

Abstract
Even though peripheral nerves regenerate well, axons are often misrouted and reinnervate inappropriate distal pathways post-injury. Misrouting most likely occurs at branch points where regenerating axons make choices. Here, we show that the accuracy of sensory axon reinnervation is enhanced by overexpression of the guidance molecule nerve growth factor (NGF) distal to the bifurcation. We used the femoral nerve as model, which contains both sensory and motor axons that intermingle in the parent trunk and distally segregate into the saphenous (SB) and motor branch (MB). Transection of the parent trunk resulted in misrouting of axon reinnervation to SB and MB. To enhance sensory axon targeting, recombinant adenovirus encoding NGF was injected along the SB close to the bifurcation one week post-injury. The accuracy of axon reinnervation was assessed by retrograde tracing at 3 or 8 weeks after nerve injury. NGF overexpression significantly increased the accuracy of SB axon reinnervation to the appropriate nerve branch, in a manner independent of enhancing axon regeneration. This novel finding provides in vivo evidence that gradient expression of neurotrophin can be used to enhance targeting of distal peripheral pathways to increase axon regeneration into the appropriate nerve branch.
Keywords: femoral transection, axon reinnervation, recombinant adenovirus, nerve growth factor, axon guidance, axon targeting, triple retrograde tracing
Introduction
Incomplete functional recovery has long been observed after surgical repair of transected peripheral nerve (Sunderland, 1978, Vordemvenne, et al., 2007). The therapeutic failure often results from misdirection of regenerating axons to target branches which lead to functionally inappropriate end organs (Brushart, et al., 2005). Many studies have investigated the underlying mechanism of post-traumatic axon pathway selection (Ahlborn, et al., 2007, Brushart, 1988, Madison, et al., 1996) to help develop a clinically relevant therapy aiming to increase the specificity of axon regeneration and enhance functional restoration.
The specificity of sensory and motor targets for axon reinnervation was observed by Cajal, who attributed it to “a neurotrophic influence which has an individual and specific character” of end organs (Cajal, 1928). Recent studies demonstrate motor axons preferentially reinnervate muscle pathway as opposed to cutaneous pathway after transection of the parent femoral nerve, even in the absence of end organ contact (Brushart, 1993, Madison, et al., 1996). Nerve grafts from denervated cutaneous nerve and ventral root preferentially supported sensory or motor axon regeneration, respectively (Hoke, et al., 2006). These results suggested some specificity of sensory and motor pathways for axon reinnervation. Schwann cells in sensory and motor nerves display differences in growth factor expression, indicating the pathway specificity is due to the specific growth factor expression. Nerve growth factor (NGF) is one of the growth factors that predominately expressed in denervated and grafted cutaneous nerve (Hoke, et al., 2006). This matches its previously reported roles in growth and guidance of sensory axons (Heron, et al., 2007, Paves and Saarma, 1997, Zhou, et al., 2006). Furthermore, overexpression of NGF within the denervated spinal cord enhanced regeneration of the calcitonin gene-related peptide (CGRP)-positive nociceptive axons across the dorsal root entry zone (Romero, et al., 2001), demonstrating a targeting role for lesioned axons.
Femoral transection is a commonly used injury model to assess the specificity of post-traumatic axon reinnervation. The femoral nerve originates from the lumbar plexus II, III, and IV, contains both sensory and motor axons that intermingle in the parent trunk and segregate into separate distal branches: one purely sensory passes subcutaneously along the tibial side of the leg and innervate skin (saphenous branch, SB), while the other leading to quadriceps contains both motor and proprioceptive axons (motor branch, MB). Transection of the parent femoral nerve results in inappropriate axon reinnervation to SB and MB (Brushart, 1988) (Fig. 1A – B). We hypothesized that overexpression of NGF along post-traumatic SB would mediate the accuracy of sensory axon reinnervation.
Figure 1.
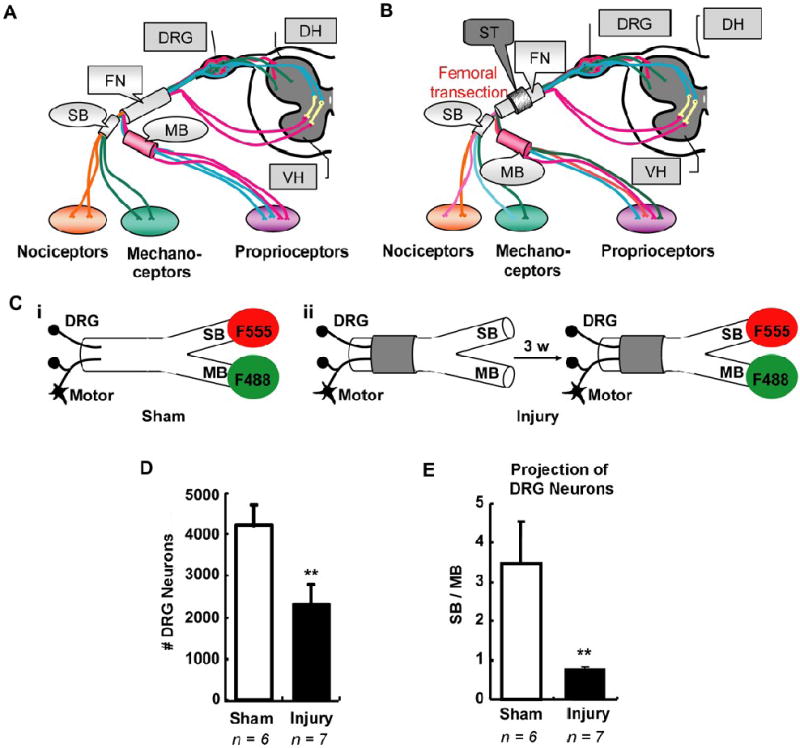
Femoral transection induced abnormal sensory neuron reinnervation. A, Schematic illustration of the adult femoral nerve, which consists of parent trunk, saphenous branch (SB) and motor branch (MB). B, Schematic illustration of axon reinnervation in transected femoral nerve. Transection of parent trunk with a lesion gap in a guidance tube usually results in inappropriate axons reinnervating either SB or MB. C, Scheme of simultaneous double retrograde tracing. F555 (red fluorescent) and F488 (green fluorescent) were independently applied at either SB or MB in sham group (i) or in injury group three weeks after femoral transection (ii). D – E, Counts of labeled DRG neurons correspond to sensory neurons innervating either SB or MB. Statistical results showed the total number of retrograde labeled neurons significantly decreased after femoral transection (D). Meanwhile, their projection towards SB or MB significant changed compared to sham group, demonstrating abnormal sensory neuron reinnervation (E). DRG, dorsal root ganglion; DH, dorsal horn; VH, ventral horn; FN, femoral nerve; SB, saphenous branch; MB, motor branch; ST, silicone tubing; F555, Dextran AlexaFluor 555; F488, Dextran AlexaFluor 488; 3 w, 3 weeks. Values represent mean ± S.E.M; n = 6 – 7. ** p < 0.01 compared to sham group, analyzed by Student’s t-test.
To investigate this hypothesis, we administrated recombinant adenovirus encoding NGF (Ad-NGF) along SB in transected femoral nerve. Sequential triple retrograde tracing was performed to assess the accuracy of sensory axon reinnervation.
Material and Methods
Experimental animals
Fifty-five female adult (250 – 300 g) Sprague Dawley rats (SD rats, from Harlan Sparague Dawley, Indianapolis, IN) were used in this study. They were organized into 9 experimental groups (Table 1). All surgical procedures and animal maintenance complied with the National Institutes of Health guidelines regarding the care and use of experimental animals and were approved by the Institutional Animal Care and Research Advisory Committee.
Table 1.
Animal groups and experimental design
| Experimental animal groups | Pre-labeling on SB (1st RL) | Femoral lesion | Administration of recombinant adenovirus | Double labeling on SB and MB (2nd RL) | |
|---|---|---|---|---|---|
| Double RL | Sham | - | - | - | + |
| Non-Ad | - | transection | - | 3 weeks post-injury | |
| Ad-NGF | - | transection | 1 week post-injury | 3 weeks post-injury | |
| Sequential triple RL | Sham | + | - | - | 4 or 9 weeks after 1st RL |
| Non-Ad | 1 week pre-injury | transection | - | 3 or 8 weeks post-injury | |
| Ad-GFP | 1 week pre-injury | transection | 1 week post-injury | 3 weeks post-injury | |
| Ad-NGF | 1 week pre-injury | transection | 1 week post-injury | 3 or 8 weeks post-injury | |
| NGF Elisa or fluorescent imaging | Ad-GFP | - | crush | immediately post-injury | - |
| Ad-NGF | - | crush | Immediately post-injury | - | |
RL, retrograde labeling; Ad, adenovirus; Non-Ad, non-adenovirus; Ad-NGF, recombinant adenovirus encoding NGF; Ad-GFP, recombinant adenovirus encoding GFP
Adenoviral vectors
Replication-defective recombinant adenoviruses (Ad) expressing NGF (Ad-NGF) and green fluorescent protein (Ad-GFP, control virus) were constructed as described previously (Romero, et al., 2000). In addition to standard deletions in E1 and E3 coding regions, all viruses also encoded the ts125 mutation in the E2a, DNA binding protein, to further reduce potential toxicity after administration in vivo (Romero and Smith, 1998). All plaque-purified adenoviruses were examined for replication-competent adenoviruses via PCR. Viruses were then amplified and purified by double cesium chloride gradient ultracentrifugation. The physical number of viral particles was determined by optical absorbency. The number of infectious particles was estimated using Adeno-X Rapid titer kit (from Clontech, Palo Alto, CA) or directly counting green fluorescent cells 48 hr after transfection of 293 cells. The viral particle (vp) to plaque-forming unit (pfu) ratio varied from 10:1 to 100:1.
Femoral transection and adenoviral administration
Deeply anesthetized animals [ketamine (67 mg/kg, i.p.) / xylazine (6.7 mg/kg, i.p.)] underwent transected injury on the left femoral nerves. Briefly, the femoral nerve was transected 1 mm distal to iliacus nerve, i.e. 6 – 8 mm proximal to the bifurcated point of SB and MB. The proximal and distal stumps were sutured into a 2-mm-long silicone tubing (1 mm inner diameter, from A-M systems, Carlsborg, WA) with one 10-0 nylon (from Ethicon, Somerville, NJ) stitch. A lesion gap about 0.5 – 0.8 mm long was maintained between the two stumps.
One week later, some injured animals were treated with Ad-NGF or Ad-GFP. Immediately before virus treatment, animals intraperitoneally received 100 μg of equally combined solution of CD-4 (W3/25) and CD-45 (MRC OX-22) antisera for transient immune suppression, as described previously (Romero and Smith, 1998). Each adenoviral administration consists of 18 injections at three sites (1 mm apart from 2 – 3 mm distal to bifurcated point) along SB. All injections were made with a beveled glass micropipette (30 – 50 μm external diameter) using a nanoliter injection device (from World Precision Instruments, Sarasota, FL). At each injection, the micropipette was adjusted at an appropriate angle to insert into the nerve vertically at a depth of 0.2 mm with a micromanipulator, and nanoliter (50.5 nl) quantity of saline solutions containing Ad-GFP (2.27 – 3.62 × 105 pfu/μl) or Ad-NGF (3.62 × 105 pfu/μl) were slowly infused into the SB at the speed of 1 nl / sec. There was 1 min pause between each injection. Our previous studies demonstrated this procedure resulted in robust transgene expression consistently throughout the injected area with little cell death or tissue damage (Romero and Smith, 1998).
Simultaneous double retrograde tracing
Both SB and MB were transected 2 mm distal to the bifurcated point, and the proximal cut end was exposed to 2 μl of 10% Dextran AlexaFluor 555 (F555, 10, 000 mw, from Invitrogen, Eugene, OR) or 10% Dextran AlexaFluor 488 (F488, 10, 000 mw, from Invitrogen, Eugene, OR). F555 and F488 were randomly chosen to apply at SB and MB, respectively, and showed similar efficacy for retrograde labeling. For convenience of description, all SB ends were mentioned to be exposed to F555 whereas all MB ends to F488 during their simultaneous usage in retrograde tracing. Forty-five minutes after tracer application, the cut ends were rinsed robustly with sterile saline and sutured with appropriate distal ends by one 10-0 nylon stitch. L2 – L4 DRGs were processed 48 hours or 7 days later.
Sequential triple retrograde tracing
Experimental animals underwent sequential triple retrograde tracing in combination with performances of femoral transection and adenoviral administration, as schemed in Fig. 2A. Briefly, SB was cut 4 mm distal to bifurcated point and the proximal end was exposed to 2 μl of 5% hydroxystilbamidine (also known as FluoroGold, FG; from Biotium, Hayward, CA) for 45 min. The cut ends were then rinsed robustly with sterile saline and sutured together by one 10-0 nylon stitch. One week later, femoral transection and adenoviral administration were performed as described above. Three or eight weeks after transection, 2nd retrograde tracing was performed using the same procedure for simultaneous double retrograde tracing. Seven days were allowed for tracer transportation, and then L2 – L4 DRGs were processed for further analysis. In sham control study, 1st and 2nd tracing were performed with an interval of 4 or 9 weeks, in the absence of femoral transection and adenoviral administration to establish baselines.
Figure 2.
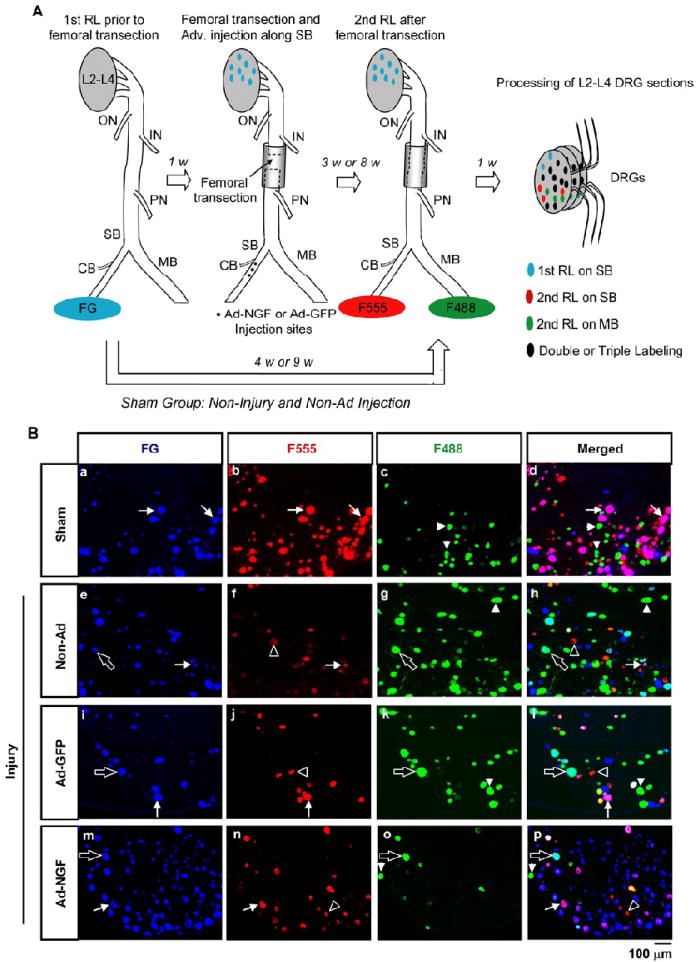
Accuracy of sensory neuron reinnervation was assessed by sequentially triple retrograde tracing. A, Schematic illustration of sequentially triple retrograde tracing. FG was applied on SB as 1st retrograde label (RL). One week later, femoral trunk was transected and the two severed stumps were sutured into silicone tubing with a 0.5 – 0.8 mm-long gap. Ad-NGF or Ad-GFP administration was performed one week post-injury. Three or eight weeks were allowed for nerve repair, and then F555 and F488 were applied at SB and MB, respectively, as 2nd RL. In sham group, femoral transection and adenoviral administration were absent between 1st RL and 2nd RL. Seven days after 2nd RL, L2 – L4 DRGs were processed for further analysis. B, Neuron labeling in different experimental groups three weeks after femoral transection. In sham group, most of FG-labeled neurons were double-labeled with F555, but rarely with F488 (a – d). In injured animals, both F555- and F488- labeling were found on FG-labeled neurons (e – p). Apparently, more co-labeling of F555 and FG was found in Ad-NGF group (m – p) compared to Ad-GFP group (i – l) and non-Ad group (e – h). Arrows, double-labeling of FG and F555; empty arrows, double-labeling of FG and F488; arrowheads, single labeling of F488; empty arrowheads, single labeling of F555. DRG, dorsal root ganglia; RL, retrograde labeling; ON, obturator nerve; IN, illiacus nerve; PN, pectineus nerve; SB, saphenous branch; MB, motor branch; CB, local cutaneous branch; F555, Dextran AlexaFluor 555; F488, Dextran AlexaFluor 488; non-Ad, non-adenovirus; Ad-NGF, recombinant adenovirus encoding NGF; Ad-GFP, recombinant adenovirus encoding GFP; Scale bar, 100 μm.
DRG processing, histology and image analysis
Animals were perfused transcardially with 0.9% NaCl, followed by 4% paraformaldehyde (PFA) in 0.1 M phosphate buffer (PB, pH 7.2). L2 – L4 DRGs were removed, post-fixed in 4% PFA for 1 hr at room temperature, and then move to 30% sucrose in 0.1 M PB at 4°C. Within one week, the DRGs were serially cryostated to 20 μm-thick sections on glass slides and reserved at -80 °C. For double immunostaining of FG and CGRP, air-dried DRG sections were simultaneously incubated with rabbit-anti-FG antibody (1:500, from Fluorochrome, Denver, CO) and mouse-anti-CGRP (1:4000, from Sigma Aldrich, St. Louis, MO) at 4°C overnight. Next day, the sections were incubated with AMCA-anti-rabbit IgG (1:50, from Jackson ImmunoResearch, West Grove, PA) and Cy5-anti-mouse IgG (1:200, from Jackson ImmunoResearch, West Grove, PA) at room temperature for 45 min. After staining, sections were coverslipped with Fluoromount-G (from SouthernBiotech, San Diego, CA) for fluorescence microscopy. The whole area of cryosections was photographed with Zeiss Conventional Inverted Microscopy System Axiovert 200M under 10x objective on a Zeiss LSM510 microscope. All labeled neurons were counted manually from the images.
Femoral crush and adenoviral administration
To evaluate the in vivo transgenic expression of NGF and GFP in post-traumatic nerve, adenoviruses were injected into crushed femoral nerve. The left femoral nerve was tightly held with Dumont forceps no. 5 (from Fine Science Tools, Heidelberg, Germany) for 1 min by twice at the site 1 mm distal to iliacus nerve. Previous studies showed that all axons at the crush site would be broken under such a procedure (Irintchev, et al., 2005). Immediately after femoral crush, Ad-GFP or Ad-NGF (~ 5 × 106 pfu/μl) was injected along SB with the same projection procedure as described above. One week later, tissue segments from the treated femoral nerve were dissected for either GFP imaging or NGF Elisa.
Fluorescent imaging
Animals were perfused transcardially with 0.9% NaCl seven days after femoral crush and Ad-GFP injections. Tissue segments of SB and MB were dissected, 4-mm-long from the bifurcation, post-fixed in 4% PFA at 4°C overnight, and then moved to 30% sucrose at 4°C for 2 – 3 days. The nerves were cryostated to 10 μm-thick sections on glass slides, coverslipped with Fluoromount-G, and photographed under 10x objective with MetaMorph Image Analysis System on a Nikon VFM microscope.
NGF Elisa
One week after femoral crush and adenoviral adenovirus injections, 3mm long tissue segments were collected from SB, MB, and femoral trunk as illustrated in Fig. 3B-i. The tissues were homogenized and then sonicated with a Branson sonifier 450 (from VWR Scientific, West Chester, PA) in icy-cold extraction buffer, which consisted of 100 mM Tris/HCl (pH 7.0), 2% bovine serum albumin (BSA), 1 M NaCl, 4 mM EDTA.Na2, 2% Triton X-100, 0.1% sodium azide, protease inhibitor cocktail (1:100, from Sigma Aldrich, St. Louis, MO) and freshly made 17 μg/ml phenylmethyl-sulphonyl fluoride (PMSF). Protein assays were performed with BCA kit (from Pierce, Rockford, IL) and 200 μg / well (20 μg / μl) of each sample was used for Elisa test with NGF Emax ImmunoAssay System (from Promega, Madison, MI). After color development, 96-well plates were read at 450 nm using a BioTech E12a microplate reader.
Figure 3.
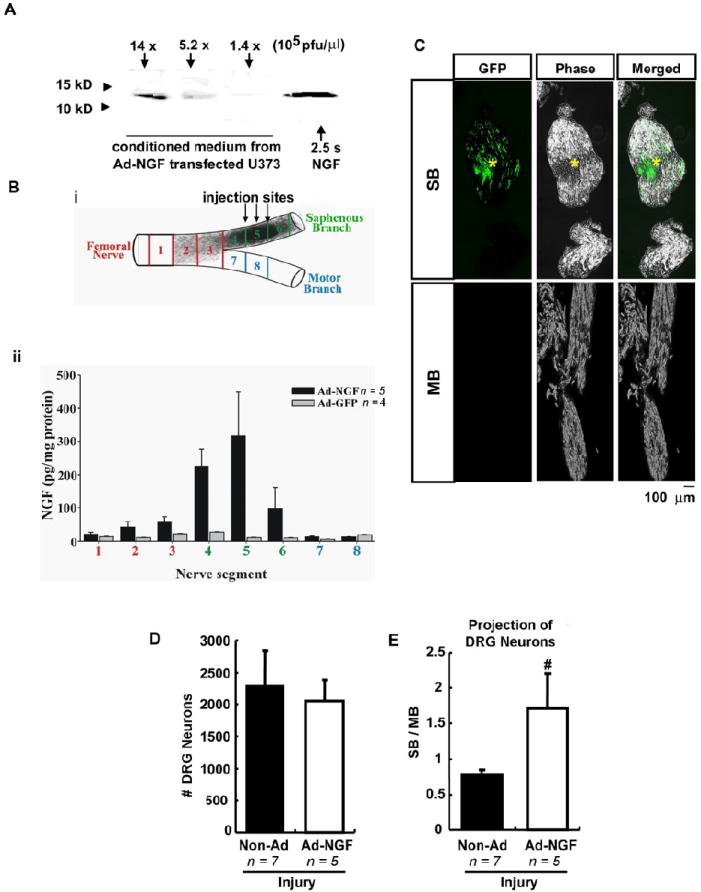
Ad-NGF administration significantly increased sensory neuron reinnervation to SB. A – C, Adenoviral-based transgenic overexpression of exogenous genes. Cultured U373 cells were infected with Ad-NGF of 1.4 × 105 pfu/μl, 5.2 × 105 pfu/μl, and 1.4 × 106 pfu/μl. Western blots of conditioned medium showed dose-dependent expression of NGF (A). For in vivo overexpression, Ad-NGF or Ad-GFP was injected along SB in the crushed femoral nerve. One week later, 3-mm long tissue segments were collected as illustrated and processed for NGF ELISA (B-i). ELISA results showed a dramatic increase of NGF expression along SB, but not MB, with the peak in Ad-NGF at the injection sites (B-ii). C, Fluorescent and phase imaging to show GFP expression along SB and MB one week after Ad-GFP administration on post-traumatic SB. D – E, One week after femoral transection, Ad-NGF was injected along SB, followed by double retrograde tracing at 3 weeks post-injury. Counts of labeled DRG neurons showed Ad-NGF treatment preferentially increased sensory neuron reinnervation to SB as opposed to MB (E) without changing the total number of regenerating sensory neurons (D). Ad-NGF, recombinant adenovirus encoding NGF; Ad-GFP, recombinant adenovirus encoding GFP; non-Ad, non-adenovirus; SB, saphenous branch; MB, motor branch. *, injection site; Scale bar, 100 μm. Values represent mean ± S.E.M; n = 5 – 7. # p < 0.05 compared to non-Ad group, analyzed by Student’s t-test.
Western blot of NGF
The transgenic in vitro expression of NGF was evaluated as previously described (Tang, et al., 2004). Briefly, 48 hr after Ad-NGF infection of U373 cells, 1 ml of culture supernatant was precipitated by 100 μl of 0.5% sodium deoxycholate and 100 μl of TCA. Proteins were pelleted by centrifugation at 14,000 rpm, washed in icy-cold 70% acetone, dried, and resuspended in 100 μl of lysis buffer, which consisted of 1% SDS in TE (10 mM Tris-Cl + 1 mM EDTA, pH 8.0) with protease inhibitor cocktail (1:10, from Sigma Aldrich, St. Louis, MO) and freshly made 17 μg/ml PMSF. After centrifuging at 14,000 rpm, protein was assayed for protein concentration with BCA kit (from Pierce, Rockford, IL), and 200 μg of each protein sample was loaded for electrophoresis, transferred to nitrocellulose membrane, and identified with rabbit anti-NGF antibody (1:500, from Accurate, Westbury, NY).
Calculation of reinnervation accuracy
The reinnervation accuracy is calculated by counting multi-labeled DRG neurons, based on sequential triple retrograde tracing. The number of FG-labeled neurons designates the number of neurons normally projecting to the SB (SB neurons), whereas those normally projecting to the other branches (non-SB neurons) lack FG-labeling. The number of F555- or F488-labeled cells represents the number of regenerating neurons projecting to either SB or MB. Dual-labeling of F555 and F488 represents axon branching into both nerves from regenerating neurons. Neurons labeled by FG but not F555 or F488 represent neuronal die back or failed regeneration of SB axons after transection. For regeneration of either SB neurons or non-SB neurons, the percentages of F555-or F488-labeling represent the percentages of axons selectively regenerating into a distinct pathway.
Statistical analysis
Mean values (± standard error of the mean; SEM) are shown throughout. Comparisons were made using One-way Analysis of Variance (ANOVA) followed by Tukey’s Post Hoc test among multiple groups or using unpaired student’s t-tests between only two groups. p-value < 0.05 were considered statistically significant.
Results
Retrograde tract tracing
To establish baseline controls for normal sensory axon distribution from either the SB or the MB of the femoral nerve (Fig. 1A), we applied distinct retrograde tracers to each branch simultaneously in normal animals. For labeling, SB and MB were cut and the proximal ends were exposed to Dextran AlexaFluor 555 (F555, red fluorescent) and Dextran AlexaFluor 488 (F488, green fluorescent) for 45 min, respectively (Fig. 1C-i). Within 48 hours, these tracers could easily be identified in dorsal root ganglia (DRG). Retrograde labeling (RL) of DRG neurons with these two colors (F555 and F488) designated sensory neurons that normally project to either SB (SB neruons) or MB (MB neurons), respectively. Cell counts showed that 1790.0 ± 235.0 sensory neurons were labeled red from F555 applied to the SB, whereas 990.0 ± 170.0 sensory neurons were labeled green corresponding to MB neurons. There were no double-labeled neurons, indicating that under normal conditions these neurons do not dually innervate both branches. These results are similar to those observed in previous study (Brushart, et al., 2005).
Since neuronal labeling depends on the amount of tracer transported from the cut site to DRGs, tracer transportation time might influence the number of labeled neurons. To assess this possibility, we waited 7 days, instead of 48 hours, for tracer transportation before DRG processing and cell counting. Neuronal counts showed the number of labeled DRG neurons dramatically increased, especially those from the SB, demonstrating more accurate retrograde labeling of DRG neurons at one week compared to 48 hours. Under these conditions, the number of DRG neurons projecting to SB (SB neurons) was ~ 3 times those projecting to MB (MB neurons) (Fig. 1D, E). These results are similar to previous observation that most retrograde tracers show maximal labeling at 1 week in the facial nerve (Choi, et al., 2002). Thus, unlike previous femoral nerve studies, we waited 7 days for tracer transport for all of the following tracing experiments in this study.
Sensory axon regeneration in transected femoral nerves
For femoral nerve lesions, the left femoral nerve was transected 6 – 8 mm proximal to the bifurcation of sensory and motor branches. The severed stumps were sutured into a guidance tube with a gap about 0.5 mm in length, to disrupt realignment of axons between the stumps (Fig. 1B, C). Three weeks after nerve repair, regeneration of sensory axons was evaluated using double retrograde tracing of the sensory and motor branches (Fig. 1C-ii). Cell counts showed the total number of retrograde labeled DRG neurons significantly decreased after femoral nerve transection compared to the sham control group (Fig. 1D), representing incomplete sensory axon regeneration. Projections from regenerating sensory neurons growing into either the sensory and motor branches (SB / MB distribution) were determined by the ratio of F555-to F488-labeled DRG neurons. There was a dramatic drop in the SB / MB ratio in injured animals compared to sham control animals (Fig. 1E), indicating misrouting of sensory axons into the sensory and motor branches during regeneration. These results are consistent with previous studies that examine regeneration of motor axons from lesioned femoral nerves (Brushart, et al., 2005).
Femoral transection at the site depicted above also influences two other nerve branches, which have been usually ignored in previous studies. One branch is a local cutaneous branch (CB) that separates from the saphenous branch between the femoral nerve bifurcation and the distal sites where tracer is applied. Thus, we are not able to trace these axons by applying retrograde tracer on the cut end of the saphenous branch. The other branch is the pectineus nerve (PN), which separates from the femoral trunk distal to the transection site but proximally to the bifurcation (Fig. 2A). Since the axons in these two branches are also cut by femoral nerve transection, it is high probable regenerating axons originally from the CB or the PN would be misrouted into either the sensory or motor branch and reduce the overall accuracy of labeling counts.
The numbers of L2 – L4 sensory neurons which project to CB and PN in normal animals were evaluated by double retrograde tracing at CB and PN. Briefly, both CB and PN were cut 2 mm distal to their branch point from the femoral trunk, proximal cut ends were treated with F555 and F488, respectively. Seven days later, L2 – L4 DRGs were processed and labeled neurons counted. Neuron counts revealed 105.0 ± 100.0 sensory neurons projected to the PN, whereas 2520.0 ± 885.0 sensory neurons projected to the CB (n = 2). Based on the high number of CB neurons, we cannot ignore that CB axons could reinnervate either SB or MB and artifactually inflate misrouting counts. Since these sensory axons cannot be distinguished from those traversing either the sensory or motor branch, using standard retrograde tracing technique, we modified our method to pre-label axons from the saphenous branch using FluoroGold (FG). This will allow us to identify all neurons that initially innervate the saphenous branch, but no other branches.
Sequential triple retrograde tracing is a necessary tool to assess the reinnervation accuracy of DRG neurons
As described above, regenerating sensory axons after femoral transection mainly originated from the cutaneous, sensory, and motor branches, which intermingled during regeneration (Fig. 2A). To evaluate the accuracy of axon reinnervation, sequential triple retrograde tracing was performed to label pre-lesion SB neurons and post-lesion reinnervating pathways, as summarized in Fig. 2A. Neuron counts from sham controls showed only 3.01 ± 0.58% of those labeled with F555 were not initially labeled by FG, demonstrating more than 96% sensitivity for this technique (Table 2). Likewise with triple labeling, the ratio of F555-to F488-labeled DRG neurons was 3.52 ± 0.31 (Fig. 4A), similar to the ratio estimated by simultaneous double retrograde tracing in sham control group (Fig. 1E). These data demonstrate that sensory axons regenerating into either the sensory or motor branches were not significantly influenced by a conditioning effect of the 1st labeling procedure. However, only 86.37 ± 4.26% of FG-labeled neurons were co-labeled via 2nd RL, indicating potential dieback of saphenous branch axons after the 1st RL (Fig. 4C). Assessment of pathway selection showed 99.4 ± 0.14% of the labeled neurons from the saphenous branch remained in the saphenous branch in sham controls (Fig. 4D). These findings suggest sequential triple retrograde tracing is necessary for measuring accuracy for sensory neuron reinnervation of either the sensory or motor nerve branches, without increasing the number of regenerating axons or their pathway selection.
Table 2.
Number of L2 – L4 DRG neurons labeled by F555 and F488, respectively, 4 or 9 weeks after FG pre-labeling in sham animals, as baseline control study for sequentially triple retrograde tracing
| Ctrl | FG+ (SB axons) |
FG- (Non-SB axons) |
|||||
|---|---|---|---|---|---|---|---|
| F555+F488- (SB only) | F488+F555- (MB only) | F555+F488+ (Dual) | F555-F488- (not regenerated) | F555+F488- (SB only) | F488+F555- (MB only) | F555+F488+ (Dual) | |
| 4 w | 2882 ± 389 | 3.8 ± 2.4 | 12.5 ± 4.3 | 453 ± 157 | 83 ± 16 | 892 ± 209 | 2.5 ± 2.5 |
| 9 w | 2156 ± 474 | 31.7 ± 13.6 | 10.0 ± 7.6 | 397 ± 96 | 810 ± 254* | 1015 ± 230 | 3.3 ± 1.7 |
Values represent mean ± S.E.M., n = 3 - 4.
p < 0.05 comparing with 4 w group, analyzed by Student’s t-test.
SB, saphenous branch; MB, motor branch; FG, FluoroGold; F555, Dextran AlexaFluor 555; F488, Dextran AlexaFluor 488
Figure 4.
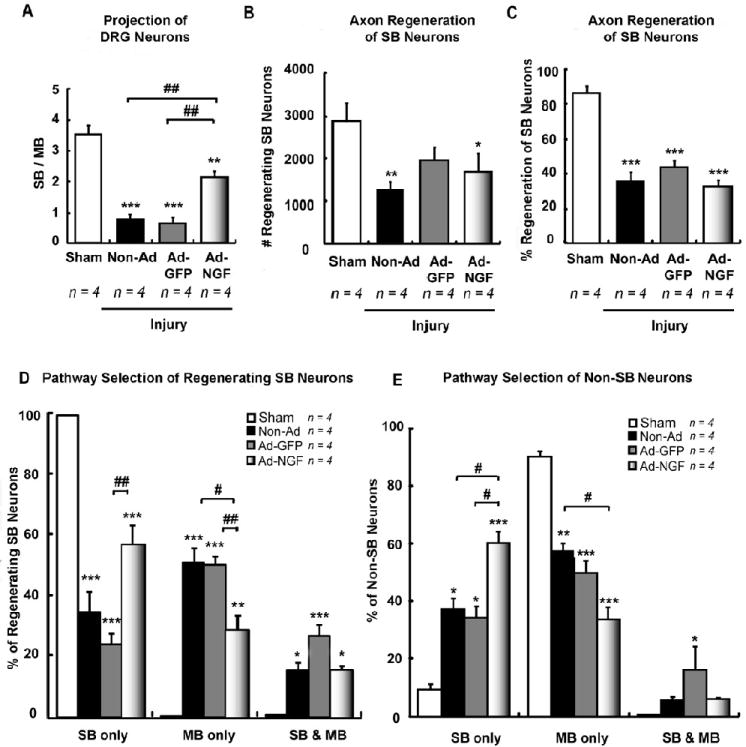
Ad-NGF administration significantly increased the accuracy of SB sensory neuron reinnervation 3 weeks post-injury. Femoral transection, adenoviral administration, and sequentially triple retrograde tracing were performed as illustrated in figure 2A. L2 – L4 DRGs were processed at 3 weeks post-injury. A, The ratio of F555 to F488 labeled DRG neurons (either in the presence or absence of FG-labeling) indicate the ratio of sensory neurons with axons extending into either SB and MB. Statistical results showed Ad-NGF administration significantly increased sensory neuron reinnervation into SB, compared to non-Ad and Ad-GFP groups. B – C, The number of F555/FG+ and F488/FG+ co-labeled neurons designate the number or percent of regenerating SB neurons. Injection of adenoviruses did not significantly affect the number of regenerating SB neurons (B) as well as the percentage of regenerating SB neurons (C). D – E, Pathway selection for sensory neuron reinnervation was identified and calculated by counting of multi-labeled DRG neurons. The percentages of F555, F488, F555 & F488 labeling corresponds to the proportion of reinnervating SB, MB, and dual pathways, respectively. Injection of Ad-NGF along post-traumatic SB significantly increased the accuracy of SB neuron reinervation (D). Interestingly, non-SB neurons also preferentially grow into SB with Ad-NGF administration (E). SB, saphenous branch; MB, motor branch; non-Ad, non-adenovirus; Ad-NGF, recombinant adenovirus encoding NGF; Ad-GFP, recombinant adenovirus encoding GFP. Values represent mean ± S.E.M; n = 4. * p < 0.05; ** p < 0.01; *** p < 0.001 compared to sham group; # p < 0.05; ## p < 0.01 compared to non-Ad or Ad-GFP group, analyzed by One-Way ANOVA followed by Tukey post hoc test.
For longer time points, however, FG may not be suitable as it was reported to produce neurotoxic effects on pre-labeled cells by 6 weeks (Naumann, et al., 2000) and began to degrade between 6 weeks to 12 weeks (Akhavan, et al., 2006). To investigate whether sequential retrograde tracing is suitable for long-term studies, 1st RL and 2nd RL were performed at an interval of 9 weeks in sham control animals (Fig. 2A). Data from retrograde labeled DRG neurons showed only 73.7 ± 5.32% of F555-labeled neurons co-labeled with FG; whereas, the number of F555-labeled neurons not colabeled by FG significantly increased, suggesting the loss of FG over extended periods of time (Table 2). As it is difficult to distinguish non-SB neurons from those that lost FG, sequential retrograde tracing is not suitable to assess long-term motor branch neuron reinnervation. However, 98.31 ± 0.51% of FG-positive neurons colabeled with F555 from SB (Fig. 5C), showing good double-labeling of cells that retained FG. These results showed sequential triple retrograde tracing is useful to assess reinnervation accuracy of the saphenous branch neurons in long-term studies, although, the overall number of regenerating axons might be slightly underestimated due to loss of FG.
Figure 5.
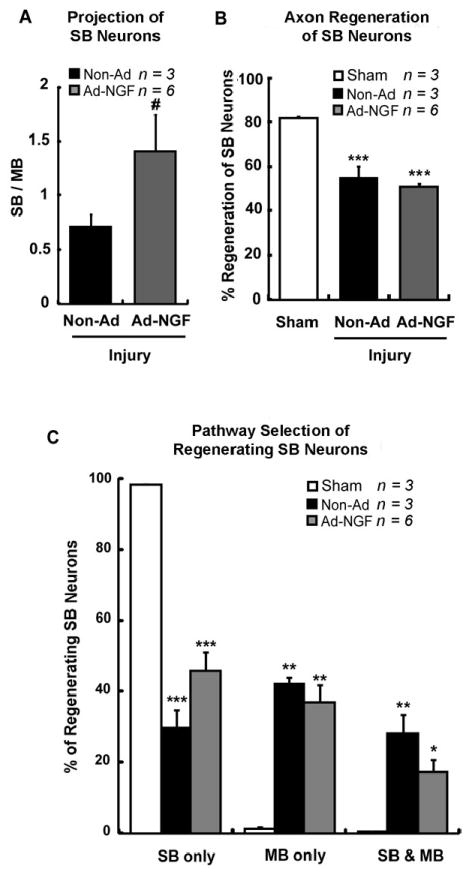
Ad-NGF administration significantly increased the accuracy of SB sensory neuron reinnervation 8 weeks post-injury. Femoral transection, adenoviral administration, and sequentially triple retrograde tracing were performed as illustrated in figure 2A. L2 – L4 DRG were processed at 8 weeks post-injury. A, The ratio of F555 to F488 labeling in FG pool corresponds to regenerating SB neuron projection between SB and MB. Ad-NGF administration significantly increased the SB neuron reinnervation to SB as opposed to MB. B, The percentage of SB neuron regeneration was not significantly changed. C, Ad-NGF administration increased SB neuron regrowth into SB, and decreased SB neuron regrowth into MB or dual branches. No significant difference was detected. SB, saphenous branch; MB, motor branch; Ad-NGF, recombinant adenovirus encoding NGF; non-Ad, non-adenovirus. Values represent mean ± S.E.M; n = 3 – 6. * p < 0.05; ** p < 0.01; *** p < 0.001 compared to sham group; # p < 0.05 compared to non-Ad group, analyzed by Students’ t-test (between only two groups) or One-Way ANOVA followed by Tukey post hoc test (among multiple groups).
Post-traumatic NGF gene therapy significantly increased sensory neuron reinnervation into the saphenous branch
Our previous studies have shown successful overexpression of exogenous genes with administration of recombinant adenoviruses (Romero and Smith, 1998, Tang, et al., 2007). In the current study, cultured astrocytoma cell line U373 was infected with recombinant adenoviruses encoding NGF (Ad-NGF). Western blots of conditioned medium showed NGF production was induced in a dose-dependent manner (Fig. 3A). To test in vivo targeted overexpression of exogenous genes, Ad-NGF and the control virus, recombinant adenoviruses encoding green fluorescent protein (Ad-GFP), were independently injected along the saphenous branch after crushing the femoral nerve. One week after viral injections, tissue segments from the SB, MB, and femoral nerve trunk (Fig. 3B-i) showed a gradient of NGF expression along the SB with a peak at the injection site (Fig. 3B-ii). Fluorescent imaging showed strong GFP expression along SB, but not MB (Fig. 3C) after injection of Ad-GFP into the SB. These results demonstrated that adenoviral administration can induce a gradient of expression for exogenous genes in target tissue area.
To examine the potential effect of NGF on axon regeneration, Ad-NGF was injected along the saphenous branch of the femoral nerve which had been transected 7 days earlier. Two weeks later, simultaneous double retrograde tracing was performed on experimental animals. Counting labeled DRG neurons showed that NGF expression significantly increased the SB/MB ratio, compared to non-adenovirus (non-Ad) treatment (Fig. 3E); eventhough, the total number of regenerating sensory axons did not change (Fig. 3D). These findings demonstrate that NGF overexpression within the saphenous branch increases axon reinnervation into the saphenous branch in a manner independent of promoting axon regeneration. Therefore, NGF can act as a guidance molecule to promote branch point entrance of sensory axons into the saphenous branch. To further evaluate the accuracy of sensory axon regrowth, sequential triple retrograde tracing was done on both pre-lesion and post-lesion pathways.
Accuracy of saphenous branch neuron reinnervation was significantly increased by post-traumatic NGF gene therapy
To evaluate the accuracy of sensory neuron reinnervation, triple sequential labeling was performed. Initially, the SB was labeled using FG, followed by transection of the femoral nerve and repair 1 week later. For sham controls the femoral nerve was not lesioned. One week after femoral transection, the saphenous branch was injected with either Ad-GFP (Ad-GFP-control group), Ad-NGF (Ad-NGF group), or no adenovirus (non-Ad control group; Fig. 2A). Two weeks later the sensory and motor branches were cut and tracer added to each proximal stump and transported to DRG neurons over a one week period. To analyze the accuracy of reinnervation, the numbers of F555-to F488-labeled DRG neurons with axons extending into either saphenous or motor branches, respectively, were counted. In non-Ad control animals, 34.4 ± 6.8% of regenerating SB neurons reinnervated SB, 50.5 ± 4.8% grew into MB, and 15.0 ± 2.8% showed dual reinnervation of both branches, resulting in low accuracy of SB neuron reinnervation post-injury. Treatment of Ad-GFP did not significantly change this innervation pattern (Fig. 4D). Likewise, in the non-Ad group, the SB/MB ratio was 0.78 ± 0.17, which was a similar ratio to that of the Ad-GFP treated group (Fig 4A). In animals receiving Ad-NGF, the proportion of SB neurons selecting the saphenous branch significantly increased to 56.7 ± 6.2%, whereas those selecting MB significantly decreased to 28.4 ± 5.1%, and dual reinnervation did not change (Fig. 4D). The SB/MB ratio, however, significantly increased to 2.15 ± 0.18 with treatment of Ad-NGF (Fig. 4A). The number of total regenerating SB neurons was not significantly changed (Fig. 4B, C). These data demonstrated overexpression of NGF within the saphenous branch significantly increased the accuracy of SB neurons reinnervation without enhancing overall axon regeneration.
Even though there was a significant increase in the accuracy of SB neuron reinnervation, we investigated whether or not non-SB neurons also preferentially regrew into the Ad-NGF treated saphenous branch. For this study, DRG neurons not labeled by FG, but positive for either F555 or F488 tracers were counted. These non-SB neurons were also shown to preferentially reinnervate the saphenous branch expressing NGF, in which more sensory axons grew into the sensory than motor branch (Fig. 4E). The proportion of dual reinnervation, however, did not significantly change (Fig. 4E). This finding indicates that overexpression of NGF induced preferential growth of both SB and non-SB neurons into the saphenous branch. However, it remains unclear whether the non-SB neurons originally project to the motor branch or the cutaneous (also sensory) branch. Since NGF-mediated sensory axon targeting showed similar tendency in SB-neurons (Fig. 4D) and in non-SB neurons (Fig. 4E), we assumed that the proportion of NGF-responsive DRG neurons which express NGF receptor, tropomyosin-related kinase A (TrkA), is roughly equal between saphenous branch and non-saphenous branches (including CB and MB), and this distribution does not change following femoral transection.
Although the majority of axons will regenerate within a 3 week time period, regeneration will continue for more than 2 months post-injury (Brushart, et al., 2002). To determine if reinnervation accuracy changes at longer time points, we examined the accuracy of sensory neuron reinnervation at 8 weeks post-injury. Again, triple retrograde tracing was performed on injured animals at an interval of 8 weeks between the femoral nerve transection and 2nd retrograde labeling (Fig. 2A). As discussed before, this long-term triple retrograde tracing is useful to estimate reinnervation of SB neurons, but not non-SB neurons, due to the loss of tracer. In the non-Ad group, the SB / MB ratio for regenerating SB neurons was calculated to be 0.71 ± 0.11, which was significantly increased to 1.41 ± 0.34 with Ad-NGF administration (Fig. 5A). Further analysis of pathway selection showed 45.7 ± 5.3% regenerating SB neurons selectively reinnervated the saphenous branch in the Ad-NGF group, which increased from 29.8 ± 4.9% in animals with no viral treatment. The proportion of neurons reinnervating MB or both pathways decreased with Ad-NGF treatment, although no significant difference was detected when compared to non-Ad group (Fig. 5C). In the Ad-NGF treated groups, selective SB reinnervation at 3 weeks is approximately 20% greater than at 8 weeks. This difference matches the general loss of FG tracer during this period. As previously observed, the total number of regenerating neurons was not significantly altered by Ad-NGF treatment compared to non-Ad treatment (Fig. 5B). These findings showed post-traumatic NGF gene therapy significantly increased the accuracy of SB neuron reinnevation within nerve repair of 8 weeks, in a manner independent of promoting axon regeneration.
Ad-NGF treatment induces preferential reinnervation of CGRP-positive DRG neurons into the saphenous branch
Calcitonin-related gene peptide (CGRP) positive DRG neurons represent a subset of nociceptive neurons that express TrkA, and require NGF for survival during development (Thippeswamy, et al., 2005). To assess how regenerating CGRP-positive neurons respond to NGF gene therapy, immunostaining for CGRP was performed on the same DRG cryosections which had been analyzed for reinnervation accuracy 3 and 8 weeks post-injury. Co-labeling of CGRP and FG represents nociceptive SB neurons and F555-and F488-labeling designates regenerating neurons project to either SB or MB, respectively (Fig. 6A). CGRP+ SB neuron reinnervation was estimated by counting the multi-labeled DRG neurons (Fig.6A). Three weeks after femoral transection and repair, approximately 25% of FG-positive (SB) DRG neurons expressed CGRP. This proportion did not change with treatment of Ad-NGF or Ad-GFP (Fig. 6C). In sham control, the percentage of SB neurons expressed CGRP showed no significant difference (~ 17%) compared to injury groups, demonstrating that femoral transected injury did not influence CGRP expression. Only about 40% of CGRP+ SB neurons appeared to regenerate post-injury, either in the presence or absence of adenoviral administration (Fig. 6D). In the non-Ad control group, the SB / MB ratio for regenerating CGRP+ SB neurons was 0.65 ± 0.15, which was significantly increased to 1.85 ± 0.48 with treatment of Ad-NGF, but not Ad-GFP (Fig. 6B). Eight weeks after nerve transection, CGRP expression in SB neurons did not change significantly compared to that after 3 weeks either in the presence or absence of Ad-NGF treatment (Fig. 6F). The percentage of regenerating CGRP+ SB neurons was 54.8 ± 4.9% in non-treated animals, not significantly different from that for nerve repair after 3 weeks. This proportion of regeneration did not significantly change with Ad-NGF treatment (Fig. 6G). However, the SB / MB ratio for the distribution of regenerating CGRP+ SB axons was significantly increased with administration of Ad-NGF comparing with non-treated animals (Fig. 6E). These data demonstrated post-traumatic NGF gene therapy significantly increased the accuracy of CGRP-positive SB axon reinnervation.
Figure 6.
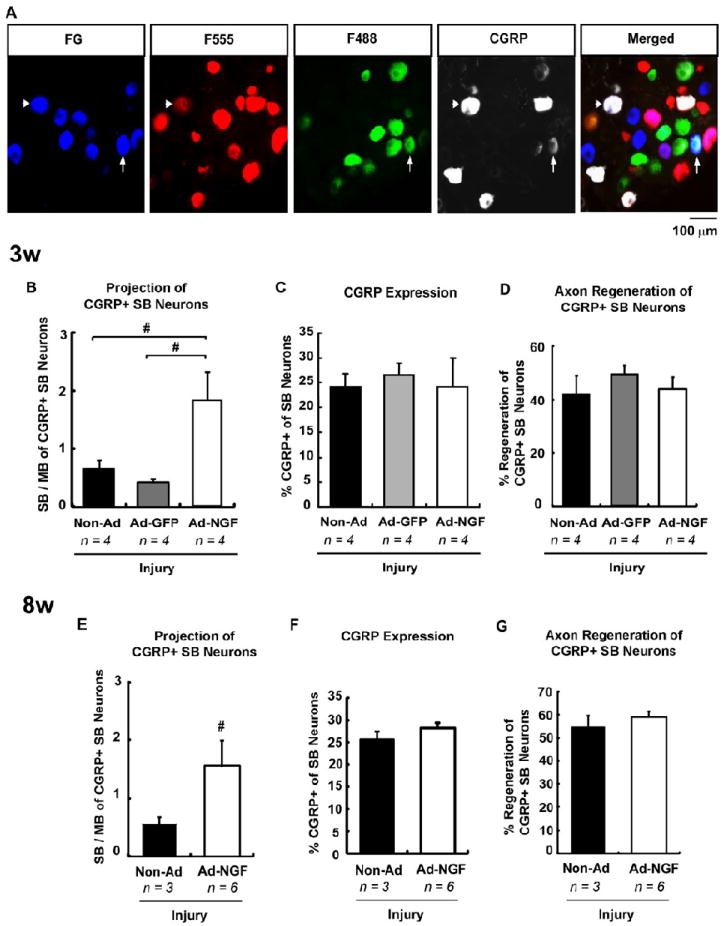
Ad-NGF administration significantly increased accuracy of CGRP-positive nociceptive neuron reinnervation. The cryosections from L2 – L4 DRG were immunostained with CGRP, a specific marker for peptidergic nociceptive axons. A, Co-labeling of FG and CGRP corresponds to CGRP+ SB neurons. F555- and F488-labeling designates neuron reinnervating SB or MB, respectively. The fluorescent images showed CGRP+ SB neurons reinnervated either SB or MB. B – D, Neuron reinnervation 3 weeks post-injury. Ad-NGF administration significantly increased the ratio of SB / MB projection of CGRP+ SB neurons (B). However, the expression of CGRP (C) and the regeneration of CGRP+ SB neurons (D) did not change significantly. E – F, Neuron reinnervation 8 weeks post-injury. The ratio of SB / MB projection of CGRP+ SB neurons significantly increased with Ad-NGF administration (E). Ad-NGF application did not influence CGRP expression (F) and regeneration of CGRP+ neurons (G). Arrowhead, CGRP+ SB neurons reinnervated SB; arrow, CGRP+ SB neurons reinnervated MB. Values represent mean ± S.E.M; n = 3 – 6. # p < 0.05 compared to non-Ad or Ad-GFP group, analyzed by Students’ t-test (between only two groups) or One-Way ANOVA followed by Tukey post hoc tests. CGRP, calcitonin gene-related peptide; SB, saphenous branch; MB, motor branch; non-Ad, non-adenovirus; Ad-NGF, recombinant adenovirus encoding NGF; Ad-GFP, recombinant adenovirus encoding GFP. Scale bar, 100 μm.
Discussion
Unlike the central nervous system, the peripheral nerve system retains a robust ability to regenerate even after complete transection. However, the recovery of function remains low, partly due to misrouting of regenerating axons to functionally inappropriate target locations. This can be attributed to the lack of appropriate environmental cues for axons to make accurate pathway choices, especially when they encounter branches. With this in mind, we set forth to evaluate the possibility of overexpressing guidance factors near branch points to enhance targeting into distal pathways. We adapted the femoral nerve lesion model to investigate potential guidance cues, because this relatively simple model could easily be used to estimate sensory and motor pathway decisions during axon regeneration. Although many molecules may direct axon growth (Charron and Tessier-Lavigne, 2007, Koncina, et al., 2007, Reber, et al., 2007), NGF was selected in the current study because of its well-known function in sensory axon development (Zhang, et al., 1994), regeneration (Romero, et al., 2001), and sprouting (Cameron, et al., 2006, Tang, et al., 2007). The expression of NGF near the branch point of the saphenous nerve established a gradient that extended into the parent trunk of the femoral nerve to attract sensory axons into that branch prior to reaching the nerve branch. Our results demonstrate the accuracy of sensory axon reinnervation in the transected femoral nerve is significantly increased using this model. This finding suggests that the application of guidance molecules can alter pathway decision during the course of peripheral nerve regeneration to reduce misrouting of axons at selective branch points. We also observed that NGF overexpression along the saphenous nerve not only induced the regeneration of these neurons back into their original branch, but also sensory neurons from the motor branch to regenerate inappropriately into the saphenous nerve branch. These neurons have a similar neurotrophin specificity but normally target a different nerve branch. Therefore, pathway selection is mostlikely dependent on multiple factors and the use of a single guidance molecule is not sufficient to enhance pathway selection of a single neuronal population.
During regeneration, sensory and motor axons are dispersed throughout the cross sectional area of the transected femoral nerve (Brushart, 1988) and motor axons preferentially reinnervate the motor rather than the saphenous branch (preferential motor reinnervation, PMR) (Brushart, 1988, Brushart, 1993), even if intentional misrotation or a gap was imposed between proximal and distal stumps (Brushart, 1988). This may be due to a specific interaction between regenerating motor axons and Schwann cells, which are thought to maintain a memory of the motor pathway or by selective pruning of the motor neurons that fail to reach their appropriate target (Redett, et al., 2005). Likewise, gene expression profiles of cutaneous sensory or ventral motor nerves display different growth factor expression patterns after injury further supporting the Schwann cell memory hypothesis (Hoke et al., 2006). However, classic PMR can be reversed by preventing muscle contact in mice (Madison, et al., 2007, Madison, et al., 2009, Uschold, et al., 2007) demonstrating an important role of the target in stabilizing appropriate connections. Madison et al., (2009) also showed that removal of Schwann cells in injured femoral nerve resulted in no significant difference of PMR while muscle contact was maintained, further demonstrating the role for muscle contact in PMR. In the absence of muscle contact, shortening the saphenous nerve pathway enhanced PMR (Uschold, et al., 2007); whereas, removal of Schwann cells abolished pathway differences for regenerating motor neurons (Madison et al., 2009). These results indicate reinnervation accuracy of motor neurons is hierarchal with muscle contact showing the highest influence most likely due to the formation and stability of nerve terminals, followed by the influence of Schwann cells in terminal branches, which may provide a sense of directionality to the regenerating axons. Considering both the muscle and Schwann cells in distal pathways can express guidance factors that diffuse along the pathway toward bifurcated point, we hypothesis that accuracy in pathway selection to appropriate targets could be enhanced by the initial influenced by Schwann cell guidance molecules in the distal pathways that are stabilized as the axons reach their appropriate targets. For our experiments, we only attempted to influence the initial pathway selection for a subpopulation of sensory axons at a single branch point by enhancing the balance of neurotrophins for that neuronal population. Our data indicates that the initial accuracy of sensory neuron reinnervation is strongly influenced by the guidance environment near branch points and supports the general assumption of a “trophic balance hypothesis”, at least for the initial pathway choice that axons make when encountering branches.
Axon regeneration in the injured peripheral nerve is a complicated process. Transection completely disrupts the continuity of endoneurial tubes at the lesion site, disrupting the pre-existing default pathways regenerating axons could follow (Nguyen, et al., 2002). Instead, sprouts continually emerge from severe axons, travel across the lesion, and enter endoneurial tubes in the distal nerve stump (Ide, 1996). Even with alignment to match original (pre-lesion) endoneurial tubes, many axons often travel laterally across the face of the distal stump before choosing a pathway, and this lateral growth prevents them from entering their original endoneurial tubes (Witzel, et al., 2005). There are two possible mechanisms by which regenerating axons can be redistributed at distal locations for retargeting. Early during the regeneration process axons arborize and send multiple axons into different endoneurial tubes and pathways. Over time, axonal sprouts that have grown into inappropriate pathways are pruned (Pruning Hypothesis); whereas, those traversing more appropriate pathways are conserved (Morris, et al., 1972, Witzel, et al., 2005). For the second mechanism, proteases produced by macrophages, Schwann cells, and fibroblasts are thought to make the basal lamina more porous allowing growth cones to change direction or grow less restricted between distal endoneurial tubes (Fu and Gordon, 1997, Ide, 1996). At distal bifurcated sites, growth cones could use localized guidance molecules to make choices to innervate specific target pathways. Such a process would be greatly influenced by the neurotrophins Schwann cells produce, and their maintenance of a “neurotrophin memory” of the motor or sensory axons they originally supported would benefit form this mechanism. Our data, however, indicated that both mechanisms play a role in pathways selection when guidance molecules are placed distal to a branch point, since we observe NGF to reduce the numbers of neurons retrogradely labeled either from the MB only or both branches. Under such a condition, axon sprouts would be enticed to preferential grow into the branch with the highest neurotrophic support. Misrouted axon sprouts would have reduced neurotrophic support as they grew into the inappropriate branch and become destabilized and retract. This effect, however, requires a neurotrophin gradient to emanate for the appropriate branch, be specific to the neuronal population of interest, and show a much lower expression within the inappropriate branch.
To induce gradient formation from a single branch, we made a series of micro-injections of Ad-NGF to produce a localized cellular source of NGF expression. The secretion and diffusion of NGF from these cells is what established the gradient. The NGF gradient near the bifurcation dramatically increased SB neuron reinnervation 3 weeks post-injury. Although the enhanced accuracy dropped slightly 7 weeks after Ad-NGF application, it remained significant different from controls (Fig. 5). These results are consistent with the temporal pattern of adenoviral-based gene overexpression, which usually diminishes rapidly after three weeks (Romero and Smith, 1998). The expression of NGF within the gradient would signal through the TrkA-receptor, which is specifically expressed on adult CGRP-positive nociceptive axons (Bergman, et al., 2000, Ernfors, et al., 1994). Thus, an expression gradient of NGF should increase pathway selection of CGRP-positive axons without altering neuropeptide expression or guidance of the other sensory axon subpopulations (Romero, et al., 2000, Tang, et al., 2004). In the present study, Ad-NGF administration selectively increased the accuracy of CGRP+ SB neuron reinnervation without changing the expression of CGRP. However, not all SB neurons expressing CGRP were attracted to NGF-overexpressing SB (Fig. 6). This indicates that some CGRP-positive axons were not influences by the NGF gradient, either because they did not come in contact with it, or they require additional factors for pathway selection. For example, the preferential targeting of motor axons also is though to requires multiple factor since it has been observed that Schwann cells within motor branches express higher levels of motor neurons selective neurotrophins (Hoke et al., 2006), and motor axons expressing polysialic acid (PSA) to allow defasciculation of axons and enhance pathway choice determination (Franz, et al., 2008). Thus, the accuracy of motor axon reinnervation appears dependent multiple factors. Whether, similar combinations of other factors would increase the selectivity of sensory neuron reinnervation need to be tested in further experiments.
Neurotrophins have been shown to promote and direct the motility of grow cones, which are located at the leading edge of developing and regenerating nerve processes (Farrar and Spencer, 2008). Cultured DRG neuronal growth cones rapidly turn and migrate toward a localized source of NGF via the local activation of TrkA (Gallo, et al., 1997). The downstream activated Cdc42/Rac/Rho protein family, PI3K, PLC-γ signaling is essential for triggering chemoattraction in an NGF gradient (Ming, et al., 1999, Patapoutian and Reichardt, 2001), that is, determining the leading direction of growth cones. Interestingly, growth cones proceed to migrate in the same direction after having contacted NGF-expressing source even if the source is then removed from its original location (Gallo, et al., 1997). This observation shows the gradient of NGF determines the migrating direction but not the termination point of growth cones. The migrating distance may be decided by both the total amount of guidance cues which had encountered growth cones in the gradient as well as the local concentration around the growth cones (Farrar and Spencer, 2008). Such integration may provide advantages in vivo to withstand defects that might otherwise result from slight disruptions of the endogenous gradients. Consistently, our previous studies show induced axon growth along performed NGF-expressing pathway over long distance, although the gradient of NGF may wave among distinct Ad-NGF injection sites (Ziemba, et al., 2008). These observations lay the groundwork for elucidating the possible mechanisms by which exogenous NGF application targets sensory axon regeneration.
In summary, we demonstrate that in vivo gene transfer of NGF near the branch point can mediate the pathway choice of regenerating sensory axons, directing them toward targeted pathway in transected nerve. This finding provides potential therapeutic strategy after severe peripheral nerve injury to guide regenerating axons to their functionally appropriate target locations.
Acknowledgments
We deeply appreciate Dr. Edward Hall and Dr. Jinhui Chen for kindly providing devices for our experiments. The current work is contributed by NIH/NINDS R01NS40592 and the Kentucky Spinal Cord and Head Injury Research Trust 6:13.
Abbreviations
- Ad
adenoviruses
- Ad-NGF
recombinant adenoviruses encoding nerve growth factor
- Ad-GFP
recombinant adenoviruses encoding green fluorescent protein
- CB
cutaneous branch
- CGRP
calcitonin gene-related peptide
- DRG
dorsal root ganglia
- F488
Dextran AlexaFluor 488
- F555
Dextran AlexaFluor 555
- FG
FluoroGold
- FN
femoral nerve
- GFP
green fluorescent protein
- MB
motor branch
- NGF
nerve growth factor
- PN
pectineus nerve
- RL
retrograde labeling
- SB
saphenous branch
- ST
silicon tubing
- TrkA
tropomyosin-related kinase A
Footnotes
Publisher's Disclaimer: This is a PDF file of an unedited manuscript that has been accepted for publication. As a service to our customers we are providing this early version of the manuscript. The manuscript will undergo copyediting, typesetting, and review of the resulting proof before it is published in its final citable form. Please note that during the production process errors may be discovered which could affect the content, and all legal disclaimers that apply to the journal pertain.
References
- 1.Ahlborn P, Schachner M, Irintchev A. One hour electrical stimulation accelerates functional recovery after femoral nerve repair. Exp Neurol. 2007;208:137–144. doi: 10.1016/j.expneurol.2007.08.005. [DOI] [PubMed] [Google Scholar]
- 2.Akhavan M, Hoang TX, Havton LA. Improved detection of fluorogold-labeled neurons in long-term studies. J Neurosci Methods. 2006;152:156–162. doi: 10.1016/j.jneumeth.2005.09.010. [DOI] [PubMed] [Google Scholar]
- 3.Bergman E, Ulfhake B, Fundin BT. Regulation of NGF-family ligands and receptors in adulthood and senescence: correlation to degenerative and regenerative changes in cutaneous innervation. Eur J Neurosci. 2000;12:2694–2706. doi: 10.1046/j.1460-9568.2000.00149.x. [DOI] [PubMed] [Google Scholar]
- 4.Brushart TM. Preferential reinnervation of motor nerves by regenerating motor axons. J Neurosci. 1988;8:1026–1031. doi: 10.1523/JNEUROSCI.08-03-01026.1988. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Brushart TM. Preferential reinnervation of motor nerves by regenerating motor axons. J Neurosci. 1988;8:1026–1031. doi: 10.1523/JNEUROSCI.08-03-01026.1988. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Brushart TM. Motor axons preferentially reinnervate motor pathways. J Neurosci. 1993;13:2730–2738. doi: 10.1523/JNEUROSCI.13-06-02730.1993. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Brushart TM. Motor axons preferentially reinnervate motor pathways. J Neurosci. 1993;13:2730–2738. doi: 10.1523/JNEUROSCI.13-06-02730.1993. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Brushart TM, Hoffman PN, Royall RM, Murinson BB, Witzel C, Gordon T. Electrical stimulation promotes motoneuron regeneration without increasing its speed or conditioning the neuron. J Neurosci. 2002;22:6631–6638. doi: 10.1523/JNEUROSCI.22-15-06631.2002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Brushart TM, Jari R, Verge V, Rohde C, Gordon T. Electrical stimulation restores the specificity of sensory axon regeneration. Exp Neurol. 2005;194:221–229. doi: 10.1016/j.expneurol.2005.02.007. [DOI] [PubMed] [Google Scholar]
- 10.Cajal Ry. Degeneration and regeneration of the nervous system. New York: Hafner; 1928. [Google Scholar]
- 11.Cameron AA, Smith GM, Randall DC, Brown DR, Rabchevsky AG. Genetic manipulation of intraspinal plasticity after spinal cord injury alters the severity of autonomic dysreflexia. J Neurosci. 2006;26:2923–2932. doi: 10.1523/JNEUROSCI.4390-05.2006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Charron F, Tessier-Lavigne M. The Hedgehog, TGF-beta/BMP and Wnt families of morphogens in axon guidance. Adv Exp Med Biol. 2007;621:116–133. doi: 10.1007/978-0-387-76715-4_9. [DOI] [PubMed] [Google Scholar]
- 13.Choi D, Li D, Raisman G. Fluorescent retrograde neuronal tracers that label the rat facial nucleus: a comparison of Fast Blue, Fluoro-ruby, Fluoro-emerald, Fluoro-Gold and DiI. J Neurosci Methods. 2002;117:167–172. doi: 10.1016/s0165-0270(02)00098-5. [DOI] [PubMed] [Google Scholar]
- 14.Ernfors P, Lee KF, Kucera J, Jaenisch R. Lack of neurotrophin-3 leads to deficiencies in the peripheral nervous system and loss of limb proprioceptive afferents. Cell. 1994;77:503–512. doi: 10.1016/0092-8674(94)90213-5. [DOI] [PubMed] [Google Scholar]
- 15.Farrar NR, Spencer GE. Pursuing a ‘turning point’ in growth cone research. Dev Biol. 2008;318:102–111. doi: 10.1016/j.ydbio.2008.03.012. Epub 2008 Mar 2020. [DOI] [PubMed] [Google Scholar]
- 16.Franz CK, Rutishauser U, Rafuse VF. Intrinsic neuronal properties control selective targeting of regenerating motoneurons. Brain. 2008;131:1492–1505. doi: 10.1093/brain/awn039. [DOI] [PubMed] [Google Scholar]
- 17.Fu SY, Gordon T. The cellular and molecular basis of peripheral nerve regeneration. Mol Neurobiol. 1997;14:67–116. doi: 10.1007/BF02740621. [DOI] [PubMed] [Google Scholar]
- 18.Gallo G, Lefcort FB, Letourneau PC. The trkA receptor mediates growth cone turning toward a localized source of nerve growth factor. J Neurosci. 1997;17:5445–5454. doi: 10.1523/JNEUROSCI.17-14-05445.1997. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Heron PM, Sutton BM, Curinga GM, Smith GM, Snow DM. Localized gene expression of axon guidance molecules in neuronal co-cultures. J Neurosci Methods. 2007;159:203–214. doi: 10.1016/j.jneumeth.2006.07.013. [DOI] [PubMed] [Google Scholar]
- 20.Hoke A, Redett R, Hameed H, Jari R, Zhou C, Li ZB, Griffin JW, Brushart TM. Schwann cells express motor and sensory phenotypes that regulate axon regeneration. J Neurosci. 2006;26:9646–9655. doi: 10.1523/JNEUROSCI.1620-06.2006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Ide C. Peripheral nerve regeneration. Neurosci Res. 1996;25:101–121. doi: 10.1016/0168-0102(96)01042-5. [DOI] [PubMed] [Google Scholar]
- 22.Irintchev A, Simova O, Eberhardt KA, Morellini F, Schachner M. Impacts of lesion severity and tyrosine kinase receptor B deficiency on functional outcome of femoral nerve injury assessed by a novel single-frame motion analysis in mice. Eur J Neurosci. 2005;22:802–808. doi: 10.1111/j.1460-9568.2005.04274.x. [DOI] [PubMed] [Google Scholar]
- 23.Koncina E, Roth L, Gonthier B, Bagnard D. Role of semaphorins during axon growth and guidance. Adv Exp Med Biol. 2007;621:50–64. doi: 10.1007/978-0-387-76715-4_4. [DOI] [PubMed] [Google Scholar]
- 24.Madison RD, Archibald SJ, Brushart TM. Reinnervation accuracy of the rat femoral nerve by motor and sensory neurons. J Neurosci. 1996;16:5698–5703. doi: 10.1523/JNEUROSCI.16-18-05698.1996. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Madison RD, Robinson GA, Chadaram SR. The specificity of motor neurone regeneration (preferential reinnervation) Acta Physiol (Oxf) 2007;189:201–206. doi: 10.1111/j.1748-1716.2006.01657.x. [DOI] [PubMed] [Google Scholar]
- 26.Madison RD, Sofroniew MV, Robinson GA. Schwann cell influence on motor neuron regeneration accuracy. Neuroscience. 2009;6:6. doi: 10.1016/j.neuroscience.2009.05.073. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Ming G, Song H, Berninger B, Inagaki N, Tessier-Lavigne M, Poo M. Phospholipase C-gamma and phosphoinositide 3-kinase mediate cytoplasmic signaling in nerve growth cone guidance. Neuron. 1999;23:139–148. doi: 10.1016/s0896-6273(00)80760-6. [DOI] [PubMed] [Google Scholar]
- 28.Morris JH, Hudson AR, Weddell G. A study of degeneration and regeneration in the divided rat sciatic nerve based on electron microscopy. IV. Changes in fascicular microtopography, perineurium and endoneurial fibroblasts. Z Zellforsch Mikrosk Anat. 1972;124:165–203. doi: 10.1007/BF00335678. [DOI] [PubMed] [Google Scholar]
- 29.Naumann T, Hartig W, Frotscher M. Retrograde tracing with Fluoro-Gold: different methods of tracer detection at the ultrastructural level and neurodegenerative changes of back-filled neurons in long-term studies. J Neurosci Methods. 2000;103:11–21. doi: 10.1016/s0165-0270(00)00292-2. [DOI] [PubMed] [Google Scholar]
- 30.Nguyen QT, Sanes JR, Lichtman JW. Pre-existing pathways promote precise projection patterns. Nat Neurosci. 2002;5:861–867. doi: 10.1038/nn905. [DOI] [PubMed] [Google Scholar]
- 31.Patapoutian A, Reichardt LF. Trk receptors: mediators of neurotrophin action. Curr Opin Neurobiol. 2001;11:272–280. doi: 10.1016/s0959-4388(00)00208-7. [DOI] [PubMed] [Google Scholar]
- 32.Paves H, Saarma M. Neurotrophins as in vitro growth cone guidance molecules for embryonic sensory neurons. Cell Tissue Res. 1997;290:285–297. doi: 10.1007/s004410050933. [DOI] [PubMed] [Google Scholar]
- 33.Reber M, Hindges R, Lemke G. Eph receptors and ephrin ligands in axon guidance. Adv Exp Med Biol. 2007;621:32–49. doi: 10.1007/978-0-387-76715-4_3. [DOI] [PubMed] [Google Scholar]
- 34.Redett R, Jari R, Crawford T, Chen YG, Rohde C, Brushart TM. Peripheral pathways regulate motoneuron collateral dynamics. J Neurosci. 2005;25:9406–9412. doi: 10.1523/JNEUROSCI.3105-05.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Romero MI, Rangappa N, Garry MG, Smith GM. Functional regeneration of chronically injured sensory afferents into adult spinal cord after neurotrophin gene therapy. J Neurosci. 2001;21:8408–8416. doi: 10.1523/JNEUROSCI.21-21-08408.2001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Romero MI, Rangappa N, Garry MG, Smith GM. Functional regeneration of chronically injured sensory afferents into adult spinal cord after neurotrophin gene therapy. J Neurosci. 2001;21:8408–8416. doi: 10.1523/JNEUROSCI.21-21-08408.2001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Romero MI, Rangappa N, Li L, Lightfoot E, Garry MG, Smith GM. Extensive sprouting of sensory afferents and hyperalgesia induced by conditional expression of nerve growth factor in the adult spinal cord. J Neurosci. 2000;20:4435–4445. doi: 10.1523/JNEUROSCI.20-12-04435.2000. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Romero MI, Rangappa N, Li L, Lightfoot E, Garry MG, Smith GM. Extensive sprouting of sensory afferents and hyperalgesia induced by conditional expression of nerve growth factor in the adult spinal cord. J Neurosci. 2000;20:4435–4445. doi: 10.1523/JNEUROSCI.20-12-04435.2000. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Romero MI, Smith GM. Adenoviral gene transfer into the normal and injured spinal cord: enhanced transgene stability by combined administration of temperature-sensitive virus and transient immune blockade. Gene Ther. 1998;5:1612–1621. doi: 10.1038/sj.gt.3300774. [DOI] [PubMed] [Google Scholar]
- 40.Romero MI, Smith GM. Adenoviral gene transfer into the normal and injured spinal cord: enhanced transgene stability by combined administration of temperature-sensitive virus and transient immune blockade. Gene Ther. 1998;5:1612–1621. doi: 10.1038/sj.gt.3300774. [DOI] [PubMed] [Google Scholar]
- 41.Sunderland S. Nerves and Nerve Injuries. 2. Churchill Livingston; New York: 1978. [Google Scholar]
- 42.Tang XQ, Cai J, Nelson KD, Peng XJ, Smith GM. Functional repair after dorsal root rhizotomy using nerve conduits and neurotrophic molecules. Eur J Neurosci. 2004;20:1211–1218. doi: 10.1111/j.1460-9568.2004.03595.x. [DOI] [PubMed] [Google Scholar]
- 43.Tang XQ, Heron P, Mashburn C, Smith GM. Targeting sensory axon regeneration in adult spinal cord. J Neurosci. 2007;27:6068–6078. doi: 10.1523/JNEUROSCI.1442-07.2007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Tang XQ, Heron P, Mashburn C, Smith GM. Targeting sensory axon regeneration in adult spinal cord. J Neurosci. 2007;27:6068–6078. doi: 10.1523/JNEUROSCI.1442-07.2007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Tang XQ, Tanelian DL, Smith GM. Semaphorin3A inhibits nerve growth factor-induced sprouting of nociceptive afferents in adult rat spinal cord. J Neurosci. 2004;24:819–827. doi: 10.1523/JNEUROSCI.1263-03.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Thippeswamy T, McKay JS, Quinn J, Morris R. Either nitric oxide or nerve growth factor is required for dorsal root ganglion neurons to survive during embryonic and neonatal development. Brain Res Dev Brain Res. 2005;154:153–164. doi: 10.1016/j.devbrainres.2004.10.011. [DOI] [PubMed] [Google Scholar]
- 47.Uschold T, Robinson GA, Madison RD. Motor neuron regeneration accuracy: balancing trophic influences between pathways and end-organs. Exp Neurol. 2007;205:250–256. doi: 10.1016/j.expneurol.2007.02.005. [DOI] [PubMed] [Google Scholar]
- 48.Vordemvenne T, Langer M, Ochman S, Raschke M, Schult M. Long-term results after primary microsurgical repair of ulnar and median nerve injuries. A comparison of common score systems. Clin Neurol Neurosurg. 2007;109:263–271. doi: 10.1016/j.clineuro.2006.11.006. [DOI] [PubMed] [Google Scholar]
- 49.Witzel C, Rohde C, Brushart TM. Pathway sampling by regenerating peripheral axons. J Comp Neurol. 2005;485:183–190. doi: 10.1002/cne.20436. [DOI] [PubMed] [Google Scholar]
- 50.Zhang L, Schmidt RE, Yan Q, Snider WD. NGF and NT-3 have differing effects on the growth of dorsal root axons in developing mammalian spinal cord. J Neurosci. 1994;14:5187–5201. doi: 10.1523/JNEUROSCI.14-09-05187.1994. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Zhou FQ, Walzer M, Wu YH, Zhou J, Dedhar S, Snider WD. Neurotrophins support regenerative axon assembly over CSPGs by an ECM-integrin-independent mechanism. J Cell Sci. 2006;119:2787–2796. doi: 10.1242/jcs.03016. [DOI] [PubMed] [Google Scholar]
- 52.Ziemba KS, Chaudhry N, Rabchevsky AG, Jin Y, Smith GM. Targeting axon growth from neuronal transplants along preformed guidance pathways in the adult CNS. J Neurosci. 2008;28:340–348. doi: 10.1523/JNEUROSCI.3819-07.2008. [DOI] [PMC free article] [PubMed] [Google Scholar]