Scholar]
- Krampera M, Pasini A, Rigo A, Scupoli MT, Tecchio C, Malpeli G, Scarpa A, Dazzi F, Pizzolo G, Vinante F. HB-EGF/HER-1 signaling in bone marrow mesenchymal stem cells: inducing cell expansion and reversibly preventing multilineage differentiation. Blood. 2005;106:59–66. doi: 10.1182/blood-2004-09-3645. [DOI] [PubMed] [Google Scholar]
- Livak KJ, Schmittgen TD. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) method. Methods. 2001;25:402–408. doi: 10.1006/meth.2001.1262. [DOI] [PubMed] [Google Scholar]
- Loza J, Carpio L, Lawless G, Marzec N, Dziak R. Role of extracellular calcium influx in EGF-induced osteoblastic cell proliferation. Bone. 1995;16:341S–347S. doi: 10.1016/8756-3282(94)00049-6. [DOI] [PubMed] [Google Scholar]
- Marie PJ, Hott M, Perheentupa J. Effects of epidermal growth factor on bone formation and resorption in vivo. Am J Physiol. 1990;258:E275–E281. doi: 10.1152/ajpendo.1990.258.2.E275. [DOI] [PubMed] [Google Scholar]
- Martineau-Doize B, Lai WH, Warshawsky H, Bergeron JJ. In vivo demonstration of cell types in bone that harbor epidermal growth factor receptors. Endocrinology. 1988;123:841–858. doi: 10.1210/endo-123-2-841. [DOI] [PubMed] [Google Scholar]
- Mundy GR. Bone remodeling. In: Favus MJ, editor. Primer on the metabolic bone diseases and disorders of mineral metabolism. Philadelphia: Lippincott Williams & Wilkins; 1999. [Google Scholar]
- Nicolas V, Nefussi JR, Collin P, Forest N. Effects of acidic fibroblast growth factor and epidermal growth factor on subconfluent fetal rat calvaria cell cultures: DNA synthesis and alkaline phosphatase activity. Bone Miner. 1990;8:145–156. doi: 10.1016/0169-6009(90)90117-X. [DOI] [PubMed] [Google Scholar]
- Pacary E, Legros H, Valable S, Duchatelle P, Lecocq M, Petit E, Nicole O, Bernaudin M. Synergistic effects of CoCl2 and ROCK inhibition on mesenchymal stem cell differentiation into neuron-like cells. J Cell Sci. 2006;119:2667–2678. doi: 10.1242/jcs.03004. [DOI] [PubMed] [Google Scholar]
- Pacary E, Tixier E, Coulet F, Roussel S, Petit E, Bernaudin M. Crosstalk between HIF-1 and ROCK pathways in neuronal differentiation of mesenchymal stem cells, neurospheres and in PC12 neurite outgrowth. Mol Cell Neurosci. 2007;35:409–423. doi: 10.1016/j.mcn.2007.04.002. [DOI] [PubMed] [Google Scholar]
- Pacary E, Petit E, Bernaudin M. Concomitant inhibition of prolyl hydroxylases and ROCK initiates differentiation of mesenchymal stem cells and PC12 towards the neuronal lineage. Biochem Biophys Res Commun. 2008;377:400–406. doi: 10.1016/j.bbrc.2008.09.145. [DOI] [PubMed] [Google Scholar]
- Peng XH, Karna P, Cao Z, Jiang BH, Zhou M, Yang L. Cross-talk between epidermal growth factor receptor and hypoxia-inducible factor-1alpha signal pathways increases resistance to apoptosis by up-regulating survivin gene expression. J Biol Chem. 2006;281:25903–25914. doi: 10.1074/jbc.M603414200. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pinkas-Kramarski R, Lenferink AE, Bacus SS, Lyass L, van de Poll ML, Klapper LN, Tzahar E, Sela M, van Zoelen EJ, Yarden Y. The oncogenic ErbB-2/ErbB-3 heterodimer is a surrogate receptor of the epidermal growth factor and betacellulin. Oncogene. 1998;16:1249–1258. doi: 10.1038/sj.onc.1201642. [DOI] [PubMed] [Google Scholar]
- Qin L, Partridge NC. Stimulation of amphiregulin expression in osteoblastic cells by parathyroid hormone requires the protein kinase A and cAMP response element-binding protein signaling pathway. J Cell Biochem. 2005;96:632–640. doi: 10.1002/jcb.20550. [DOI] [PubMed] [Google Scholar]
- Qin L, Tamasi J, Raggatt L, Li X, Feyen JH, Lee DC, Dicicco-Bloom E, Partridge NC. Amphiregulin is a novel growth factor involved in normal bone development and in the cellular response to parathyroid hormone stimulation. J Biol Chem. 2005;280:3974–3981. doi: 10.1074/jbc.M409807200. [DOI] [PubMed] [Google Scholar]
- Raheja LF, Genetos DC, Yellowley CE (2009) The effect of oxygen tension on the long-term osteogenic differentiation and MMP/TIMP expression of human mesenchymal stem cells. Cells Tissues Organs (in press) [DOI] [PubMed]
- Rankin EB, Giaccia AJ. The role of hypoxia-inducible factors in tumorigenesis. Cell Death Differ. 2008;15:678–685. doi: 10.1038/cdd.2008.21. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Richard DE, Berra E, Pouyssegur J. Nonhypoxic pathway mediates the induction of hypoxia-inducible factor 1alpha in vascular smooth muscle cells. J Biol Chem. 2000;275:26765–26771. doi: 10.1074/jbc.M003325200. [DOI] [PubMed] [Google Scholar]
- Schneider MR, Dahlhoff M, Herbach N, Renner-Mueller I, Dalke C, Puk O, Graw J, Wanke R, Wolf E. Betacellulin overexpression in transgenic mice causes disproportionate growth, pulmonary hemorrhage syndrome, and complex eye pathology. Endocrinology. 2005;146:5237–5246. doi: 10.1210/en.2005-0418. [DOI] [PubMed] [Google Scholar]
- Schneider MR, Mayer-Roenne B, Dahlhoff M, Proell V, Weber K, Wolf E, Erben RG. High cortical bone mass phenotype in betacellulin transgenic mice is EGFR dependent. J Bone Miner Res. 2009;24:455–466. doi: 10.1359/jbmr.081202. [DOI] [PubMed] [Google Scholar]
- Schultz K, Fanburg BL, Beasley D. Hypoxia and hypoxia-inducible factor-1alpha promote growth factor-induced proliferation of human vascular smooth muscle cells. Am J Physiol Heart Circ Physiol. 2006;290:H2528–H2534. doi: 10.1152/ajpheart.01077.2005. [DOI] [PubMed] [Google Scholar]
- Swarthout JT, Doggett TA, Lemker JL, Partridge NC. Stimulation of extracellular signal-regulated kinases and proliferation in rat osteoblastic cells by parathyroid hormone is protein kinase C-dependent. J Biol Chem. 2001;276:7586–7592. doi: 10.1074/jbc.M007400200. [DOI] [PubMed] [Google Scholar]
- Tanaka H, Yamamoto M, Hashimoto N, Miyakoshi M, Tamakawa S, Yoshie M, Tokusashi Y, Yokoyama K, Yaginuma Y, Ogawa K. Hypoxia-independent overexpression of hypoxia-inducible factor 1alpha as an early change in mouse hepatocarcinogenesis. Cancer Res. 2006;66:11263–11270. doi: 10.1158/0008-5472.CAN-06-1699. [DOI] [PubMed] [Google Scholar]
- VandePol C, Schlaeger D, Wong GL. Mitogenic responses to and binding of insulin-like growth factor 1 and/or epidermal growth factor by bone cells. Bone Miner. 1989;5:371–382. doi: 10.1016/0169-6009(89)90014-7. [DOI] [PubMed] [Google Scholar]
- Wan C, Gilbert SR, Wang Y, Cao X, Shen X, Ramaswamy G, Jacobsen KA, Alaql ZS, Eberhardt AW, Gerstenfeld LC, Einhorn TA, Deng L, Clemens TL. Activation of the hypoxia-inducible factor-1alpha pathway accelerates bone regeneration. Proc Natl Acad Sci USA. 2008;105:686–691. doi: 10.1073/pnas.0708474105. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wang Y, Wan C, Deng L, Liu X, Cao X, Gilbert SR, Bouxsein ML, Faugere MC, Guldberg RE, Gerstenfeld LC, Haase VH, Johnson RS, Schipani E, Clemens TL. The hypoxia-inducible factor alpha pathway couples angiogenesis to osteogenesis during skeletal development. J Clin Invest. 2007;117:1616–1626. doi: 10.1172/JCI31581. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Watanabe T, Shintani A, Nakata M, Shing Y, Folkman J, Igarashi K, Sasada R. Recombinant human betacellulin. Molecular structure, biological activities, and receptor interaction. J Biol Chem. 1994;269:9966–9973. [PubMed] [Google Scholar]
- Wong RW, Kwan RW, Mak PH, Mak KK, Sham MH, Chan SY. Overexpression of epidermal growth factor induced hypospermatogenesis in transgenic mice. J Biol Chem. 2000;275:18297–18301. doi: 10.1074/jbc.M001965200. [DOI] [PubMed] [Google Scholar]
- Wouters BG, Koritzinsky M. Hypoxia signalling through mTOR and the unfolded protein response in cancer. Nat Rev Cancer. 2008;8:851–864. doi: 10.1038/nrc2501. [DOI] [PubMed] [Google Scholar]
- Xian CJ. Roles of epidermal growth factor family in the regulation of postnatal somatic growth. Endocr Rev. 2007;28:284–296. doi: 10.1210/er.2006-0049. [DOI] [PubMed] [Google Scholar]
- Yarden Y. The EGFR family and its ligands in human cancer. Signalling mechanisms and therapeutic opportunities. Eur J Cancer. 2001;37(Suppl 4):S3–S8. doi: 10.1016/S0959-8049(01)00230-1. [DOI] [PubMed] [Google Scholar]
- Yarden Y, Sliwkowski MX. Untangling the ErbB signalling network. Nat Rev Mol Cell Biol. 2001;2:127–137. doi: 10.1038/35052073. [DOI] [PubMed] [Google Scholar]
- Yarram SJ, Tasman C, Gidley J, Clare M, Sandy JR, Mansell JP. Epidermal growth factor and calcitriol synergistically induce osteoblast maturation. Mol Cell Endocrinol. 2004;220:9–20. doi: 10.1016/j.mce.2004.04.005. [DOI] [PubMed] [Google Scholar]
- Yi T, Lee HL, Cha JH, Ko SI, Kim HJ, Shin HI, Woo KM, Ryoo HM, Kim GS, Baek JH. Epidermal growth factor receptor regulates osteoclast differentiation and survival through cross-talking with RANK signaling. J Cell Physiol. 2008;217:409–422. doi: 10.1002/jcp.21511. [DOI] [PubMed] [Google Scholar]
- Zhu J, Jia X, Xiao G, Kang Y, Partridge NC, Qin L. EGF-like ligands stimulate osteoclastogenesis by regulating expression of osteoclast regulatory factors by osteoblasts: implications for osteolytic bone metastases. J Biol Chem. 2007;282:26656–26664. doi: 10.1074/jbc.M705064200. [DOI] [PubMed] [Google Scholar]