

Abstract
Deep brain stimulation (DBS) has recently emerged as a potential treatment for medically intractable neuropsychiatric disorders. Pilot clinical studies with encouraging results have been performed with DBS of the ventral anterior internal capsule (VAIC) and subgenual cingulate white matter (Cg25WM) for the treatment of obsessive-compulsive disorder and depression. However, little is known about the underlying response of individual neurons, or the networks they are connected to, when DBS is applied to the VAIC or Cg25WM. This review summarizes current understanding of the response of axons to DBS, and discusses the general brain network architectures thought to underlie neuropsychiatric disorders. We also employ diffusion tensor imaging tractography to better understand the axonal trajectories surrounding DBS electrodes implanted in the VAIC or Cg25WM. Finally, we attempt to reconcile various data sets by presenting generalized hypotheses on potential therapeutic mechanisms of DBS for neuropsychiatric disease.
Keywords: Deep Brain Stimulation, Psychiatric Disorders, Depression, Obsessive Compulsive Disorder, Tractography, Neurostimulation, Neuromodulation, Review
2. INTRODUCTION
Neuropsychiatric disorders such as major depression and obsessive compulsive disorder (OCD) are severely disabling conditions that interfere with the ability to work, interact socially, or live independently (1, 2). Depression is associated with disturbances in attention, motivation, sleep, motor and mental speed, as well as recurrent thoughts of death (3, 4). OCD is characterized by repetitive behaviors (compulsions) performed to neutralize the anxiety induced by undesired thoughts (obsessions) (5). Depression and OCD are among the most common neuropsychiatric conditions, affecting 5% and 2% respectively, of adult Americans each year (4, 6–8).
Although most patients suffering from depression and OCD are treated effectively with a combination of medications (e.g., serotonin reuptake inhibitors) and/or cognitive behavioral therapy, 20 to 30% of patients fail to respond to treatment (4, 9) and, consequently, remain severely disabled. Despite the effectiveness of ablative surgical procedures (e.g., anterior capsulotomy, anterior cingulotomy, limbic leucotomy, and subcaudate tractotomy) for treating intractable depression and OCD, their destructive and irreversible natures prevent them from being used as a conventional form of treatment (10, 11).
Benabid and colleagues pioneered the concept of using high frequency cathodic electrical stimulation delivered directly into the surgical target to mimic the effect of a surgical lesion (12, 13). Their work led to the development of the commercial clinical technology known as deep brain stimulation (DBS), a reversible stimulation paradigm for treating intractable neurologic disorders (14–16). DBS delivers electrical currents to the brain using electrodes connected to implanted pulse generators. These electrodes are permanently placed into specific anatomical targets and stimulation is adjusted by varying the stimulation mode (monopolar, multipolar), active contact(s), pulse amplitude and duration, and stimulation frequency. DBS represents an established therapy for movement disorders (e.g., Parkinson’s disease, essential tremor, dystonia) (17–19). However, the application of DBS for the treatment of neuropsychiatric disorders remains an experimental procedure, tested on a relatively small number of patients (1, 4, 8, 20–25).
3. DBS FOR DEPRESSION AND OCD
Limited understanding of the abnormalities in brain circuitry that are responsible for neuropsychiatric disorders, together with a limited availability of animal models, have hindered the identification of optimal stimulation targets for neuropsychiatric DBS. The surgical targets reported to date have been empirically selected from existing hypotheses on the pathophysiology of neuropsychiatric disease (16). Specifically, metabolic and imaging studies have been used to identify areas of the brain with abnormal activity levels in depression and OCD (5, 9, 26). Currently, stimulation of the ventral anterior internal capsule (VAIC) (1, 7, 14, 27–29) and the nucleus accumbens (2) are being evaluated for the treatment of both OCD and depression. Similarly, DBS of the subgenual cingulate cortex white matter (Cg25WM) is being evaluated for the treatment of depression (4).
Two of the larger DBS studies for neuropsychiatric disorders were performed by Mayberg et al. (4) and Greenberg et al (1). Mayberg et al. (4) reported remission or near remission of depression symptoms in three of six patients after six months of chronic stimulation of the fiber tracts near the subgenual anterior cingulate (Cg25). Continued antidepressant responses in four patients beyond the period of active stimulation showed long-term improvements in mood as well as cognitive ability. Greenberg et al. (1) extended on the work of Nuttin et al. (27) and reported gains in real-world functioning in a group of six of ten OCD patients after chronic stimulation of the VAIC and ventral striatum. Together, these and other clinical studies (2, 5, 27), demonstrate the potential for future use of DBS in the treatment of neuropsychiatric disorders.
Clearly, larger-scale randomized blinded clinical trials are necessary to address fundamental questions such as the most appropriate surgical target site, the most efficacious stimulation paradigms, as well as long-term clinical outcomes. Despite these limitations, existing evidence suggests that severely debilitated individuals can substantially benefit from DBS technology. However, one big question remains: how does DBS for the treatment of neuropsychiatric disorders work?
4. NEURAL RESPONSE TO DBS ELECTRIC FIELDS
The electric field generated by DBS is a three-dimensionally complex phenomenon that is distributed throughout the brain (30, 31). This electric field is applied to the three-dimensional geometry of the surrounding neural processes (i.e., axons and dendrites) surrounding the electrode. The response of an individual neuron to the applied field is related to the second derivative of the extracellular potential distribution along each neural process (32, 33). In turn, each neuron (or neural process) surrounding the electrode will be subject to both depolarizing and hyperpolarizing stimulation effects (34, 35). Extracellular stimulation can cause a neuron to be either activated or suppressed in different ways and in different neural processes. The specific neuronal response will depend on its positioning with respect to the electrode as well as the stimulation parameters. Neurons directly affected by the stimulation will impact network activity, and the network response will depend on the types of neurons affected and their interconnections.
In general, three classes of neurons can be affected by the stimulation: local cells, afferent inputs, and fibers of passage. Local cells represent neurons that have their cell body in close proximity to the electrode. Afferent inputs represent neurons that project axon terminals to the region of the electrode and whose axon terminals make synaptic connections with local cells. Fibers of passage represent neurons where both the cell body and axon terminals are far from the electrode, but the axonal process of the neuron traces a path that comes in close proximity to the electrode.
To date, it remains unclear which class, or combinations of different neuron classes, stimulated by DBS are responsible for its therapeutic benefit. Multiple studies in neural modeling (36–38), neural recording (39–41), microdialysis (42, 43), and functional imaging (44–46) have attempted to define the mechanisms of action of DBS. In-vitro and in-vivo experimental studies have shown DBS induced inhibition of somatic activity in local cells by activating axons of pre-synaptic inhibitory afferent inputs (47) and/or by blocking voltage-gated membrane channels (48). For this reason, DBS has been thought to produce therapeutic benefits by inhibiting neurons around the electrode. However, theoretical models and multiple experimental studies have found evidence of efferent axonal activation of local cells during therapeutic stimulation (38, 39, 41, 49). In an attempt to reconcile these conflicting experimental findings, theoretical studies have shown that although the soma of local cells can be inhibited by DBS, stimulus-dependent efferent axonal depolarization results in axonal firing and efferent signal transduction (37, 38). These findings suggest that understanding the axonal response to DBS is paramount to understanding the network effects induced by DBS.
While the general mechanisms of DBS remain unknown and continuously debated, we propose that DBS-induced modulation of axonal activity may be an important aspect of these mechanisms. Understanding the axonal response to DBS in the context of neuropsychiatric disorders is of great importance because current surgical targets for electrode implantation lie within white matter areas. Although the direct effects of DBS on axonal activity will continue to be investigated over the coming years, several important findings have been elucidated. Theoretical models suggest that typical DBS parameter settings generate consistent axonal activation in response to supra-threshold stimulation (30, 50). These theoretical predictions of activation are corroborated by empirical and experimental measurements of stimulation spread into fiber tracts that generate behavioral outcomes consistent with axonal activation (e.g. corticospinal tract activation and subsequent muscle contractions with subthalamic nucleus DBS). Typically, stimulation-induced blocking of axonal activity begins only after stimulation frequencies have reached kilohertz levels (51). However, in vitro studies have observed axonal block in response to typical (~100 Hz) DBS frequencies (52). Irrespective of whether axons are activated or inhibited by DBS, defining the specific fiber tracts that are modulated by DBS is a first step toward deciphering its therapeutic mechanisms.
5. DBS TRACTOGRAPHY
Diffusion-tensor imaging (DTI) is a non-invasive imaging technique that can be used to define axonal trajectories through white matter areas of the brain (53–55). In an attempt to understand the geometry and relative positioning of various fiber tracts pertinent to DBS for neuropsychiatric disorders, we used DTI tractography to define axonal pathways surrounding the typical sites of therapeutic stimulation for depression (4) (Figure 1) and OCD (1) (Figure 2). We used the Wakana et al. (56) DTI brain atlas as the basis for our tractography analysis and used SCIrun/BioTensor (SCI Institute, University of Utah) to visualize our results. We defined a region of interest (ROI) with a 40 mm cube centered on each stimulating DBS electrode. Three-by-three tensors describing the direction and magnitude of water diffusion within each voxel in the ROI were used to calculate a set of eigenvectors that described the primary axonal trajectory in each voxel (57–60). Centered on the stimulating contact of the DBS electrode, we placed a 10 mm cube containing 125 equidistantly distributed seed points for streamline tractography (55). Given the high stimulus amplitudes commonly used in DBS for neuropsychiatric applications (14, 27, 29), the 10 mm cube was chosen to be representative of the general spread of direct axonal stimulation (30).
Figure 1.
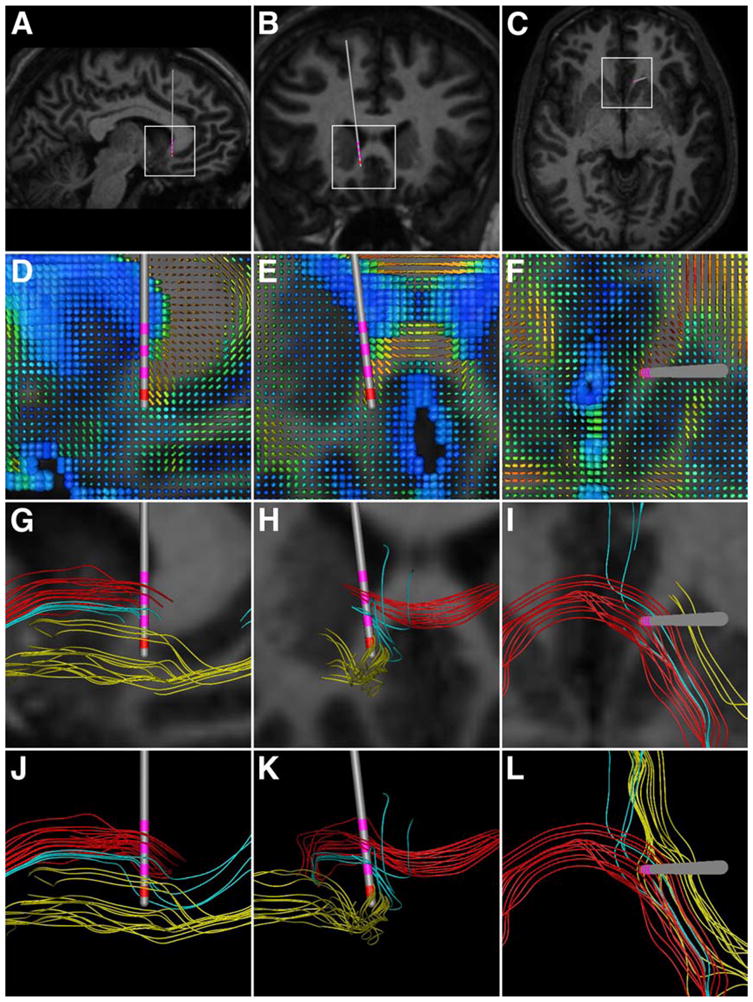
Typical DBS electrode location and fiber trajectories around the subgenual cingulate white matter target for depression. The columns show sagittal, coronal, and axial projections, respectively. (A–C) Region of interest (white rectangular boxes) defined, and DBS electrode (Medtronic 3387) location. Contact 0 represented the center of the 10 mm cube seed region for tractography. (D–F) DTI tensors in the region of interest. The colors depict the individual fractional anisotropy values of the tensors (blue–0; red–1), while the shape and size describe directionality and magnitude, respectively. (G–I) Individual fiber pathways with the corresponding MR. The colors depict three distinct fibers trajectories. (J–L) Fiber pathways without the underlying MR.
Figure 2.
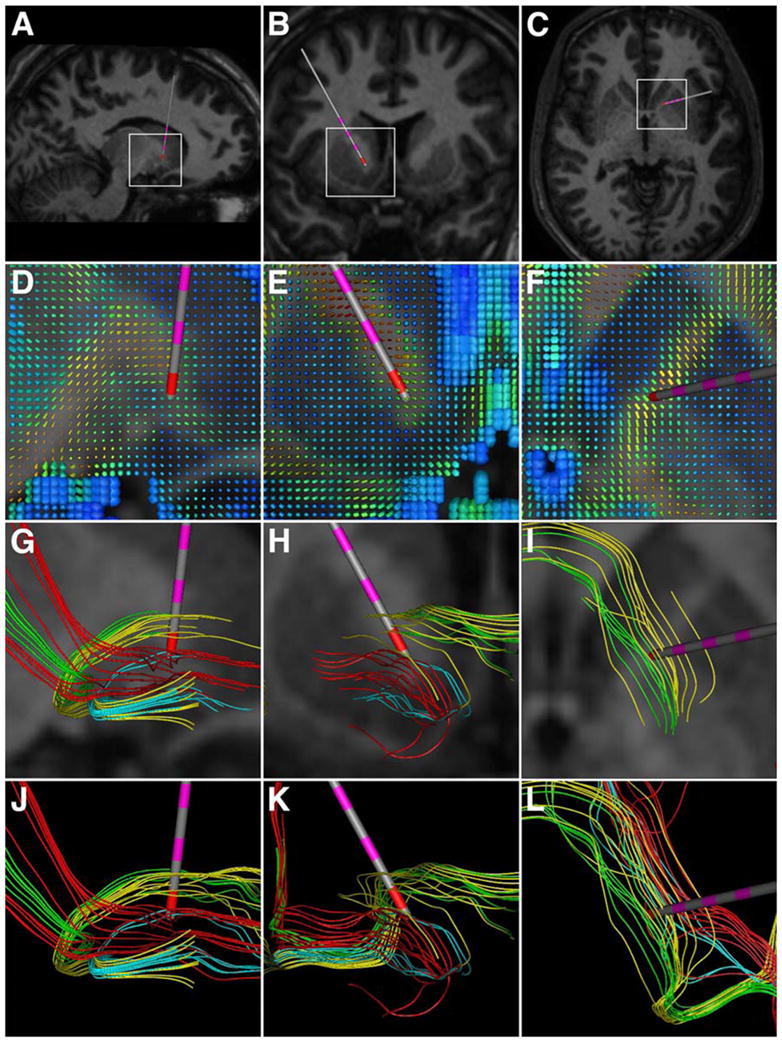
Typical DBS electrode location and fiber trajectories around the ventral anterior internal capsule target for OCD. The columns show sagittal, coronal, and axial projections, respectively. (A–C) Region of interest (white rectangular boxes) defined, and DBS electrode (Medtronic 3887) location. Contact 0 represented the center of the 10 mm cube seed region for tractography. (D–F) DTI tensors in the region of interest. The colors depict the individual fractional anisotropy values of the tensors (blue–0; red–1), while the shape and size describe directionality and magnitude, respectively. (G–I) Individual fiber pathways with the corresponding MR. The colors depict four distinct fibers trajectories. (J–L) Fiber pathways without the underlying MR.
Individual fiber pathways within the stimulating electrode regions for depression and OCD are shown in Figures 1 and 2, respectively. Our results suggest three general axonal trajectories within the region of stimulation for Cg25WM DBS (Figure 1), and four for VAIC DBS (Figure 2). Analysis of the tractography data for both the Cg25WM or VAIC suggests that DBS would activate fiber pathways connecting the regions of orbitofrontal and subgenual anterior cingulate cortices with the ventral striatum (sometimes passing through the anterior pallidum), as well as inter-hemispheric connections. It should be noted that these streamline tractography results suffer from numerous limitations (e.g. normal subject brain, neglect of higher order tensor characterization to accommodate fiber crossings, and difficultly in determining terminations). However, the fundamental purpose of this analysis was to demonstrate the numerous fiber trajectories that surround DBS electrodes and the complex anatomical pathways that exist between brain regions affected by DBS.
Recently, Johansen-Berg et al. (61) used probabilistic tractography to define the likely connectivity of cingulate regions stimulated by DBS. Probabilistic tractography provides statistical details on how likely a given streamline is to connect one brain region to another. Their results show that the effects of Cg25WM DBS may be mediated via strong connections to orbitofrontal, anterior mid cingulate, hypothalamus, accumbens, and amygdala/hippocampus regions.
Our results (Figures 1 and 2) and the work of Johansen-Berg et al. (62) represent necessary first steps to identifying fiber pathways responsible for the therapeutic benefits of DBS for neuropsychiatric disorders. However to create a more complete picture, we (as a community) will need to integrate several additional pieces of information that can be derived from DTI and functional imaging data specifically acquired in neuropsychiatric patients treated with DBS. First, probabilistic tractography can be used to statistically define the most pertinent connected anatomical regions (61). Second, the fiber trajectories connecting these pertinent anatomical regions that also pass in close proximity to the DBS electrode can be defined (Figures 1, 2). Third, DTI-based electric field models can be used to solve for the voltage distribution generated by DBS along each fiber trajectory (30). Fourth, multi-compartment cable models of axons can be used to predict the spatial extent of action potential generation in response to specific stimulation parameter settings (62). Finally, correlation analyses can be performed with functional imaging data (fMRI and/or PET) to identify directly stimulated fiber populations, and their corresponding cortical and/or subcortical regions altered by the stimulation, that are also associated with clinical benefit or side effects. If such an exercise was performed on enough patients it may be possible to statistically define the “actual target” of the stimulation. This information would be highly important in defining an optimal electrode implantation location, and motivate the use of patient-specific tractography in pre-operative surgical planning.
6. NETWORK COMPONENTS RELEVANT FOR DEPRESSION AND OCD
Depression and OCD are system-level disorders that cause a dysregulation of activity in cortico-striato-thalamocortical (CSTC) circuits (4, 8, 22, 63–66). Depression symptoms are associated with abnormal activity in the ventral striatum as well as orbitofrontal and Cg25 cortices (4). Normalization of activity in local (e.g., Cg25) and remote (e.g., prefrontal cortex) areas of the brain following successful DBS treatment suggest that the antidepressant benefits of DBS are correlated with reversal of the abnormalities found at baseline (4, 14). Similarly, increased symptomatic OCD episodes have been shown to be highly correlated with increased neuronal activity in the caudate nucleus, putamen, globus pallidus, inferior frontal gyrus, posterior cingulate gyrus, thalamus, and hippocampus (67–69). DBS of the VAIC, associated with therapeutic benefits for OCD, modulates activity in the dorsal striatum, Cg25, ventral globus pallidus, medial orbitofrontal cortex, and thalamus (5). These functional imaging results from OCD and depression make a clear case for DBS affecting multiple brain regions and show that stimulating a single region can generate complex changes throughout the interconnected network.
It has been hypothesized that the pathogenesis of depression and OCD may be caused by a combination of genetic defects and environmental factors that reduce the effectiveness of synaptic transmission in serotonergic, noradrenergic and dopaminergic pathways (70, 71). Antidepressant drugs, effective in treating both depression and OCD, increase the amount of neurotransmitter exposed to post-synaptic neurons by delaying neurotransmitter degradation (72) and/or reuptake (73, 74), slowly increasing serotonin receptor sensitivity and therefore the effectiveness of serotonergic synapses (71, 75). Thus, it is possible that DBS provides its therapeutic effects by manipulating serotonergic pathways. Serotonin receptors and transporters implicated in depression and OCD are highly expressed in the ventral striatum (76–78) while striatal activity is lower in OCD patients than in healthy individuals (79). The VAIC has fibers of passage connecting the subgenual cingulate and orbitofrontal cortices with striatum (Figure 2), and it is possible that stimulation of those fibers is modulating serotonin levels.
It has also been speculated that dysfunction of the nucleus accumbens, resulting in a disability to adequately modulate circuitry in the amygdala, basal ganglia, and prefrontal cortex, could be the origin of anxiety-disorders, depression, and OCD (2). The high stimulation amplitudes (6.5–10.5V) required to achieve therapeutic benefits when stimulating the VAIC suggest that the observed outcomes might have been the result of current spread to other nuclei (14, 29). Furthermore, lesions to the ventro-caudal part of the internal capsule, which are typically required for successful capsulotomies, are likely to include the nucleus accumbens (2). In turn, Greenberg et al. (1) concluded that the stimulating contacts closest to the nucleus accumbens might provide better therapeutic effects. Sturm et al. (2) confirmed significant symptom improvements with unilateral high-frequency stimulation of the right nucleus accumbens, indicating a major role for this nucleus as a central relay between the amygdaloid complex, basal ganglia, mediodorsal thalamus and the prefrontal cortex.
It is well known that the amygdaloid complex, especially the lateral nucleus, is involved in anxiety and fear reactions (80, 81). Pathological information flow from the lateral amygdaloid nucleus can be propagated to baso-lateral and central amygdaloid nuclei before converging in the accumbens via both, the ventral amygdalofugal pathway and the extended amygdale (82, 83). The main efferents of the accumbens innervate the pallidum, striatum, mediodorsal thalamus, prefrontal and cingulate cortices, as well as mesolimbic dopaminergic areas. The nucleus accumbens could thus represent a central location for impulse-propagation from the amygdaloid complex to the basal ganglia, mediodorsal thalamus and prefrontal cortex, all of which are implicated in the pathophysiology of OCD (84). Inputs from the amygdaloid complex to the nucleus accumbens “gate” both fronto-striatal and hippocampo-striatal circuitry (2, 85–87). Thus, disruption of pathological impulse-flow by means of chronic high-frequency stimulation of the nucleus accumbens might explain the therapeutic benefits of DBS in both depression and OCD patients.
A multi-circuit model of OCD (Figure 3) has been defined to help develop a rationale for surgical intervention (8). The model emphasizes the functional interconnections between the orbitofrontal and anterior cingulate cortices, medial, dorsomedial, and anterior thalamic nuclei, Papez circuit, anterior cingulate cortex, and nucleus accumbens (8). This model suggests that obsessive-compulsive symptoms appear when striato-pallido-thalamic activity is abnormally decreased or when orbitofronto-thalamic activity is abnormally increased. The OCD model is characterized by three main building blocks. The first block involves an excitatory glutamatergic positive feedback loop between the orbital cortex, the prefrontal cortex, and the dorso-medial thalamic nucleus, passing through the anterior limb of the internal capsule. The second block is an inhibitory GABAergic loop between the orbital and prefrontal cortices, the ventral caudate, the dorsomedial pallidum, and the anterior, dorsomedial and intralaminar thalamic nuclei. This component is thought to mediate the excitatory orbito-frontal-thalamic loop. The third block is a loop between the limbic system and the circuit of Papez (from the hippocampus to the mammillary body by means of the fornix, and onto the anterior thalamic nuclei). This model contains many projections from the anterior cingulate cortex to the nucleus accumbens in the striatum. According to this model, OCD symptoms could occur when striato-pallido-thalamic activity decreases and fails to modulate the activity in the positive fronto-thalamic loop (79).
Figure 3.

OCD model. The first component involves an excitatory and reciprocal positive feedback corticothalamic loop, in which the orbitofrontal and prefrontal cortices project onto the dorsomedial thalamus via the anterior limb of the internal capsule. The corticothalamic projection is mediated primarily by glutamate and aspartate, and the reciprocal thalamocortical projection is glutamate-mediated. The second component involves the orbitofrontal cortex, the ventral caudate, the dorsomedial pallidum, and the anterior, dorsomedial, and intralaminar nuclei of the thalamus. This component is thought to serve as a modulator for the orbitofrontal-thalamic loop trough inhibition from the dorsomedial pallidum to the thalamus by means of gamma-aminobutyric acid. This component is also thought to involve inhibitory serotonergic projections from the dorsal raphe nuclei of the midbrain to the ventral striatum. The third component involves a pathway between the limbic system and the circuit of Papez. This pathway projects from the hippocampal formation to the mammillary body by means of the fornix and continues on to the anterior thalamic nuclei and finally onto the cingulate gyrus. Reproduced with permission from (8).
An analogous model of depression (Figure 4) implicates failure to coordinate CSTC (including amygdale, hippocampus, and hypothalamic-pituitary axis) interactions as a result of overactive paralimbic regions and hypoactive dorsal cortical sites (8, 88). This model proposes that depressive behaviors are modulated by explicit interacting regions. Depression is thus not simply a dysfunction of any single one of these regions, but a failure in coordination of interactions between brain nuclei in these regions (88). This depression model involves dorsal (attention-cognition), ventral (vegetative-circadian), and rostral (regulatory) compartments (8, 67, 88). The dorsal compartment mediates the cognitive aspects of negative emotion. It involves the premotor and prefrontal cortices, as well as the dorsal segment of the anterior cingulate cortex. The ventral compartment involves the Cg25, as well as the insula and orbitofrontal cortex. This compartment is known to mediate circadian and vegetative aspects of depression. The rostral compartment involves the pregenual anterior cingulate cortex, the amygdala, and the hypothalamic pituitary axis. It is thought to regulate the overall network activity by facilitating the interaction between the dorsal and ventral compartments. Metabolism in this region may predict antidepressant response in acutely depressed patients (88).
Figure 4.

Depression model. The dorsal compartment is principally involved with the motor and cognitive aspects of depression. Projections to the dorsal striatum from prefrontal, dorsal anterior cingulate, and premotor cortices continue on to the thalamus by means of its projections from the dorsomedial portion of the pallidum. Thalamocortical projections close the loop. The ventral compartment, involved with affective aspects of depression, is composed of a closed loop involving paralimbic and subcortical components as well as regions of the brainstem. Specifically, the subgenual anterior cingulate (Cg25), orbitofrontal and insular cortices, project onto the ventral striatum, medial/rostral pallidum, and finally to the anterior and dorsomedial thalamus. The ventral compartment communicates with regions of the ventral compartment through the rostral and dorsal anterior cingulate, caudate-putamen, mediodorsal thalamus, and posterior cingulate. The modulatory compartment is thought to regulate relative ventral-dorsal component activity. It consists of the pregenual anterior cingulate, the amygdala, and the hypothalamic-pituitary axis. The amygdala drives activity in the ventral compartment and the pregenual anterior cingulate sends inhibitory projections to both compartments. The hippocampus, in turn, modulates activity in hypothalamic axis. Thus, reciprocal inhibitory connections between dorsal and ventral compartments combined with amygdala hyperactivity and abnormal hippocampal activity could lead to depression symptoms. Reproduced with permission from (8).
The models described above show that neuropsychiatric disorders appear to be rooted in the dysfunction of highly complex networks. Development of schematic diagrams of the pertinent brain regions associated with the disease processes are of fundamental importance in designing intervention strategies. However, analogous to movement disorders, it must be recognized that the actual brain networks are complex dynamical systems, and modulation of one node in the network can cause cascades of nonintuitive changes throughout the entire network on multiple time scales. Thus, DBS provides a unique and exciting opportunity to expand our understanding of the networks involved in neuropsychiatric disorders.
7. NETWORK MECHANISMS OF DBS IN DEPRESSION AND OCD
Although neural responses to electrical stimulation have been well characterized for single cells over short time scales, there is currently little understanding of the cellular mechanisms responsible for network responses to DBS in neuropsychiatric disease. In general, the slow progression of the therapeutic effects of stimulation (e.g., only limited improvements in patient mood and anxiety are evident with acute DBS) suggests that acute disruption of pathological network activity may not be the principal mechanism of DBS for neuropsychiatric disease (1, 2, 4, 14). For the most part, improvement of depression and OCD symptoms requires months of chronic stimulation (1, 4, 89), resulting in progressive symptom improvement and normalization of metabolic activity (9). Sleep disturbances typically normalize within the first week of chronic DBS, while energy, interest, and psychomotor improvements occur within a few weeks. These are followed by increased interest and pleasure in social interactions, as well as improved ability in planning and task initiation and completion. Furthermore, progressive symptom worsening can be observed if stimulation is turned off. Interestingly, the metabolic changes observed with DBS (90) linger after chronic stimulation (4). These results suggest that DBS reproduces the therapeutic benefits of antidepressants by inducing long-term changes in network activity trough continuous excitation/inhibition of serotonergic and noradrenergic pathways (91).
Given the overlap in symptom control, the similar neural circuitry affected by the stimulation, and the progressive improvement in symptoms over long-term stimulation, it is possible that DBS has similar underlying therapeutic mechanisms in both OCD and depression. One logical hypothesis is that DBS for neuropsychiatric disease has acute (fast) and chronic (delayed) therapeutic components. Acute improvements would most likely be the result of an immediate disruption of pathologic activity in serotonergic and noradrenergic circuits, whereas chronic improvements would be the result of long-term changes in synaptic effectiveness and/or synaptic connectivity (92).
In addition to acute and chronic changes, DBS may induce local and remote effects (93). PET studies have shown that successful treatment of both depression and OCD are associated with normalization of abnormal (hyper/hypo) basal metabolism and induced metabolic changes in remote (previously-normal) regions of the brain (3). Specific brain functions can be associated with distinct regions of the brain or with highly parallel, reciprocal, and overlapping processing networks, where the function of each neuronal group is determined by its dominant inputs (94, 95). By altering these inputs as a consequence of disease and/or slowly-developing brain lesions, large-scale reorganization of the brain (i.e., not confined to changes within the lesion area) can be induced (96, 97). This type of cerebral reorganization has been shown to be proportional to the progression of neuropsychiatric disease (94). Diaschisis has been described in the basal ganglia and internal capsule, suggesting that pathological activity within cerebral loops is a possible mechanism for the remote metabolic abnormalities observed in neuropsychiatric disease (4, 98, 99). Although the correlation between metabolic pattern changes and recovery is uncertain, it is likely that modulation of metabolic activity through cerebral loops is responsible for some (if not all) recovery. However, it remains unclear how this DBS-induced modulation generates chronic therapeutic effects.
Acute changes in activation levels in remote areas could occur as an indirect consequence of trans-synaptic effects or direct activation of projection neurons (100). In depression, acute symptom improvements are consistent with acute deactivation of hyperactive Cg25 and areas monosynaptically connected to it (4). Mayberg et al. (4) suggested that suppression of abnormally-elevated baseline activity in Cg25 could result from DBS-induced activation of inhibitory GABAergic afferents and/or high frequency stimulation induced synaptic or metabolic failure. However, it is also possible that stimulation directly affected the network dynamics of those cerebral loops passing through Cg25 in ways that cannot be simply understood with block diagram network models (100, 101).
Acute disruption of pathological activity can be achieved by inducing changes in neuronal membrane excitability, imbalances between excitatory and inhibitory inputs (e.g., as a result of changes in tonic inhibition), and/or short/long-term potentiation/depression (100, 102). Conversely, anatomical changes (e.g., formation of new synapses, growth of new connections into different regions of the brain) and synaptic potentiation/depression may produce changes in brain circuitry associated with delayed therapeutic benefits. Several activity-dependent mechanisms, which depend on prolonged stimulation, may cooperate to achieve synaptic plasticity that could underlie the therapeutic effects of DBS. Thus, long-term benefits may be the result of global changes in neural network properties (i.e., synaptic plasticity).
Below is a hypothetical example of how DBS induced activity could result in long-term network changes. Supra-threshold high frequency DBS continuously activates a large number of axons and increases glutamatergic and serotonergic exposure in the striatum (Figures 1 and 2). Activation of serotonin receptors results in protein kinase phosphorylation and activation of second messenger systems that regulate gene transcription (103, 104). Activated genes induce long-term potentiation (LTP) as well as the growth of synaptic connections (105). The high-frequency stimulation also induces spine enlargement and increases the number of glutamate receptors (106). These synaptic changes then impact the excitability of the striatum and alter its integration of cortical inputs. In parallel, analogous changes in cortical regions could also be occurring (100). The sum result would be a substantial alteration of the CSTC network dynamics, alleviating the pathological activity patterns. Clearly this generalized hypothesis is untested, speculative, and missing important details (i.e. how CSTC network dynamics actually work), but given our current understanding of the system, we believe it is plausible.
8. FUTURE DIRECTIONS
The preliminary clinical effectiveness of DBS for intractable neuropsychiatric disorders makes it a promising therapeutic alternative for a large number of patients (4, 8). However, our limited understanding of both the pathophysiology of neuropsychiatric disorders and the effects of DBS on the nervous system, make wide-scale use of DBS technology a tenuous proposition from a scientific perspective. However, clinical outcomes will drive clinical adoption of this technology, and similar to DBS for movement disorders, scientists will have to play catch up.
Although a large number of DBS procedures have been performed worldwide, only a few have been intended for treatment of neuropsychiatric disorders (10), and as such, study populations have been small. The limited documentation of parameter adjustments utilized in these studies makes it difficult to identify the direct effects induced by various device settings. As such, the optimal target and stimulation parameters remain unclear. Interestingly, data suggests that stimulation of the nucleus accumbens may play a critical role in normalization of activity and increased therapeutic benefits in both depression and OCD (9, 65, 69). In addition, it is also possible that optimal therapeutic benefits might be achieved by stimulating a combination of targets within the network, potentially allowing a more complex manipulation of the entire neural circuitry involved in neuropsychiatric disorders. However, before embarking on speculative analyses of alternative targets, it may be more prudent to perform larger double blinded studies, with longer follow-up periods to clearly demonstrate the efficacy of DBS with currently accepted stimulation paradigms.
Scientifically, an almost endless number of questions remain to be addressed. For example, a fascinating aspect of DBS for neuropsychiatric disorders is the relatively slow progression of therapeutic benefits, even though acute effects can often be seen in the operating room. Another interesting question is why, if DBS replicates the effects of antidepressant drugs, has it been effective in treating patients where antidepressants have failed? These questions and many more will make for interesting discussion and debate as the scientific and clinical worlds converge to elucidate the therapeutic mechanisms of DBS for neuropsychiatric disorders.
Acknowledgments
This work was supported by funding from the National Institutes of Health (NS050449, NS059736). The authors also thank Susumu Mori for providing the diffusion tensor brain atlas data set and Christopher Maks for assistance in the tractography analysis.
Abbreviations
- DBS
Deep brain stimulation
- VAIC
ventral anterior internal capsule
- Cg25
subgenual anterior cingulate
- Cg25WM
subgenual cingulate white matter
- OCD
obsessive compulsive disorder
- DTI
diffusion-tensor imaging
- ROI
region of interest
- CSTC
cortico-striato-thalamocortical
- LTP
long-term potentiation
References
- 1.Greenberg BD, Malone DA, Friehs GM, Rezai AR, Kubu CS, Malloy PF, Salloway SP, Okun MS, Goodman WK, Rasmussen SA. Three-year outcomes in deep brain stimulation for highly resistant obsessive-compulsive disorder. Neuropsychopharmacology. 2006;31(11):2384–93. doi: 10.1038/sj.npp.1301165. [DOI] [PubMed] [Google Scholar]
- 2.Sturm V, Lenartz D, Koulousakis A, Treuer H, Herholz K, Klein JC, Klosterkotter J. The nucleus accumbens: a target for deep brain stimulation in obsessive-compulsive- and anxiety-disorders. J Chem Neuroanat. 2003;26(4):293–9. doi: 10.1016/j.jchemneu.2003.09.003. [DOI] [PubMed] [Google Scholar]
- 3.Mayberg HS, Brannan SK, Tekell JL, Silva JA, Mahurin RK, McGinnis S, Jerabek PA. Regional metabolic effects of fluoxetine in major depression: serial changes and relationship to clinical response. Biol Psychiatry. 2000;48(8):830–43. doi: 10.1016/s0006-3223(00)01036-2. [DOI] [PubMed] [Google Scholar]
- 4.Mayberg HS, Lozano AM, Voon V, McNeely HE, Seminowicz D, Hamani C, Schwalb JM, Kennedy SH. Deep brain stimulation for treatment-resistant depression. Neuron. 2005;45(5):651–60. doi: 10.1016/j.neuron.2005.02.014. [DOI] [PubMed] [Google Scholar]
- 5.Rauch SL, Dougherty DD, Malone D, Rezai A, Friehs G, Fischman AJ, Alpert NM, Haber SN, Stypulkowski PH, Rise MT, Rasmussen SA, Greenberg BD. A functional neuroimaging investigation of deep brain stimulation in patients with obsessive-compulsive disorder. J Neurosurg. 2006;104(4):558–65. doi: 10.3171/jns.2006.104.4.558. [DOI] [PubMed] [Google Scholar]
- 6.Aouizerate B. Bipolar disorder and so called neurotic symptoms. Encephale. 2006;32(Pt 2):S33–7. doi: 10.1016/s0013-7006(06)76176-0. [DOI] [PubMed] [Google Scholar]
- 7.Gabriels L, Cosyns P, Nuttin B, Demeulemeester H, Gybels J. Deep brain stimulation for treatment-refractory obsessive-compulsive disorder: psychopathological and neuropsychological outcome in three cases. Acta Psychiatr Scand. 2003;107(4):275–82. [PubMed] [Google Scholar]
- 8.Kopell BH, Greenberg B, Rezai AR. Deep brain stimulation for psychiatric disorders. J Clin Neurophysiol. 2004;21(1):51–67. doi: 10.1097/00004691-200401000-00007. [DOI] [PubMed] [Google Scholar]
- 9.Aouizerate B, Cuny E, Martin-Guehl C, Guehl D, Amieva H, Benazzouz A, Fabrigoule C, Allard M, Rougier A, Bioulac B, Tignol J, Burbaud P. Deep brain stimulation of the ventral caudate nucleus in the treatment of obsessive-compulsive disorder and major depression. Case report. J Neurosurg. 2004;101(4):682–6. doi: 10.3171/jns.2004.101.4.0682. [DOI] [PubMed] [Google Scholar]
- 10.Cosgrove GR. Deep brain stimulation and psychosurgery. J Neurosurg. 2004;101(4):574–5. doi: 10.3171/jns.2004.101.4.0574. discussion 575–6. [DOI] [PubMed] [Google Scholar]
- 11.Sakas DE, I, Panourias G. Rostral cingulate gyrus: A putative target for deep brain stimulation in treatment-refractory depression. Med Hypotheses. 2006;66(3):491–4. doi: 10.1016/j.mehy.2005.07.036. [DOI] [PubMed] [Google Scholar]
- 12.Benabid AL, Pollak P, Gervason C, Hoffmann D, Gao DM, Hommel M, Perret JE, de Rougemont J. Long-term suppression of tremor by chronic stimulation of the ventral intermediate thalamic nucleus. Lancet. 1991;337(8738):403–6. doi: 10.1016/0140-6736(91)91175-t. [DOI] [PubMed] [Google Scholar]
- 13.Benabid AL, Pollak P, Louveau A, Henry S, de Rougemont J. Combined (thalamotomy and stimulation) stereotactic surgery of the VIM thalamic nucleus for bilateral Parkinson disease. Appl Neurophysiol. 1987;50(1–6):344–6. doi: 10.1159/000100803. [DOI] [PubMed] [Google Scholar]
- 14.Abelson JL, Curtis GC, Sagher O, Albucher RC, Harrigan M, Taylor SF, Martis B, Giordani B. Deep brain stimulation for refractory obsessive-compulsive disorder. Biol Psychiatry. 2005;57(5):510–6. doi: 10.1016/j.biopsych.2004.11.042. [DOI] [PubMed] [Google Scholar]
- 15.Gross RE, Lozano AM. Advances in neurostimulation for movement disorders. Neurol Res. 2000;22(3):247–58. doi: 10.1080/01616412.2000.11740667. [DOI] [PubMed] [Google Scholar]
- 16.Lipsman N, Neimat JS, Lozano AM. Deep brain stimulation for treatment-refractory obsessive-compulsive disorder: the search for a valid target. Neurosurgery. 2007;61(1):1–11. doi: 10.1227/01.neu.0000279719.75403.f7. discussion 11–3. [DOI] [PubMed] [Google Scholar]
- 17.Benabid AL, Pollak P, Gao D, Hoffmann D, Limousin P, Gay E, Payen I, Benazzouz A. Chronic electrical stimulation of the ventralis intermedius nucleus of the thalamus as a treatment of movement disorders. J Neurosurg. 1996;84(2):203–14. doi: 10.3171/jns.1996.84.2.0203. [DOI] [PubMed] [Google Scholar]
- 18.Obeso JA, Olanow CW, Rodriguez-Oroz MC, Krack P, Kumar R, Lang AE. Deep-brain stimulation of the subthalamic nucleus or the pars interna of the globus pallidus in Parkinson’s disease. N Engl J Med. 2001;345(13):956–63. doi: 10.1056/NEJMoa000827. [DOI] [PubMed] [Google Scholar]
- 19.Vidailhet M, Vercueil L, Houeto JL, Krystkowiak P, Benabid AL, Cornu P, Lagrange C, Tezenas du Montcel S, Dormont D, Grand S, Blond S, Detante O, Pillon B, Ardouin C, Agid Y, Destee A, Pollak P. Bilateral deep-brain stimulation of the globus pallidus in primary generalized dystonia. N Engl J Med. 2005;352(5):459–67. doi: 10.1056/NEJMoa042187. [DOI] [PubMed] [Google Scholar]
- 20.Giacobbe P, Kennedy SH. Deep brain stimulation for treatment-resistant depression: a psychiatric perspective. Curr Psychiatry Rep. 2006;8(6):437–44. doi: 10.1007/s11920-006-0048-5. [DOI] [PubMed] [Google Scholar]
- 21.Greenberg BD, Price LH, Rauch SL, Friehs G, Noren G, Malone D, Carpenter LL, Rezai AR, Rasmussen SA. Neurosurgery for intractable obsessive-compulsive disorder and depression: critical issues. Neurosurg Clin N Am. 2003;14(2):199–212. doi: 10.1016/s1042-3680(03)00005-6. [DOI] [PubMed] [Google Scholar]
- 22.Greenberg BD, Rezai AR. Mechanisms and the current state of deep brain stimulation in neuropsychiatry. CNS Spectr. 2003;8(7):522–6. doi: 10.1017/s109285290001899x. [DOI] [PubMed] [Google Scholar]
- 23.Jimenez F, Velasco F, Salin-Pascual R, Hernandez JA, Velasco M, Criales JL, Nicolini H. A patient with a resistant major depression disorder treated with deep brain stimulation in the inferior thalamic peduncle. Neurosurgery. 2005;57(3):585–93. doi: 10.1227/01.neu.0000170434.44335.19. discussion 585–93. [DOI] [PubMed] [Google Scholar]
- 24.Kuhn J, Huff W, Lee SH, Lenartz D, Sturm V, Klosterkotter J. Deep brain stimulation in the treatment of psychiatric disorders. Fortschr Neurol Psychiatr. 2007;75(8):447–57. [Google Scholar]
- 25.Sartorius A, Henn FA. Deep brain stimulation of the lateral habenula in treatment resistant major depression. Med Hypotheses. 2007 doi: 10.1016/j.mehy.2007.03.021. [DOI] [PubMed] [Google Scholar]
- 26.Saxena S, Brody AL, Ho ML, Alborzian S, Ho MK, Maidment KM, Huang SC, Wu HM, Au SC, Baxter LR., Jr Cerebral metabolism in major depression and obsessive-compulsive disorder occurring separately and concurrently. Biol Psychiatry. 2001;50(3):159–70. doi: 10.1016/s0006-3223(01)01123-4. [DOI] [PubMed] [Google Scholar]
- 27.Nuttin B, Cosyns P, Demeulemeester H, Gybels J, Meyerson B. Electrical stimulation in anterior limbs of internal capsules in patients with obsessive-compulsive disorder. Lancet. 1999;354(9189):1526. doi: 10.1016/S0140-6736(99)02376-4. [DOI] [PubMed] [Google Scholar]
- 28.Nuttin BJ, Gabriels L, van Kuyck K, Cosyns P. Electrical stimulation of the anterior limbs of the internal capsules in patients with severe obsessive-compulsive disorder: anecdotal reports. Neurosurg Clin N Am. 2003;14(2):267–74. doi: 10.1016/s1042-3680(02)00117-1. [DOI] [PubMed] [Google Scholar]
- 29.Nuttin BJ, Gabriels LA, Cosyns PR, Meyerson BA, Andreewitch S, Sunaert SG, Maes AF, Dupont PJ, Gybels JM, Gielen F, Demeulemeester HG. Long-term electrical capsular stimulation in patients with obsessive-compulsive disorder. Neurosurgery. 2003;52(6):1263–72. doi: 10.1227/01.neu.0000064565.49299.9a. discussion 1272–4. [DOI] [PubMed] [Google Scholar]
- 30.Butson CR, Cooper SE, Henderson JM, McIntyre CC. Patient-specific analysis of the volume of tissue activated during deep brain stimulation. Neuroimage. 2007;34(2):661–70. doi: 10.1016/j.neuroimage.2006.09.034. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.McIntyre CC, Mori S, Sherman DL, Thakor NV, Vitek JL. Electric field and stimulating influence generated by deep brain stimulation of the subthalamic nucleus. Clin Neurophysiol. 2004;115(3):589–95. doi: 10.1016/j.clinph.2003.10.033. [DOI] [PubMed] [Google Scholar]
- 32.McNeal DR. Analysis of a model for excitation of myelinated nerve. IEEE Trans Biomed Eng. 1976;23(4):329–37. doi: 10.1109/tbme.1976.324593. [DOI] [PubMed] [Google Scholar]
- 33.Rattay F. Analysis of models for external stimulation of axons. IEEE Trans Biomed Eng. 1986;33(10):974–7. doi: 10.1109/TBME.1986.325670. [DOI] [PubMed] [Google Scholar]
- 34.McIntyre CC, Grill WM. Excitation of central nervous system neurons by nonuniform electric fields. Biophys J. 1999;76(2):878–88. doi: 10.1016/S0006-3495(99)77251-6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Rattay F. The basic mechanism for the electrical stimulation of the nervous system. Neuroscience. 1999;89(2):335–46. doi: 10.1016/s0306-4522(98)00330-3. [DOI] [PubMed] [Google Scholar]
- 36.Grill WM, McIntyre CC. Extracellular excitation of central neurons: implications for the mechanisms of deep brain stimulation. Thalamus & Related Systems. 2001;1(3):269–277. [Google Scholar]
- 37.McIntyre CC, Grill WM, Sherman DL, Thakor NV. Cellular effects of deep brain stimulation: model-based analysis of activation and inhibition. J Neurophysiol. 2004;91(4):1457–69. doi: 10.1152/jn.00989.2003. [DOI] [PubMed] [Google Scholar]
- 38.Miocinovic S, Parent M, Butson CR, Hahn PJ, Russo GS, Vitek JL, McIntyre CC. Computational analysis of subthalamic nucleus and lenticular fasciculus activation during therapeutic deep brain stimulation. J Neurophysiol. 2006;96(3):1569–80. doi: 10.1152/jn.00305.2006. [DOI] [PubMed] [Google Scholar]
- 39.Anderson ME, Postupna N, Ruffo M. Effects of high-frequency stimulation in the internal globus pallidus on the activity of thalamic neurons in the awake monkey. J Neurophysiol. 2003;89(2):1150–60. doi: 10.1152/jn.00475.2002. [DOI] [PubMed] [Google Scholar]
- 40.Dostrovsky JO, Lozano AM. Mechanisms of deep brain stimulation. Mov Disord. 2002;17(Suppl 3):S63–8. doi: 10.1002/mds.10143. [DOI] [PubMed] [Google Scholar]
- 41.Hashimoto T, Elder CM, Okun MS, Patrick SK, Vitek JL. Stimulation of the subthalamic nucleus changes the firing pattern of pallidal neurons. J Neurosci. 2003;23(5):1916–23. doi: 10.1523/JNEUROSCI.23-05-01916.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Boulet S, Lacombe E, Carcenac C, Feuerstein C, Sgambato-Faure V, Poupard A, Savasta M. Subthalamic stimulation-induced forelimb dyskinesias are linked to an increase in glutamate levels in the substantia nigra pars reticulata. J Neurosci. 2006;26(42):10768–76. doi: 10.1523/JNEUROSCI.3065-06.2006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Windels F, Bruet N, Poupard A, Feuerstein C, Bertrand A, Savasta M. Influence of the frequency parameter on extracellular glutamate and gamma-aminobutyric acid in substantia nigra and globus pallidus during electrical stimulation of subthalamic nucleus in rats. J Neurosci Res. 2003;72(2):259–67. doi: 10.1002/jnr.10577. [DOI] [PubMed] [Google Scholar]
- 44.Jech R, Urgosik D, Tintera J, Nebuzelsky A, Krasensky J, Liscak R, Roth J, Ruzicka E. Functional magnetic resonance imaging during deep brain stimulation: a pilot study in four patients with Parkinson’s disease. Mov Disord. 2001;16(6):1126–32. doi: 10.1002/mds.1217. [DOI] [PubMed] [Google Scholar]
- 45.Perlmutter JS, Mink JW, Bastian AJ, Zackowski K, Hershey T, Miyawaki E, Koller W, Videen TO. Blood flow responses to deep brain stimulation of thalamus. Neurology. 2002;58(9):1388–94. doi: 10.1212/wnl.58.9.1388. [DOI] [PubMed] [Google Scholar]
- 46.Rezai AR, Lozano AM, Crawley AP, Joy ML, Davis KD, Kwan CL, Dostrovsky JO, Tasker RR, Mikulis DJ. Thalamic stimulation and functional magnetic resonance imaging: localization of cortical and subcortical activation with implanted electrodes. Technical note. J Neurosurg. 1999;90(3):583–90. doi: 10.3171/jns.1999.90.3.0583. [DOI] [PubMed] [Google Scholar]
- 47.Dostrovsky JO, Levy R, Wu JP, Hutchison WD, Tasker RR, Lozano AM. Microstimulation-induced inhibition of neuronal firing in human globus pallidus. J Neurophysiol. 2000;84(1):570–4. doi: 10.1152/jn.2000.84.1.570. [DOI] [PubMed] [Google Scholar]
- 48.Beurrier C, Bioulac B, Audin J, Hammond C. High-frequency stimulation produces a transient blockade of voltage-gated currents in subthalamic neurons. J Neurophysiol. 2001;85(4):1351–6. doi: 10.1152/jn.2001.85.4.1351. [DOI] [PubMed] [Google Scholar]
- 49.Maurice N, Thierry AM, Glowinski J, Deniau JM. Spontaneous and evoked activity of substantia nigra pars reticulata neurons during high-frequency stimulation of the subthalamic nucleus. J Neurosci. 2003;23(30):9929–36. doi: 10.1523/JNEUROSCI.23-30-09929.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.McIntyre CC, Grill WM. Extracellular stimulation of central neurons: influence of stimulus waveform and frequency on neuronal output. J Neurophysiol. 2002;88(4):1592–604. doi: 10.1152/jn.2002.88.4.1592. [DOI] [PubMed] [Google Scholar]
- 51.Bhadra N, Kilgore KL. High-frequency electrical conduction block of mammalian peripheral motor nerve. Muscle Nerve. 2005;32(6):782–90. doi: 10.1002/mus.20428. [DOI] [PubMed] [Google Scholar]
- 52.Jensen AL, Durand DM. Suppression of axonal conduction by sinusoidal stimulation in rat hippocampus in vitro. J Neural Eng. 2007;4(2):1–16. doi: 10.1088/1741-2560/4/2/001. [DOI] [PubMed] [Google Scholar]
- 53.Basser PJ, Mattiello J, LeBihan D. MR diffusion tensor spectroscopy and imaging. Biophys J. 1994;66(1):259–67. doi: 10.1016/S0006-3495(94)80775-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.D’Arceuil HE, Westmoreland S, de Crespigny AJ. An approach to high resolution diffusion tensor imaging in fixed primate brain. Neuroimage. 2007;35(2):553–65. doi: 10.1016/j.neuroimage.2006.12.028. [DOI] [PubMed] [Google Scholar]
- 55.Mori S, Crain BJ, Chacko VP, van Zijl PC. Three-dimensional tracking of axonal projections in the brain by magnetic resonance imaging. Ann Neurol. 1999;45(2):265–9. doi: 10.1002/1531-8249(199902)45:2<265::aid-ana21>3.0.co;2-3. [DOI] [PubMed] [Google Scholar]
- 56.Wakana S, Jiang H, Nagae-Poetscher LM, van Zijl PC, Mori S. Fiber tract-based atlas of human white matter anatomy. Radiology. 2004;230(1):77–87. doi: 10.1148/radiol.2301021640. [DOI] [PubMed] [Google Scholar]
- 57.Le Bihan D, Mangin JF, Poupon C, Clark CA, Pappata S, Molko N, Chabriat H. Diffusion tensor imaging: concepts and applications. J Magn Reson Imaging. 2001;13(4):534–46. doi: 10.1002/jmri.1076. [DOI] [PubMed] [Google Scholar]
- 58.Lori NF, Akbudak E, Shimony JS, Cull TS, Snyder AZ, Guillory RK, Conturo TE. Diffusion tensor fiber tracking of human brain connectivity: aquisition methods, reliability analysis and biological results. NMR Biomed. 2002;15(7–8):494–515. doi: 10.1002/nbm.779. [DOI] [PubMed] [Google Scholar]
- 59.Nimsky C, Ganslandt O, Merhof D, Sorensen AG, Fahlbusch R. Intraoperative visualization of the pyramidal tract by diffusion-tensor-imaging-based fiber tracking. Neuroimage. 2006;30(4):1219–29. doi: 10.1016/j.neuroimage.2005.11.001. [DOI] [PubMed] [Google Scholar]
- 60.Tournier JD, Calamante F, King MD, Gadian DG, Connelly A. Limitations and requirements of diffusion tensor fiber tracking: an assessment using simulations. Magn Reson Med. 2002;47(4):701–8. doi: 10.1002/mrm.10116. [DOI] [PubMed] [Google Scholar]
- 61.Johansen-Berg H, Gutman DA, Behrens TE, Matthews PM, Rushworth MF, Katz E, Lozano AM, Mayberg HS. Anatomical Connectivity of the Subgenual Cingulate Region Targeted with Deep Brain Stimulation for Treatment-Resistant Depression. Cereb Cortex. 2007 doi: 10.1093/cercor/bhm167. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62.Chaturvedi A, Butson CR, Cooper SE, McIntyre CC. Subthalamic nucleus deep brain stimulation: accurate axonal threshold prediction with diffusion tensor based electric field models. Conf Proc IEEE Eng Med Biol Soc; 2006. pp. 1240–3. [DOI] [PubMed] [Google Scholar]
- 63.Greenberg BD, Altemus M, Murphy DL. The role of neurotransmitters and neurohormones in obsessive-compulsive disorder. International Review of Psychiatry. 1997;9:31–44. [Google Scholar]
- 64.Modell JG, Mountz JM, Curtis GC, Greden JF. Neurophysiologic dysfunction in basal ganglia/limbic striatal and thalamocortical circuits as a pathogenetic mechanism of obsessive-compulsive disorder. J Neuropsychiatry Clin Neurosci. 1989;1(1):27–36. doi: 10.1176/jnp.1.1.27. [DOI] [PubMed] [Google Scholar]
- 65.Rauch SL. Neuroimaging and neurocircuitry models disorders. Neurosurg Clin N Am. 2003;14(2):213–23. vii–viii. doi: 10.1016/s1042-3680(02)00114-6. [DOI] [PubMed] [Google Scholar]
- 66.Saxena S, Brody AL, Schwartz JM, Baxter LR. Neuroimaging and frontal-subcortical circuitry in obsessive-compulsive disorder. Br J Psychiatry Suppl. 1998;(35):26–37. [PubMed] [Google Scholar]
- 67.Mayberg HS. Depression. In: Mazziotta JC, Toga AW, Frackowiak RS, editors. Brain Mapping: The Disorders. Academic Press; 2000. [Google Scholar]
- 68.Mayberg HS. Neurobiology of Anxiety. In: Mazziotta JC, Toga AW, Frackowiak RS, editors. Brain Mapping: The Disorders. Academic Press; 2000. [Google Scholar]
- 69.Schlaepfer TE, Cohen MX, Frick C, Kosel M, Brodesser D, Axmacher N, Joe AY, Kreft M, Lenartz D, Sturm V. Deep Brain Stimulation to Reward Circuitry Alleviates Anhedonia in Refractory Major Depression. Neuropsychopharmacology. 2007 doi: 10.1038/sj.npp.1301408. [DOI] [PubMed] [Google Scholar]
- 70.Delcomyn F. Foundations of Neurobiology. W. H. Freeman and Company; New York: 1998. [Google Scholar]
- 71.Kandel ER. Disorders of mood: Depression, mania, and anxiety disorders. In: Kandel ER, Schwartz J, Jessell T, editors. Principles of Neural Science. Elsevier; 2000. [Google Scholar]
- 72.Lestra C, d’Amato T, Ghaemmaghami C, Perret-Liaudet A, Broyer M, Renaud B, Dalery J, Chamba G. Biological parameters in major depression: effects of paroxetine, viloxazine, moclobemide, and electroconvulsive therapy. Relation to early clinical outcome. Biol Psychiatry. 1998;44(4):274–80. doi: 10.1016/s0006-3223(97)00389-2. [DOI] [PubMed] [Google Scholar]
- 73.Matsumoto K, Puia G, Dong E, Pinna G. GABA(A) receptor neurotransmission dysfunction in a mouse model of social isolation-induced stress: possible insights into a non-serotonergic mechanism of action of SSRIs in mood and anxiety disorders. Stress. 2007;10(1):3–12. doi: 10.1080/10253890701200997. [DOI] [PubMed] [Google Scholar]
- 74.Saxena S, Brody AL, Ho ML, Zohrabi N, Maidment KM, Baxter LR., Jr Differential brain metabolic predictors of response to paroxetine in obsessive-compulsive disorder versus major depression. Am J Psychiatry. 2003;160(3):522–32. doi: 10.1176/appi.ajp.160.3.522. [DOI] [PubMed] [Google Scholar]
- 75.Insel TR, Winslow JT. Neurobiology of obsessive compulsive disorder. Psychiatr Clin North Am. 1992;15(4):813–24. [PubMed] [Google Scholar]
- 76.Camarena B, Rinetti G, Cruz C, Hernandez S, de la Fuente JR, Nicolini H. Association study of the serotonin transporter gene polymorphism in obsessive-compulsive disorder. Int J Neuropsychopharmacol. 2001;4(3):269–72. doi: 10.1017/S1461145701002516. [DOI] [PubMed] [Google Scholar]
- 77.Hoyer D, Pazos A, Probst A, Palacios JM. Serotonin receptors in the human brain. I. Characterization and autoradiographic localization of 5-HT1A recognition sites. Apparent absence of 5-HT1B recognition sites. Brain Res. 1986;376(1):85–96. doi: 10.1016/0006-8993(86)90902-9. [DOI] [PubMed] [Google Scholar]
- 78.Welch JM, Lu J, Rodriguiz RM, Trotta NC, Peca J, Ding JD, Feliciano C, Chen M, Adams JP, Luo J, Dudek SM, Weinberg RJ, Calakos N, Wetsel WC, Feng G. Cortico-striatal synaptic defects and OCD-like behaviours in Sapap3-mutant mice. Nature. 2007;448(7156):894–900. doi: 10.1038/nature06104. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 79.Haber SN, Fudge JL, McFarland NR. Striatonigrostriatal pathways in primates form an ascending spiral from the shell to the dorsolateral striatum. J Neurosci. 2000;20(6):2369–82. doi: 10.1523/JNEUROSCI.20-06-02369.2000. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 80.LeDoux JE. Emotion circuits in the brain. Annu Rev Neurosci. 2000;23:155–84. doi: 10.1146/annurev.neuro.23.1.155. [DOI] [PubMed] [Google Scholar]
- 81.Shumyatsky GP, Tsvetkov E, Malleret G, Vronskaya S, Hatton M, Hampton L, Battey JF, Dulac C, Kandel ER, Bolshakov VY. Identification of a signaling network in lateral nucleus of amygdala important for inhibiting memory specifically related to learned fear. Cell. 2002;111(6):905–18. doi: 10.1016/s0092-8674(02)01116-9. [DOI] [PubMed] [Google Scholar]
- 82.Alheid GF, Beltramino CA, De Olmos JS, Forbes MS, Swanson DJ, Heimer L. The neuronal organization of the supracapsular part of the stria terminalis in the rat: the dorsal component of the extended amygdala. Neuroscience. 1998;84(4):967–96. doi: 10.1016/s0306-4522(97)00560-5. [DOI] [PubMed] [Google Scholar]
- 83.de Olmos JS, Heimer L. The concepts of the ventral striatopallidal system and extended amygdala. Ann N Y Acad Sci. 1999;877:1–32. doi: 10.1111/j.1749-6632.1999.tb09258.x. [DOI] [PubMed] [Google Scholar]
- 84.Saxena S, Rauch SL. Functional neuroimaging and the neuroanatomy of obsessive-compulsive disorder. Psychiatr Clin North Am. 2000;23(3):563–86. doi: 10.1016/s0193-953x(05)70181-7. [DOI] [PubMed] [Google Scholar]
- 85.Grace AA. Cortical regulation of subcortical dopamine systems and its possible relevance to schizophrenia. J Neural Transm Gen Sect. 1993;91(2–3):111–34. doi: 10.1007/BF01245228. [DOI] [PubMed] [Google Scholar]
- 86.Mulder AB, Hodenpijl MG, Lopes da Silva FH. Electrophysiology of the hippocampal and amygdaloid projections to the nucleus accumbens of the rat: convergence, segregation, and interaction of inputs. J Neurosci. 1998;18(13):5095–102. doi: 10.1523/JNEUROSCI.18-13-05095.1998. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 87.Rosenkranz JA, Grace AA. Modulation of basolateral amygdala neuronal firing and afferent drive by dopamine receptor activation in vivo. J Neurosci. 1999;19(24):11027–39. doi: 10.1523/JNEUROSCI.19-24-11027.1999. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 88.Mayberg HS. Limbic-cortical dysregulation: a proposed model of depression. J Neuropsychiatry Clin Neurosci. 1997;9(3):471–81. doi: 10.1176/jnp.9.3.471. [DOI] [PubMed] [Google Scholar]
- 89.Cosyns P, Gabriels L, Nuttin B. Deep brain stimulation in treatment refractory obsessive compulsive disorder. Verh K Acad Geneeskd Belg. 2003;65(6):385–99. discussion 399–400. [PubMed] [Google Scholar]
- 90.Van Laere K, Nuttin B, Gabriels L, Dupont P, Rasmussen S, Greenberg BD, Cosyns P. Metabolic imaging of anterior capsular stimulation in refractory obsessive-compulsive disorder: a key role for the subgenual anterior cingulate and ventral striatum. J Nucl Med. 2006;47(5):740–7. [PubMed] [Google Scholar]
- 91.Vaidya VA, Duman RS. Depresssion--emerging insights from neurobiology. Br Med Bull. 2001;57:61–79. doi: 10.1093/bmb/57.1.61. [DOI] [PubMed] [Google Scholar]
- 92.Hallett M. Plasticity. In: Mazziotta JC, Toga AW, Frackowiak RS, editors. Brain Mapping: The Disorders. Academic Press; 2000. [Google Scholar]
- 93.Davis KD, Taub E, Houle S, Lang AE, Dostrovsky JO, Tasker RR, Lozano AM. Globus pallidus stimulation activates the cortical motor system during alleviation of parkinsonian symptoms. Nat Med. 1997;3(6):671–4. doi: 10.1038/nm0697-671. [DOI] [PubMed] [Google Scholar]
- 94.Chollet F, Weiller C. Recovery of Neurological Function. In: Mazziotta JC, Toga AW, Frackowiak RS, editors. Brain Mapping: The Disorders. Academic Press; 2000. [Google Scholar]
- 95.Donoghue JP. Plasticity of adult sensorimotor representations. Curr Opin Neurobiol. 1995;5(6):749–54. doi: 10.1016/0959-4388(95)80102-2. [DOI] [PubMed] [Google Scholar]
- 96.Seitz RJ, Huang Y, Knorr U, Tellmann L, Herzog H, Freund HJ. Large-scale plasticity of the human motor cortex. Neuroreport. 1995;6(5):742–4. doi: 10.1097/00001756-199503270-00009. [DOI] [PubMed] [Google Scholar]
- 97.Wunderlich G, Knorr U, Herzog H, Kiwit JC, Freund HJ, Seitz RJ. Precentral glioma location determines the displacement of cortical hand representation. Neurosurgery. 1998;42(1):18–26. doi: 10.1097/00006123-199801000-00005. discussion 26–7. [DOI] [PubMed] [Google Scholar]
- 98.Meneghetti G, Vorstrup S, Mickey B, Lindewald H, Lassen NA. Crossed cerebellar diaschisis in ischemic stroke: a study of regional cerebral blood flow by 133Xe inhalation and single photon emission computerized tomography. J Cereb Blood Flow Metab. 1984;4(2):235–40. doi: 10.1038/jcbfm.1984.32. [DOI] [PubMed] [Google Scholar]
- 99.Rougemont D, Baron JC, Lebrun-Grandie P, Bousser MG, Cabanis E, Laplane D. Cerebral blood flow and oxygen extraction in lacunar hemiplegia. Semi-quantitative study using oxygen 15 and emission tomography. Rev Neurol (Paris) 1983;139(4):277–82. [PubMed] [Google Scholar]
- 100.McCracken CB, Grace AA. High-frequency deep brain stimulation of the nucleus accumbens region suppresses neuronal activity and selectively modulates afferent drive in rat orbitofrontal cortex in vivo. J Neurosci. 2007;27(46):12601–10. doi: 10.1523/JNEUROSCI.3750-07.2007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 101.Montgomery EB., Jr Dynamically coupled, high-frequency reentrant, non-linear oscillators embedded in scale-free basal ganglia-thalamic-cortical networks mediating function and deep brain stimulation effects. Nonlinear Studies. 2004;11(3):385–421. [Google Scholar]
- 102.Ziemann U, Hallett M, Cohen LG. Mechanisms of deafferentation-induced plasticity in human motor cortex. J Neurosci. 1998;18(17):7000–7. doi: 10.1523/JNEUROSCI.18-17-07000.1998. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 103.Kandel ER. Cellular mechanisms of learning and the biological basis of individuality. In: Kandel ER, Schwartz J, Jessell T, editors. Principles of Neural Science. Elsevier; 2000. [Google Scholar]
- 104.Ward RP, Dorsa DM. Molecular and behavioral effects mediated by Gs-coupled adenosine A2a, but not serotonin 5-Ht4 or 5-Ht6 receptors following antipsychotic administration. Neuroscience. 1999;89(3):927–38. doi: 10.1016/s0306-4522(98)00364-9. [DOI] [PubMed] [Google Scholar]
- 105.Kupfermann I. Learning and Memory. In: Kandel ER, Schwartz J, Jessell T, editors. Principles of Neural Science. Elsevier; 1991. [Google Scholar]
- 106.Bennett MR. The concept of long term potentiation of transmission at synapses. Progress in Neurobiology. 2000;60(2):109–137. doi: 10.1016/s0301-0082(99)00006-4. [DOI] [PubMed] [Google Scholar]