

Abstract
The characterization of adolescence as a time of “storm and stress” remains an open debate. Intense and frequent negative affect during this period has been hypothesized to explain the increased rates of affective disorders, suicide, and accidental death during this time of life. Yet some teens emerge from adolescence with minimal turmoil. We provide a neurobiological model of adolescence that proposes an imbalance in the development of subcortical limbic (e.g., amygdala) relative to prefrontal cortical regions as a potential mechanism for heightened emotionality during this period. Empirical support for this model is provided from recent behavioral and human imaging studies on the development of emotion regulation. We then provide examples of environmental factors that may exacerbate imbalances in amygdala-ventrofrontal function increasing risk for anxiety related behaviors. Finally we present data from human and mouse studies to illustrate how genetic factors may enhance or diminish this risk. Together, these studies provide a converging methods approach for understanding the highly variable stress and turmoil experienced in adolescence.
Keywords: Adolescence, brain, development, genetics, mouse, imaging, environment
Adolescence has been considered, almost by definition, a period of heightened stress (Spear, 2000) due to the many changes experienced concomitantly, including physical maturation, drive for independence, increased salience of social and peer interactions, and brain development (Blakemore, 2008; Casey, Getz, & Galvan, 2008a; Casey, Jones, & Hare, 2008b). Although new-found independence can be stimulating it may also lead to feelings of being overwhelmed by change, which has historically led some researchers to characterize adolescence as ridden with ‘storm and stress’ (Hall, 1904). The controversial ‘storm and stress’ viewpoint is bolstered by reports that the onset of many psychiatric illnesses increases sharply from childhood to adolescence (Compas, Orosan, & Grant, 1993; Kessler, et al., 2005) and by the alarming US health statistics on mortality associated with this time of life. In this paper we highlight recent empirical behavioral, imaging and genetic findings to help explain why some teens are at greater risk for storm and stress during this developmental period. This perspective considers both risk and resilience during adolescence.
Adolescence is characterized by physical maturation of the brain and body, giving rise to intense psychological and physical change. One primary class of psychological change typical of adolescents is an intensification of emotional experiences. These heightened emotional experiences have been argued to be the basis of psychopathology and suicidal behavior. Adolescence is the most common time of life for psychiatric illness to emerge (Kessler et al., 2005), with reported anxiety reaching its lifetime peak (Abe & Suzuki, 1986) and suicide being the fourth leading cause of death (Eaton et al., 2008). These findings, in combination with adult data showing anxiety disorders to be the most common class of psychiatric illness (Kessler et al., 2005), underscores the importance of understanding the developmental and neurobiological substrates that give rise to anxious states and related pathologies (Paus et al., 2008; Pine et al., 2001; Steinberg, 2008).
A number of cognitive and neurobiological hypotheses have been postulated for why adolescence may be a period of heightened turmoil and stress. In a review of the literature on human adolescent brain development, Yurgelun-Todd (2007) suggests that development through the adolescent years is associated with progressively greater efficiency of cognitive control capacities. This efficiency is described as dependent on maturation of the prefrontal cortex as evidenced by increased activity within focal prefrontal regions (Rubia et al., 2000; Tamm, Menon, & Reiss, 2002) and diminished activity in irrelevant brain regions (Brown et al., 2005; Durston et al., 2006).
The general pattern, of improved cognitive control with maturation of the prefrontal cortex (Crone et al 2007), suggests a linear increase in development from childhood to adulthood. Yet the behaviors observed during adolescence represent a nonlinear change that can be distinguished from childhood and adulthood, as evidenced by the National Center for Health Statistics on adolescent behavior and mortality. If immature prefrontal cortex were the basis for teen behavior then children should look remarkably similar or even worse than adolescents, given their less developed prefrontal cortex and cognitive abilities. Thus, immature prefrontal function alone, cannot account for adolescent behavior.
To understand this developmental period, transitions into and out of adolescence are necessary for distinguishing distinct attributes of this period of development (Casey et al. 2008a; 2008b). Any theoretical model of adolescence must account for nonlinear changes–such as deflections or inflections during adolescence relative to both childhood and adulthood. Our laboratory has proposed such a model of brain development that accounts for the unique affective behavioral changes that arise during adolescence (Casey, Getz & Galvan, 2008a; Casey, Jones, & Hare, 2008b; Somerville, Jones, & Casey, 2010).
Our characterization of adolescence goes beyond exclusive association of teen behavior with immaturity of the prefrontal cortex. Instead we suggest that limbic subcortical and prefrontal top-down control regions must be considered together. The cartoon illustrates different developmental trajectories for these systems, with limbic systems developing earlier than prefrontal control regions (see Figure 1). This ‘imbalance’ model proposes that during adolescence, differential timing of brain development induces a disparity between the structural and functional maturity of brain systems critical to affective processing (e.g., subcortical limbic regions including the amygdala), relative to cortical regions of the brain important in control over emotional responses (e.g., the prefrontal cortex). Differential developmental timing of these regions is consistent with nonhuman primate and human postmortem studies showing that the prefrontal cortex is one of the last brain regions to mature (Bourgeois, Goldman-Rakic & Rakic, 1994; Huttenlocher, 1979) while subcortical and sensorimotor regions develop sooner.
Figure 1.

Neurobiological model depicting later development of top down prefrontal regions relative to subcortical limbic regions involved in emotional processes. This imbalance in development of these systems is suggested to be at the core of aberrant teen behavior in contrast to the popular view of adolescent behavior being due to the protracted development of the prefrontal cortex alone (Adapted from Somerville et al., 2010).
Our model is similar in part to other recent models of adolescent development (Ernst, Pine & Hardin, 2006; Nelson, Leibenluft, McClure, & Pine, 2005; Steinberg, 2008) suggesting heightened emotionality during adolescence due to less top-down regulation. However, the present model differs in that it emphasizes the dynamic interplay between subcortical and cortical brain systems across development that results in heightened sensitivity to both appetitive and aversive cues, and accounts for the nonlinearity of adolescent behavior change by integrating findings from children, adolescents and adult into the model.
Empirical support for our imbalance model is provided from recent behavioral and human imaging studies on the development of emotion regulation. We then provide examples of environmental factors that may exacerbate imbalances in amygdala-ventrofrontal function increasing risk for anxiety related behaviors. Finally we present data from human and mouse studies to illustrate how genetic factors may exacerbate or diminish this risk. Together, these studies provide a converging methods approach for understanding the highly variable stress and turmoil experienced in adolescence.
Empirical Studies
Brain Development
The development of the prefrontal cortex is believed to play an important role in the maturation of higher cognitive abilities and goal oriented behavior (Casey, Tottenham, & Fossella 2002b; Casey et al., 1997a). Many paradigms have been used, together with fMRI, to assess the neurobiological basis of these abilities, including go/nogo, flanker, stop signal, and anti saccade tasks (Bunge et al 2002; Casey et al., 1997b; Casey, Giedd, & Thomas, 2000a; Durston et al., 2003; Luna et al., 2001). Collectively, these studies show that children recruit distinct but often larger, more diffuse prefrontal regions when performing these tasks than do adults. The pattern of activity within brain regions central to task performance (i.e., that correlate with cognitive performance) become more focal or fine-tuned with age, while regions not correlated with task performance diminish in activity with age. This pattern has been observed across both cross-sectional (Brown et al., 2005) and longitudinal studies (Durston et al., 2006) and across a variety of paradigms.
Although neuroimaging studies cannot definitively characterize the mechanism of such developmental changes (e.g. dendritic arborization, synaptic pruning) the findings reflect development within, and refinement of, projections to and from, activated brain regions with maturation and suggest that these changes occur over a protracted period of time (Brown et al., 2005; Bunge, Dudukovic, Thomason, Vaidya, Gabrieli, 2002; Casey et al., 1997a; Casey, Thomas, et al., 2002a; Crone, Donohue, Honomichl, Wendelken & Bunge, 2006; Luna, et al., 2001; Moses et al., 2002; Schlaggar et al., 2002; Tamm et al., 2002; Thomas, Hunt, Vizueta, Sommer, Durston, Yang, 2004; Turkletaub, Gareau, Flowers, Zeffiro & Eden, 2003).
Differential recruitment of prefrontal and subcortical regions has been reported across a number of developmental fMRI studies (Casey et al., 2002b; Luna et al 2001; Monk et al., 2003; Thomas et al., 2004). Outside of the functional neuroimaging literature, there is evidence to suggest a differential relative maturity of subcortical limbic brain structures as compared to prefrontal regions, which may be most pronounced during adolescence. Evidence for the continued pruning of prefrontal cortical synapses well into development has been established in both nonhuman primates and humans (Huttenlocher, 1997; Rakic, Bourgeois, Eckenhoff, Zecevic, & Goldman-Rakic, 1986), with greater regional differentiation suggested in the human brain (Huttenlocher, 1997) such that cortical sensory and subcortical areas undergo dynamic synaptic pruning earlier than higher-order association areas. This conceptualization of cortical development is consistent with anatomical MRI work demonstrating protracted pruning of gray matter in higher-order prefrontal areas that continue through adolescence (e.g., Giedd et al., 1999) relative to subcortical regions. Volumetric analyses of the human amygdala shows a substantially reduced slope of change magnitude relative to cortical areas in 4–18 year olds (Giedd et al., 1996). Taken together, these findings suggest a protracted developmental time course of the prefrontal cortex relative to subcortical regions.
Intense and frequent negative affect common during the early adolescent years, (Pine et al., 2001; Silveri et al., 2004; Steinberg, 2005) has led to a number of imaging studies of affect in adolescents (Baird et al., 1999; Killgore et al., 2005; Thomas et al., 2001; Yang et al., 2002). These show that the amygdala is engaged by affective cues, with exaggerated response magnitudes in adolescents relative to children or adults (Ernst et al., 2005; Guyer et al., 2008; 2009; Monk et al., 2003; Rich et al., 2006; Williams et al., 2006). In concert with these findings is the established role of the prefrontal cortex in the regulation of emotive behavior and its protracted maturation throughout adolescence (Galvan et al., 2006; Monk et al., 2003).
These findings suggest that exaggerated emotional reactivity during adolescence might increase the need for top-down control and put individuals with less control at greater risk for poor outcomes. To test this hypothesis, we examined the association between emotion regulation and frontoamygdala circuitry in 60 children, adolescents, and adults with an emotional go-nogo paradigm (Hare et al. 2005) and functional magnetic resonance imaging (fMRI). We went beyond examining the magnitude of neural activity and focused on neural adaptation within this circuitry across time with functional magnetic resonance imaging (Hare et al., 2008). Because individual differences in emotional reactivity might put some teens at greater risk during this sensitive transition in development, we also assessed everyday anxiety using the Speilberger Trait Anxiety Inventory.
Our results showed that adolescents have an initial, exaggerated amygdala response to cues that signal threat (fearful faces) relative to children and adults (see Figure 2 above). This age-related difference decreased with repeated exposures to the stimuli. Anonymous self report anxiety ratings predicted the extent of adaptation or habituation in the amygdala to empty threat. Individuals with higher trait anxiety showed less habituation over repeated exposures. This failure to habituate was associated with less functional connectivity between ventral prefrontal cortex (vPFC) and amygdala.
Figure 2. Exaggerated Amydala Response in Adolescents.
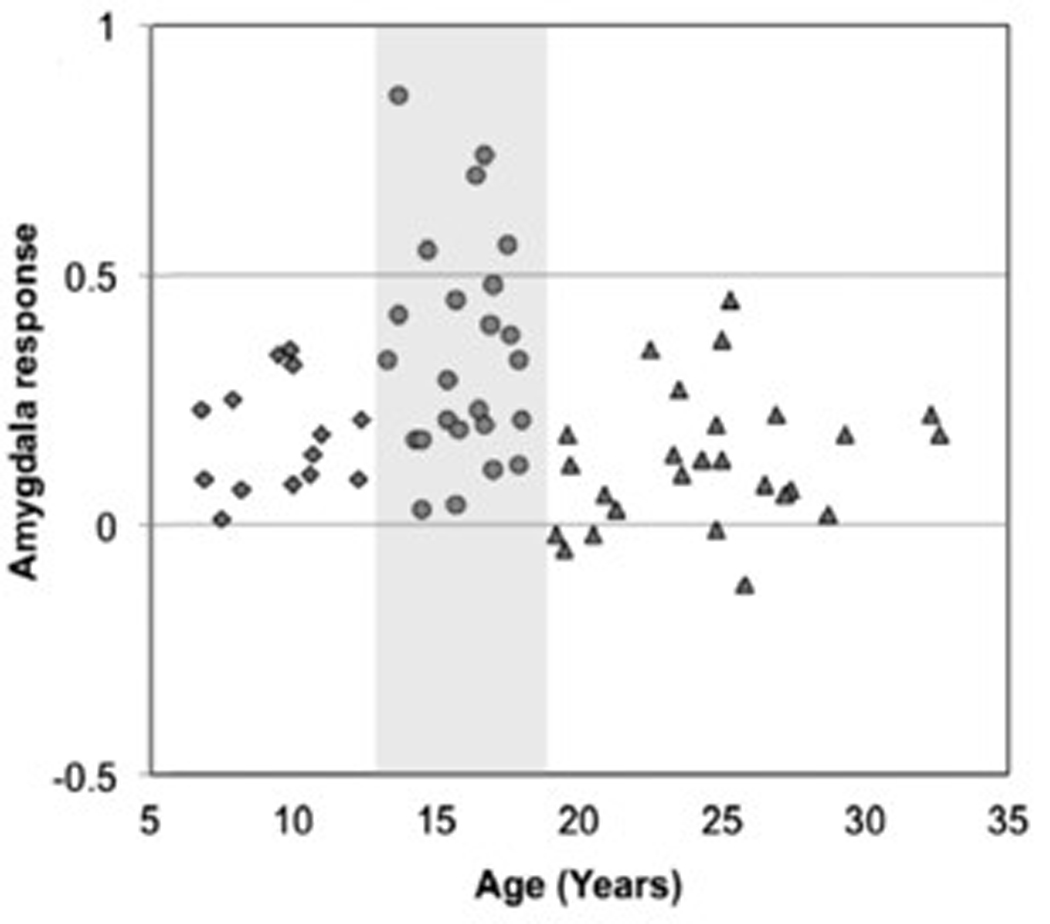
Amygdala response to empty threat (fearful faces) as a function of age. Adapted from Hare et al., 2008
This observed amygdala-vPFC network, showing imbalanced activity in adolescents, is consistent with a wide variety of work in animal (Baxter et al., 2000; Milad & Quirk, 2002) and human samples (Delgado et al., 2006; Etkin et al., 2006; Haas et al., 2007; Johnstone et al., 2007; Urry et al., 2006), implicating an inverse relationship between these structures that govern affective output. In particular, increased response in the vPFC is inversely correlated with responding in the amygdala, and predicts behavioral outcomes such as fear extinction (Gottfried & Dolan, 2004; Phelps et al., 2004), downregulation of autonomic responses (Phelps et al., 2004) and more positive interpretations of emotionally ambiguous information (Kim, Somerville et al., 2003; 2004). Therefore, it is not surprising that the particular circuitry observed to show an ‘imbalance’ in adolescents and giving rise to heightened emotional behavior is that of the amygdala and vPFC.
Environmental Factors
A number of studies have shown the significance of environmental factors such as stress and early adversity on brain and behavior (Liston et al., 2006; 2009; Tottenham et al., in press) and risk for psychopathology (Breslau et al., 1998; Kessler, Sonnega, Bromet, Hughes, & Nelson, 1995). Trauma exposure is a particularly potent environmental risk factor for anxiety and depression (Brown, 1993; Kendler, Hettema, Butera, Gardner, & Prescott, 2003; McCauley, Kern, Kolodner, Dill, & Schroeder, 1997). A recent study by our group examined the effects of a naturally occurring disaster on affective processing of cues of threat. Specifically, we used functional magnetic resonance imaging to assess the impact of proximity to the disaster of September 11, 2001, on amygdala function in 22 healthy young adults.
Our findings suggest that more than three years after the terrorist attacks, bilateral amygdale activity in response to viewing fearful faces compared to calm ones was higher in individuals who were within 1.5 miles of the World Trade Center on 9/11, relative to those who were living more than 200 miles away (all were living in the New York metropolitan area at time of scan). This effect was statistically driven by time since worst trauma in lifetime and intensity of worst trauma, as indicated by reported symptoms at time of the trauma (see Figure 4). These data are consistent with a model of heightened amygdala reactivity following high-intensity trauma exposure, with relatively slow recovery.
Figure 4. Amygdala activity and proximity to the WTC on September 11th, 2001.
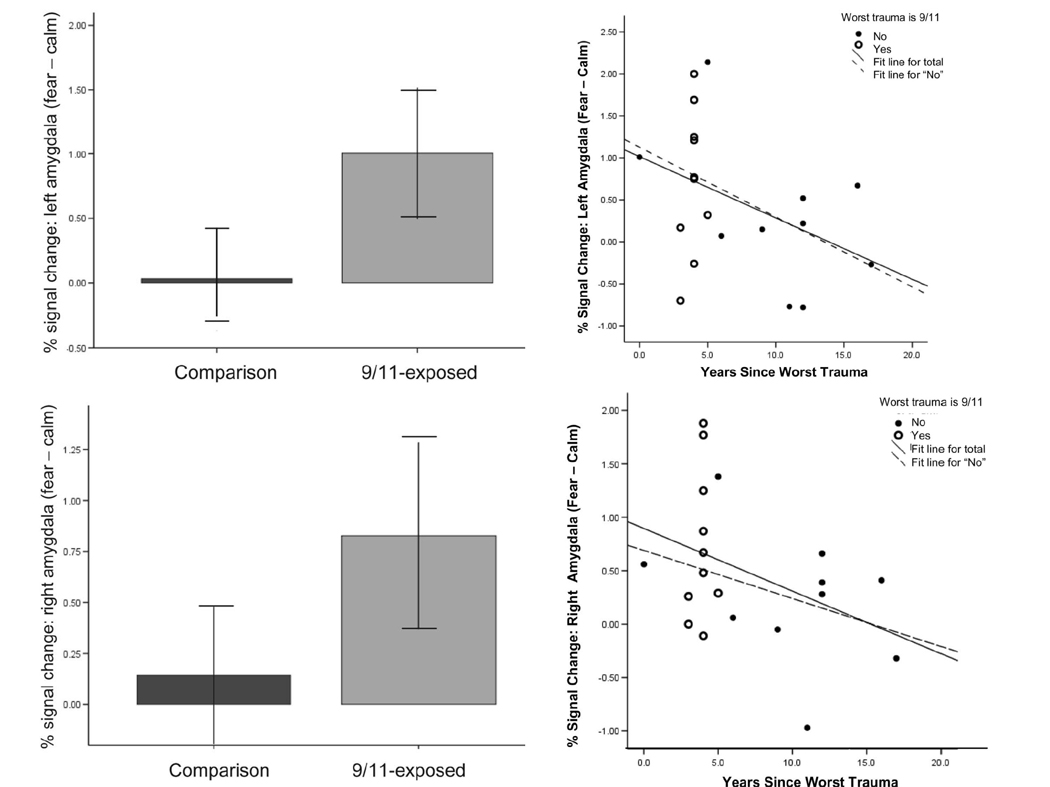
Fearful emotional faces elicited greater left (top panel) and right (bottom panel) amygdale activity in the 9/11-exposed group relative to a comparison group (p < 05). Amygdala activity was negatively correlated with time since worst trauma in lifetime in left (r = .46, p < .05 and right amygdale ((r = .45, p < .06). Adapted from Ganzel et al. (2007).
In the context of our model of adolescence, individuals who experience trauma during this period or have experienced multiple traumas may be especially vulnerable for developing symptoms of anxiety and depression as teens. In other words, while heightened emotional reactivity is typical during the period of adolescence, failure to suppress that emotional reactivity is associated with symptoms of anxiety. The large variability observed in our developmental studies of emotion regulation may in part be due to variation in individuals’ experiences. An imbalance in amygdala-PFC coupling has been implicated in the pathophysiology of psychiatric illnesses (mood and anxiety disorders) in adult (Blair et al., 2008; Drevets, 2003; Johnstone et al., 2007); and developing populations (Guyer et al., 2009; McClure et al., 2007; Monk et al., 2008; Rich et al., 2006; Pine, 2007) showing greater amygdala relative to prefrontal activity. As such, improving our understanding of the development of these circuits and the source of biased responding in some adolescents over others will facilitate our understanding of the most commonly experienced psychiatric illnesses of this developmental period (i.e., anxiety and depression).
Genetic Factors
A number of human genetic studies have begun to identify candidate genes that may play a role in increased risk for anxiety and depression. The main avenues for understanding gene function in these disorders have been in behavioral genetics on one end and on the other end, molecular mouse model. Attempts to bridge these approaches have used brain imaging to conveniently link anatomical abnormalities seen in knockout/transgenic mouse models and abnormal patterns of brain activity seen in humans. Recently we completed a study using human and mouse behavioral genetic together with imaging genetics. Each of these approaches alone, provide limited information on gene function in complex human behavior such as emotion regulation and its dysregulation in psychopathology, but together, they are forming bridges between animal models and human psychiatric disorders.
In this study, we utilized a common single nucleotide polymorphism (SNP) in the brain-derived neurotrophic factor (BDNF) gene that leads to a valine (Val) to methionine (Met) substitution at codon 66 (Val66Met). In an inbred genetic knock-in mouse strain that expresses the variant BDNF allele to recapitulate the specific phenotypic properties of the human polymorphism in vivo, we found the BDNF Val66Met genotype was associated with treatment resistant forms of anxiety-like behavior (Chen et al. 2006). A key feature of anxiety is impaired learning of cues that signal safety versus threat and unlearning of cues that signal threat when the association no longer exists (i.e., extinction). Thus, the objective of our study was to test if the Val66Met genotype could impact extinction learning in our mouse model, and if such findings could be generalized to human populations.
We examined the impact of the variant BDNF on fear conditioning and extinction paradigms (Soliman et al 2010). Approximately 70 mice and 70 humans were tested. The mice include 17 BDNFVal/Val, 33 BDNFVal/Met and 18 BDNFMet/Met. The human sample included 36 Met allele carriers (31 BDNFVal/Met and 5 BDNFMet/Met) and 36 nonMet allele carriers group-matched on age, gender and ethnic background. Fear conditioning consisted of pairing a neutral cue with an unconditioned aversive stimulus until the cue itself took on properties of the unconditioned stimulus (US) of an impending aversive event. The extinction procedure consisted of repeated presentations of the cue (i.e., conditioned stimulus or CS) alone. Behavioral responses of percentage of time freezing in the mouse and amplitude of the galvanic skin response in the human were the dependent measures. In addition, we collected brain imaging data using fMRI with the human sample.
Our findings showed no effects of BDNF genotype on fear conditioning in the mice or humans as measured by freezing behavior to the conditioned stimulus in the mice (F(2,65) = 1.58, p < 0.22) and by skin conductance response in humans to the cue predicting the aversive stimulus relative to a neutral cue (F(1,70) = 0.67, p < 0.42). However, both the mice and humans showed slower extinction in Met allele carriers than in nonMet allele carriers as shown in Figure 5A and B below. Moreover, human functional magnetic resonance imaging data provide neuroanatomical validation of the cross-species translation. Specifically, we show alterations in frontoamygdala circuitry, shown to support fear conditioning and extinction in previous rodent (Myers & Davis, 2002; Quirk et al., 2003;Milad & Quirk, 2002;LeDoux, 2000) and human (LaBar et al., 1998; Schiller et al., 2008; Delgado et al., 2008; Kalischet al., 2006; Gottfried & Dolan, 2004; Phelps et al., 2004) studies, as a function of BDNF genotype. Met allele carriers show less ventromedial prefrontal cortical (vmPFC) activity during extinction relative to nonMet allele carriers (Figure 5C), but greater amygdala activity relative to nonMet allele carriers (Figure 5D). These findings suggest that cortical regions essential for extinction in animals and humans (Quirk et al., 2003; Gottfried & Dolan, 2004; Lebron et al., 2004) are less responsive in Met allele carriers during extinction. Moreover, amygdala recruitment, that should show diminished activity during the extinction (Phelps et al 2004) remains elevated in Met allele carriers.
Figure 5. Altered behavior and neural circuitry underlying extinction in mice and humans with BDNF Val66Met.

Impaired extinction in Met allele carriers (Val/Met and Met/Met) as a function of time in 68 mice (A) and 72 humans (B) as indexed by percent time freezing in mice and skin conductance response (SCR) in humans to the conditioned stimulus when it was no longer paired with the aversive stimulus. (C) Brain activity as indexed by percent change in MR signal during extinction in the ventromedial prefrontal cortex (vmPFC) by genotype (xyz = −4, 24, 3), with Met allele carriers having significantly less activity than Val/Val homozygotes [VM < VV = blue], image threshold p < 0.05, corrected. (D) Genotypic differences in left amygdala activity during extinction (xyz = −25, 2, −20) in 70 humans, with Met allele carriers having significantly greater activity than Val/Val homozygotes [VM > VV = orange], image threshold p < 0.05, corrected. *p < 0.05. **MM were included in the analysis with VM, but plotted separately to see dose response. All results are presented as a mean ± SEM. VV = Val/Val; VM = Val/Met; MM = Met/Met (From Soliman et al. 2010).
These findings are provocative as they provide an example of bridging human behavioral and imaging genetics with a molecular mouse model to suggest a role for BDNF in anxiety disorders. Moreover, these data suggest impaired learning of cues that signal safety versus threat, and in the efficacy of treatments that rely on extinction mechanisms such as exposure therapy. In the context of our model of adolescence, individuals with the BDNF Met allele may be more vulnerable for developing symptoms of anxiety and depression as teens, in that they show less vPFC activity and greater amygdala activity to repeated exposure to empty threat. These genetic data provide an example then of how an imbalance in amygdala-PFC coupling during typical development could be exacerbated and lead to clinical symptoms of anxiety.
Conclusions
Taken together, the findings synthesized here indicate that increased risk in adolescence for storm and stress is associated with different developmental trajectories of subcortical emotional systems and cortical control regions. This differential development may lead to an imbalance in control by subcortical regions over prefrontal ones and heightened emotional reactivity. Although elevated emotional reactivity is typical during the period of adolescence, failure to suppress that emotional reactivity with time is associated with symptoms of anxiety. The large variability observed in our developmental studies of emotion regulation may in part be due to variation in individuals’ environmental and genetic background. Both environmental and genetic factors can exacerbate the imbalance between limbic and control regions and in turn lead to greater storm and stress in some individuals over others. Together, our studies provide a converging methods approach for understanding the highly variable stress and turmoil experienced in adolescence (see potential promises and challenges of the application of genetic imaging to developmental research are described by Casey, Soliman, Bath and Glatt, 2010). Important future directions will be to consider the interaction among genetic, environmental and developmental factors in sufficiently large samples to directly test theses effects from a developmental perspective.
Figure 3. Habituation of Amygdala Response is associated with Anxiety and Less vPFC activity.

Trait anxiety scores were negatively correlated with habituation (decrease from early to late trials) of amygdale activity (r=.447; p .001). Amygdala habituation was calculated by subtracting activity in late trials from activity in early trials. (A) Region of the left amygdala that correlated with trait anxiety. (B) Scatter plot of the correlation between trait anxiety and amygdala habituation. The y-axis represents MR signal in the left amygdala for early_late trials. The x-axis represents trait anxiety score. There was negative functional coupling between the amygdala and the ventral prefrontal cortex (vPFC). The magnitude of activity in vPFC and the strength of the connectivity between vPFC and the amygdala were negatively correlated with amygdala habituation (r =.559 p .001). (Right) Scatterplot of vPFC-amygdala connectivity values versus amygdala habituation. The y-axis represents MR signal in the left amygdala for early-late trials. The x-axis represents Z-scored vPFC-amygdala connectivity values. Adapted from Hare et al. 2008.
Acknowledgments
This work was supported in part by NIMH 1R01 MH73175 NIDA R01 DA018879, NIMH P50 MH62196, the Mortmer D. Sackler family, the Dewitt-Wallace fund, and by the Weill Cornell Medical College Citigroup Biomedical Imaging Center and Imaging Core.
References
- Abe K, Suzuki T. Prevalence of some symptoms in adolescence and maturity: Social phobias, anxiety symptoms, episodic illusions and ideas of reference. Psychopathology. 1986;19:200–205. doi: 10.1159/000284448. [DOI] [PubMed] [Google Scholar]
- Baird AA, Gruber SA, Fein DA, Maas LC, Steingard RJ, Renshaw PF, Cohen BM, Yurgelun-Todd DA. Functional magnetic resonance imaging of facial affect recognition in children and adolescents. Journal of American Academy of Child and Adolescent Psychiatry. 1999;38:195–199. doi: 10.1097/00004583-199902000-00019. [DOI] [PubMed] [Google Scholar]
- Baxter MG, Parker A, Lindner CC, Izquierdo AD, Murray EA. Control of response selection by reinforcer value requires interaction of amygdala and orbital prefrontal cortex. Journal of Neuroscience. 2000;20:4311–4319. doi: 10.1523/JNEUROSCI.20-11-04311.2000. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Blakemore S-J. The social brain in adolescence. Nature Reviews Neuroscience. 2008;9:267–277. doi: 10.1038/nrn2353. [DOI] [PubMed] [Google Scholar]
- Blair K, Shaywitz J, Smith BW, Rhodes R, Geraci M, Jones M, McCaffrey D, Vythilingam M, Finger E, Mondillo K, Jacobs M, Charney DS, Blair RJ, Drevets WC, Pine DS. Response to emotional expressions in generalized social phobia and generalized anxiety disorder: evidence for separate disorders. American Journal of Psychiatry. 2008;165:1193–1202. doi: 10.1176/appi.ajp.2008.07071060. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bourgeois JP, Goldman-Rakic PS, Rakic P. Synaptogenesis in the prefrontal cortex of rhesus monkeys. Cerebral Cortex. 1994;4:78–96. doi: 10.1093/cercor/4.1.78. [DOI] [PubMed] [Google Scholar]
- Breslau N, Kessler RC, Chilcoat HD, Schultz LR, Davis GC, Andreski P. Trauma and posttraumatic stress disorder in the community: the 1996 Detroit Area Survey of Trauma. Archives of General Psychiatry. 1998;55:626–632. doi: 10.1001/archpsyc.55.7.626. [DOI] [PubMed] [Google Scholar]
- Brown TT, Lugar HM, Coalson RS, Miezin FM, Petersen SE, Schlaggar BL. Developmental changes in human cerebral functional organization for word generation. Cerebral Cortex. 2005;15:275–290. doi: 10.1093/cercor/bhh129. [DOI] [PubMed] [Google Scholar]
- Brown G. Life events and affective disorder: Replications and limitations. Psychosomatic Medicine. 1993;55:248–259. doi: 10.1097/00006842-199305000-00003. [DOI] [PubMed] [Google Scholar]
- Bunge SA, Dudukovic NM, Thomason ME, Vaidya CJ, Gabrieli JD. Immature frontal lobe contributions to cognitive control in children: evidence from fMRI. Neuron. 2002;33:301–311. doi: 10.1016/s0896-6273(01)00583-9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Casey BJ, Trainor RJ, Orendi JL, Schubert AB, Nystrom LE, Giedd JN, et al. A developmental functional MRI study of prefrontal activation during performance of a go-no-go task. Journal of Cognitive Neuroscience. 1997a;9:835–847. doi: 10.1162/jocn.1997.9.6.835. [DOI] [PubMed] [Google Scholar]
- Casey BJ, Castellanos FX, Giedd JN, Marsh WL, Hamburger SD, Schubert AB, et al. Implication of right frontostriatal circuitry in response inhibition and attention-deficit/hyperactivity disorder. J American Academy of Child & Adolescent Psychiatry. 1997b;36:374–383. doi: 10.1097/00004583-199703000-00016. [DOI] [PubMed] [Google Scholar]
- Casey BJ, Giedd JN, Thomas KM. Structural and functional brain development and its relation to cognitive development. Biological Psychology. 2000a;54:241–257. doi: 10.1016/s0301-0511(00)00058-2. [DOI] [PubMed] [Google Scholar]
- Casey BJ, Soliman F, Bath KG, Glatt CE. Imaging Genetics and Development: Possibilities and Challenges. Human Brain Mapping. doi: 10.1002/hbm.21047. in press. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Casey BJ, Thomas KM, Davidson MC, Kunz K, Franzen PL. Dissociating striatal and hippocampal function developmentally with a stimulus-response compatibility task. Journal of Neuroscience. 2002a;22:8647–8652. doi: 10.1523/JNEUROSCI.22-19-08647.2002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Casey BJ, Tottenham N, Fossella J. Clinical, imaging, lesion and genetic approaches toward a model of cogntitive control. Developmental Psychobiology. 2002b;40:237–254. doi: 10.1002/dev.10030. [DOI] [PubMed] [Google Scholar]
- Casey BJ, Getz S, Galvan A. The adolescent brain. Developmental Review. 2008a;28(1):62–77. doi: 10.1016/j.dr.2007.08.003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Casey BJ, Jones RM, Hare T. The adolescent brain. Annals of the New York Academy of Sciences. 2008b;1124:111–126. doi: 10.1196/annals.1440.010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chen ZY, Jing D, Bath KG, Ieraci A, Khan T, Siao CJ, Herrera DG, Toth M, Yang C, McEwen BS, Hempstead BL, Lee FS. Genetic variant BDNF (Val66Met) polymorphism alters anxiety-related behavior. Science. 2006;314:140–143. doi: 10.1126/science.1129663. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Compas BE, Orosan PG, Grant KE. Adolescent stress and coping: Implications for psychopathology during adolescence. Journal of Adolescence. 1993;16:331–349. doi: 10.1006/jado.1993.1028. [DOI] [PubMed] [Google Scholar]
- Crone E, Donohue S, Honomichl R, Wendelken C, Bunge S. Brain regions mediating flexible rule use during development. Journal of Neuroscience. 2006;26:11239–11247. doi: 10.1523/JNEUROSCI.2165-06.2006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Crone EA, van der Molen MW. Development of decision making in school-aged children and adolescents: evidence from heart rate and skin conductance analysis. Child Dev. 2007;78(4):1288–1301. doi: 10.1111/j.1467-8624.2007.01066.x. [DOI] [PubMed] [Google Scholar]
- Delgado MR, Olsson A, Phelps EA. Extending animal models of fear conditioning to humans. Biological Psychology. 2006;73:39–48. doi: 10.1016/j.biopsycho.2006.01.006. [DOI] [PubMed] [Google Scholar]
- Delgado MR, Nearing KI, LeDoux JE, Phelps EA. Neuron. 2008 Sep 1;59:829. doi: 10.1016/j.neuron.2008.06.029. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Drevets WC. Neuroimaging abnormalities in the amgydala in mood disorders. Annals of the NY Academy of Science. 2003;985:42044. doi: 10.1111/j.1749-6632.2003.tb07098.x. [DOI] [PubMed] [Google Scholar]
- Durston S, Davidson MC, Thomas KM, Worden MS, Tottenham N, Martinez A, et al. Parametric manipulation of conflict and response competition using rapid mixed-trial event-related fMRI. Neuroimage. 2003;20:2135–2141. doi: 10.1016/j.neuroimage.2003.08.004. [DOI] [PubMed] [Google Scholar]
- Durston S, Davidson MC, Tottenham N, Galvan A, Spicer J, Fossella J, et al. A shift from diffuse to focal cortical activity with development. Developmental Science. 2006;1:18–20. doi: 10.1111/j.1467-7687.2005.00454.x. [DOI] [PubMed] [Google Scholar]
- Eaton LK, Kann L, Kinchen S, Shanklin S, Ross J, Hawkins J, et al. Risk Behavior Surveillance - United States, 2007, surveillance summaries. Morbidity and Mortality Weekly Report. 2008;57(SS04):1–131. [PubMed] [Google Scholar]
- Ernst M, Nelson EE, Jazbec S, McClure EB, Monk CS, Leibenluft E, et al. Amygdala and nucleus accumbens in responses to receipt and omission of gains in adults and adolescents. Neuroimage. 2005;25:1279–1291. doi: 10.1016/j.neuroimage.2004.12.038. [DOI] [PubMed] [Google Scholar]
- Ernst M, Pine DS, Hardin M. Triadic model of the neurobiology of motivated behavior in adolescence. Psychol Med. 2006;36(3):299–312. doi: 10.1017/S0033291705005891. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Etkin A, Egner T, Peraza DM, Kandel ER, Hirsch J. Resolving emotional conflict: a role for the rostral anterior cingulated cortex in modulating activity in the amygdala. Neuron. 2006;51:871–882. doi: 10.1016/j.neuron.2006.07.029. [DOI] [PubMed] [Google Scholar]
- Galvan A, Hare TA, Parra CE, Penn J, Voss H, Glover G, et al. Earlier development of the accumbens relative to orbitofrontal cortex might underlie risk-taking behavior in adolescents. Journal of Neuroscience. 2006;26:6885–6892. doi: 10.1523/JNEUROSCI.1062-06.2006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ganzel B, Casey BJ, Glover G, Voss HU, Temple E. The aftermath of 9/11: effect of intensity and recency of trauma on outcome. Emotion. 2007;7:227–238. doi: 10.1037/1528-3542.7.2.227. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Giedd JN, Blumenthal J, Jeffries NO, Castellanos FX, Liu H, et al. Brain development during childhood and adolescence: A longitudinal MRI study. Nature Neuroscience. 1999;2:861–863. doi: 10.1038/13158. [DOI] [PubMed] [Google Scholar]
- Giedd JN, Snell JW, Lange. N, Rajapakse JC, Casey BJ, Kozuch PL, et al. Quantitative magnetic resonance imaging of human brain development: Ages 4–18. Cerebral Cortex. 1996;6:551–560. doi: 10.1093/cercor/6.4.551. [DOI] [PubMed] [Google Scholar]
- Gottfried JA, Dolan RJ. Human orbitofrontal cortex mediates extinction learning while accessing conditioned representations of value. Nature Neuroscience. 2004;7:1144–1152. doi: 10.1038/nn1314. [DOI] [PubMed] [Google Scholar]
- Guyer AE, Monk CS, McClure-Tone EB, Nelson EE, Roberson-Nay R, Adler A, et al. A developmental examination of amygdala response to facial expressions. Journal of Cognitive Neuroscience. 2008;20(9):1565–1582. doi: 10.1162/jocn.2008.20114. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Guyer AE, McClure-Tone EB, Shiffrin ND, Pine DS, Nelson EE. Probing the neural correlates of anticipated peer evaluation in adolescence. Child Development. 2009;80:1000–1015. doi: 10.1111/j.1467-8624.2009.01313.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Haas BW, Omura K, Constable RT, Canil T. Emotional conflict and neuroticism: personalitydependent activation in the amygdala and subgenual anterior cingulate. Behavioral Neuroscience. 2007;121:249–256. doi: 10.1037/0735-7044.121.2.249. [DOI] [PubMed] [Google Scholar]
- Hall GS. Adolescence: In psychology and its relation to physiology, anthropology, sociology, sex, crime, religion, and education (Vol. I & II) Englewood Cliffs, NJ: Prentice-Hall; 1904. [Google Scholar]
- Hare TA, Tottenham N, Galvan A, Voss HU, Glover GH, Casey BJ. Biological substrates of emotional reactivity and regulation in adolescence during an emotional go-nogo task. Biol Psychiatry. 2008;63(10):927–934. doi: 10.1016/j.biopsych.2008.03.015015. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hare TA, Tottenham N, Davidson MC, Glover GH, Casey BJ. Contributions of amygdala and striatal activity in emotion regulation. Biological Psychiatry. 2005;57:624–632. doi: 10.1016/j.biopsych.2004.12.038. [DOI] [PubMed] [Google Scholar]
- Huttenlocher PR. Synaptic density in human frontal cortex - developmental changes and effects of aging. Brain Research. 1979;163:195–205. doi: 10.1016/0006-8993(79)90349-4. [DOI] [PubMed] [Google Scholar]
- Huttenlocher PR. Regional differences in synaptogenesis in human cerebral cortex. Journal of Comparative Neurology. 1997;387:167–178. doi: 10.1002/(sici)1096-9861(19971020)387:2<167::aid-cne1>3.0.co;2-z. [DOI] [PubMed] [Google Scholar]
- Johnstone T, van Reekum CM, Urry HL, Kalin NH, Davidson RJ. Failure to regulate: Counterproductive recruitment of top-down prefrontal-subcortical circuitry in major depression. Journal of Neuroscience. 2007;27(33):8877–8884. doi: 10.1523/JNEUROSCI.2063-07.2007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kalisch R, et al. J Neurosci. 2006 Sep 13;26:9503. doi: 10.1523/JNEUROSCI.2021-06.2006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kendler KS, Hettema JM, Butera F, Gardner CO, Prescott CA. Life event dimension of loss, humiliation, entrapment, and danger in the prediction of onsets of major depression and generalized anxiety. Archives of General Psychiatry. 2003;60:789–796. doi: 10.1001/archpsyc.60.8.789. [DOI] [PubMed] [Google Scholar]
- Kessler RC, Berglund P, Delmer O, Jin R, Merikangas KR, Walters EE. Lifetime prevalence and age-of-onset distributions of DSM-IV disorders in the National Comorbidity Survey Replication. Archives of General Psychiatry. 2005;62:593–602. doi: 10.1001/archpsyc.62.6.593. [DOI] [PubMed] [Google Scholar]
- Kessler RC, Sonnega A, Bromet E, Hughes M, Nelson CB. Posttraumatic stress disorder in the National Comborbidity Survey. Archives of General Psychiatry. 1995;52:1048–1060. doi: 10.1001/archpsyc.1995.03950240066012. [DOI] [PubMed] [Google Scholar]
- Killgore WD, Yurgelun-Todd DA. Social anxiety predicts amygdala activation in adolescents viewing fearful faces. Neuroreport. 2005;16:1671–1675. doi: 10.1097/01.wnr.0000180143.99267.bd. [DOI] [PubMed] [Google Scholar]
- Kim H, Somerville LH, Johnstone T, Polis S, Alexander AL, Shin LM, Whalen PJ. Contextual modulation of amygdala responsivity to surprised faces. Journal of Cognitive Neuroscience. 2004;16:1730–1745. doi: 10.1162/0898929042947865. [DOI] [PubMed] [Google Scholar]
- Kim H, Somerville LH, Jonhstone T, Alexander AL, Whalen PJ. Inverse amygdala and medial prefrontal cortex responses to surprised faces. Neuroreport. 2003;14:2317–2322. doi: 10.1097/00001756-200312190-00006. [DOI] [PubMed] [Google Scholar]
- LaBar KS, Gatenby JC, Gore JC, LeDoux JE. Phelps, Neuron E. A. 1998 May;20:937. doi: 10.1016/s0896-6273(00)80475-4. [DOI] [PubMed] [Google Scholar]
- Lebron K, Milad MR, Quirk GJ. Learn Mem. 2004 Sep–Oct;11:544. doi: 10.1101/lm.78604. [DOI] [PubMed] [Google Scholar]
- LeDoux JE. Annu Rev Neurosci. 2000;23:155. doi: 10.1146/annurev.neuro.23.1.155. [DOI] [PubMed] [Google Scholar]
- Liston C, McEwen BS, Casey BJ. Psychosocial stress reversibly disrupts prefrontal processing and attentional control. Proceedings of the National Academy of Science. 2009;106:912–917. doi: 10.1073/pnas.0807041106. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Luna B, Sweeney JA. The emergence of collaborative brain function: FMRI studies of the development of response inhibition. Annals of the New York Academy of Sciences. 2004;1021:296–309. doi: 10.1196/annals.1308.035. [DOI] [PubMed] [Google Scholar]
- Luna B, Thulborn KR, Munoz DP, Merriam EP, Garver KE, Minshew NJ, et al. Maturation of widely distributed brain function subserves cognitive development. Neuroimage. 2001;13:786–793. doi: 10.1006/nimg.2000.0743. [DOI] [PubMed] [Google Scholar]
- McCauley J, Kern DE, Kolodner K, Dill L, Schroeder AF, DeChant HK, Ryden J, Derogatis LR, Bass EB. Clinical characteristics of women with a history of childhood abuse: unhealed wounds. JAMA. 277:1362–1368. [PubMed] [Google Scholar]
- McClure EB, Monk CS, Nelson EE, Parrish JM, Adler A, Blair RJ, Fromm S, Charney DS, Leibenluft E, Ernst M, Pine DS. Abnormal attention modulation of fear circuit function in pediatric generalized anxiety disorder. Archives of General Psychiatry. 2007;64:97–106. doi: 10.1001/archpsyc.64.1.97. [DOI] [PubMed] [Google Scholar]
- Milad MR, Quirk GJ. Neurons in medial prefrontal cortex signal memory for fear extinction. Nature. 2002;420:70–74. doi: 10.1038/nature01138. [DOI] [PubMed] [Google Scholar]
- Monk CS, McClure EB, Nelson EE, Zarahn E, Bilder RM, Leibenluft E, et al. Adolescent immaturity in attention-related brain engagement to emotional facial expressions. Neuroimage. 2003;20:420–428. doi: 10.1016/s1053-8119(03)00355-0. [DOI] [PubMed] [Google Scholar]
- Monk CS, Telzer EH, Mogg K, Bradley BP, Mai X, Louro HM, Chen G, McClure-Tone EB, Ernst M, Pine DS. Amygdala and ventrolateral prefrontal cortex activation to masked angry faces in children and adolescents with generalized anxiety disorder. Archives of General Psychiatry. 2008;65:568–577. doi: 10.1001/archpsyc.65.5.568. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Moses P, Roe K, Buxton RB, Wong EC, Frank LR, Stiles J. Functional MRI of global and local processing in children. Neuroimage. 2002;16:415–424. doi: 10.1006/nimg.2002.1064. [DOI] [PubMed] [Google Scholar]
- Nelson EE, Leibenluft E, McClure EB, Pine DS. The social re-orientation of adolescence: A neuroscience perspective on the process and its relation to psychopathology. Psychological Medicine. 2005;35:163–174. doi: 10.1017/s0033291704003915. [DOI] [PubMed] [Google Scholar]
- Paus T, Keshavan M, Giedd JN. Why do many psychiatric disorders emerge during adolescence? Nature Reviews Neuroscience. 2008;9:947–957. doi: 10.1038/nrn2513. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Phelps EA, Delgado MR, Nearing KI, LeDoux JE. Extinction learning in humans: role of the amygdale and vmPFC. Neuron. 2004;43:897–905. doi: 10.1016/j.neuron.2004.08.042. [DOI] [PubMed] [Google Scholar]
- Pine DS, Cohen P, Brook JS. Emotional reactivity and risk for psychopathology among adolescents. CNS Spectrum. 2001;6(1):27–35. doi: 10.1017/s1092852900022860. [DOI] [PubMed] [Google Scholar]
- Pine DS. Research review: a neuroscience framework for pediatric anxiety disorders. Journal of Child Psychology and Psychiatry. 2007:631–648. doi: 10.1111/j.1469-7610.2007.01751.x. [DOI] [PubMed] [Google Scholar]
- Quirk GJ, Likhtik E, Pelletier JG, Pare D. J Neurosci. 2003 Sep 24;23:8800. doi: 10.1523/JNEUROSCI.23-25-08800.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rakic P, Bourgeois JP, Eckenhoff MF, Zecevic N, Goldman-Rakic PS. Concurrent overproduction of synapses in diverse regions of the primate cerebral cortex. Science. 1986;232:232–235. doi: 10.1126/science.3952506. [DOI] [PubMed] [Google Scholar]
- Rich BA, Vinton DT, Roberson-Nay R, Hommer RE, Berghorst LH, McClure EB, Fromm SJ, Pine DS, Leibenluft E. Limbic hyperactivation during processing of neutral facial expressions in children with bipolar disorder. PNAS. 2006;103:8900–8905. doi: 10.1073/pnas.0603246103. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rubia K, Overmeyer S, Taylor E, Brammer M, Williams SC, Simmons A, et al. Functional frontalisation with age: mapping neurodevelopmental trajectories with fMRI. Neuroscience and Biobehavioral Reviews. 2000;24:13–19. doi: 10.1016/s0149-7634(99)00055-x. [DOI] [PubMed] [Google Scholar]
- Schiller D, Levy I, Niv Y, LeDoux JE, Phelps EA. J Neurosci. 2008 Nov 5;28:11517. doi: 10.1523/JNEUROSCI.2265-08.2008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Schlaggar BL, Brown TT, Lugar HM, Visscher KM, Miezin FM, Petersen SE. Functional neuroanatomical differences between adults and school-age children in the processing of single words. Science. 2002;296:1476–1479. doi: 10.1126/science.1069464. [DOI] [PubMed] [Google Scholar]
- Silveri MM, Tzilos GK, Pimentel PJ, Yurgelun-Todd DA. Trajectories of adolescent emotional and cognitive development: effects of sex and risk for drug use. Annals of the New York Academy of Sciences. 2004;1021:363–370. doi: 10.1196/annals.1308.046. [DOI] [PubMed] [Google Scholar]
- Soliman F, Glatt CE, Bath KG, Levita L, Jones RM, Pattwell SS, Jing D, Tottenham N, Amso D, Sommerville L, et al. A genetic variant BDNF polymorphism alters extinction learning in both mouse and human. Science. doi: 10.1126/science.1181886. in press. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Somerville LH, Jones RM, Casey BJ. A time of change: Behavioral and neural correlates of adolescent sensitivity to appetitive and aversive environmental cues. Brain & Cognition. 2010;72:124–133. doi: 10.1016/j.bandc.2009.07.003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Spear LP. The adolescent brain and age-related behavioral manifestations. Neuroscience and Biobehavioral Reviews. 2000;24:417–463. doi: 10.1016/s0149-7634(00)00014-2. [DOI] [PubMed] [Google Scholar]
- Steinberg L. A social neuroscience perspective on adolescent risk-taking. Dev Rev. 2008;28:78–106. doi: 10.1016/j.dr.2007.08.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Steinberg L. Cognitive and affective development in adolescence. TICS. 2005;9(2):69–74. doi: 10.1016/j.tics.2004.12.005. [DOI] [PubMed] [Google Scholar]
- Tamm L, Menon V, Reiss AL. Maturation of brain function associated with response inhibition. Journal of the American Academy of Child and Adolescent Psychiatry. 2002;41:1231–1238. doi: 10.1097/00004583-200210000-00013. [DOI] [PubMed] [Google Scholar]
- Thomas KM, Hunt RH, Vizueta N, Sommer T, Durston S, Yang Y, et al. Evidence of developmental differences in implicit sequence learning: an FMRI study of children and adults. Journal of Cognitive Neuroscience. 2004;16:1339–1351. doi: 10.1162/0898929042304688. [DOI] [PubMed] [Google Scholar]
- Thomas KM, Drevets WC, Dahl RE, Ryan ND, Birmaher B, Eccard CH, Axelson D, Whalen PJ, Casey BJ. Amygdala response to fearful faces in anxious and depressed children. Archives of General Psychiatry. 2001;58:1057–1063. doi: 10.1001/archpsyc.58.11.1057. [DOI] [PubMed] [Google Scholar]
- Tottenham N, Hare TA, Quinn BT, McCarry TW, Nurse M, Gilhooly T, Millner A, Galvan A, Davidson MC, Eigsti IM, Thomas KM, Freed PJ, Booma ES, Gunnar MR, Altemus M, Aronson J, Casey BJ. Prolonged institutional rearing is associated with atypically large amygdala volume and difficulties in emotion regulation. Developmental Science. 13:46–61. doi: 10.1111/j.1467-7687.2009.00852.x. in press. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Turkeltaub PE, Gareau L, Flowers DL, Zeffiro TA, Eden GF. Development of neural mechanisms for reading. Nature Neuroscience. 2003;6:767–773. doi: 10.1038/nn1065. [DOI] [PubMed] [Google Scholar]
- Urry HL, van Reekum CM, Johnstone T, Kalin NH, Thurow ME, Schaefer HS, et al. Amygdala and ventromedial prefrontal cortex are inversely coupled during regulation of negative affect and predict diurnal pattern of cortisol secretion among older adults. Journal of Neuroscience. 2006;26(16):4415–4425. doi: 10.1523/JNEUROSCI.3215-05.2006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Williams LM, Brown KJ, Palmer D, Liddell BJ, Kemp AH, Olivieri G, et al. The mellow years? Neural basis of improving emotional stability with age. Journal of Neuroscience. 2006;26(24):6422–6430. doi: 10.1523/JNEUROSCI.0022-06.2006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yang TT, Menon V, Eliez S, Blasey C, White CD, Reid AJ, Gotlib IH, Reiss AL. Neuroreport. 2002;13:1737–1741. doi: 10.1097/00001756-200210070-00009. [DOI] [PubMed] [Google Scholar]
- Yurgelun-Todd D. Emotional and cognitive changes during adolescence. Current Opinion in Neurobiology. 2007;17:251–257. doi: 10.1016/j.conb.2007.03.009. [DOI] [PubMed] [Google Scholar]