

Abstract
Normal pregnancy is associated with significant hemodynamic changes and vasodilation in the uterine and systemic circulation in order to meet the metabolic demands of the mother and developing fetus. Hypertension in pregnancy (HTN-Preg) and preeclampsia (PE) are major complications and life-threatening conditions to both the mother and fetus. PE is precipitated by various genetic, dietary and environmental factors. Although the initiating events of PE are unclear, inadequate invasion of cytotrophoblasts into the uterine artery is thought to reduce uteroplacental perfusion pressure and lead to placental ischemia/hypoxia. Placental hypoxia induces the release of biologically active factors such as growth factor inhibitors, anti-angiogenic proteins, inflammatory cytokines, reactive oxygen species, hypoxia-inducible factors, and antibodies to vascular angiotensin II receptor. These bioactive factors affect the production/activity of various vascular mediators in the endothelium, smooth muscle and extracellular matrix, leading to severe vasoconstriction and HTN. As an endothelial cell disorder, PE is associated with decreased vasodilator mediators such as nitric oxide, prostacyclin and hyperpolarizing factor and increased vasoconstrictor mediators such as endothelin, angiotensin II and thromboxane A2. PE also involves enhanced mechanisms of vascular smooth muscle contraction including intracellular free Ca2+ concentration ([Ca2+]i), and [Ca2+]i sensitization pathways such as protein kinase C, Rho-kinase and mitogen-activated protein kinase. Changes in extracellular matrix composition and matrix metalloproteases activity also promote vascular remodeling and further vasoconstriction in the uterine and systemic circulation. Characterization of the predisposing risk factors, the biologically active factors, and the vascular mediators associated with PE holds the promise for early detection, and should help design specific genetic and pharmacological tools for the management of the vascular dysfunction associated with HTN-Preg.
Keywords: cytokines, endothelium, growth factors, placenta, preeclampsia, vascular smooth muscle
INTRODUCTION
Normal pregnancy (Norm-Preg) is associated with increased heart rate, plasma volume and cardiac output, as well as decreased vascular resistance and late gestational decrease in blood pressure (BP). These beneficial hemodynamic changes maintain sufficient blood supply and nutrients to the growing fetus, and are in most part due to increased amount/activity of endothelium-derived vasodilators such as nitric oxide.
In 5–10% of pregnancies, women develop one of four forms of hypertension in pregnancy (HTN-Preg): chronic HTN that predates pregnancy, preeclampsia (PE), chronic HTN with superimposed PE, and nonproteinuric gestational HTN [1,2]. In the Antenatal Care Trial by the World Health Organization (WHO), 9.1% of pregnant women had either PE or gestational HTN [3]. PE is characterized by HTN (BP ≥140/90 mmHg) and proteinuria (300 mg protein in urine sample). If untreated, PE can lead to eclampsia, a life-threatening maternal neurovascular complication with severe HTN and convulsions [4]. Although PE represents a major cause of maternal and fetal morbidity/mortality, the underlying mechanisms have not been clearly identified. Analysis of the plasma, body fluids, and postpartum placenta from PE women has helped in determining some of the biochemical factors involved. Also, studies in animal models of HTN-Preg have demonstrated prominent changes in the renal control mechanisms of BP and highlighted the role of glomerular injury and alteration in kidney function as major causes of proteinuria and HTN [5]. Alterations in the neurohumoral and vascular control mechanisms of BP as well as changes in lipid metabolism and blood coagulation have also been observed in PE and in animal models of HTN-Preg. Several genetic, dietary and environmental risk factors are thought to cause poor placentation, placental ischemia/hypoxia, and increased release of biologically active factors which eventually cause generalized vascular dysfunction and HTN-Preg (Fig. 1). This review discusses the genetic, medical and environmental risk factors, and outlines the various circulating factors and vascular mediators underlying the pathology of PE. The review also provides insight into potential approaches for the diagnosis and management of the vascular dysfunction associated with PE.
Fig. 1.
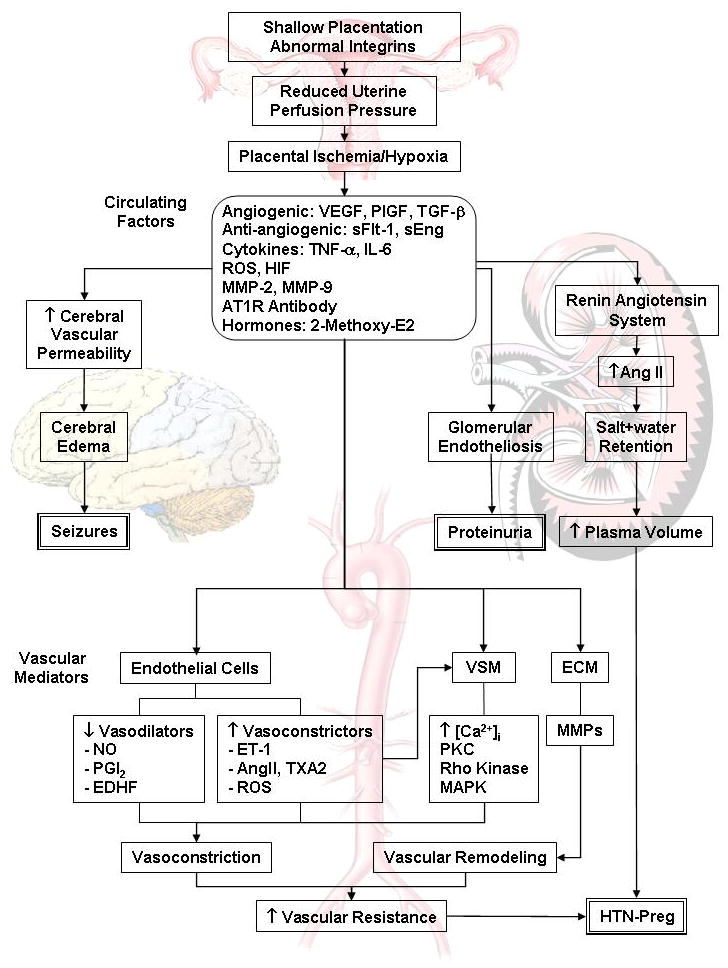
Cellular mechanisms of HTN-Preg. Shallow placentation and RUPP during late pregnancy trigger the release of biologically active factors, which affect the systemic blood vessels and lead to generalized vasoconstriction, increased vascular resistance and HTN-Preg. The bioactive factors could also injure the kidney leading to increased plasma volume and severe HTN, as well as glomerular endotheliosis and proteinuria, leading to clinically manifested PE. Increases in cerebral vascular permeability and edema lead to seizures and life-threatening eclampsia.
Genetic Risk Factors of PE
Maternal Genes
Female offspring of a pregnancy complicated by PE may have a higher risk of developing PE in their own pregnancies, suggesting potential inheritance of genes that enhance maternal susceptibility to PE [6]. Genome-wide scans and linkage studies have identified loci on chromosomes 1, 2p13, 2q, 3p, 4q, 9, 10q, 11q23-24, 12q, 15q, 18 and 22q, suggesting that PE is a multigenic disorder [7] (Table 1). Genes encoding sialic acid binding Ig-like lectin 6 (Siglec-6), a potential leptin receptor, as well as pappalysin-2 (PAPP-A2), a protease that cleaves insulin-like growth factor (IGF) binding proteins, are over-expressed in PE [8]. On the other hand, comparison of the genetic content of placental tissue from PE patients and matched control pregnancies found that a large number of the 36 major genes implicated in PE exhibit down-regulation as early as the first trimester. Also, 12 out of the 36 PE genes are involved in maternal-fetal immune dysregulation and/or altered decidualization, suggesting compromised placentation and impaired trophoblast invasion during the first trimester [9]. Some of the genes encoding different components of the renin-angiotensin system (RAS) such as the angiotensinogen (AGT) and angiotensin type 1 receptor (AT1R), as well as the endothelial nitric oxide synthase (NOS) gene have shown variations in HTN-Preg. For instance, the presence of the T allele in the M235T polymorphism in the AGT gene may serve as a link to high BP during pregnancy [10]. Also, the G894T polymorphism in the NOS3 gene has been linked to symptoms of the metabolic syndrome, a possible precursor to PE [11]. Interestingly, the presence of the T allele in the T677C polymorphism in the 5,10-methylenetetrahydrofolate reductase (MTHFR) (NADPH) gene, which is responsible for converting homocysteine to methionine, is more prevalent in PE subjects compared to controls. Mutations in the MTHFR gene can also lead to homocysteinuria, which is observed in PE and intrauterine growth retardation (IUGR) pregnancies [12]. Other candidate genes for PE have been screened in the Genetics of Pre-eclampsia Consortium (GOPEC) study [13].
Table 1.
Representative Maternal Genes Associated with PE
| Gene Symbol | Full Name | Gene Map Locus | Function | Polymorphism | MIM Number | Reference |
|---|---|---|---|---|---|---|
| AGT | Angiotensinogen | 1q42-q43 | Converted to Ang I then Ang II, which maintains blood pressure; Linked to pathogenesis of hypertension | M235T | 106150 | [10] |
| F5 | Coagulation factor V | 1q23 | Produces proteins for hemostasis; Provides resistance to activated protein C | R485K, M385T | 612309 | [14] |
| MTHFR | 5,10-methylenetetrahydrofolate reductase (NADPH) | 1p36.3 | Produces enzyme that converts amino acid homocysteine to methionine | T677C | 607093 | [15] |
| NOS3 | Nitric Oxide Synthase 3 | 7q36 | Synthesizes NO in endothelium and maintains vascular homeostasis | C786T, T894G (linked to symptoms of metabolic syndrome), 4 a/b VNTR | 163729 | [11] |
| PAPP-A2 | Pappalysin-2 Pregnancy-associated plasma protein A2 | 1q23-q25 | Encodes proteases that cleave insulin-like growth factor-binding proteins, resulting in local activation of IGF signaling pathways | [8, 16] | ||
|
PEE1 PEE2 PEE3 |
PE/eclampsia 1 PE/eclampsia 2 PE/eclampsia 3 |
2p13 2p25 9p13 |
Genome scans from Australia, Iceland and Finland suggest that these 3 loci may have significant linkage to PE; Exact function of gene still unknown | 189800 609402 609403 |
[17] | |
| Siglec-6 | Sialic acid binding Ig-like lectin 6 | 19q13.3 | Receptor for the protein leptin; Tempo of expression is related to progression of labor | 604405 | [8, 18] |
Paternal Genes
In addition to maternal genes, evidence suggests a paternal component in the genetic inheritance of PE. For instance, PE is common in hydatid-form mole-pregnancies in which all the fetal chromosomes originate from the father [12]. Also, men born from PE pregnancies are more likely to father a PE pregnancy. PE is also common in pregnancies where the father, and not the mother, was born from a PE-Preg [19].
Fetal Genes
Another factor in the development of HTN-Preg is fetal genes. PE is more common in women homozygous for the inhibitory killer Ig-like receptor (KIR) haplotypes (AA) than in women homozygous for the stimulator KIR halpotypes (BB), and the effect is strongest if the fetus is homozygous for the human leukocyte antigen class II (HLA-C2) haplotype [20]. Also, studies in mice have shown that the cyclin-dependent kinase inhibitor cdkn1c (p57Kip2) is a major regulator of embryonic growth, and mutations of this gene are associated with placental architecture and histological changes indicative of the reduced utero-placental blood flow found in HTN-Preg. Also, pregnant mice expressing wild-type levels of p57Kip2 develop a PE-like condition when carrying p57kip2-deficient pups [7], supporting a role of fetal genes in the development of HTN-Preg.
Ethnic Background
Ethnic background may also be a factor in the development of PE. A study of pregnant women in New York found that the incidence rates of PE are highest in African-American women, and lowest in Asian women [21]. Also, African-American and Hispanic women had higher rates of PE compared to Caucasian and Asian women. A more recent New York study indicated that the rates of PE per 100 hospitalizations for delivery were Caucasian 2.0, Hispanic 3.0, African American 3.3, and Other 2.3, suggesting racial discrepancies in the incidence of PE [22]. The differences in ethnic prevalence of PE may be related to the socioeconomic status, dietary traditions, and medical comorbidities.
Socioeconomic Background
The incidence of PE is higher among women of unfortunate socioeconomic status and level of education. The 2008 Generation R Study observed 3547 women from the Netherlands experiencing their first pregnancies and analyzed the way in which socioeconomic factors influenced the outcome of their pregnancies. Overall, pregnancies in women with lower education background showed an increased risk of PE compared with pregnancies in women with higher education. Lower education was associated with multiple pregnancies, family history of PE, financial difficulties, psychopathology, and physically demanding working conditions [23]. These socioeconomic conditions may also influence the quality of nutrition available to women, another important risk factor for PE.
Dietary Risk Factors
Calcium, Magnesium, Potassium
Increased calcium (Ca2+) intake has been associated with lower incidence of HTN-Preg. However, the beneficial effects of Ca2+ supplementation are not universally supported, possibly due to the differences in age and socioeconomic status among women participating in the studies [24]. In one study supporting the benefits of Ca2+ supplementation, subjects who ingested Ca2+ tablets 2 g/day had a 66.7% lower risk of PE and a 44.9% lower risk of premature delivery. However, in women who developed PE there was no significant reduction in the severity of the disease in the Ca2+ supplementation compared to placebo group [25]. Other studies suggest that increasing Ca2+ intake during pregnancy does not prevent PE, but may minimize the severity of the disease [26]. In the WHO randomized controlled clinical trials of Ca2+ supplementation during pregnancy, Ca2+ did not decrease the risk of PE, but did help in lowering the severity of the condition [27]. Studies in pregnant ewes also showed that restriction of Ca2+ intake is associated with decreased uterine blood flow, increased BP and elevated urine protein; all symptoms similar to those of PE [28]. Thus, Ca2+ intake may have some benefits against PE and should be carefully monitored in the diets of pregnant women. Magnesium (Mg2+) is another dietary mineral with potential neurovascular effects during pregnancy. Mg2+ is a cofactor of enzymes that regulate temperature and protein synthesis and maintain electrical potential in nerves and muscle membrane [29]. Observational studies based on medical records indicated that Mg2+ supplementation during pregnancy was associated with a reduced risk of PE. Also, increased maternal Mg2+ intake correlated with a reduced risk of IUGR and higher birth weight. Diets high in potassium and fiber are also associated with a reduced risk of HTN [30].
Vitamins
Vitamin D, a major regulator of Ca2+ absorption and metabolism, may exhibit dietary advantages in pregnant women. Vitamin D deficiency in early pregnancy is an independent risk factor for PE. Vitamin D levels are lower in PE compared to Norm-Preg women [31]. Also, vitamin D-deficient mice have an increase in renin expression and plasma AngII, which are common characteristics of HTN [32]. An imbalance between oxidants and antioxidants may also play a pathogenic role in PE. Vitamin E is antioxidant, and as with other antioxidants, the serum levels of vitamin E are lower in PE than Norm-Preg women. Also, studies have shown decreased plasma levels of Vitamin C in women with PE, and Vitamin C and E supplementation may decrease the incidence of PE in women at high risk [33]. Some studies have shown lower serum levels of vitamin A in PE than Norm-Preg women in the third trimester. However, because of its potential teratogenic effects, the effects of vitamin A supplementation have not been examined in clinical trials. Folic acid, a coenzyme required for the assembly of nucleic acids, is essential for placental growth. In a recent study, women who received folic acid supplementation had a 63% decreased risk for PE compared to women with no supplementation [34]. Decreased dietary intake of folic acid and B12 results in increased plasma levels of homocysteine, a condition associated with a high risk of heart and vascular disease, as well as the risk of development of PE and IUGR [35].
Other Environmental Risk Factors of HTN-Preg
Lead
Chronic environmental exposure to low levels of lead affects the electrical and mechanical properties of the heart, alters vascular smooth muscle (VSM) function and may promote the development of HTN. Studies have demonstrated a dose-dependent relationship between lead exposure and HTN-Preg, such that there is a 4% increase in the risk of HTN per 0.05 mg/m3 increase in the seasonal average lead level [36]. Some lead is taken by the bones, where it is stored and accumulated with age. Therefore, women with history of past lead exposure or chronic low level exposure may have a higher risk of HTN-Preg, even after a long period of no exposure [36, 37].
Smoking
Despite the well-known negative and teratogenic effects of cigarette smoking on the outcome of pregnancy, some studies suggest that women who are frequently exposed to cigarette smoke may have reduced risk of PE [23, 38]. The mechanism of this paradoxical effect is unclear, but may involve angiogenic growth factors. As described below, soluble fms-like tyrosine kinase-1 (sFlt-1) is an anti-angiogenic factor and an antagonist protein that binds vascular endothelial growth factor (VEGF) and placental growth factor (PlGF) [39]. In PE there is disproportionate increase in sFlt-1 and decrease in VEGF and PlGF. Cigarette smoking could ameliorate PE by decreasing sFlt-1 and possibly increasing PlGF levels. However, while maternal plasma sFlt-1 levels are lower in smokers than non-smokers, the differences are less pronounced in cases of PE [38]. Further testing on the relation between cigarette smoke and the risk of PE may help to provide insight into the pathogenesis of the disease.
Comorbidities and Risk of PE
Obesity
Obesity is a prevalent disorder in the United States. As of 2005, the WHO estimated that at least 400 million adults are considered obese and this number is expected to increase at least two-fold in the next decade [40]. Obesity is linked to complications of pregnancy including PE. Changes in lipid metabolism may play a role in the endothelial cell (EC) damage characteristic of PE. PE women have elevated plasma levels of triglycerides, increased serum levels of free fatty acids, and higher placental concentrations of cholesterol, phospholipids, and lipid peroxides [41]. Increased intake of sucrose and polyunsaturated fatty acids is associated with high risk of PE [1]. Polyunsaturated fat is easily oxidized and therefore enhances the oxidative environment that exists in PE.
Adipokines, specific hormones secreted in adipose tissue, play a role in obesity-related inflammation and may serve as biomarkers in PE [42,43]. Adiponectin is an adipokine with anti-inflammatory and anti-artherogenic properties that increases insulin sensitivity. Some studies have shown that adiponectin levels decrease with increasing body mass index (BMI) and linked this deficiency to the insulin resistance characteristic of PE [42]. Other studies showed no difference in adiponectin levels among women of differing BMIs [43], but greater levels in normal weight PE than Norm-preg women [42,43]. Only in severe PE did overweight women show a decrease in adiponectin levels compared to normal weight women [43]. These conflicting data suggested that there may be an initial increase in the release of adiponectin in PE women as an attempt to reduce excess fat accumulation in tissues. These elevated levels of adiponectin may suppress the expression of adhesion molecules in vascular ECs and cytokine production from macrophages in order to minimize the inflammation associated with PE. However, when the PE women are obese, this inflammation-minimizing mechanism may begin to fail and the adiponectin concentrations decrease. Increased levels of another adipokine, leptin, and increased insulin resistance in obese women may also contribute to the decrease in adiponectin levels and spur the onset of severe PE [43].
Ethnic differences in obesity may influence the incidence of PE. According to the Institute of Medicine’s guidelines for weight gain during pregnancy, African-American women are most likely to be overweight prior to pregnancy and to exceed the suggested weight gain during pregnancy. Caucasian women are most likely to gain the required weight during pregnancy, while Asian women are most likely to gain less than the required weight [44]. Thus, ethnicities most likely to gain weight during pregnancy correlate with those most at risk of developing PE, supporting the notion that weight gain during pregnancy increases the risk of PE.
Diabetes
Diabetes is associated with EC dysfunction and may increase the risk of PE. Also, metabolic syndrome is characterized by glucose intolerance, dyslipidemia, insulin resistance, diabetes, obesity and HTN. Patients with metabolic syndrome have endothelial dysfunction, and their blood vessels could be more sensitive to circulating factors released during HTN-Preg. Increased sugar consumption in pregnant women may lead to hyperglycemia, which in turn could inhibit endothelium-dependent vasorelaxation. In a recent study, PE was one of the leading outcomes when pregnant women received glucose supplements as part of glucose-tolerance testing [45]. Also, during Norm-Preg, women experience brief insulin resistance due to increased glycemia after a meal and exhibit symptoms of elevated inflammatory response such as increased plasma levels of plasminogen activator inhibitor-1, TNF-α and C-reactive protein. In gestational diabetes and PE these factors increase to a greater extent. Gestational diabetes is also associated with higher BMI and increased risk of PE [46].
Medical History
PE and HTN-Preg are also associated with renal or cardiac disease, PE in a previous pregnancy, urinary tract infection, extreme maternal age (<16 or >40 years old), and twin pregnancy [47]. Risk factors of HTN-Preg also include a previous neonatal macrosomia, history of reproductive tract surgery, and antepartum hemorrhage. Primiparity and a history of chronic respiratory conditions may also increase the risk of PE [3].
Abnormal Placentation as an Initiating Event of HTN-Preg
During the early stages of Norm-Preg, the cytotrophoblasts invade the uterine spiral arteries, progressively replacing ECs, medial elastic tissue, VSM, and neural tissue. The uterine spiral arteries progressively transform from small muscular arterioles to low resistance large capacitance vessels, and by the end of the second trimester, they become dilated tubes lined by cytotrophoblasts, which ensure sufficient blood supply and nutrition to the growing fetus. During PE, abnormal expression of the cell adhesion molecules integrins and apoptosis of cytotrophoblasts lead to shallow invasion of the uterine spiral arteries to only the superficial layers of the decidua [48]. The inadequate vascular remodeling of the uterine spiral arteries does not meet the fetal blood flow and nutrition demands, resulting in IUGR [49,50]. Placental ischemia may also induce the release of bioactive circulating factors that cause the harmful changes in the maternal circulation and EC function observed in PE. Animal models of HTN-Preg have been useful in understanding the role of placental ischemia as an initiating event of the elevated BP in PE. Although induction of uteroplacental ischemia during pregnancy has shown variable effects in different species, it is considered an important animal model of HTN-Preg. Reduction of uteroplacental perfusion pressure (RUPP) in late pregnant sheep, dogs, rabbits and rats results in a hypertensive state that resembles PE [51] (Table 2).
Table 2.
Representative Animal Models for Hypertension in Pregnancy
| Model and Species | Technique | Manifestations | Other Findings Relevant to PE | Ref | |||
|---|---|---|---|---|---|---|---|
| BP (mmHg) | Proteinuria (mg/24 hr) | Pup Weight (g) | Litter size (# of pups) | ||||
|
RUPP Sprague-Dawley Rat |
Two groups: Norm-Preg RUPP On gestation day 13, a silver clip (0.203mm ID) is placed around the abdominal aorta and another clip (0.1mm ID) is placed on the main uterine branches of the ovarian arteries. |
100±2 130±3 |
n/a | 2.68±0.12 2.14±0.17 |
12.7±0.5 7.7±1.3 |
RUPP rats demonstrate increased expression of antiangiogenic factors, such as sFlt-1 (Norm-Preg: ~70 pg/ml; RUPP ~675 pg/ml), and elevated plasma cytokines, such as TNF-α (Norm-Preg: ~8 pg/ml; RUPP: ~18 pg/ml). RUPP rats also demonstrate changes in vascular responses to vasoconstrictors and vasodilators. | [51] |
|
TNF-α, IL-6 Sprague-Dawley Rat |
Two Groups: Norm-Preg Cytokine-infused On gestation day 14, rats are infused IV with either TNF-α or IL-6 200 ng/kg/day for 5 days. Control rats are infused with normal 0.9% saline. | 96±3 123±3 |
15.6±2.4 88.2±9.6 |
10.4±0.6 2.6±0.2 |
10.4±0.6 7.3±0.5 |
Cytokine-infused pregnant rats demonstrate increased vascular responses to vasoconstrictors and inhibition of endothelium-dependent NO-cGMP vascular relaxation pathway. | [52] |
|
L-NAME Sprague-Dawley Rats |
Four groups: Virgin Virgin+L-NAME Pregnant Pregnant+L-NAME On gestation day 15, pregnant and age-matched virgin rats received L-NAME 4 mg/kg/day in drinking water. |
118±3 125±6 113±5 172±6 |
n/a | n/a | n/a | Reduction of NO production during late pregnancy is associated with increased contraction and [Ca2+]I in vascular smooth muscle and may lead to the increased vascular resistance associated with HTN-preg. | [53] |
|
2-ME Deficient Mouse |
Four groups: Comt+/+ Comt+/+ + RO41-0960 Comt−/− Comt −/− with 2-ME On gestation day 10, mice were injected subcutaneously daily with 10 ng of 2-ME (COMT−/− mice), with 25 mg/kg COMT inhibitor Ro41- 0960 (wild-type mice), or with placebo (olive oil). |
105 122 121 108 |
(~Albumin/creatine) 1.0 1.48 1.50 1.20 |
(~weights of placentae) 0.094 0.085 0.085 0.090 |
(~number of dead embryos) 1 - 2.5 1 |
Decreased levels of 2-ME are associated with symptoms of PE. 2-ME may have utility as a plasma and urine diagnostic marker of PE, and may also serve as a therapeutic supplement to prevent or treat PE. | [54] |
In all animal models, measurements are taken on day 19–20 of gestation. n/a, not available
Other animals models of HTN-Preg include the DOCA/saline pregnant rat model demonstrating significant changes in the rennin-angiotensin system and the vascular NO pathway [55], the BPH/5 mouse which demonstrates spontaneous late-gestational proteinuria and progressive glomerulosclerosis [56], and the transfer of activated th1 cells into Norm-Preg mice leading to glomerulonephritis and proteinuria [57].
Biologically Active Factors Involved in HTN-Preg
Placental hypoxia is thought to induce the release of various bioactive factors. Increased plasma and vascular tissue levels of these bioactive factors during pregnancy could cause EC and VSM dysfunction, severe vasoconstriction and PE in humans (Table 3) and HTN-Preg in rats (Table 4). Placental trophoblasts may be a potential source of the factors released into the maternal circulation during PE. The syncytiotrophoblast connecting the placenta to the mother’s circulation is composed of a multi-nucleated syncytium with an extensive microvillous brush. In Norm-Preg, the syncytiotrophoblast microvillous membranes (STBMs) appear normal and break off slightly due to necrosis. Although the release of these free-floating STBMs into the maternal circulation could alter the structure of ECs, they do not cause extensive damage during Norm-Preg. However, during PE the STBMs are much thinner and have an unusual club-like form, making them susceptible to more frequent breakage with more debilitating results [58]. A complex purified from STBMs has been shown to inhibit the proliferation of cultured human ECs [59]. Also, studies on human umbilical vein ECs co-cultured with STBM cells isolated from normal and PE placentas have suggested that endothelial cell-to-cell junctions are sensitive targets for STBM-derived factors, leading to increased EC permeability [60]. Other studies do not support the STBM deportation theory and have shown that intraluminal perfusion of isolated myometrial arteries of Norm-Preg women with STBM concentrations up to 100 times those reported in PE does not affect bradykinin-induced dilation or cause significant damage to the endothelium [61].
Table 3.
Plasma Levels and Effects of Bioactive Factors and Vascular Mediators during Normal Pregnancy (Norm-Preg) and Preeclampsia (PE) in Humans.
| Bioactive Factor | Norm-Preg | PE | Site of Action | Effect | Reference |
|---|---|---|---|---|---|
| VEGF (pg/mL) | 184 | 396 | Endothelium | Vasorelaxation | [62] |
| sFlt-1 (pg/mL) | 339.4 | 7247 | Endothelium | Anti VEGF | [63] |
| Endoglin (ng/mL) | 9.8 | 46.4 | Endothelium | Anti TGF-β | [64] |
| TNF-α (pg/mL) IL-6 (pg/mL) |
3.31 4.9±1.1 |
4.68 16.5±2.1 |
Endothelium Endothelium |
EC Dysfunction EC Dysfunction |
[65] [66] |
| ROS | ↑ | ↑↑ | Endothelium | EC Dysfunction | [67] |
| HIF | ↑ | ↑↑ | DNA | Gene Transcription | [68] |
| MMP-2 (ng/mL) MMP-9 (ng/mL) |
779±194 121±190.5 |
783±322.3 262.2±398.6 |
ECM, ECM |
Remodeling Remodeling |
[69] [69] |
| 2-ME | ↑ | ↓ | Estrogen receptor | Vasodilator | [54] |
| Vascular Mediator | |||||
| NO (plasma nitrate/nitrite) (μmol/L) | 13.0±4.3 | 18.1±6.2 ↑, --, ↓ |
VSM | Vasorelaxation | [70] |
| PGI2 | ↑ | ↓ | VSM | Vasorelaxation | [71] |
| EDHF | ↑ | ↓ | VSM | Vasorelaxation | [72] |
| ET-1 (pmol/L) | 12±1.0 | 22.6±2.0 | VSM Endothelium |
Vasoconstriction Vasorelaxation |
[73] |
| AngII (fmol/mL) | 30.2 | 53.3 | VSM | Vasoconstriction | [74] |
↑↑, Greatly increased; ↑, Increased; ↓, Decreased; --, No change, relative to nonpregnant.
Table 4.
Plasma, Vascular Levels and Effects of Bioactive Factors and Vascular Mediators during Norm-Preg and HTN-Preg in rats.
| Bioactive Factor | Norm-Preg | HTN-Preg | Site of Action | Effect | Reference |
|---|---|---|---|---|---|
| VEGF (pg/mL) | 830±33 | 594±34 | Endothelium | Vasorelaxation | [75] |
| sFlt-1 (pg/mL) | 82±26 | 660±270 | Endothelium | Anti VEGF | [51] |
| Endoglin (ng/mL) | 8 | 17 | Endothelium | Anti TGF-β | [76] |
| TNF-α (pg/mL) IL-6 (pg/mL) |
8±1 36.6±7 |
48±13 104.5±28.6 |
Endothelium Endothelium |
EC Dysfunction EC Dysfunction |
[77] [78] |
| ROS | ↑ | ↑↑ | Endothelium | EC Dysfunction | [51] |
| HIF | ↑ | ↑↑ | DNA | Gene Transcription | [51] |
| Vascular Mediator | |||||
| NO (urinary nitrate/nitrite) (μmol/24 hr) | 46.4±5.3 | 49.8±6.4 ↑, --, ↓ |
VSM | Vasorelaxation | [79] |
| PGI2 | ↑ | ↓ | VSM | Vasorelaxation | [80] |
| ET-1 (pmol/L) | 1.7 | 2.2 | VSM | Vasoconstriction, | [81] |
| AngII | ↑ | ↑ | VSM | Vasoconstriction | [82] |
| VSM [Ca2+]i - Basal (nM) - PHE (10−5M) (nM) - AngII (10−7M) (nM) |
63 ± 5 149 ±8 149 ±8 |
109 ± 8 234 ±1 225 +/− 9 |
Myofilaments Myofilaments Myofilaments |
Basal Tone Contraction Contraction |
[83] [83] [84] |
↑↑, Greatly increased; ↑, Increased; ↓, Decreased; --, No change, relative to nonpregnant.
Some of the bioactive factors suggested to be released during PE include the pro-angiogenic factors plasma VEGF, PlGF, and transforming growth factor β (TGFβ), and anti-angiogenic factors such as sFlt1 and soluble endoglin (sEng). Other potential factors in PE include cytokines, oxidative stress, hypoxia-inducible factors (HIFs), matrix metalloproteases (MMPs), sex hormones and antibodies to vascular AT1R (Fig. 2).
Fig. 2.
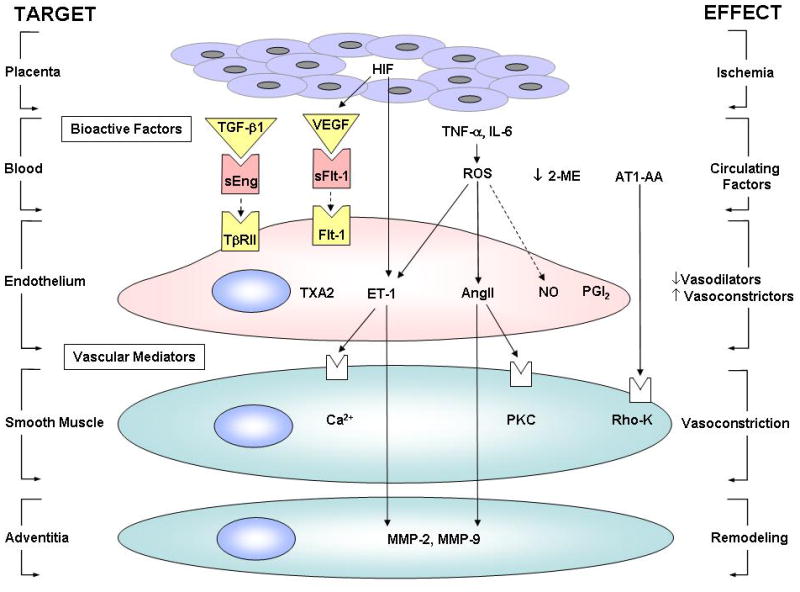
Interactions between biologically active factors and vascular mediators during HTN-Preg. Ischemia in the placenta triggers the release of circulating factors in the blood, followed by a decrease in vasodilation in the endothelium, an increase in vasoconstriction in vascular smooth muscle and vascular remodeling in the adventitia. sEng is a TGF-β antagonist that blocks the receptor TβRII, and sFlt-1 is a VEGF antagonist that blocks the Flt-1 receptor. Solid arrows indicate stimulation. Interrupted arrows indicate inhibition.
Vascular Endothelial Growth Factor (VEGF)
VEGF is a glycoprotein that acts as a mitogen and mediator of EC growth and tissue neovascularization. VEGF expression is induced by hypoxia in various cell types, including ECs and VSM cells. Studies suggest that serum VEGF immunoreactivity is suppressed during Norm-Preg [85]. In PE, elevated serum VEGF levels may be involved in EC activation/dysfunction [86]. Maternal plasma VEGF increases before the clinical onset of PE and is further elevated during the vasoconstriction phase of the disease. It is possible that the hyperdynamic circulation in the latent phase of PE may induce vascular shear stress, which in turn increases circulating VEGF. Because VEGF normally acts as a vasodilator, its increase may represent an unsuccessful vascular rescue response during P E [87]. VEGF may cause vasodilation by stimulating the NO-cyclic guanosine monophosphate (cGMP) vascular relaxation pathway. VEGF increases [Ca2+]i, which promotes calmodulin binding to eNOS, increases eNOS activity and stimulates NO production [88]. It is important to note that despite the increased serum levels of total VEGF, the levels of free-flowing VEGF may decrease in cases of PE compared to Norm-Preg [85]. In non-pregnant mice, infusion of antibodies against VEGF to cause a 50% reduction of VEGF leads to glomerular endotheliosis and proteinuria similar to what is seen in PE [39]. Also, patients receiving VEGF antagonists for cancer treatments develop HTN, proteinuria and endothelial activation [39]. Thus the decrease of the freely circulating VEGF, rather than the increased total VEGF, may be involved in the pathogensis of PE.
Placental Growth Factor (PlGF)
PlGF is another pro-angiogenic factor of the VEGF family that is associated with EC growth. Although the physiological role of PlGF is not completely understood, it may cause vasodilation in the peripheral vasculature and, thereby, modulates regional blood flow and vascular resistance [89]. During Norm-Preg, circulating PlGF concentrations are more than 40 times greater than VEGF, but PlGF has only 1/10th the affinity for the Flt-1 receptor compared to VEGF. During PE, circulating PlGF levels decrease while the levels of its antagonist protein, sFlt-1, increase [89]. Because the sFlt-/PlGF concentration ratio is much higher in PE than Norm-Preg women as early as the second trimester [90], it can be used as an early indicator of the onset of PE.
Soluble fms-like tyrosine kinase-1 (sFlt-1)
sFlt-1 is a VEGF and PlGF antagonist protein that binds VEGF and PlGF and prevents their interaction with their endogenous receptors (Fig. 2). Compared with Norm-Preg women, circulating levels of sFlt-1 are increased in PE women [75,91] secondary to placental ischemia/hypoxia [51]. The increase in sFlt-1 paired with a decrease in VEGF and PlGF promote proteinuria, HTN, and glomerular endotheliosis, the hallmarks of PE [39]. Administration of sFlt-1 to pregnant rats induces albuminurea, HTN, and renal pathologic changes in the form of glomerular endotheliosis [64]. Also, removal of sFlt-1 from the supernatant of PE tissue culture restores EC function and angiogenesis to normal levels [63]. Interestingly, Redman and coworkers suggested an association between trisomy 13 and PE and predicted the involvement of fetal factors and proteins encoded by genes located on chromosome 13, the same chromosome where the sFlt-1 gene is found [92,93].
Endoglin
Endoglin is a transmembrane receptor for transforming growth factor-β (TGF-β) that is expressed on ECs. Placenta-derived soluble endoglin (sEng) is an anti-angiogenic protein with a potential role in PE [94]. sEng inhibits TGF-β1 signaling in ECs and blocks TGF-β1 mediated activation of eNOS and vasodilation, suggesting that dysregulated TGF-β signaling is involved in the pathogenesis of PE [95] (Fig. 2). sEng is elevated in sera of PE women 2 to 3 months before the onset of clinical signs, correlates with the severity of PE, and falls after delivery. Thus, an increase in the serum level of sEng together with increased sFlt-1:PlGF ratio could be predictive of PE [96]. This is supported by reports that in pregnant rats, sEng increases vascular permeability, induces HTN [95] and potentiates the vascular effects of sFlt-1 to induce a severe PE-like state, with the development of IUGR and symptoms of HELLP syndrome including hemolysis, elevated liver enzymes and low platelet count [96]. However, the use of sFlt-1 and sEng as mediators or predictors of PE should be viewed with extreme caution as the levels of sFlt-1 and sEng may not be altered in some women or experimental animals with PE/HTN-Preg as compared to Norm-Preg.
Cytokines
Cytokines such as tumor necrosis factor-α (TNF-α), interleukin (IL)-6, and IL-8, are involved in cell proliferation and hypoxia-mediated cell activation. Hypoxia induces marked increases in IL-6 and IL-8 synthesis in VSM cells [78]. RUPP during pregnancy and the ensuing placental ischemia are thought to increase the release of TNF-α and IL-6 into the maternal circulation, leading to EC dysfunction, generalized vascular changes and HTN [97]. The plasma levels of TNF-α and IL-6 are elevated in women with PE [97]. In contrast, IL-10, which may be involved in the maintenance of pregnancy by inducing corpus luteum maturation and progesterone production, is decreased in PE women [98]. Cytokines may affect various vascular mediators (Fig. 2). TNF-α induces structural and functional alterations in ECs, enhances the formation of endothelin-1 (ET-1), reduces acetylcholine-induced vasodilatation and destabilizes eNOS mRNA [77]. Also, both TNF-α and IL-6 induce VEGF expression in ECs [99]. Infusion of cytokines has been used to develop animal models of HTN-Preg (Table 2). Chronic infusion of TNF-α or IL-6 in late pregnant rats results in marked elevation of renal vascular resistance and BP [52,100]. Also, vascular contraction is enhanced and endothelium-dependent vascular relaxation is reduced in TNF-α and IL-6 infused pregnant rats compared with Norm-Preg rats possibly due to inhibition of the endothelial NO-cGMP pathway [52].
Reactive Oxygen Species (ROS)
While some ROS may be normal and necessary during Norm-Preg, excessive ROS has been implicated in PE/HTN-Preg. Excessive oxidative stress could provide the link between localized RUPP and the generalized EC dysfunction in PE [101]. Hypoxia increases the production of superoxide anion (O2•−) and hydrogen peroxide in ECs and VSM cells [102]. During PE, free radicals and placental lipid peroxides are increased. Also, a deficiency in the plasma levels of the antioxidant ascorbate is typical of PE [103]. It has been shown that brachial artery flow-mediated and endothelium-dependent vasodilation is reduced in women who previously developed PE compared with controls. Administration of ascorbic acid increases flow-mediated vasodilation in women who previously developed PE but not in controls, suggesting that the EC dysfunction in women with previous PE is possibly due to increased oxidative stress and ROS [104]. However, whether antioxidants are beneficial in preventing the impaired EC function associated with PE needs to be further examined [105].
Neutrophils are another source of ROS and EC damage in PE. Stimulated neutrophils from women with PE produce more O2•− compared to neutrophils from Norm-Preg women. Neutrophils also produce NO, which can protect cells from O2•−-induced damage in Norm-Preg. However, in PE, O2•− production may dominate the neutrophil and EC NO production, resulting in EC injury [67]. Also, the plasma levels of lipid peroxidation markers is increased in women with PE, supporting the hypothesis that poor uteroplacental perfusion results in increased synthesis of placental free radical and maternal oxidative stress [106].
Recent research suggests an association between ROS and the severe endoplasmic reticulum (ER) stress initiated by deficient remodeling of the uterine arteries during PE. When there is a deficiency in the formation of maternal spiral arteries supplying the placenta, the arteries spontaneously constrict. If the arteries constrict enough, ischemia-reperfusion (I/R) injury will result. I/R injury leads to an increase in ROS, which decreases the amount of stored Ca2+ and intracellular ATP and produces misfolded proteins. These misfolded proteins create ER stress, which the cell attempts to fix via the unfolded protein response (UPR). During low-grade ER stress, the UPR promotes phosphorylation of eukaryotic initiation factor 2, which prevents the binding of the initiator to the ribosome and reduces cell proliferation. Although this is meant as a cell survival technique, prolonged exposure to UPR during pregnancy can be harmful to the mother and the fetus. If the ER stress is severe enough, the UPR may require the cells to undergo apoptosis. As a result, STBM particles enter the maternal circulation, impair vascular EC function, and lead to PE and IUGR [107].
O2•− and peroxynitrite may promote the vascular dysfunction in PE through increased degradation of HIF-1α [108] and stimulation of matrix metalloproteases MMP-1, -2, and -9 in VSM [109]. ROS can also increase VSM [Ca2+]i by stimulating the influx of extracellular Ca2+, inhibiting Ca2+-ATPase activity, or promoting inositol triphosphate (IP3)-induced Ca2+ release from intracellular stores. ROS also activate Ca2+-dependent protein kinase C (PKC) and promote vasoconstriction by activating Rho-A/Rho-kinase signaling mechanisms [110].
Hypoxia Inducible Factors (HIF)
Hypoxia promotes the induction of HIF-1α and HIF-1β, which are quickly degraded after reoxygenation. Once it is produced, HIF-1 binds to the enhancer region of genes that encode glycolytic enzymes and VEGF [111]. Because placental hypoxia likely plays a role in PE, the role of HIFs in the human placenta has been investigated. The expression of HIF-1α and HIF-2α, but not HIF-1β, is selectively increased in PE placenta [68]. The molecular mechanisms underlying the changes in HIF expression and the genes affected during PE are unclear. However, a HIF-1α binding site exists in the promoter region of the ET-1 gene and acts, in combination with other transcription factors, to regulate ET-1 expression [112]. Upregulation of HIF-1 also enhances the expression of VEGF [108].
Matrix Metalloproteases (MMP)
MMPs are zinc-dependent endopeptidases capable of degrading extracellular matrix proteins and processing a number of bioactive molecules. MMPs may play a role in the vascular dysfunction associated with HTN-preg. MMP-2, the main collagenolytic enzyme in the umbilical cord artery (UCA) wall, cleaves big ET-1 into ET-11-32, a potent vasodilator, and inhibits both relaxin and thrombin. Some studies suggest an increase in MMP-2 levels in PE [113]. Other studies have demonstrated reduced levels of MMP-2 in UCA and reduced collagen breakdown in the arterial wall, and suggested that the accumulation of collagen, along with reduction of elastin in UCA, decrease the blood flow to the fetus in women with PE [114]. Hypoxia increases MMP-9 mRNA and enzymatic activity 3-to 4-fold in ECs [111]. The higher MMP-9 activity in HTN-Preg compared with Norm-Preg suggests that MMP-9 may also play a role in the pathophysiology of PE. MMP may have a bidirectional relationship with VEGF, such that VEGF upregulates MMP expression, and membrane type-1 MMP promotes VEGF expression [115].
2-Methoxy-estradiol (2-ME) Deficiency
2-ME, a natural metabolite of E2, is elevated during the third trimester of Norm-Preg but is markedly decreased in women with severe PE (Fig. 2). Also, pregnant mice deficient in catechol-O-methyltransferase (COMT) demonstrate a decrease in 2-ME and show a PE-like phenotype. Administration of 2-ME ameliorated all PE-like features and suppressed placental hypoxia, HIF-1α expression and sFLT-1 elevation in the COMT deficient pregnant mice [54]. Thus, measurement of 2-ME in plasma and urine may be a useful biomarker for the diagnosis of PE. Also, because of its ability to reverse key abnormalities in PE, 2-ME supplementation may serve as a potential therapeutic measure [54].
Vascular Mediators in HTN-Preg
Vascular tone is maintained by a balance between vasodilator and vasoconstrictor mediators. The release of biologically active factors in response to placental hypoxia/ischemia could result in EC and VSM dysfunction and influence the release of various vascular mediators (Fig. 1). PE and other pregnancy-related complications, such as HELLP syndrome, may be associated with “generalized endotheliosis” and abnormal proliferation of ECs. The PE-associated glomerular endotheliosis in the kidney could result in renal injury. In PE, the ECs lining the glomerular capillaries are swollen and hypertrophied, leaving less space for erythrocytes and giving the glomeruli their bloodless appearance. Glomerular endotheliosis could be manifested as a decrease in both glomerular filtration rate (GFR) and renal plasma flow (RPF), resulting in increased blood urea nitrogen and creatinine levels similar to that of non-preganant women. The decrease in VEGF levels found in PE is partly responsible for glomerular endotheliosis. VEGF is produced in the glomerulus by podocytes and is essential for the preservation of general endothelial wellness, but also plays a key role in the initiation of fenestrae, the small pores in the ECs that allow the transfer of molecules from the glomerular capillaries into the urinary space. The lack of fenestrae contributes to the reduction of GFR during PE. At the dame time, subendothelial deposits of fibrin or fibrinogen-derivatives produces more filtrate in the extracellular space, which further reduces the GFR. The combination of a lack of fenestrae and the excessive production of sFlt-1 seen in women with PE inhibits VEGF signaling and eventually leads to the clinical complications of endotheliosis and proteinuria. Once the fetus is delivered, all glomerular changes accompanying endotheliosis are reversed within weeks as is the HTN and proteinuria [116,117]. Generalized endotheliosis in PE could also affect the systemic blood vessels leading to increased vascular resistance and HTN. More specifically, endotheliosis could affect the blood vessels of the brain leading to seizures or the blood vessels of the liver leading to HELLP syndrome. The EC dysfunction could also involve changes in the release of vasodilators, such as NO, prostacyclin (PGI2), and endothelium-derived hyperpolarizing factor (EDHF), or vasoconstrictor mediators, such as ET-1 and AngII in human (Table 3) and in rats (Table 4).
Decreased Endothelium Derived Vasodilators in PE
Nitric Oxide (NO)
NO is a potent vasodilator and relaxant of VSM. NO is produced from the transformation of L-arginine to L-citrulline by neuronal nNOS, inducible iNOS, and endothelial eNOS. Low oxygen tension in vivo results in a decrease in eNOS mRNA expression and NO production [111]. The vascular changes during Norm-Preg have been attributed, in part, to the increased NO synthesis by various maternal cells including vascular ECs [118]. NOS expression and activity are increased in human uterine artery during Norm-Preg [119]. Also, the plasma concentration and urinary excretion of cGMP, a second messenger of NO, are increased in Norm-Preg. The production of cGMP markedly increases during the first trimester when the maternal circulation is rapidly vasodilating, while NO production is not proportionately elevated, suggesting additional sources of cGMP [120].
Measurements of total NO production during HTN-Preg did not produce consistent findings. Some studies have shown that the concentration of nitrite/nitrate, stable byproducts of NO, is reduced in the sera of PE women [121], while other studies have shown increased levels [122]. The difference in the nitrite/nitrate levels during PE could be related to dietary nitrate intake. However, in a study of women with PE during which dietary nitrate/nitrite intake was controlled, reduction in NO production was not consistently observed [120]. The apparent dissociation between whole body NO production and the decreased vascular relaxation during HTN-Preg suggests tissue-specific differences in NOS expression and NO bioavailability. For instance, the expression of NOS isoforms may not be different in the placentas of Norm-Preg and PE women [123]. However, in late pregnant rats, the amount of renal eNOS is decreased by 39% while iNOS and nNOS increase by 31% and 25%, respectively [124].
Although the total nitrate/nitrite production may not be decreased in the plasma of PE women, the bioavailability of NO to produce vascular relaxation may be reduced. Ascorbate is essential for the decomposition of S-nitrosothiols and the release of NO. Ascorbate deficiency is typical of PE plasma and might result in decreased decomposition rates of S-nitrosothiols [33]. This is consistent with the observation that PE plasma contains higher concentrations of total S-nitrosothiols and S-nitrosoalbumin than plasma taken during Norm-Preg, possibly due to insufficient NO release from these major NO reservoirs in PE [103].
While measurements of total nitrate/nitrite did not produce consistent findings in PE, specific reduction in vascular endothelial NO production may be associated with PE [125]. In support of this theory, NOS blockade with Nω-nitro-L-arginine methyl ester (L-NAME) during mid to late gestation in rats produces pathological changes similar to those observed in women with PE including renal vasoconstriction, proteinuria, thrombocytopenia and IUGR [126,127]. Also, BP is markedly increased in pregnant rats treated with L-NAME compared with virgin rats treated with equal doses of L-NAME [127] (Table 2).
Prostacyclin (PGI2)
PGI2 is an anti-platelet aggregator and a vasodilator produced from the metabolism of arachidonic acid by the cyclooxygenase (COX)-1 and COX-2. Endothelium-derived PGI2 may contribute to the hemodynamic and vascular changes during pregnancy. During Norm-Preg, the plasma and urinary levels of 6-keto-prostaglandin F1α (PGF1α), a hydration product of PGI2, are increased [128]. Hypoxia has been shown to down-regulate COX-1 [129], and may result in the alterations in PGI2 production observed in PE. During severe PE, the plasma and urinary concentrations of PGF1α are lower than in Norm-Preg, suggesting that overall PGI2 synthesis is diminished. Endothelial PGI2 production may also be decreased in PE [130]. Interestingly, in vascular strips of RUPP rats, acetylcholine-induced relaxation is not completely inhibited by L-NAME, suggesting that changes in other endothelium-derived vasodilators such as PGI2 may play a role in HTN-preg [71].
Endothelium-Derived Hyperpolarizing Factor
EDHF is a major relaxing factor particularly in the small resistance vessels. Studies have shown that EDHF is the predominant relaxing factor in female mice, while NO and PGI2 are the dominant vasodilators in male mouse vessels [131]. An increase in EDHF production/activity may also play a role in the vascular adaptation during Norm-Preg. Studies on the uterine vascular beds of pregnant rats have suggested that EDHF release is activated by a delayed rectifier type of voltage-sensitive K+ channel [132] and causes vasodilation by affecting myoendothelial gap junctions alone or in combination with H2O2 or cytochrome P-450 epoxygenase, a metabolite of arachidonic acid. Although EDHF is not reduced by chronic hypoxia [133,134], its release may be impaired during PE [135,136].
While NO, PGI2 and EDHF have different vasodilator mechanisms, they may affect each other. PGI2 analogue may upregulate eNOS and NO production in ECs [137]. PGI2 may also activate the inward rectifier K+ channels through the release of EDHF [138].
Increased Endothelium Derived Vasoconstrictors in PE
Endothelin-1 (ET-1)
ET-1 is produced by ECs and causes marked vasoconstriction. Hypoxia induces the synthesis and secretion of ET-1 from ECs [112]. ET-1 production also appears to be increased in women with PE [139], perhaps due to activation of AT1R [73]. ET-1 secretion is 4- to 8-fold higher in umbilical cord ECs of PE compared to Norm-Preg women [140]. Also, the concentration of immunoreactive ET-1 is elevated in the plasma of PE women and rapidly returns to a normal value within 48 h of delivery, suggesting that ET-1 may contribute to the vasoconstriction associated with PE [141]. Typically, plasma ET-1 levels are highest during the later stages of PE, suggesting that ET-1 may not be involved in the initiation of PE, but rather in the progression into the malignant hypertensive phase of the disease [73].
ET-1 interacts with ETA receptor (ETAR) and ETB2R in VSM to cause vascular contraction. Under normoxic conditions, ET-1 increases VSM [Ca2+]i and contraction by stimulating ETAR-mediated Ca2+ release from the intracellular stores and PKC-dependent inhibition of K+ channels and subsequent membrane depolarization and Ca2+ influx through voltage-gated channels. In hypoxic conditions, up to 80% of ET-1 induced vascular contraction is independent of [Ca2+]i and PKC [142]. ET-1 also activates ETB1R in ECs and causes the release of NO, PGI2, and EDHF and promotes vascular relaxation [143]. ET-1 via ETBR may also mediate the reduced myogenic reactivity and vasodilation of renal arteries and the glomerular hyperfiltration during pregnancy in rats [144,145]. The role of ET-1 and its receptors in the vascular dysfunction associated with PE remains an important area of investigation.
Angiotensin II (AngII)
AngII is an important regulator of BP and electrolyte homeostasis. Hypoxia causes an increase in plasma levels of AngII in rabbits [146]. Also, local levels of ACE, AT1R and angiotensin type 2 receptor (AT2R) are increased during chronic hypoxia [146]. Plasma Ang-(1–7) levels and most other components of RAS, except ACE, are reduced in PE compared with Norm-Preg women. Only plasma AngII remains elevated in PE. In PE, the decreased plasma Ang-(1–7) and elevated AngII may contribute to HTN [147].
AngII causes vascular remodeling by promoting vasoconstriction, vascular growth, and inflammation. AngII also activates the Rho/Rho-kinase system, a prominent regulator of VSM contraction, the cytoskeleton, and vascular remodeling [148]. The response to AngII is decreased during Norm-Preg partly due to inactivation of AngII by angiotensinase in serum and placenta. Angiotensinase activity is lower in sera of women with severe PE leading to decreased degradation and increased sensitivity and vascular responses to AngII [149].
A fraction of circulating IgG antibodies may activate the AT1R and is considered a factor in PE (Fig. 2). The plasma level of AT1R autoantibodies (AT1-AA) rises in PE and subsides within 6 weeks after delivery. AT1-AA activity may promote Ca2+ signaling and VSM contraction. AT1-AA induced mobilization of intracellular Ca2+ also initiates a signaling cascade that leads to activation of NF-κB and activator protein-1 and increased tissue factor expression, which in turn cause vasoconstriction and lead to HTN [150].
Cytokines such as TNF-α and IL-6 stimulate AngII production in the female reproductive tract [151] and upregulate the expression of AT1R in VSM cells [152]. The vasoconstrictor effects of AngII are mediated via AT1R in VSM. AngII also induces the production of ET-1, which further promotes vasoconstriction [153]. AngII also stimulates AT2R in ECs and promotes the release of NO and PGI2 [154,155]. Although AngII infusion in mice does not change EDHF-mediated vascular relaxation [156], administration of an ACE inhibitor or an AT1R antagonist improves EDHF-mediated vascular relaxation in rats [157,158].
Thromboxane A2
TXA2, a potent vasoconstrictor and platelet aggregator, was thought to play a role in PE. The levels of TXB2, a metabolite of TXA2, are higher in PE than Norm-Preg women [159], and earlier studies suggested that the levels of TXA2 or its metabolites might be used to determine the severity of PE. However, experimental studies suggest that enhanced production of TXA2 may not play a major role in mediating the HTN and renal vasoconstriction produced by RUPP in pregnant rats [160]. Some clinical studies suggested the use of a low dose of aspirin to prevent PE [161]; however, a randomized controlled clinical trial on women at high risk for PE showed no benefit of low dose aspirin as a preventive measure [162].
Increased Vascular Smooth Muscle Mediators in PE
VSM Ca2+
[Ca2+]i is a major determinant of vascular tone. VSM activation is triggered by increases in [Ca2+]i due to IP3-induced Ca2+ release and ryanodine-sensitive Ca2+-induced Ca2+ release from the intracellular stores. Vasoconstrictor agonists also stimulate Ca2+ entry through ligand-gated and voltage-gated Ca2+ channels. Endothelium-derived relaxing factors act on VSM to inhibit phospholipase C, open K+ channels, or stimulate [Ca2+]i extrusion, and thereby decrease [Ca2+]i. EC dysfunction is associated with decreased release of relaxing factors and decreased VSM Ca2+ extrusion mechanisms. EC dysfunction also causes an increase in VSM contracting factors such as ET-1, which increases [Ca2+]i. VSM contraction in response to hypoxia is also mediated by increased [Ca2+]i [163]. It is generally thought that vasoconstrictors may play a role in PE. In rat aortic VSM cells, the basal and agonist-stimulated [Ca2+]i are reduced in Norm-Preg compared with virgin rats, but significantly elevated in pregnant rats treated with L-NAME [83] (Table 4). Also, AngII and KCl cause greater increases in VSM contraction and [Ca2+]i in renal VSM from RUPP compared with Norm-Preg rats [84]. However, studies on myometrial and subcutaneous resistance vessels have shown that microvessel reactivity to high KCl depolarizing solution, phenylephrine (PHE) and AngII are not increased in PE compared with Norm-Preg women, and suggested that this may be an unlikely mechanism of the increased vascular resistance in PE [164].
VSM Protein Kinase C (PKC)
PKC plays a role in VSM contraction. Activation of PKC by phorbol esters causes sustained contraction of VSM with no detectable change in [Ca2+]i, suggesting that PKC increases the Ca2+ sensitivity of the contractile proteins. In the uterine artery from pregnant sheep and the aorta of late pregnant rats there is a decreased contraction associated with a decrease in vascular PKC activity [165,166]. Also, the expression and subcellular redistribution of the Ca2+-dependent α-PKC and the Ca2+-independent δ- and ζ-PKC are reduced in aortic VSM of late pregnant compared with nonpregnant rats [166,167].
Increased VSM PKC expression/activity has been identified in HTN [168,169]. Also, the Ca2+ sensitivity of VSM contractile myofilaments is increased in women with PE, suggesting that VSM PKC activity may be involved in the vascular changes observed in PE [170]. PKC may also play a role in the changes in AngII and AT1R-mediated signaling associated with PE. Studies on cultured neonatal rat cardiomyocytes have shown that IgG in plasma from PE women enhances AT1R-mediated chronotropic response, and treatment of cardiomyocytes with the PKC inhibitor calphostin C prevented the stimulatory effect of PE IgG. Also, examination of VSM cells with confocal microscopy have shown colocalization of purified IgG from PE women and AT1R antibody. These studies concluded that PE women develop stimulatory AT1-AA, a process that appears to be mediated via PKC [171]. Also, we have shown that the expression and activity of α- and δ-PKC isoforms are enhanced in L-NAME treated compared with Norm-Preg rats, suggesting a role of α- and δ-PKC in the increased vasoconstriction and vascular resistance observed in HTN-Preg [166,167].
VSM Rho/Rho-Kinase
Rho is a family of small GTP-binding proteins that are involved in many cellular functions including cell proliferation, migration, cytoskeletal reorganization and contraction. Rho-kinase is activated by GTP binding and inactivated by hydrolyzing GTP to GDP, and this process is influenced by growth factors and vasoactive substances [172]. The Ca2+ sensitivity of the contractile elements is increased in subcutaneous resistance arteries from women with PE compared with Norm-Preg and nonpregnant women, suggesting a role of Rho-kinase in PE [170]. Also, an increase in AngII activity during PE may activate Rho-kinase and promote vasoconstriction [173]. However, other studies have shown that the expression of Rho-kinase mRNA may be downregulated in umbilical arteries of PE women [174], making it important to further investigate the role of Rho-kinase in the vascular changes associated with PE.
Diagnostic Elements and Biomarkers of PE
In addition to the hallmarks of PE, such as HTN and proteinuria, women with PE may develop edema in the face, hands, and feet, and experience sudden weight gain. The edema in PE may be related to the renal dysfunction and consequent sodium and fluid retention [117]. Other diagnostic elements and biomarkers of PE have been suggested.
Immune Response
The status of the immune system can be useful in predicting the risk of PE in early stages of pregnancy. The immune system fights off foreign pathogens through the activation of the alternate complement pathway, and activation of the Bb fragment could initiate vascular injury and fetal demise. PE women show higher levels of Bb than Norm-Preg women [175]. In support of this theory, the mother’s abnormal immune response to paternal antigens is prominent in PE. For example, PE is more common during a first conception, after the mother has switched partners, in women who use barrier contraception, and in cases of donated gametes. Conversely, conditions associated with suppressed immune response, such as HIV-related T-cell immune deficiency, are associated with a low rate of PE [20]. An increase in inflammatory factors in PE may also represent an abnormal maternal immunological response, comprising a change in the role of monocytes and natural killer cells, altered release of circulating cytokines, and activation of proinflammatory AT1R [20].
Platelet Count
The mean platelet volume (MPV) may be an important predictor of PE in pregnant women. Longitudinal studies have shown that women who develop PE have higher MPV than Norm-Preg women. Also, an increase in MPV can be detected as early as 4.6 weeks prior to the appearance of the hallmark symptoms of PE [176].
Uric Acid
The levels of uric acid could also be a potential predictor of PE. During Norm-Preg, uric acid levels decrease initially, but then gradually increase over gestational time. However, during PE the uric acid levels begin to increase as soon as 10 weeks earlier in the gestational period, and even precedes the MPV predictor. Also, rats with experimentally enhanced uric acid levels show higher incidence of renal and vascular disease. Elevated uric acid levels could contribute to the reduced production of NO and the altered endothelial function in PE women. Also, uric acid stimulates human monocytes to produce the pro-inflammatory cytokines TNF-α, IL-6 and IL-1β, which are elevated in PE [177]. Careful studies are needed to further evaluate the use of uric acid levels as a predictor of PE.
Amniotic Fluid
Because of the dangers to the fetus inherent from collection of the amniotic fluid, some studies utilized amniotic fluid from amniocyte banks for retrospective analysis. In one study, amniotic fluid samples were tested for insulin-like factor (INSL) 3, a peptide hormone made by the fetal testis and responsible for the first transabdominal phase of testicular descent. The results showed that, in male fetuses only, INSL3 levels were elevated in asymptomatic women who subsequently developed PE [178]. These results indicate that careful monitoring of hormones in the amniotic fluid may be used as a potential indicator of PE.
Fetal Development
Ultrasound technique can also monitor changes in the progress of fetal development and may detect IUGR early. For instance, ultrasounds have shown that fetuses with significant decrease in thymic size are prone to developing IUGR [179].
Circulating Factors
Other potential biomarkers, including the circulating factors described above, can also help in the prediction of PE. For example, the levels of VEGF, sFlt-1:PlGF, inflammatory cytokines, ROS and 2-ME can be monitored for early detection of PE (Table 3).
Uterine artery Doppler screening
Uterine artery Doppler screening at 23 weeks of pregnancy has been used to predict PE, with an abnormal finding defined as early diastolic bilateral uterine artery notching in the waveform. However, about 50% of mothers with uterine artery notching do not develop PE. Plasma levels of fibrinolytic markers including plasma levels of tissue-type plasminogen activator (t-PA), plasminogen activator inhibitor-1 (PAI-1), plasminogen activator inhibitor-2 (PAI-2), plasmin-α2-antiplasmin (PAP), D-dimers have also been used in conjunction with uterine artery Doppler screening as predictors of PE. Uterine artery notching at 23 weeks of gestation is associated with increased PAI-1 levels. Within the uterine artery notching group, mothers who developed PE had increased t-PA levels and decreased PAI-2 levels, although there was no net change in fibrinolysis as measured by D-dimer levels. These findings have led to the conclusion that no single fibrinolytic marker is helpful in determining pregnancy outcome in women with uterine artery notching, but t-PA and PAI-2 are worthy of study in a multifactorial algorithm [180].
Management of HTN-Preg
Overall, the current management of PE is mainly symptomatic, and the most effective way to reverse the manifestations is induction of delivery. Management of mild to moderate HTN-Preg consists of bed rest and anti-hypertensive drugs such as methyldopa, hydralazine, β-blockers and nifedipine, which have few side effects on the mother and fetus [181]. The antihypertensive drugs mainly decrease the risk of developing acute HTN, but may not have a clear effect on fetal mortality, premature birth, or fetal growth restriction. It is important to note that some of the contraindicated anti-hypertensive medications during PE include angiotensin receptor blockers (ARBs) and the angiotensin converting enzyme (ACE) inhibitors. If PE worsens into eclampsia, maintenance of airway patency and prevention of fluid aspiration is the first measure, then anticonvulsants such as magnesium sulfate infusion should be given. High doses of magnesium sulfate infusion may create side effects, such as increased postpartum bleeding, decreased neuromuscular transmission, increased respiratory depression, and maternal death. Lower dose intravenous magnesium sulfate regimens reduce these side-effects, while still working just as effectively as higher doses against eclampsia [182].
Potential Molecular Targets for Management of PE
Future approaches for management of HTN-Preg should focus on preventing the disease rather than treating the symptoms. Some of the potential approaches include methods to minimize placental hypoxia, utilize cytokine antagonists, and develop drugs that increase the release of vasodilators such as NO or block the release or the effects of vasoconstrictors such as ET-1. For example, recent trials have shown promising effects of supplements of L-arginine, a precursor of NO, in non-proteinuric gestational HTN [183], but little effects on the glomerular injury associated with PE [184]. Control of dietary and environmental factors including dietary calcium and vitamin D could also reduce the incidence of PE in susceptible women. Advanced gene analysis may also provide an alternative gene therapy approach and a specific measure to prevent PE.
Current research focuses on altering the levels of angiogenic factors in order to reverse the manifestations of PE. Experimental studies using recombinant VEGF-121 therapy on rats reversed PE-like phenotypes by replacing natural VEGF lost to sFlt-1 antagonism without causing any perceived effects on the fetus or placenta. VEGF-121 treatments also produced significant changes in gene expression by reversing the effects of genes related to angiogenesis, hypoxia, inflammation, and coagulation [185].
Research has also suggested possible management of PE by reducing the TNF-α inflammation of the endothelium with Digibind, a polyclonal-fragmented Digoxin-immune Fab antibody raised in sheep. The antibody decreases inflammation by down-regulating endothelial Na+/K+-ATPase activity, leading to improved maternal symptoms and increased fetoplacental perfusion [186].
Conclusion
Genetic predisposition, dietary and environmental factors affect the normal development of the placenta and may lead to placental ischemia/hypoxia and increase the release of bioactive growth factors, cytokines, ROS, HIF-1 and antibodies to AT1R. These bioactive factors in turn decrease the vasodilator mediators and increase the production of vasoconstrictor mediators leading to severe vasoconstriction, increased vascular resistance and HTN-Preg. Understanding the cellular mediators of PE should provide more specific and efficient measures for the prevention, diagnosis and treatment of this disorder.
Acknowledgments
This work was supported by grants from National Heart, Lung, and Blood Institute (HL-65998 and HL-70659) and The Eunice Kennedy Shriver National Institute of Child Health and Human Development (HD-60702).
List of Abbreviations
- AngII
angiotensin II
- BP
blood pressure
- [Ca2+]i
intracellular free Ca2+ concentration
- cGMP
cyclic guanosine monophosphate
- COX
cyclooxygenase
- EC
endothelial cell
- ECM
extracellular matrix
- EDHF
endothelium-derived hyperpolarizing factor
- ET-1
endothelin-1
- HIF
hypoxia-inducible transcription factor
- HTN-Preg
hypertension in pregnancy
- IL
interleukin
- L-NAME
Nω-nitro-L-arginine methyl ester
- MMP
matrix metalloproteinase
- 2-ME
2-methoxy-estradiol
- NO
nitric oxide
- Norm-Preg
normal pregnant
- NOS
NO synthase
- O2−•
superoxide anion
- PE
preeclampsia
- PGI2
prostacyclin
- PHE
phenylephrine
- PKC
protein kinase C
- PlGF
placental growth factor
- RAS
renin-angiotensin system
- ROS
reactive oxygen species
- RUPP
reduced uterine perfusion pressure
- sEng
soluble endoglin
- sFlt-1
soluble fms-like tyrosine kinase-1
- STBM
syncytiotrophoblast microvillous membranes
- TNF-α
tumor necrosis factor-α
- TXA2
thromboxane A2
- UCA
umbilical cord artery
- VEGF
vascular endothelial growth factor
- VSM
vascular smooth muscle
References
- 1.Clausen T, Slott M, Solvoll K, Drevon CA, Vollset SE, Henriksen T. High intake of energy, sucrose, and polyunsaturated fatty acids is associated with increased risk of preeclampsia. Am J Obstet Gynecol. 2001;185:451–458. doi: 10.1067/mob.2001.116687. [DOI] [PubMed] [Google Scholar]
- 2.Roberts JM, Gammill HS. Preeclampsia: recent insights. Hypertension. 2005;46(6):1243–1249. doi: 10.1161/01.HYP.0000188408.49896.c5. [DOI] [PubMed] [Google Scholar]
- 3.Villar J, Carroli G, Wojdyla D, Abalos E, Giordano D, Ba’aqeel H, Farnot U, Bergsjø P, Bakketeig L, Lumbiganon P, Campodónico L, Al-Mazrou Y, Lindheimer M, Kramer M World Health Organization Antenatal Care Trial Research Group. Preeclampsia, gestational hypertension and intrauterine growth restriction, related or independent conditions? Am J Obstet Gynecol. 2006;194(4):921–931. doi: 10.1016/j.ajog.2005.10.813. [DOI] [PubMed] [Google Scholar]
- 4.Stennett AK, Khalil RA. Neurovascular mechanisms of hypertension in pregnancy. Curr Neurovasc Res. 2006;3(2):131–148. doi: 10.2174/156720206776875885. [DOI] [PubMed] [Google Scholar]
- 5.Sholook MM, Gilbert JS, Sedeek MH, Huang M, Hester RL, Granger JP. Systemic hemodynamic and regional blood flow changes in response to chronic reductions in uterine perfusion pressure in pregnant rats. Am J Physiol Heart Circ Physiol. 2007;293(4):H2080–H2084. doi: 10.1152/ajpheart.00667.2007. [DOI] [PubMed] [Google Scholar]
- 6.Skjaerven R, Vatten LR, Wilcox AJ, Ronning T, Irgens LM, Lie RT. Recurrence of pre-eclampsia across generations: exploring fetal and maternal genetic components in a population based cohort. Obst Gynecol Surv. 2006;61(3):162–163. doi: 10.1136/bmj.38555.462685.8F. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Cross JC. The genetics of pre-eclampsia: a feto-placental or maternal problem? Clin Genet. 2003;64:96–103. doi: 10.1034/j.1399-0004.2003.00127.x. [DOI] [PubMed] [Google Scholar]
- 8.Winn VD, Gormley M, Paquet AC, Kjar-Sorensen K, Kramer A, Rumer KK, Haimov-Kochman R, Yeh RF, Overgaard MT, Varki A, Oxvig C, Fisher SJ. Severe preeclampsia-related changes in gene expression at the maternal-fetal interface include sialic acid-binding immunoglobulin-like lectin-6 and pappalysin-2. Endocrinology. 2009;150(1):452–62. doi: 10.1210/en.2008-0990. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Founds SA, Conley YP, Lyons-Weiler JF, Jeyabalan A, Hogge WA, Conrad KP. Altered global gene expression in first trimester placentas of women destined to develop preeclampsia. Placenta. 2009;30(1):15–24. doi: 10.1016/j.placenta.2008.09.015. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Zafarmand MH, Franx A, Sabour S, Van Der Schouw YT, Grobbee DE, De Leeuw PW, Bots ML. The M235T variant of the angiotensinogen gene is related to development of self-reported hypertension during pregnancy: the Prospect-EPIC Cohort Study. Hypertens Res. 2008;31:1299–1305. doi: 10.1291/hypres.31.1299. [DOI] [PubMed] [Google Scholar]
- 11.Piccoli JC, Gottlieb MG, Castro L, Bodanese LC, Manenti ER, Bogo MR, Peres A, Rocha MI, Cruz IB. Association between 894>T endothelial nitric oxide synthase gene polymorphisms and metabolic syndrome. Arq Bras Endocrinol Metabol. 2008;52:1367–1373. doi: 10.1590/s0004-27302008000800027. [DOI] [PubMed] [Google Scholar]
- 12.Sohda S, Arinami T, Hamada H, Yamada N, Hamaguchi H, Kubo T. Methylenetetrahydrofolate reductase polymorphism and pre-eclampsia. J Med Genet. 1997;34:525–526. doi: 10.1136/jmg.34.6.525. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Genetics of Pre-eclampsia (GOPEC) consortium. Babies, pre-eclamptic mothers and grandparents: a three-generation phenotyping study. J Hypertens. 2007;25(4):849–854. doi: 10.1097/HJH.0b013e32803fb634. [DOI] [PubMed] [Google Scholar]
- 14.Duckers C, Simioni P, Spiezia L, Radu C, Gavasso S, Rosing J, Castoldi E. Low plasma levels of tissue factor pathway inhibitor in patients with congenital factor V deficiency. Blood. 2008;112(9):3615–3623. doi: 10.1182/blood-2008-06-162453. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Mayor-Olea A, Callejón G, Palomares AR, Jiménez AJ, Gaitán MJ, Rodríguez A, Ruiz M, Reyes-Engel A. Human genetic selection on the MTHFR 677C>T polymorphism. BMC Med Genet. 2008;9:104. doi: 10.1186/1471-2350-9-104. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Nishizawa H, Pryor-Koishi K, Suzuki M, Kato T, Kogo H, Sekiya T, Kurahashi H, Udagawa Y. Increased levels of pregnancy-associated plasma protein-A2 in the serum of pre-eclamptic patients. Mol Hum Reprod. 2008;14(10):595–602. doi: 10.1093/molehr/gan054. [DOI] [PubMed] [Google Scholar]
- 17.Fitzpatrick E, Göring HH, Liu H, Borg A, Forrest S, Cooper DW, Brennecke SP, Moses EK. Fine mapping and SNP analysis of positional candidates at the preeclampsia susceptibility locus (PREG1) on chromosome 2. Hum Biol. 2004;76(6):849–862. doi: 10.1353/hub.2005.0017. [DOI] [PubMed] [Google Scholar]
- 18.Brinkman-Van der Linden EC, Hurtado-Ziola N, Hayakawa T, Wiggleton L, Benirschke K, Varki A, Varki N. Human-specific expression of Siglec-6 in the placenta. Glycobiology. 2007;17(9):922–931. doi: 10.1093/glycob/cwm065. [DOI] [PubMed] [Google Scholar]
- 19.Nejatizadeh A, Stobdan T, Malhotra N, Qadar Pasha MA. The genetic aspects of pre-eclampsia: Achievments and limitations. Biochem Genet. 2008;46:451–479. doi: 10.1007/s10528-008-9163-9. [DOI] [PubMed] [Google Scholar]
- 20.Ward K. Searching for genetic factors underlying pre-eclampsia: recent progress and persistent challenges. Minerva Ginecol. 2008;60(5):399–419. [PubMed] [Google Scholar]
- 21.Rosenberg TJ, Garbers S, Lipkind H, Chiasson MA. Maternal obesity and diabetes as risk factors for adverse pregnancy outcomes: differences among 4 racial/ethnic groups. Am J Public Health. 2005;95(9):1545–1551. doi: 10.2105/AJPH.2005.065680. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Tanaka M, Jaamaa G, Kaiser M, Hills E, Soim A, Zhu M, Shcherbatykh IY, Samelson R, Bell E, Zdeb M, McNutt LA. Racial disparity in hypertensive disorders of pregnancy in New York State: a 10-year longitudinal population-based study. Am J Public Health. 2007;97(1):163–170. doi: 10.2105/AJPH.2005.068577. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Silva LM. Low socioeconomic status is a risk factor for preeclampsia: the Generation R Study. J Hypertens. 2008;26(6):1200–1208. doi: 10.1097/HJH.0b013e3282fcc36e. [DOI] [PubMed] [Google Scholar]
- 24.Thomas M, Weisman SM. Calcium supplementation during pregnancy and lactation: effects on the mother and the fetus. Am J Obstet Gynecol. 2006;194(4):937–945. doi: 10.1016/j.ajog.2005.05.032. [DOI] [PubMed] [Google Scholar]
- 25.Kumar A, Salam GD, Batra S, Singh C, Shukla DK. Calcium supplementation for the prevention of pre-eclampsia. Int J Gynaecol Obstet. 2008;104(1):32–36. doi: 10.1016/j.ijgo.2008.08.027. [DOI] [PubMed] [Google Scholar]
- 26.Oken E, Ning Y, Rifas-Shiman SL, Rich-Edwards JW, Olsen SF, Gillman MW. Diet during pregnancy and risk of preeclampsia or gestational hypertension. Ann Epidemiol. 2007;17(9):663–668. doi: 10.1016/j.annepidem.2007.03.003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Villar J, Abdel-Aleem H, Merialdi M, Mathai M, Ali MM, Zavaleta N, Purwar M, Hofmeyr J, Nguyen TN, Campódonico L, Landoulsi S, Carroli G, Lindheimer M World Health Organization Calcium Supplementation for the Prevention of Preeclampsia Trial Group. World Health Organization randomized trial of calcium supplementation among low calcium intake pregnant women. Am J Obstet Gynecol. 2006;194(3):639–649. doi: 10.1016/j.ajog.2006.01.068. [DOI] [PubMed] [Google Scholar]
- 28.Prada JA, Tsang RC, Clark KE. Hypocalcemia and pregnancy-induced hypertension produced by low-calcium diet. Hypertension. 1994;23(6):695–702. doi: 10.1161/01.hyp.23.6.695. [DOI] [PubMed] [Google Scholar]
- 29.Roberts JM, Balk JL, Bodnar LM, Belizan JM, Bergel E, Martinez A. Nutrition as a preventive strategy against adverse pregnancy outcomes. J Nutr. 2003;133:1684S–1692S. doi: 10.1093/jn/133.5.1684S. [DOI] [PubMed] [Google Scholar]
- 30.Frederick IO, Williams MA, Dashow E, Kestin M, Zhang C, Leisenring WM. Dietary fiber, potassium, magnesium and calcium in relation to the risk of preeclampsia. J Reprod Med. 2005;50:332–344. [PubMed] [Google Scholar]
- 31.Bodnar LM, Catov JM, Simhan HN, Holick MF, Powers RW, Roberts JM. Maternal vitamin D deficiency increases the risk of preeclampsia. J Clin Endocrinol Metab. 2007;92(9):3517–3522. doi: 10.1210/jc.2007-0718. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Li YC, Kong J, Wei M, Chen ZF, Liu SQ, Cao LP. 1,25-Dihydroxyvitamin D(3) is a negative endocrine regulator of the renin-angiotensin system. J Clin Invest. 2002;110(2):229–238. doi: 10.1172/JCI15219. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Holmes VA, McCance DR. Could antioxidant supplementation prevent pre-eclampsia? Proc Nutr Soc. 2005;64:491–501. doi: 10.1079/pns2005469. [DOI] [PubMed] [Google Scholar]
- 34.Wen SW, Chen XK, Rodger M, White RR, Yang Q, Smith GN, Sigal RJ, Perkins SL, Walker MC. Folic acid supplementation in early second trimester and the risk of preeclampsia. Am J Obstet Gynecol. 2008;198:45.e1–7. doi: 10.1016/j.ajog.2007.06.067. [DOI] [PubMed] [Google Scholar]
- 35.LaMarca BD, Gilbert J, Granger JP. Recent progress toward the understanding of the pathophysiology of hypertension during preeclampsia. Hypertension. 2008;51:982–988. doi: 10.1161/HYPERTENSIONAHA.107.108837. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Chen XK, Yang Q, Smith G, Krewski D, Walker M, Wen SW. Environmental lead level and pregnancy-induced hypertension. Environ Res. 2006;100:424–430. doi: 10.1016/j.envres.2005.07.006. [DOI] [PubMed] [Google Scholar]
- 37.Vigeh M, Yokoyama K, Mazaheri M, Beheshti S, Ghazizadeh S, Sakai T, Morita Y, Kitamura F, Araki S. Relationship between increased blood lead and pregnancy hypertension in women without occupational lead exposure in Tehran, Iran. Arch Environ Health. 2004;59(2):70–75. doi: 10.3200/AEOH.59.2.70-75. [DOI] [PubMed] [Google Scholar]
- 38.Jeyabalan A, Powers RW, Durica AR, Harger G, Roberts JM, Ness RB. Cigarette smoke exposure and angiogenic factors in pregnancy and preeclampsia. Am J Hypertens. 2008;21(8):943–947. doi: 10.1038/ajh.2008.219. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Mutter WP, Karumanchi SA. Molecular mechanisms of preeclampsia. Microvasc Res. 2008;75(1):1–8. doi: 10.1016/j.mvr.2007.04.009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Yogev Y, Visser GHA. Obesity, gestational diabetes and preganancy outcome. Semin Fetal Neonatal Med. 2009;14:77–84. doi: 10.1016/j.siny.2008.09.002. [DOI] [PubMed] [Google Scholar]
- 41.Ray J, Diamond P, Singh G, Bell C. Brief overview of maternal triglycerides as a risk factor for pre-eclampsia. BJO G. 2006;113:379–386. doi: 10.1111/j.1471-0528.2006.00889.x. [DOI] [PubMed] [Google Scholar]
- 42.Ramsay JE, Jamieson N, Greer IA, Sattar N. Paradoxical elevation in adiponectin concentrations in women with preeclampsia. Hypertension. 2003;42(5):891–894. doi: 10.1161/01.HYP.0000095981.92542.F6. [DOI] [PubMed] [Google Scholar]
- 43.Hendler I, Blackwell SC, Mehta SH, Whitty JE, Russell E, Sorokin Y, Cotton DB. The levels of leptin, adiponectin, and resistin in normal weight, overweight, and obese pregnant women with and without preeclampsia. Am J Obstet Gynecol. 2005;193:979–983. doi: 10.1016/j.ajog.2005.06.041. [DOI] [PubMed] [Google Scholar]
- 44.Leddy MA, Power ML, Schulkin J. The impact of maternal obesity on maternal and fetal health. Rev Obstet Gynecol. 2008;1(4):170–178. [PMC free article] [PubMed] [Google Scholar]
- 45.HAPO Study Cooperative Research Group. Metzger BE, Lowe LP, Dyer AR, Trimble ER, Chaovarindr U, Coustan DR, Hadden DR, McCance DR, Hod M, McIntyre HD, Oats JJ, Persson B, Rogers MS, Sacks DA. Hyperglycemia and adverse pregnancy outcomes. N Engl J Med. 2008;358(19):1991–2002. doi: 10.1056/NEJMoa0707943. [DOI] [PubMed] [Google Scholar]
- 46.Carpenter MW. Gestational Diabetes, Pregnancy Hyppertension, and Late Vascular Disease. Diabetes Care. 2007;30(Suppl 2):S246–S250. doi: 10.2337/dc07-s224. [DOI] [PubMed] [Google Scholar]
- 47.Duckitt K, Harrington D. Risk factors for pre-eclampsia at antenatal booking: systematic review of controlled studies. BM J. 2005;330(7491):565. doi: 10.1136/bmj.38380.674340.E0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Fisher SJ. The placental problem: linking abnormal cytotrophoblast differentiation to the maternal symptoms of preeclampsia. Reprod Biol Endocrinol. 2004;2:53. doi: 10.1186/1477-7827-2-53. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Alexander BT. Fetal programming of hypertension. Am J Physiol Regul Integr Comp Physiol. 2006;290(1):R1–R10. doi: 10.1152/ajpregu.00417.2005. [DOI] [PubMed] [Google Scholar]
- 50.Ojeda NB, Grigore D, Alexander BT. Intrauterine growth restriction: fetal programming of hypertension and kidney disease. Adv Chronic Kidney Dis. 2008;15(2):101–106. doi: 10.1053/j.ackd.2008.01.001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Gilbert JS, Ryan MJ, LaMarca BB, Sedeek M, Murphy SR, Granger JP. Pathophysiology of hypertension during preeclampsia: linking placental ischemia with endothelial dysfunction. Am J Physiol Heart Circ Physiol. 2008;294(2):H541–H550. doi: 10.1152/ajpheart.01113.2007. [DOI] [PubMed] [Google Scholar]
- 52.Orshal JM, Khalil RA. Reduced endothelial NO-cGMP-mediated vascular relaxation and hypertension in IL-6-infused pregnant rats. Hypertension. 2004;43:434–444. doi: 10.1161/01.HYP.0000113044.46326.98. [DOI] [PubMed] [Google Scholar]
- 53.Murphy JG, Fleming JB, Cockrell KL, Granger JP, Khalil RA. [Ca2+]I signaling in renal arterial smooth muscle cells of pregnant rat is enhanced during inhibition of NO synthesis. Am J Physiol Regul Integr Comp Physiol. 2001;280(1):R87–R99. doi: 10.1152/ajpregu.2001.280.1.R87. [DOI] [PubMed] [Google Scholar]
- 54.Kanasaki K, Palmsten K, Sugimoto H, Ahmad S, Hamano Y, Xie L, Parry S, Augustin HG, Gattone VH, Folkman J, Strauss JF, Kalluri R. Deficiency in catechol-O-methyltransferase and 2-methoxyoestradiol is associated with pre-eclampsia. Nature. 2008;453(7198):1117–1121. doi: 10.1038/nature06951. [DOI] [PubMed] [Google Scholar]
- 55.Uddin MN, Agunanne E, Horvat D, Puschett JB. Alterations in the renin-angiotensin system in a rat model of human preeclampsia. Am J Nephrol. 2009;31(2):171–177. doi: 10.1159/000267099. [DOI] [PubMed] [Google Scholar]
- 56.Davisson RL, Hoffmann DS, Butz GM, Aldape G, Schlager G, Merrill DC, Sethi S, Weiss RM, Bates JN. Discovery of a spontaneous genetic mouse model of preeclampsia. Hypertension. 2002;39(2 Pt 2):337–342. doi: 10.1161/hy02t2.102904. [DOI] [PubMed] [Google Scholar]
- 57.Zenclussen AC. A novel mouse model for preeclampsia by transferring activated th1 cells into normal pregnant mice. Methods Mol Med. 2006;122:401–412. doi: 10.1385/1-59259-989-3:401. [DOI] [PubMed] [Google Scholar]
- 58.Smárason AK, Sargent IL, Starkey PM, Redman CW. The effect of placental syncytiotrophoblast microvillous membranes from normal and pre-eclamptic women on the growth of endothelial cells in vitro. Br J Obstet Gynaecol. 1993;100(10):943–949. doi: 10.1111/j.1471-0528.1993.tb15114.x. [DOI] [PubMed] [Google Scholar]
- 59.Kertesz Z, Hurst G, Ward M, Willis AC, Caro H, Linton EA, Sargent IL, Redman CW. Purification and characterization of a complex from placental syncytiotrophoblast microvillous membranes which inhibits the proliferation of human umbilical vein endothelial cells. Placenta. 1999;20(1):71–79. doi: 10.1053/plac.1998.0351. [DOI] [PubMed] [Google Scholar]
- 60.Wang Y, Lewis DF, Gu Y, Zhang Y, Alexander JS, Granger DN. Placental trophoblast-derived factors diminish endothelial barrier function. J Clin Endocrinol Metab. 2004;89(5):2421–2428. doi: 10.1210/jc.2003-031707. [DOI] [PubMed] [Google Scholar]
- 61.Van Wijk MJ, Boer K, Nisell H, Smarason AK, Van Bavel E, Kublickiene KR. Endothelial function in myometrial resistance arteries of normal pregnant women perfused with syncytiotrophoblast microvillous membranes. Br J Obstet Gynecol. 2001;108:967–972. doi: 10.1111/j.1471-0528.2001.00221.x. [DOI] [PubMed] [Google Scholar]
- 62.Shaarawy M, Al-Sokkary F, Sheba M, Wahba O, Kandil HO, Abdel-Mohsen I. Angiogenin and vascular endothelial growth factor in pregnancies complicated by preeclampsia. Int J Gynaecol Obstet. 2005;88:112–117. doi: 10.1016/j.ijgo.2004.10.005. [DOI] [PubMed] [Google Scholar]
- 63.Stepan H, Geide A, Faber R. Soluble fms-like tyrosine kinase 1. N Engl J Med. 2004;351(21):2241–2242. doi: 10.1056/NEJM200411183512123. [DOI] [PubMed] [Google Scholar]
- 64.Levine RJ, Lam C, Qian C, Yu KF, Maynard SE, Sachs BP, Sibai BM, Epstein FH, Romero R, Thadhani R, Karumanchi SA CPEP Study Group. Soluble endoglin and other circulating antiangiogenic factors in preeclampsia. N Engl J Med. 2006;355:992–1005. doi: 10.1056/NEJMoa055352. [DOI] [PubMed] [Google Scholar]
- 65.Hayashi M, Ueda Y, Yamaguchi T, Sohma R, Shibazaki M, Ohkura T, Inaba N. Tumor Necrosis Factor-a in the placenta is not elevated in the pre-eclamptic patients despite its elevation in peripheral blood. Am J Reprod Immunol. 2005;53:113–119. doi: 10.1111/j.1600-0897.2005.00253.x. [DOI] [PubMed] [Google Scholar]
- 66.Casart YC, Tarrazzi K, Camejo MI. Serum levels of interleukin-6, interleukin-1beta and human chorionic gonadotropin in pre-eclamptic and normal pregnancy. Gynecol Endocrinol. 2007;23(5):300–303. doi: 10.1080/09513590701327638. [DOI] [PubMed] [Google Scholar]
- 67.Tsukimori K, Fukushima K, Tsushima A, Nakano H. Generation of reactive oxygen species by neutrophils and endothelial cell injury in normal and preeclamptic pregnancies. Hypertension. 2005;46:696–700. doi: 10.1161/01.HYP.0000184197.11226.71. [DOI] [PubMed] [Google Scholar]
- 68.Rajakumar A, Whitelock KA, Weissfeld LA, Daftary AR, Markovic N, Conrad KP. Selective overexpression of the hypoxia-inducible transcription factor, HIF-2alpha, in placentas from women with preeclampsia. Biol Reprod. 2001;64:499–506. doi: 10.1093/biolreprod/64.2.499. [DOI] [PubMed] [Google Scholar]
- 69.Narumiya H, Zhang Y, Fernandez-Patron C, Guilbert LJ, Davidge ST. Matrix metalloproteinase-2 is elevated in the plasma of women with preeclampsia. Hypertens Pregnancy. 2001;20(2):185–194. doi: 10.1081/PRG-100106968. [DOI] [PubMed] [Google Scholar]
- 70.Ranta V, Viinikka L, Halmesmäki E, Ylikorkala O. Nitric oxide production with preeclampsia. Obstet Gynecol. 1999;93(3):442–445. doi: 10.1016/s0029-7844(98)00465-7. [DOI] [PubMed] [Google Scholar]
- 71.Crews JK, Herrington JN, Granger JP, Khalil RA. Decreased endothelium-dependent vascular relaxation during reduction of uterine perfusion pressure in pregnant rat. Hypertension. 2000;35:367–372. doi: 10.1161/01.hyp.35.1.367. [DOI] [PubMed] [Google Scholar]
- 72.Kenny LC, Baker PN, Kendall DA, Randall MD, Dunn WR. Differential mechanisms of endothelium-dependent vasodilator responses in human myometrial small arteries in normal pregnancy and pre-eclampsia. Clin Sci (Lond) 2002;103:67–73. doi: 10.1042/cs1030067. [DOI] [PubMed] [Google Scholar]
- 73.Clark BA, Halvorson L, Sachs B, Epstein FH. Plasma endothelin levels in preeclampsia: elevation and correlation with uric acid levels and renal impairment. Am J Obstet Gynecol. 1992;166(3):962–968. doi: 10.1016/0002-9378(92)91372-h. [DOI] [PubMed] [Google Scholar]
- 74.Brosnihan KB, Neves LA, Anton L, Joyner J, Valdes G, Merrill DC. Enhanced expression of Ang-(1–7) during pregnancy. Braz J Med Biol Res. 2004;37:1255–1262. doi: 10.1590/s0100-879x2004000800017. [DOI] [PubMed] [Google Scholar]
- 75.Maynard SE, Min JY, Merchan J, Lim KH, Li J, Mondal S, Libermann TA, Morgan JP, Sellke FW, Stillman IE, Epstein FH, Sukhatme VP, Karumanchi SA. Excess placental soluble fms-like tyrosine kinase 1 (sFlt1) may contribute to endothelial dysfunction, hypertension, and proteinuria in preeclampsia. J Clin Invest. 2003;111:649–658. doi: 10.1172/JCI17189. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 76.Purwosunu Y, Sekizawa A, Farina A, Wibowo N, Koide K, Okazaki S, Nakamura M, Okai T. Evaluation of physiological alterations of the placenta through analysis of cell-free messenger ribonucleic acid concentrations of angiogenic factors. Am J Obstet Gynecol. 2008;198(1):124.e1–7. doi: 10.1016/j.ajog.2007.06.079. [DOI] [PubMed] [Google Scholar]
- 77.LaMarca BB, Bennett WA, Alexander BT, Cockrell K, Granger JP. Hypertension produced by reductions in uterine perfusion in the pregnant rat: role of tumor necrosis factor-alpha. Hypertension. 2005;46(4):1022–1025. doi: 10.1161/01.HYP.0000175476.26719.36. [DOI] [PubMed] [Google Scholar]
- 78.LaMarca BD, Ryan MJ, Gilbert JS, Murphy SR, Granger JP. Inflammatory cytokines in the pathophysiology of hypertension during preeclampsia. Curr Hypertens Rep. 2007;9(6):480–485. doi: 10.1007/s11906-007-0088-1. [DOI] [PubMed] [Google Scholar]
- 79.Alexander BT, Kassab SE, Miller MT, Abram SR, Reckelhoff JF, Bennett WA, Granger JP. Reduced uterine perfusion pressure during pregnancy in the rat is associated with increases in arterial pressure and changes in renal nitric oxide. Hypertension. 2001;37(4):1191–1195. doi: 10.1161/01.hyp.37.4.1191. [DOI] [PubMed] [Google Scholar]
- 80.Wang Y, Walsh S, Guo J, Zhang J. The imbalance between thromboxane and prostacyclin in preeclampsia. Curr Hypertens. 1991;165:1695–1700. doi: 10.1016/0002-9378(91)90017-l. [DOI] [PubMed] [Google Scholar]
- 81.LaMarca BB, Cockrell K, Sullivan E, Bennett W, Granger JP. Role of endothelin in mediating tumor necrosis factor-induced hypertension in pregnant rats. Hypertension. 2005;46(1):82–86. doi: 10.1161/01.HYP.0000169152.59854.36. [DOI] [PubMed] [Google Scholar]
- 82.Lamarca B, Wallukat G, Llinas M, Herse F, Dechend R, Granger JP. Autoantibodies to the angiotensin type I receptor in response to placental ischemia and tumor necrosis factor-alpha in pregnant rats. Hypertension. 2008;52(6):1168–1172. doi: 10.1161/HYPERTENSIONAHA.108.120576. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 83.Murphy JG, Fleming JB, Cockrell KL, Granger JP, Khalil RA. [Ca2+]i signaling in renal arterial smooth muscle cells of pregnant rat is enhanced during inhibition of NOS. Am J Physiol Regul Integr Comp Physiol. 2001;280(1):R87–R99. doi: 10.1152/ajpregu.2001.280.1.R87. [DOI] [PubMed] [Google Scholar]
- 84.Murphy JG, Herrington JN, Granger JP, Khalil RA. Enhanced [Ca2+]i in renal arterial smooth muscle cells of pregnant rats with reduced uterine perfusion pressure. Am J Physiol Heart Circ Physiol. 2003;284(1):H393–H403. doi: 10.1152/ajpheart.00247.2002. [DOI] [PubMed] [Google Scholar]
- 85.Munaut C, Lorquet S, Pequeux C, Blacher S, Berndt S, Frankenne F, Foidart JM. Hypoxia is responsible for soluble vascular endothelial growth factor receptor-1 (VEGFR-1) but not for soluble endoglin induction in villous trophoblast. Hum Reprod. 2008;23(6):1407–1415. doi: 10.1093/humrep/den114. [DOI] [PubMed] [Google Scholar]
- 86.Akercan F, Cirpan T, Terek MC, Ozcakir HT, Giray G, Sagol S, Karadadas N. The immunohistochemical evaluation of VEGF in placenta biopsies of pregnancies complicated by preeclampsia. Arch Gynecol Obstet. 2008;277(2):109–114. doi: 10.1007/s00404-007-0430-5. [DOI] [PubMed] [Google Scholar]
- 87.Bosio PM, Wheeler T, Anthony F, Conroy R, O’herlihy C, McKenna P. Maternal plasma vascular endothelial growth factor concentrations in normal and hypertensive pregnancies and their relationship to peripheral vascular resistance. Am J Obstet Gynecol. 2001;184:146–152. doi: 10.1067/mob.2001.108342. [DOI] [PubMed] [Google Scholar]
- 88.Serraf A, Aznag H, Baudet B, Détruit H, Séccatore F, Mazmanian MG, Planché C. Pulmonary vascular endothelial growth factor and nitric oxide interaction during total cardiopulmonary bypass in neonatal pigs. J Thorac Cardiovasc Surg. 2003;125:1050–1057. doi: 10.1067/mtc.2003.402. [DOI] [PubMed] [Google Scholar]
- 89.Osol G, Celia G, Gokina N, Barron C, Chien E, Madala M, Luksha L, Kublickiene K. Placental growth factor is a potent vasodilator of rat and human resistance arteries. Am J Physiol Heart Circ Physiol. 2008;294:H1381–H1387. doi: 10.1152/ajpheart.00922.2007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 90.Kim SY, Ryu HM, Yang JH, Kim MY, Han JY, Kim JO, Chung JH, Park SY, Lee MH, Kim DJ. Increased sFlt-1 to PlGF ratio in women who subsequently develop preeclampsia. J Korean Med Sci. 2007;22(5):873–877. doi: 10.3346/jkms.2007.22.5.873. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 91.Woolcock J, Hennessy A, Xu B, Thornton C, Tooher J, Makris A, Ogle R. Soluble Flt-1 as a diagnostic marker of pre-eclampsia. Aust NZJ Obstet Gynaecol. 2008;48:64–70. doi: 10.1111/j.1479-828X.2007.00804.x. [DOI] [PubMed] [Google Scholar]
- 92.Boyd PA, Lindenbaum RH, Redman C. Pre-eclampsia and trisomy 13: a possible association. Lancet. 1987;2:425–427. doi: 10.1016/s0140-6736(87)90960-3. [DOI] [PubMed] [Google Scholar]
- 93.Tuohy JF, James DK. Pre-eclampsia and trisomy 13. Br J Obstet Gynaecol. 1992;99:891–894. doi: 10.1111/j.1471-0528.1992.tb14436.x. [DOI] [PubMed] [Google Scholar]
- 94.Hirashima C, Ohkuchi A, Matsubara S, Suzuki H, Takahashi K, Usui R, Suzuki M. Alteration of serum soluble endoglin levels after the onset of preeclampsia is more pronounced in women with early-onset. Hypertens Res. 2008;31(8):1541–1548. doi: 10.1291/hypres.31.1541. [DOI] [PubMed] [Google Scholar]
- 95.Venkatesha S, Toporsian M, Lam C, Hanai J, Mammoto T, Kim YM, Bdolah Y, Lim KH, Yuan HT, Libermann TA, Stillman IE, Roberts D, D’Amore PA, Epstein FH, Sellke FW, Romero R, Sukhatme VP, Letarte M, Karumanchi SA. Soluble endoglin contributes to the pathogenesis of preeclampsia. Nat Med. 2006;2:642–649. doi: 10.1038/nm1429. [DOI] [PubMed] [Google Scholar]
- 96.ten Dijke P, Goumans MJ, Pardali E. Endoglin in angiogenesis and vascular diseases. Angiogenesis. 2008;11(1):79–89. doi: 10.1007/s10456-008-9101-9. [DOI] [PubMed] [Google Scholar]
- 97.Conrad KP, Miles TM, Benyo DF. Circulating levels of immunoreactive cytokines in women with preeclampsia. Am J Reprod Immunol. 1998;40:102–111. doi: 10.1111/j.1600-0897.1998.tb00398.x. [DOI] [PubMed] [Google Scholar]
- 98.Hashii K, Fujiwara H, Yoshioka S, Kataoka N, Yamada S, Hirano T, Mori T, Fujii S, Maeda M. Peripheral blood mononuclear cells stimulate progesterone production by luteal cells derived from pregnant and non-pregnant women: possible involvement of interleukin-4 and interleukin-10 in corpus luteum function and differentiation. Hum Reprod. 1998;13(10):2738–2744. doi: 10.1093/humrep/13.10.2738. [DOI] [PubMed] [Google Scholar]
- 99.Gadonski G, LaMarca BB, Sullivan E, Bennett W, Chandler D, Granger JP. Hypertension produced by reductions in uterine perfusion in the pregnant rat: role of interleukin 6. Hypertension. 2006;48(4):711–716. doi: 10.1161/01.HYP.0000238442.33463.94. [DOI] [PubMed] [Google Scholar]
- 100.Alexander BT, Cockrell KL, Massey MB, Bennett WA, Granger JP. Tumor necrosis factor-α-induced hypertension in pregnant rats results in decreased renal neuronal nitric oxide synthase expression. Am J Hypertens. 2002;15:12–19. doi: 10.1016/s0895-7061(01)02255-5. [DOI] [PubMed] [Google Scholar]
- 101.Sedeek M, Gilbert JS, LaMarca BB, Sholook M, Chandler DL, Wang Y, Granger JP. Role of reactive oxygen species in hypertension produced by reduced uterine perfusion in pregnant rats. Am J Hypertension. 2008;21(10):1152–1156. doi: 10.1038/ajh.2008.239. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 102.Sato H, Sato M, Kanai H, Uchiyama T, Iso T, Ohyama Y, Sakamoto H, Tamura J, Nagai R, Kurabayashi M. Mitochondrial reactive oxygen species and c-Src play a critical role in hypoxic response in vascular smooth muscle cells. Cardiovasc Res. 2005;67:714–722. doi: 10.1016/j.cardiores.2005.04.017. [DOI] [PubMed] [Google Scholar]
- 103.Tyurin VA, Liu SX, Tyurina YY, Sussman NB, Hubel CA, Roberts JM, Taylor RN, Kagan VE. Elevated levels of S-nitrosoalbumin in preeclampsia plasma. Circ Res. 2001;88(11):1210–1215. doi: 10.1161/hh1101.092179. [DOI] [PubMed] [Google Scholar]
- 104.Chambers JC, Fusi L, Malik IS, Haskard DO, De Swiet M, Kooner JS. Association of maternal endothelial dysfunction with preeclampsia. JAM A. 2001;285:1607–1612. doi: 10.1001/jama.285.12.1607. [DOI] [PubMed] [Google Scholar]
- 105.Roberts JM, Speer P. Antioxidant therapy to prevent preeclampsia. Semin Nephrol. 2004;24:557–564. doi: 10.1016/s0270-9295(04)00126-3. [DOI] [PubMed] [Google Scholar]
- 106.Parra M, Rodrigo R, Barja P, Bosco C, Fernández V, Muñoz H, Soto-Chacón E. Screening test for preeclampsia through assessment of uteroplacental blood flow and biochemical markers of oxidative stress and endothelial dysfunction. Am J Obstet Gynecol. 2005;193(4):1486–1491. doi: 10.1016/j.ajog.2005.02.109. [DOI] [PubMed] [Google Scholar]
- 107.Yung HW, Calabrese S, Hynx D, Hemmings BA, Cetin I, Charnock-Jones DS, Burton GJ. Evidence of placental translation inhibition and endoplasmic reticulum stress in the etiology of human intrauterine growth restriction. Am J Pathol. 2008;173:451–462. doi: 10.2353/ajpath.2008.071193. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 108.Chang TC, Huang CJ, Tam K, Chen SF, Tan KT, Tsai MS, Lin TN, Shyue SK. Stabilization of hypoxia-inducible factor-1{alpha} by prostacyclin under prolonged hypoxia via reducing reactive oxygen species level in endothelial cells. J Biol Chem. 2005;280(44):36567–36574. doi: 10.1074/jbc.M504280200. [DOI] [PubMed] [Google Scholar]
- 109.Egi K, Conrad NE, Kwan J, Schulze C, Schulz R, Wildhirt SM. Inhibition of inducible nitric oxide synthase and superoxide production reduces matrix metalloproteinase-9 activity and restores coronary vasomotor function in rat cardiac allografts. Eur J of Cardiothorac Surg. 2004;26:262–269. doi: 10.1016/j.ejcts.2004.04.037. [DOI] [PubMed] [Google Scholar]
- 110.Jin L, Ying Z, Webb RC. Activation of Rho/Rho kinase signaling pathway by reactive oxygen species in rat aorta. Am J Physiol Heart Circ Physiol. 2004;287:H1495–H1500. doi: 10.1152/ajpheart.01006.2003. [DOI] [PubMed] [Google Scholar]
- 111.Faller DV. Endothelial Cell Responses to Hypoxic Stress. Clin Exp Pharmacol Physiol. 1999;26:74–84. doi: 10.1046/j.1440-1681.1999.02992.x. [DOI] [PubMed] [Google Scholar]
- 112.Yamashita K, Discher DJ, Hu J, Bishopric NH, Webster KA. Molecular regulation of the endothelin-1 gene by hypoxia. Contributions of hypoxia-inducible factor-1, activator protein-1, GATA-2, AND p300/CBP. J Biol Chem. 2001;276(16):12645–12653. doi: 10.1074/jbc.M011344200. [DOI] [PubMed] [Google Scholar]
- 113.Kelly BA, Bond BC, Poston L. Aortic adaptation to pregnancy: elevated expression of matrix metalloproteinases-2 and -3 in rat gestation. Mol Hum Reprod. 2004;10:331–337. doi: 10.1093/humrep/gah045. [DOI] [PubMed] [Google Scholar]
- 114.Galewska Z, Bañkowski E, Romanowicz L, Jaworski S. Pre-eclampsia (EPH-gestosis)-induced decrease of MMP-s content in the umbilical cord artery. Clin Chim Acta. 2003;335(1–2):109–115. doi: 10.1016/s0009-8981(03)00296-1. [DOI] [PubMed] [Google Scholar]
- 115.Wang H, Olszewski B, Rosebury W, Wang D, Robertson A, Keiser JA. Impaired angiogenesis in SHR is associated with decreased KDR and MT1-MMP expression. Biochem Biophys Res Commun. 2004;315:363–368. doi: 10.1016/j.bbrc.2004.01.059. [DOI] [PubMed] [Google Scholar]
- 116.Stillman IE, Karumanchi SA. The glomerular injury of preeclampsia. J Am Soc Nephrol. 2007;18:2281–2284. doi: 10.1681/ASN.2007020255. [DOI] [PubMed] [Google Scholar]
- 117.Baumwell S, Karumanchi SA. Pre-eclampsia: clinical manifestations and molecular mechanisms. Nephron Clin Pract. 2007;106(2):c72–81. doi: 10.1159/000101801. [DOI] [PubMed] [Google Scholar]
- 118.Anumba DO, Robson SC, Boys RJ, Ford GA. Nitric oxide activity in the peripheral vasculature during normotensive and preeclamptic pregnancy. Am J Physiol. 1999;277:H848–H854. doi: 10.1152/ajpheart.1999.277.2.H848. [DOI] [PubMed] [Google Scholar]
- 119.Nelson SH, Steinsland OS, Wang Y, Yallampalli C, Dong YL, Sanchez JM. Increased nitric oxide synthase activity and expression in the human uterine artery during pregnancy. Circ Res. 2000;87:406–411. doi: 10.1161/01.res.87.5.406. [DOI] [PubMed] [Google Scholar]
- 120.Conrad KP, Kerchner LJ, Mosher MD. Plasma and 24-h NO(x) and cGMP during normal pregnancy and preeclampsia in women on a reduced NO(x) diet. Am J Physiol. 1999;277:F48–F57. doi: 10.1152/ajprenal.1999.277.1.F48. [DOI] [PubMed] [Google Scholar]
- 121.Mutlu-Turkoglu U, Aykac-Toker G, Ibrahimoglu L, Ademoglu E, Uysal M. Plasma nitric oxide metabolites and lipid peroxide levels in preeclamptic pregnant women before and after delivery. Gynecol Obstet Invest. 1999;48:247–250. doi: 10.1159/000010192. [DOI] [PubMed] [Google Scholar]
- 122.Schiessl B, Strasburger C, Bidlingmaier M, Mylonas I, Jeschk eU, Kainer F, Friese K. Plasma- and urine concentrations of nitrite/nitrate and cyclic Guanosinemonophosphate in intrauterine growth restricted and preeclamptic pregnancies. Arch Gynecol Obstet. 2006;274(3):150–154. doi: 10.1007/s00404-006-0149-8. [DOI] [PubMed] [Google Scholar]
- 123.Conrad KP, Davis AK. Nitric oxide synthase activity in placentae from women with pre-eclampsia. Placenta. 1995;16:691–699. doi: 10.1016/0143-4004(95)90013-6. [DOI] [PubMed] [Google Scholar]
- 124.Alexander BT, Miller MT, Kassab S, Novak J, Reckelhoff JF, Kruckeberg WC, Granger JP. Differential expression of renal nitric oxide synthase isoforms during pregnancy in rats. Hypertension. 1999;33(1 Pt 2):435–439. doi: 10.1161/01.hyp.33.1.435. [DOI] [PubMed] [Google Scholar]
- 125.Roberts JM. Endothelial dysfunction in preeclampsia. Semin Reprod Endocrinol. 1998;16:5–15. doi: 10.1055/s-2007-1016248. [DOI] [PubMed] [Google Scholar]
- 126.Molnar M, Suto T, Toth T, Hertelendy F. Prolonged blockade of nitric oxide synthesis in gravid rats produces sustained hypertension, proteinuria, thrombocytopenia, and intrauterine growth retardation. Am J Obstet Gynecol. 1994;170:1458–1466. doi: 10.1016/s0002-9378(94)70179-2. [DOI] [PubMed] [Google Scholar]
- 127.Khalil RA, Crews JK, Novak J, Kassab S, Granger JP. Enhanced vascular reactivity during inhibition of nitric oxide synthesis in pregnant rats. Hypertension. 1998;31:1065–1069. doi: 10.1161/01.hyp.31.5.1065. [DOI] [PubMed] [Google Scholar]
- 128.Suzuki Y, Hattori T, Kajikuri J, Yamamoto T, Suzumori K, Itoh T. Reduced function of endothelial prostacyclin in human omental resistance arteries in pre-eclampsia. J Physiol. 2002;545:269–277. doi: 10.1113/jphysiol.2002.022384. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 129.Fike CD, Kaplowitz MR, Pfister SL. Arachidonic acid metabolites and an early stage of pulmonary hypertension in chronically hypoxic newborn pigs. Am J Physiol Lung Cell Mol Physiol. 2003;284:L316–L323. doi: 10.1152/ajplung.00228.2002. [DOI] [PubMed] [Google Scholar]
- 130.Bowen RS, Zhang Y, Gu Y, Lewis DF, Wang Y. Increased phospholipase A2 and thromboxane but not prostacyclin production by placental trophoblast cells from normal and preeclamptic pregnancies cultured under hypoxia condition. Placenta. 2005;26(5):402–409. doi: 10.1016/j.placenta.2004.07.007. [DOI] [PubMed] [Google Scholar]
- 131.Scotland RS, Madhani M, Chauhan S, Moncada S, Andresen J, Nilsson H, Hobbs AJ, Ahluwalia A. Investigation of vascular responses in endothelial nitric oxide synthase/cyclooxygenase-1 double-knockout mice: key role for endothelium-derived hyperpolarizing factor in the regulation of blood pressure in vivo. Circulation. 2005;111(6):796–803. doi: 10.1161/01.CIR.0000155238.70797.4E. [DOI] [PubMed] [Google Scholar]
- 132.Fulep EE, Vedernikov YP, Saade GR, Garfield RE. The role of endothelium-derived hyperpolarizing factor in the regulation of the uterine circulation in pregnant rats. Am J Obstet Gynecol. 2001;185:638–642. doi: 10.1067/mob.2001.117665. [DOI] [PubMed] [Google Scholar]
- 133.Petersson J, Zygmunt PM, Jonsson P, Hogestatt ED. Characterization of endothelium- dependent relaxation in guinea pig basilar artery - effect of hypoxia and role of cytochrome P450 mono-oxygenase. J Vasc Res. 1998;35:285–294. doi: 10.1159/000025595. [DOI] [PubMed] [Google Scholar]
- 134.Resta TC, Walker BR. Chronic hypoxia selectively augments endothelium-dependent pulmonary arterial vasodilation. Am J Physiol. 1996;270:H888–H896. doi: 10.1152/ajpheart.1996.270.3.H888. [DOI] [PubMed] [Google Scholar]
- 135.Kenny LC, Baker PN, Kendall DA, Randall MD, Dunn WR. Differential mechanisms of endothelium-dependent vasodilator responses in human myometrial small arteries in normal pregnancy and pre-eclampsia. Clin Sci (Lond) 2002;103:67–73. doi: 10.1042/cs1030067. [DOI] [PubMed] [Google Scholar]
- 136.Luksha L, Nisell H, Luksha N, Kublickas M, Hultenby K, Kublickiene K. Endothelium-derived hyperpolarizing factor in preeclampsia: heterogeneous contribution, mechanisms, and morphological prerequisites. Am J Physiol Regul Integr Comp Physiol. 2008;294(2):R510–R519. doi: 10.1152/ajpregu.00458.2007. [DOI] [PubMed] [Google Scholar]
- 137.Niwano K, Arai M, Tomaru K, Uchiyama T, Ohyama Y, Kurabayashi M. Transcriptional stimulation of the eNOS gene by the stable prostacyclin analogue beraprost is mediated through cAMP-responsive element in vascular endothelial cells: close link between PGI2 signal and NO pathways. Circ Res. 2003;93(6):523–530. doi: 10.1161/01.RES.0000091336.55487.F7. [DOI] [PubMed] [Google Scholar]
- 138.Orie NN, Fry CH, Clapp LH. Evidence that inward rectifier K+ channels mediate relaxation by the PGI2 receptor agonist cicaprost via a cyclic AMP-independent mechanism. Cardiovasc Res. 2006;69:107–115. doi: 10.1016/j.cardiores.2005.08.004. [DOI] [PubMed] [Google Scholar]
- 139.Bernardi F, Constantino L, Machado R, Petronilho F, Dal-Pizzol F. Plasma nitric oxide, endothelin-1, arginase and superoxide dismutase in pre-eclamptic women. J Obstet Gynaecol Res. 2008;34(6):957–963. doi: 10.1111/j.1447-0756.2008.00860.x. [DOI] [PubMed] [Google Scholar]
- 140.Kourembanas S, McQuillan LP, Leung GK, Faller DV. Nitric oxide regulates the expression of vasoconstrictors and growth factors by vascular endothelium under both normoxia and hypoxia. J Clin Invest. 1993;92:99–104. doi: 10.1172/JCI116604. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 141.Taylor RN, Varma M, Teng NN, Roberts JM. Women with preeclampsia have higher plasma endothelin levels than women with normal pregnancies. J Clin Endocrinol Metab. 1990;71:1675–1677. doi: 10.1210/jcem-71-6-1675. [DOI] [PubMed] [Google Scholar]
- 142.Weigand L, Sylvester JT, Shimoda LA. Mechanisms of endothelin-1-induced contraction in pulmonary arteries from chronically hypoxic rats. Am J Physiol Lung Cell Mol Physiol. 2006;290(2):L284–L290. doi: 10.1152/ajplung.00449.2004. [DOI] [PubMed] [Google Scholar]
- 143.Giardina JB, Green GM, Rinewalt AN, Granger JP, Khalil RA. Role of endothelin B receptors in enhancing endothelium-dependent nitric oxide-mediated vascular relaxation during high salt diet. Hypertension. 2001;37:516–523. doi: 10.1161/01.hyp.37.2.516. [DOI] [PubMed] [Google Scholar]
- 144.Conrad KP, Gandley RE, Ogawa T, Nakanishi S, Danielson LA. Endothelin mediates renal vasodilation and hyperfiltration during pregnancy in chronically instrumented conscious rats. Am J Physiol. 1999;276:F767–F776. doi: 10.1152/ajprenal.1999.276.5.F767. [DOI] [PubMed] [Google Scholar]
- 145.Gandley RE, Conrad KP, McLaughlin MK. Endothelin and nitric oxide mediate reduced myogenic reactivity of small renal arteries from pregnant rats. Am J Physiol Regul Integr Comp Physiol. 2001;280:R1–R7. doi: 10.1152/ajpregu.2001.280.1.R1. [DOI] [PubMed] [Google Scholar]
- 146.Chassagne C, Eddahibi S, Adamy C, Rideau D, Marotte F, Dubois-Randé JL, Adnot S, Samuel JL, Teiger E. Modulation of angiotensin II receptor expression during development and regression of hypoxic pulmonary hypertension. Am J Respir Cell Mol Biol. 2000;22(3):323–332. doi: 10.1165/ajrcmb.22.3.3701. [DOI] [PubMed] [Google Scholar]
- 147.Merrill DC, Karoly M, Chen K, Ferrario CM, Brosnihan KB. Angiotensin-(1–7) in normal and preeclamptic pregnancy. Endocrine. 2002;18:239–245. doi: 10.1385/ENDO:18:3:239. [DOI] [PubMed] [Google Scholar]
- 148.Wesselman JP, De Mey JG. Angiotensin and cytoskeletal proteins: role in vascular remodeling. Curr Hypertens Rep. 2002;4:63–70. doi: 10.1007/s11906-002-0055-9. [DOI] [PubMed] [Google Scholar]
- 149.Mizutani S, Tomodak Y. Effects of placental proteases on maternal and fetal blood pressure in normal pregnancy and preeclampsia. Am J Hypertens. 1996;9(6):591–597. doi: 10.1016/0895-7061(96)00016-7. [DOI] [PubMed] [Google Scholar]
- 150.Dechend R, Homuth V, Wallukat G, Kreuzer J, Park JK, Theuer J, Juepner A, Gulba DC, Mackman N, Haller H, Luft FC. AT(1) receptor agonistic antibodies from preeclamptic patients cause vascular cells to express tissue factor. Circulation. 2000;101(20):2382–2387. doi: 10.1161/01.cir.101.20.2382. [DOI] [PubMed] [Google Scholar]
- 151.Wijayagunawardane MP, Gabler C, Killian G, Miyamoto A. Tumor necrosis factor alpha in the bovine oviduct during the estrous cycle: messenger RNA expression and effect on secretion of prostaglandins, endothelin-1, and angiotensin II. Biol Reprod. 2003;69:1341–1346. doi: 10.1095/biolreprod.103.017327. [DOI] [PubMed] [Google Scholar]
- 152.Wassmann S, Stumpf M, Strehlow K, Schmid A, Schieffer B, Böhm M, Nickenig G. Interleukin-6 induces oxidative stress and endothelial dysfunction by overexpression of the angiotensin II type 1 receptor. Circ Res. 2004;94(4):534–541. doi: 10.1161/01.RES.0000115557.25127.8D. [DOI] [PubMed] [Google Scholar]
- 153.Cingolani HE, Villa-Abrille MC, Cornelli M, Nolly A, Ennis IL, Garciarena C, Suburo AM, Torbidoni V, Correa MV, Camiliónde Hurtado MC, Aiello EA. The positive inotropic effect of angiotensin II: role of endothelin-1 and reactive oxygen species. Hypertension. 2006;47(4):727–734. doi: 10.1161/01.HYP.0000208302.62399.68. [DOI] [PubMed] [Google Scholar]
- 154.Cheng ZJ, Vapaatalo H, Mervaala E. Angiotensin II and vascular inflammation. Med Sci Monit. 2005;11:RA194–RA205. [PubMed] [Google Scholar]
- 155.Askari B, Ferrerib NR. Regulation of prostacyclin synthesis by angiotensin II and TNF-α in vascular smooth muscle. Prostaglandins. 2001;63:175–187. doi: 10.1016/s0090-6980(01)00098-3. [DOI] [PubMed] [Google Scholar]
- 156.Wang D, Chabrashvili T, Borrego L, Aslam S, Umans JG. Angiotensin II infusion alters vascular function in mouse resistance vessels: roles of superoxide and endothelium. J Vasc Res. 2006;43:109–119. doi: 10.1159/000089969. [DOI] [PubMed] [Google Scholar]
- 157.Goto K, Fujii K, Kansui Y, Iida M. Changes in endothelium-derived hyperpolarizing factor in hypertension and ageing: Response to chronic treatment with renin-angiotensin system inhibitors. Clin Exp Pharmacol Physiol. 2004;31:650–655. doi: 10.1111/j.1440-1681.2004.04054.x. [DOI] [PubMed] [Google Scholar]
- 158.Deja MA, Gołba KS, Widenka K, Mrozek R, Biernat J, Kolowca M, Malinowski M, Woś S. Angiotensin-converting enzyme inhibitors reveal non-NO-, non-prostacycline-mediated endothelium-dependent relaxation in internal thoracic artery of hypertensive patients. Int J Cardiol. 2005;102(3):455–460. doi: 10.1016/j.ijcard.2004.05.050. [DOI] [PubMed] [Google Scholar]
- 159.Wang Y, Walsh SW, Kay HH. Placental lipid peroxides and thromboxane are increased and prostacyclin is decreased in women with preeclampsia. Am J Obstet Gynecol. 1999;167:946–949. doi: 10.1016/s0002-9378(12)80017-2. [DOI] [PubMed] [Google Scholar]
- 160.Llinás MT, Alexander BT, Seedek M, Abram SR, Crell A, Granger JP. Enhanced thromboxane synthesis during chronic reductions in uterine perfusion pressure in pregnant rats. Am J Hypertension. 2002;15(9):793–797. doi: 10.1016/s0895-7061(02)02975-8. [DOI] [PubMed] [Google Scholar]
- 161.Coomarasamy A, Papaioannou S, Gee H, Khan KS. Aspirin for the prevention of preeclampsia in women with abnormal uterine artery doppler: a meta-analysis. Obstet Gynecol. 2001;98:861–866. doi: 10.1016/s0029-7844(01)01569-1. [DOI] [PubMed] [Google Scholar]
- 162.Coomarasamy A, Honest H, Papaioannou S, Gee H, Khan KS. Aspirin for prevention of preeclampsia in women with historical risk factors: a systematic review. Obstet Gynecol. 2003;101(6):1319–1332. doi: 10.1016/s0029-7844(03)00169-8. [DOI] [PubMed] [Google Scholar]
- 163.de Berrazueta JR. The role of calcium in the regulation of normal vascular tone and in arterial hypertension. Rev Esp Cardiol. 1999;52(3):25–33. [PubMed] [Google Scholar]
- 164.Wimalasundera RC, Thom SA, Regan L, Hughes AD. Effects of vasoactive agents on intracellular calcium and force in myometrial and subcutaneous resistance arteries isolated from preeclamptic, pregnant, and nonpregnant woman. Am J Obstet Gynecol. 2005;192(2):625–632. doi: 10.1016/j.ajog.2004.07.040. [DOI] [PubMed] [Google Scholar]
- 165.Magness RR, Rosenfeld CR, Carr BR. Protein kinase C in uterine and systemic arteries during ovarian cycle and pregnancy. Am J Physiol. 1991;260(3):E464–E470. doi: 10.1152/ajpendo.1991.260.3.E464. [DOI] [PubMed] [Google Scholar]
- 166.Kanashiro CA, Cockrell KL, Alexander BT, Granger JP, Khalil RA. Pregnancy-associated reduction in vascular protein kinase C activity rebounds during inhibition of NO synthesis. Am J Physiol Regul Integr Comp Physiol. 2000;278(2):R295–R303. doi: 10.1152/ajpregu.2000.278.2.R295. [DOI] [PubMed] [Google Scholar]
- 167.Kanashiro CA, Alexander BT, Granger JP, Khalil RA. Ca2+-insensitive vascular protein kinase C during pregnancy and NOS inhibition. Hypertension. 1999;34:924–930. doi: 10.1161/01.hyp.34.4.924. [DOI] [PubMed] [Google Scholar]
- 168.Horowitz A, Menice CB, Laporte R, Morgan KG. Mechanisms of smooth muscle contraction. Physiol Rev. 1996;76(4):967–1003. doi: 10.1152/physrev.1996.76.4.967. [DOI] [PubMed] [Google Scholar]
- 169.Salamanca DA, Khalil RA. Protein kinase C isoforms as specific targets for modulation of vascular smooth muscle function in hypertension. Biochem Pharmacol. 2005;70(11):1537–1547. doi: 10.1016/j.bcp.2005.07.017. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 170.VanWijk MJ, Boer K, van der Meulen ET, Bleker OP, Spaan JA, VanBavel E. Resistance artery smooth muscle function in pregnancy and preeclampsia. Am J Obstet Gynecol. 2002;186(1):148–154. doi: 10.1067/mob.2002.119184. [DOI] [PubMed] [Google Scholar]
- 171.Wallukat G, Homuth V, Fischer T, Lindschau C, Horstkamp B, Jüpner A, Baur E, Nissen E, Vetter K, Neichel D, Dudenhausen JW, Haller H, Luft FC. Patients with preeclampsia develop agonistic autoantibodies against the angiotensin AT1 receptor. J Clin Invest. 1999;103:945–952. doi: 10.1172/JCI4106. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 172.Hu E. Recent patents on Rho signaling pathway as therapeutic target for cardiovascular diseases. Recent Patents Cardiovasc Drug Discov. 2006;1(3):249–263. doi: 10.2174/157489006778777034. [DOI] [PubMed] [Google Scholar]
- 173.Kataoka C, Egashira K, Inoue S, Takemoto M, Ni W, Koyanagi M, Kitamoto S, Usui M, Kaibuchi K, Shimokawa H, Takeshita A. Important role of Rho-kinase in the pathogenesis of cardiovascular inflammation and remodeling induced by long-term blockade of nitric oxide synthesis in rats. Hypertension. 2002;39(2):245–250. doi: 10.1161/hy0202.103271. [DOI] [PubMed] [Google Scholar]
- 174.Friel AM, Sexton DJ, O’reilly MW, Smith TJ, Morrison JJ. Rho A/Rho kinase: human umbilical artery mRNA expression in normal and pre eclamptic pregnancies and functional role in isoprostane-induced vasoconstriction. Reproduction. 2006;132(1):169–176. doi: 10.1530/rep.1.01088. [DOI] [PubMed] [Google Scholar]
- 175.Lynch AM, Murphy JR, Byers T, Gibbs RS, Neville MC, Giclas PC, Salmon JE, Holers VM. Alternative complement pathway activation gragment Bb in early pregnancy as a predictor of preeclampsia. Am J Obstet Gynecol. 2008;198(4):385.e1–9. doi: 10.1016/j.ajog.2007.10.793. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 176.Dundar O, Yoruk P, Tutuneu L, Erikci AA, Muheu M, Ergur AR, Atay V, Mungen E. Longitudinal study of platelet size changes in gestation and predictive power of elevated MPV in development of pre-eclampsia. Prenat Diagn. 2008;28:1052–1056. doi: 10.1002/pd.2126. [DOI] [PubMed] [Google Scholar]
- 177.Bainbridge SA, Roberts JM. Uric acid as a pathotgenic factor in preeclampsia. Placenta. 2008;22:S67–S72. doi: 10.1016/j.placenta.2007.11.001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 178.Anand-Ivell R, Ivell R, Driscoll D, Manson J. Insulin-like factor 3 levels in amniotic fluid of human male fetuses. Hum Reprod. 2008;23(5):1180–1186. doi: 10.1093/humrep/den038. [DOI] [PubMed] [Google Scholar]
- 179.Cromi A, Ghezzi F, Raffaelli R, Bergamini V, Siesto G, Bolis P. Ultrasonographic measurement of thymus size in IUGR fetuses: a marker of the fetal immunoendocrine response to malnutrition. Ultrasound Obstet Gynecol. 2009;33(4):421–426. doi: 10.1002/uog.6320. [DOI] [PubMed] [Google Scholar]
- 180.Hunt BJ, Missfelder-Lobos H, Parra-Cordero M, Fletcher O, Parmar K, Lefkou E, Lees CC. Pregnancy outcome and fibrinolytic, endothelial and coagulation markers in women undergoing uterine artery Doppler screening at 23 weeks. J Thromb Haemost. 2009;7(6):955–961. doi: 10.1111/j.1538-7836.2009.03344.x. [DOI] [PubMed] [Google Scholar]
- 181.Abalos E, Duley L, Steyn DW, Henderson-Smart DJ. Antihypertensive drug therapy for mild to moderate hypertension during pregnancy. Cochrane Database Syst Rev. 2007;(1):CD002252. doi: 10.1002/14651858.CD002252.pub2. [DOI] [PubMed] [Google Scholar]
- 182.Chowdhury JR, Chaudhuri S, Bhattacharyya N, Biswas PK, Panpalia M. Comparison of intramuscular magnesium sulfate with low dose intravenous magnesium sulfate regimen for treatment of eclampsia. J Obstet Gynaecol Res. 2009;35(1):119–125. doi: 10.1111/j.1447-0756.2008.00842.x. [DOI] [PubMed] [Google Scholar]
- 183.Facchinetti F, Saade GR, Neri I, Pizzi C, Longo M, Volpe A. L-arginine supplementation in patients with gestational hypertension: a pilot study. Hypertens Pregnancy. 2007;26(1):121–130. doi: 10.1080/10641950601147994. [DOI] [PubMed] [Google Scholar]
- 184.Hladunewich MA, Derby GC, Lafayette RA, Blouch KL, Druzin ML, Myers BD. Effect of L-arginine therapy on the glomerular injury of preeclampsia: a randomized controlled trial. Obstet Gynecol. 2006;107(4):886–895. doi: 10.1097/01.AOG.0000207637.01824.fe. [DOI] [PubMed] [Google Scholar]
- 185.Li Z, Zhang Y, Ying Ma J, Kapoun AM, Shao Q, Kerr I, Lam A, O’Young G, Sannajust F, Stathis P, Schreiner G, Karumanchi SA, Protter AA, Pollitt NS. Recombinant vascular endothelial growth factor 121 attenuates hypertension and improves kidney damage in rat model of preeclampsia. Hypertension. 2007;50:686–692. doi: 10.1161/HYPERTENSIONAHA.107.092098. [DOI] [PubMed] [Google Scholar]
- 186.Wang Y, Lewis DF, Adair CD, Gu Y, Mason L, Kipikasa JH. Digibind attenuates cytokine TNFα-induced endothelial inflammatory response: potential benefit role of Digibind in preeclampsia. J Perinatol. 2009;29(3):195–200. doi: 10.1038/jp.2008.222. [DOI] [PMC free article] [PubMed] [Google Scholar]