

Abstract
Epithelial cell migration is an essential part of embryogenesis and tissue regeneration, yet their migration is least understood. Using our three-dimensional (3D) motility analysis, migrating epithelial cells formed an atypical polarized cell shape with the nucleus leading the cell front and a contractile cell rear. Migrating epithelial cells exerted traction forces to deform both the anterior and posterior extracellular matrix toward the cell body. The cell leading edge exhibited a myosin II-dependent retrograde flow with the magnitude and direction consistent with surrounding network deformation. Interestingly, on a two-dimensional substrate, myosin IIA-deficient cells migrated faster than wild-type cells, but in a 3D gel, these myosin IIA-deficient cells were unpolarized and immobile. In contrast, the migration rates of myosin IIB-deficient cells were similar to wild-type cells. Therefore, myosin IIA, not myosin IIB, is required for 3D epithelial cell migration.
Main Text
Diverse arrays of cells are capable of migrating, but not all cells migrate in the same way. Many cells crawl asynchronously with a distinct membrane extension at the leading edge and a cell body retraction to move the bulky nucleus forward while others rapidly glide on the surface with a constant cell shape (1). At the leading edge, the constant retrograde flow of actin network transmits forces needed for cell movement. This transient engagement between force generating actin machinery and the surrounding matrix is a molecular clutch that regulates the efficiency of cell migration (2).
Despite the different types of cell migration, non-muscle myosin II emerged as the underlying core machinery that drives cell movement. Particularly, the distinct roles of myosin II isoforms in cell migration became evident. Of three myosin isoforms (A, B, C), the more ubiquitously expressed isoforms A and B are distinctly localized in migrating cells. The absence or reduced amount of myosin IIA caused increased protrusion, in most cases, increased motility and reduced retrograde flow in spreading cells (3). In contrast, myosin IIB was essential for persistent cell migration and cell polarization (3). However, the roles of myosin II isoforms in generating traction forces in a three-dimensional matrix have not been elucidated.
We analyzed epithelial cell migration in a three-dimensional collagen matrix. In contrast to previously described motile cells, these single cells maintained a constant cell shape with the wider region containing the nucleus and leading the cell body (Fig. 1 A, Movie S1 of the Supporting Material). These cells migrated at 3.8 ± 0.3 μm/h (Fig. 3 D) without significant changes in the cell body length (<15% of the total cell length, Fig. S1 D). Many cells migrated without membrane activity at the leading edge, suggesting that rapidly extending and retracting protrusions are not required for cell migration.
Figure 1.
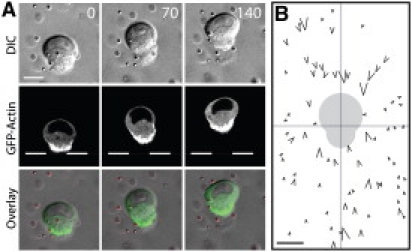
Epithelial cells migrate with a constant polarized shape in a 3D matrix. (A) A GFP-actin expressing MDCK epithelial cell migrating in a 3D collagen matrix. (B) Traction force generated by migrating cells deform surrounding matrix. The individual tracer particles around 10 migrating cells were superimposed relative to the cell centroid (center). The arrow direction and size reflects the direction and magnitude of the particle motion. Gray cell shadow is the relative position of migrating cells. Scale bar 10 μm. Time in minutes.
Figure 3.
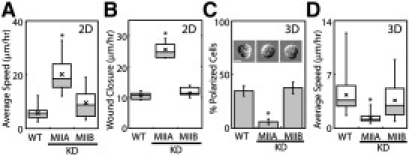
Epithelial migration is myosin IIA-dependent. (A) Box plot of the rates of single cell migration of wild-type (n = 33), myosin IIA (n = 11) or B knockdown (KD) cells (n = 21) on a 2D surface. ∗P < 0.001. (B) Box plot of wound closure rate of wild-type (n = 9), myosin IIA (n = 13) or B knockdown cells (n = 10). ∗P < 0.001. (C) The percent of polarized cells in a 3D gel, for wild-type (n = 98), myosin IIA (n = 83) and IIB knockdown cells (n = 80). Top: cell phenotypes. Scale bar 10 μm. ∗P < 0.001. (D) Box plot of the average speed of wild-type (n = 38), myosin IIA (n = 54), and B (n = 64) knockdown cells in a 3D matrix. ∗P <0.001.
In the absence of a well-defined protrusion, the actin network was distributed throughout the cortex of migrating cells, but was highly concentrated at the trailing edge (Fig. 1 A), suggesting that the cell rear may act as a contractile structure. To examine the traction forces exerted by migrating cells, we quantitatively analyzed the surrounding matrix deformations by embedding fluorescent particles in the matrix as fiducial markers. Both anterior and posterior particles moved toward migrating cells (Fig. 1 B, Fig. S1 C), suggesting that both the front and rear of the migrating cell exert pulling forces. Based on particle displacements, migrating cells exerted ∼100 nN of traction force (see Methods in the Supporting Material). Similar matrix deformation patterns were observed with epithelial cells migrating in Matrigel (Fig. S1 A and B). This is a stark contrast to the amoeboid mode of migration in which cells extend leading protrusions to push the matrix away from the cell body (4). Our results are more consistent with the mesenchymal mode of migration in which cells retract leading protrusions to pull the matrix toward the cell body (5), albeit without the distinct extension and retraction cycles required for net cell displacement.
It was not immediately clear as to how migrating cells generate the observed anterior and posterior matrix deformation. The focal adhesion protein, paxillin, accumulated at the trailing edge (Fig. S1 E). This residual matrix adhesion may be sufficient for migrating cells to drag the posterior matrix, and this is consistent with the observed posterior matrix deformation.
To elucidate how the cell leading edge deforms the anterior matrix, we analyzed the movement of tracer particles that came in direct contact with the surface of migrating cells. Once the particle became coupled to the leading edge, the particle moved rapidly along the cell cortex toward the cell rear (Fig. 2 A black arrow, Movie S2), and either remained there or was released into the matrix. The retrograde flow was consistent with the matrix deformation at the immediate vicinity of cell cortex (Fig. 1 B and S1 B).
Figure 2.
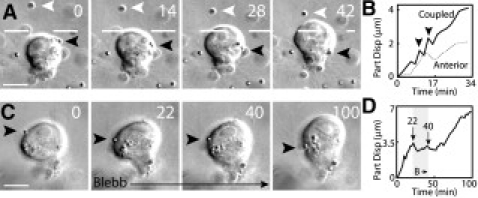
Myosin II dependent retrograde flow of migrating cell in a 3D matrix. (A) The particle movement along the cell cortex. Black and white arrowheads track the coupled and anterior particle, respectively. (B) The displacement of the coupled (Coupled) and anterior (Anterior) particle. (C) Blebbistatin prevents the retrograde flow on the cell cortex (black arrow). (D) The particle displacement before, during and after blebbistatin treatment. Scale bar 10 μm. Time in minutes.
This retrograde movement was greater than the deformation of the anterior collagen network (Fig. 2 B), suggesting that the cellular force was not fully translated into matrix deformation. Interestingly, the retrograde flow and anterior particle displacement occasionally stalled (Fig. 2 B, arrowheads). The increasing load on the retrograde flow due to increasing matrix strain could induce sudden matrix fracture or release, which is consistent with load-and-fail cycles predicted by a molecular clutch model (6). In the absence of well-defined protrusions, these results suggest that retrograde flow generated by the cortex of migrating cells exerts traction forces to deform the anterior matrix.
Since traction forces are often generated by actin-myosin contractions, we examined the roles of nonmuscle myosin II in cell migration using blebbistatin, a small molecule inhibitor of myosin II ATPase activity. The retrograde flow immediately ceased during the brief blebbistatin treatment and this effect was reversible (Fig. 2 C and D, Movie S3), suggesting that myosin II activity was required for the generation of retrograde flow. With the addition of blebbistatin, the migrating cells immediately stopped, and lost their polarized morphology and actin accumulation at the cell rear (Fig. S2 A, Movie S4). Furthermore, blebbistatin treatment caused the immediate release of both anterior and posterior particles away from the cell, indicating the loss of traction force (Fig. S2 B). A small but significant matrix relaxation was observed after the addition of cytochalasin D (Fig. S2 B), which may be due to the residual activity of myosin II or myosin II-independent forces. Together, these data suggest that the myosin II-dependent retrograde flow engages with the surrounding matrix to exert traction forces to advance cell body.
Previous studies have shown that two myosin II isoforms (A and B) play distinct roles in cell polarization and migration on a two-dimensional substrate (3). In a three-dimensional matrix, however, both myosin II isoforms were concentrated at the cell rear of migrating cells (Fig. S3 A). Using maximum projection images to visualize the three-dimensional organization of myosin network, we found that both myosin II isoforms also formed a ring structure at the central region of the cell body (Fig. S3 A). Furthermore, the phosphorylated (active) myosin light chain was concentrated at the cell cortex and trailing edge (Fig. S3 B), raising possible roles for myosin II in retrograde flow and the return of cortical flow to the cell front, respectively. In a well-defined fibroblast lamellipodia, the retrograde flow is in part generated by myosin IIA (7). Therefore, a similar mechanism may drive the retrograde flow in migrating epithelial cells in a three-dimensional matrix.
To distinguish the roles of each myosin II isoform in epithelial cell migration, we stably silenced myosin IIA or B (Fig. S4 A and B). On a two-dimensional surface, myosin IIA-depleted cells had larger lamella and fewer actin stress fibers than untransfected wild-type cells or myosin IIB-depleted cells (Fig. S4 C). Surprisingly, myosin IIB-deficient cells did not exhibit any morphological phenotype or actin organization defects (Fig. S4 C).
To determine if these morphological or actin organization defects translated into functional deficiencies, we analyzed the role of myosin II isoforms using cell migration assays. On a coverslip, myosin IIA depleted cells migrated faster than wild-type cells (Fig. 3 A). The increased migration rate was more pronounced when cells were forced to migrate in a more directed manner during a two-dimensional wound healing assay (Fig. 3 B). In contrast, myosin IIB knockdown did not affect single cell migration or wound closure rate of cells on a coverslip. One explanation may be that myosin IIA can functionally compensate for the loss of myosin IIB. However, this is unlikely since a previous study demonstrated that the overexpression of myosin IIA could not fully rescue the phenotype of myosin IIB depleted cells (8).
Unlike the phenotypes on a two-dimensional surface, in a three-dimensional matrix, the majority of myosin IIA-depleted cells remained round and unpolarized whereas myosin IIB-depleted cells exhibited a normal phenotype (Fig. 3 C, Fig. S4 D). Only 6% of myosin IIA knockdown cells were polarized whereas >30% of wild-type or myosin IIB knockdown cells were polarized (Fig. 3 C). Consistent with this, myosin IIA knockdown cells migrated slower than wild-type or myosin IIB knockdown cells (Fig. 3 D, Movies S5, S6, and S7). The expression of exogeneous myosin IIA that is insensitive to siRNA was able to rescue this phenotype (Fig. S4 E and F), suggesting that cell migration defects were solely due to loss of myosin IIA, and not a consequence of off target silencing. Although myosin IIA depletion increased the rate of cell migration on a two-dimensional surface, in a three-dimensional matrix, loss of myosin IIA resulted in a drastically reduced cell migration speed.
What drives this dramatic switch in the mechanisms of cell migration on a two-dimensional surface and in a three-dimensional matrix? One possibility is that matrix elasticity plays a key role in this morphological switch. To test this hypothesis, instead of embedding cells in the matrix, cells were seeded on the surface of a collagen matrix, thus providing an elastic two-dimensional substrate. In contrast to migration in a three-dimensional matrix, but similar to that on a stiffer glass coverslip, myosin IIA knockdown cells on an elastic matrix migrated quickly with broad lamellipodia (Fig. S5). This result suggests that the stiffness of the matrix alone is not sufficient for the phenotypic switch from migration on a two- to in a three-dimensional environment. The adaptation of cell morphology to a three-dimensional matrix may require additional matrix cues on the cell dorsal surface (9), a key stimulus for cell intercalation during embryogenesis (10).
Our motility assays revealed that, unlike any other cell types, epithelial cell migration in a three-dimensional matrix does not require extension-retraction cycles. Rather, the cells polarize to form a constant cell shape with retrograde motion along the cell cortex. This cell polarization, retrograde flow, and therefore migration, is solely dependent on myosin IIA. Interestingly, glioma cell invasion, which uses extensive protrusions, also depends on myosin IIA activity (11), underscoring the critical roles that myosin IIA plays in the migration of diverse cell types. Since the pattern of matrix deformation surrounding migrating cells is consistent with the retrograde flow, and both traction forces and retrograde flow are dependent on myosin II, our data suggest that retrograde flow is responsible for the generation of traction forces (Fig. S1 F). Interestingly, this retrograde flow is similar to the myosin IIA-dependent actin flow in other migrating cells on a coverslip (7). Therefore, retrograde flow may be a universal mechanism to generate traction forces for migrating cells in all matrix dimensions.
Supporting Material
Methods, five figures, and seven movies are available at http://www.biophysj.org/biophysj/supplemental/S0006-3495(10)00303-6.
Supporting Material
Acknowledgments
We thank Dr. Rick Horwitz for pSuper and mCherry myosin plasmids.
This work was supported by University of California Davis Startup and Cancer Research Coordinating Committee Fund.
References and Footnotes
- 1.Ridley A.J., Schwartz M.A., Horwitz A.R. Cell migration: integrating signals from front to back. Science. 2003;302:1704–1709. doi: 10.1126/science.1092053. [DOI] [PubMed] [Google Scholar]
- 2.Wang Y.L. Flux at focal adhesions: slippage clutch, mechanical gauge, or signal depot. Sci. STKE. 2007;2007:pe10. doi: 10.1126/stke.3772007pe10. [DOI] [PubMed] [Google Scholar]
- 3.Conti M.A., Adelstein R.S. Nonmuscle myosin II moves in new directions. J. Cell Sci. 2008;121:11–18. doi: 10.1242/jcs.007112. [DOI] [PubMed] [Google Scholar]
- 4.Wyckoff J.B., Pinner S.E., Sahai E. ROCK- and myosin-dependent matrix deformation enables protease-independent tumor-cell invasion in vivo. Curr. Biol. 2006;16:1515–1523. doi: 10.1016/j.cub.2006.05.065. [DOI] [PubMed] [Google Scholar]
- 5.Bloom R.J., George J.P., Wirtz D. Mapping local matrix remodeling induced by a migrating tumor cell using three-dimensional multiple-particle tracking. Biophys. J. 2008;95:4077–4088. doi: 10.1529/biophysj.108.132738. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Chan C.E., Odde D.J. Traction dynamics of filopodia on compliant substrates. Science. 2008;322:1687–1691. doi: 10.1126/science.1163595. [DOI] [PubMed] [Google Scholar]
- 7.Cai Y., Biais N., Sheetz M.P. Nonmuscle myosin IIA-dependent force inhibits cell spreading and drives F-actin flow. Biophys. J. 2006;91:3907–3920. doi: 10.1529/biophysj.106.084806. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Vicente-Manzanares M., Zareno J., Horwitz A.F. Regulation of protrusion, adhesion dynamics, and polarity by myosins IIA and IIB in migrating cells. J. Cell Biol. 2007;176:573–580. doi: 10.1083/jcb.200612043. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Beningo K.A., Dembo M., Wang Y.L. Responses of fibroblasts to anchorage of dorsal extracellular matrix receptors. Proc. Natl. Acad. Sci. USA. 2004;101:18024–18029. doi: 10.1073/pnas.0405747102. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Keller R., Shook D., Skoglund P. The forces that shape embryos: physical aspects of convergent extension by cell intercalation. Phys. Biol. 2008;5:15007. doi: 10.1088/1478-3975/5/1/015007. [DOI] [PubMed] [Google Scholar]
- 11.Beadle C., Assanah M.C., Canoll P. The role of myosin II in glioma invasion of the brain. Mol. Biol. Cell. 2008;19:3357–3368. doi: 10.1091/mbc.E08-03-0319. (r) [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.