

Abstract
Epigenetic mechanisms (Box 1) are considered to play major gene-regulatory roles in development, differentiation and disease. However, the relative importance of epigenetics in defining the mammalian transcriptome in normal and disease states is unknown. The mammalian genome contains only a few model systems where epigenetic gene regulation has been shown to play a major role in transcriptional control. These model systems are important not only to investigate the biological function of known epigenetic modifications but also to identify new and unexpected epigenetic mechanisms in the mammalian genome. Here we review recent progress in understanding how epigenetic mechanisms control imprinted gene expression.
Introduction
The last few years have seen a tremendous breakthrough in high-throughput sequencing technologies that allow histone and DNA modifications, transcription factors, and RNA polymerases to be exactly localized throughout the genome relative to expressed or silent genes [1-3]. These studies provide important information about the chromatin state of expressed or silent genes, but give no insight into whether the chromatin state is the cause or effect of changes in gene expression. An alternative approach is to apply these mapping techniques to genes that are known to be subject to specific epigenetic regulation, to identify chromatin and transcription features of these genes and then to test the relevance of these features for gene expression. In this way a large number of epigenetic gene-regulatory models (such as yeast mating-type switching, transgene position effect variegation, transposon silencing, and centromere silencing) have been proposed as models to probe how the chromatin state controls expressed and silent genes [4]. In mammals, systems showing mono-allelic expression in diploid cells such as X-chromosome inactivation and genomic imprinting show very clear evidence of epigenetic regulation that restricts expression to one parental allele in diploid cells. Here we overview recent progress in using genomic imprinting (Box 2) as a model of epigenetic gene regulation in mammals, which indicates that epigenetic mechanisms do not always follow the most straightforward route to regulate gene expression.
Box 2 Genomic imprinting background.
While most genes in mammalian diploid somatic cells are expressed from both parental chromosomes (i.e., show bi-allelic expression), imprinted genes show parental-specific mono-allelic expression [6,51,52•]. In contrast to X-chromosome inactivation, the sex of the offspring is not important, instead, imprinted expression is dictated by the parental origin of the chromosome and affected genes show the same parental-specific expression in both male and female offspring. To date 131 maternally or paternally expressed imprinted genes have been identified in mice and many of these control essential functions in embryonic development (http://www.mousebook.org/catalog.php?catalog=imprinting [49••]). Notably, although imprinted gene expression can vary in development, differentiation and disease; the parental origin of the expressed allele does not. For example, the mouse Igf2r (Insulin-like growth factor type 2 receptor) gene shows bi-allelic expression in early pre-implantation embryos that changes to maternal-specific expression in early post-implantation embryos, and in post-mitotic neurons only, Igf2r reverts to bi-allelic expression [53,54]. The key to understanding how imprinted expression can show tissue-specific or developmental variation, was the appreciation of two features: (i) that DNA methylation imprints (Box 1) are not gene-specific but instead, repress a long-range regulatory element (known as the imprint control element or region: ICE or ICR) that represses clusters of genes in cis (Box 1), and (ii), that the repressor activity of the ICE is developmentally and tissue-specifically regulated [55,56]. Thus, the ICE that is universally present in all somatic cells is ‘imprinted’ on one chromosome by DNA methylation or ‘not imprinted’ on the other chromosome, and genes regulated by the non-imprinted unmethylated ICE, show imprinted expression in some tissues and bi-allelic expression in others.
It has recently been appreciated that multiple mechanistic steps lead to the stable inheritance of epigenetic phenotypes through cell division [5••]. This new operational definition of epigenetics proposes three logical steps: first an ‘epigenator signal’ from outside the cell is required to trigger an intracellular pathway, second an ‘epigenetic initiator’ responds to this pathway in a locus-specific manner, and third, ‘epigenetic maintainers’ change the chromatin state at the locus recognized by the initiator and maintain this state throughout subsequent cell divisions. While little is yet known of epigenator signals, epigenetic initiators include DNA-binding proteins and non-coding RNAs, while the epigenetic maintainers include the well-known processes of DNA methylation, histone modifications, histone variants, or nucleosome remodelling [5••]. With this operational definition of epigenetics in mind, we overview here recent progress in understanding how genomic imprinting induces parental-specific gene expression.
The key role of the imprint control element (ICE) in genomic imprinting
In mammals, genomic imprinting acts as a block to parthenogenesis (Box 1) and mouse embryos carrying two maternal genomes normally die at mid-gestation [6]. However, bi-maternal mice can be generated from one normal haploid maternal genome and one immature haploid maternal genome genetically manipulated to delete two ICEs (Box 1), which normally gain a paternal methylation imprint during spermatogenesis [7••]. In these bi-maternal mice one haploid chromosome set obtained from mature oocytes carries normal maternal imprints and shows typical maternal-specific imprinted expression (Figure 1). The other genetically manipulated haploid chromosome set was transferred from an immature oocyte and so lacks maternal imprints. The combination of a lack of maternal imprints plus a lack of two normally paternally methylated ICEs not only fully substitutes for a paternally inherited chromosome set, but also shows that no other paternally methylated ICE regulates essential genes during development (note that Figure 1 only shows chromosome 7, the other deleted paternally methylated ICE in these bi-maternal mice lies on chromosome 12). While this appears to be a complicated story there is a simple explanation that highlights the two key features of the ICE: (i) On one parental chromosome the unmethylated ICE is a cis-acting repressor. Thus, deletion of the unmethylated ICE will relieve gene repression. In Figure 1, deletion of the normally paternally methylated ICE relieves Igf2 repression on the maternal chromosome. (ii) On the other parental chromosome a gametic DNA methylation imprint acquired during male or female gametogenesis represses ICE function. In Figure 1 expression of Cdkn1c from a maternal chromosome requires maternal ICE methylation. Note that deletion of the methylated ICE will not change gene expression.
Box 1 Glossary
- Epigenetics
A heritable phenotype resulting from changes in a chromosome without alterations in the DNA sequence [5••].
- Cis/Trans effect
‘in cis’ refers to an action on the same chromosome, ‘in trans’ refers to an action on other chromosomes.
- Genomic imprinting
An epigenetic mechanism leading to parental-specific expression normally affecting small clusters of genes (Box 2).
- ICE
Imprint control element is a short DNA element whose epigenetic state controls imprinted expression of all genes in one imprinted cluster (Box 2).
- Imprint
An epigenetic mark on the ICE which distinguishes the parental alleles of an imprinted gene cluster in the gametes. So far only DNA methylation has been shown to act as the imprint and the imprinted chromosome is here defined as the allele that gains ICE DNA methylation in one parental gamete (Box 2).
- Macro (long) ncRNA
A new class of non-coding (nc) RNAs with regulatory potential defined as >200 bp whose function does not depend on processing into short/micro/pi RNAs [43-45,33].
- Non-coding (nc) RNA
RNAs not translated into a protein but since they could ‘code’ for a function the alternative terms, non-protein-coding (npc) or untranslated (ut) are more accurate [50].
- Parthenogenesis
In mammals, reproduction without fertilization in female oocytes requiring doubling of the oocyte haploid chromosome set.
- Retrogene
An expressed retrotransposed gene copy that retains its protein-coding capacity.
Figure 1.

Bypassing paternal imprints to generate bi-maternal mice. On mouse chromosome 7, a paternal DNA methylation imprint (Me/blue circle) represses the ICE and allows expression of Igf2 from the paternal chromosome in normal diploid embryonic cells (arrow). Igf2 is not expressed from a maternal chromosome that has an active unmethylated ICE (lollipop). A similar but opposite situation occurs in a neighboring imprinted cluster on this chromosome, where expression of Cdkn1c depends on a maternally methylated ICE (Me/red circle). Note that chromosome 7 contains only one of the two normally paternally methylated ICEs deleted to generate bi-maternal mice [7••]. DNA methylation imprints on ICEs are erased in primordial germ cells of the developing gonad and in females these imprints are reacquired during oocyte maturation. Chromosomes in immature oocytes lack maternal ICE imprints and, if they also genetically lack Pat-ICEs (oblique rectangle) that are normally modified by paternal gametic DNA methylation, then this haploid chromosome set will have a maternal origin with the imprinted expression pattern of the paternal genome [7••].
The unmethylated ice is a cis-acting repressor
To date there are three known ways in which the unmethylated ICE acts as an ‘epigenetic initiator’ to repress genes in cis. Moreover, variations in imprinted expression arise because these initiator mechanisms can show tissue- and developmental-specific regulation. In the first identified mechanism, an analysis of the Igf2 imprinted cluster on mouse chromosome 7 (Figure 2a) shows the unmethylated ICE acts as an insulator that binds the zinc finger transcription factor CTCF to block the access of Igf2 to enhancers that lie downstream of the H19 ncRNA [8]. Recently, COHESIN, a protein complex essential for sister chromatid cohesion, was shown to bind the same sites in mammalian genomes as CTCF [9••,10••]. Both CTCF and COHESIN have now been shown to be necessary to induce specific chromatin loops necessary for Igf2 silencing on the maternal chromosome [11,12••]. In the second identified mechanism, an analysis of two imprinted clusters (Igf2r and Kcnq1) shows the unmethylated ICE contains an active promoter for a non-coding RNA that represses multiple genes in cis (Figure 2b). In the Igf2r imprinted cluster the 108 kb long Airn ncRNA represses three genes spread over 300 kb in cis, while in the Kcnq1 imprinted cluster the 90 kb long Kcnq1ot1 ncRNA represses in cis, 11 genes spread over 800 kb [13,14]. The Airn ncRNA when shortened to 3% is unable to repress genes in its imprinted cluster in all tested tissues. However, in the Kcnq1 cluster while most genes are not repressed by a shortened Kcnq1ot1 ncRNA, the Cdkn1c gene remains repressed in some, but not all tissues [15•]. This indicates that in some tissues, either a short form of the Kcnq1ot1 ncRNA is sufficient for silencing Cdkn1c, or, that two distinct mechanisms operate in this cluster. In the third example (Figure 2c), analysis of the H13 imprinted cluster shows the unmethylated ICE contains an active promoter for the Mcts2 retrogene [16••]. Expression of Mcts2 or the unmethylated ICE itself causes H13 to use internal polyadenylation sites and the resulting short transcripts lack enzyme activity. Although superficially similar to the situation in the Igf2r and Kcnq1 imprinted cluster, the transposed Mcts2 retrogene retains an open-reading-frame and protein-coding capacity.
Figure 2.
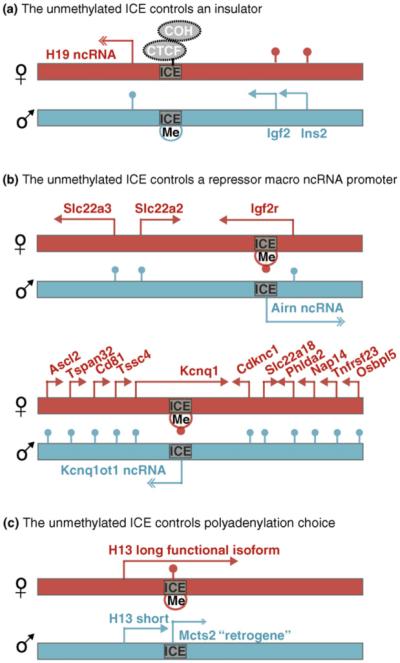
The unmethylated ICE is a cis-acting repressor. Three examples of how the unmethylated ICE can repress mRNA genes in cis are known. (a) The unmethylated ICE in the mouse Igf2 cluster on chromosome 7 forms an insulator on the maternal chromosome by binding CTCF and COHESIN (COH) proteins, which blocks the access of Igf2 to enhancers located downstream to the H19 ncRNA [8,12••]. (b) The unmethylated ICE in the mouse Igf2r imprinted gene cluster on chromosome 17 (top) and in the mouse Kcnq1 imprinted gene cluster on chromosome 7 (bottom) contains an active promoter, respectively, for the Airn and Kcnq1ot1 macro ncRNAs. Both these ncRNAs repress multiple genes in cis on the paternal chromosome [13,14,15•]. (c) The unmethylated ICE in the mouse H13 (Minor histocompatibility antigen H13 encoding a signal-peptide peptidase) imprinted cluster on chromosome 2 contains the active promoter for the Mcts2 retrogene, and either the unmethylated ICE or Mcts2 expression induces premature polyadenylation of H13 transcripts that lack enzyme activity [16••]. The maps are not drawn to scale and show imprinted expression in the visceral yolk sac (A), for placenta (B) and in adult brain (C); genes showing bi-allelic expression are not indicated. Arrow: expressed gene, Double-headed arrow: expressed ncRNA or retrogene, lollipop: silent gene, Me/blue circle: paternal gametic methylation imprint, Me/red circle: maternal gametic methylation imprint.
Epigenetic initiators at the unmethylated ICE have been shown to control parental-specific silencing in all types of mouse tissue (references in Figure 2). However, recent studies indicate that epigenetic maintainers responding to the epigenetic initiator may differ in mouse embryonic/adult tissues and extra-embryonic tissues. The latter comprise the placenta and membranes of mouse embryos that are short-lived tissues and only present during the 21-day embryonic gestation period [17•]. For example, in placenta the Airn ncRNA represses three genes Slc22a3, Slc22a2, and Igf2r (Figure 2). In embryo and adult tissues, only Igf2r is repressed by Airn. In the placenta, Airn was recently shown to recruit G9A (KMT1C) a histone H3-K9-dimethylase to silence Slc22a3 (Slc22a2 could not be tested as it is not expressed before G9A null embryos die at 10.5 dpc) [18••]. Igf2r was not affected by the loss of G9A, indicating Slc22a3 and Igf2r are silenced by different mechanisms in placenta. The Kcnq1ot1 ncRNA was also recently shown to localize to a nuclear compartment with Polycomb group (PcG) proteins that induce a contracted state in the paternal allele necessary for silencing of some, but not all genes in extra-embryonic tissues, but not in embryonic tissues [19••].
An earlier analysis of mice deficient in the EED Polycomb protein, similarly showed loss of imprinted expression of only 4/14 tested genes in extra-embryonic tissues but no effect in embryonic tissues [20]. These data indicate that histone modifications play a partial role as epigenetic maintainers of extra-embryonic imprinted expression. In contrast, histone modifications have not yet been shown to play an epigenetic maintainer role in mouse embryonic tissues. Genome-wide maps of repressive histone H3K9me3 marks in embryonic stem (ES) cells have shown they are focally restricted to the DNA methylated ICE and do not spread throughout imprinted clusters [21••]. We have recently shown that the Igf2r gene is devoid of Polycomb-dependent H3K27me3 modifications when it shows imprinted expression in differentiated embryonic cells. Moreover, although this mark is present in undifferentiated embryonic stem (ES) cells, it is lost from both parental alleles when Igf2r gains imprinted expression in differentiated ES cells [22••,23••]. This contrasts with tissue-specific silent genes that are contained in broad domains of H3K27me3 in differentiated embryonic cells [24••]. Notably, very few genes repressed in cis by the unmethylated ICE, subsequently gain promoter DNA methylation. The mouse Igf2r promoter and Cdkn1c CpG island promoters are an exception and both are methylated when repressed (Figure 2). However, both of these genes are silenced by their respective macro-ncRNAs in the absence of genome-wide DNA methylation [25-28]. A new model for generating imprinted expression based on kinetic experiments at the Igf2r imprinted cluster demonstrated that the repressed allele maintained low-level basal expression, and that imprinted expression arose from an expression bias between the two parental alleles [23••]. While it remains to be tested how general this expression-bias model is, it may explain the widespread lack of repressive marks on genes repressed by the active ICE, and the frequent finding that the expression of imprinted genes becomes biallelic, but is reduced to basal levels in the absence of ICE methylation [29-31].
Gametically acquired DNA methylation imprints repress ICE function
The situation on the one parental chromosome carrying the methylated ICE can be reduced to the question of how an ICE gains DNA methylation in one gamete and avoids methylation in the other gamete. Since the targeting and maintenance of DNA methylation patterns in mammals is poorly understood, ICE methylation offers an important model system. To date 15 ICEs have been provisionally identified based on their gain of maternal methylation during oogenesis (12 examples) or paternal methylation during spermatogenesis (3 examples) [32]; however, only 6 of these have been genetically shown to have ICE activity [33•]. The de novo and maintenance DNA methyltransferase complexes that are responsible for genome-wide DNA methylation in somatic tissues [34•] are also responsible for gametic ICE methylation. In particular, the germline specific DNMT3A2 de novo methyltransferase isoform is necessary for ICE DNA methylation in male and female gametes, while the DNMT3L accessory protein is required specifically for maternal gametic ICE methylation, but not for paternal ICE methylation [35,36]. DNMT3L has been shown to play an important role in targeting DNA methylation to regions containing histone H3 that lacks K4 methylation [37••]. Recently, an H3K4 demethylase (KDM1B) was also shown to be necessary for establishing maternal ICE methylation in 4/7 tested ICEs, but played no role in methylation acquisition of 2/3 tested paternally methylated ICEs [38••]. Once ICE methylation imprints have been acquired in male and female gametes, maintenance of a subset of methylated ICE requires additional proteins such as the germ cell and oocyte-specific nuclear PGC7/Stella protein, and a KRAB zinc finger protein encoded by the Zfp57 locus [39••,40••].
While these experiments have clearly identified some of the key players in the acquisition and maintenance of methylation imprints, they do not explain how methylation is targeted to ICEs that lack obvious sequence-specificity apart from a high CpG content and the presence of tandem direct repeats in some ICEs [32,41]. One possibility suggested from an analysis of the ICE in the mouse Gnas imprinted cluster, is that overlapping transcription from a protein-coding mRNA gene may be required for oocyte (maternal) ICE methylation [42••]. All the 12 known maternally methylated ICEs [32] are overlapped by protein-coding genes and many of these were also shown to be transcribed in oocytes [42••], indicating this could be a general epigenetic initiator mechanism leading to maternal-specific ICE methylation. These data are not contradicted by the above demonstration that H3K4 demethylation may be a prerequisite for DNA methyltransferase targeted via DNMT3L [38••], as H3K4 methylation is not distributed equally through transcribed genes, but is concentrated focally at promoters and enhancers [1].
Conclusions
The analysis of epigenetic mechanisms at imprinted gene clusters has yielded a wealth of information particularly at the level of the locus-specific epigenetic initiator, where the unexpected appears to rule. The discovery of the ICE, which is a long-range cis-acting repressor that is itself repressed by DNA methylation, not only sheds light on the domain regulation of genes, but also demonstrates that one biological role of DNA methylation in mammals is to express genes normally repressed by default. The subsequent discovery that the unmethylated ICE can repress genes in cis by different mechanisms including insulator formation and macro ncRNA or retrogene expression also has a wider implication for mammalian genome biology. We do not know yet if methyl-sensitive long-range regulatory elements are common features of the mammalian genome outside of imprinted clusters. Macro ncRNAs that have long been associated with imprinted gene clusters [33•] are, however, now being seen as part of the genome-wide regulatory repertoire in mammals [43-45]. Notably, macro ncRNAs have also been shown to regulate non-imprinted disease phenotypes [46••,47,48••]. It is yet too early to know if genomic imprinting provides an accurate epigenetic regulatory model of tissue-specific silencing or, whether it better reflects abnormal gene silencing occurring, for example, in aging or in tumor development. However, since the analysis of imprinted gene clusters has been unevenly concentrated on 6 of the known 26 regions containing imprinted genes [33•,49••], we can be certain they will continue to provide one of the best discovery models for epigenetic gene regulation in mammals.
Acknowledgements
We thank the whole Barlow Group for lively discussions on the themes presented here and Michael Jantsch, Florian Pauler and Quanah Hudson for reading the manuscript. The authors are supported by EU-FW6 IP ‘HEROIC’ (LSHG-CT-2005-018883), The EU-FW6 NoE ‘THE EPIGENOME’ (LSHG-Ct-2004-053433) and the FWF Modulators of RNA Fate and Function (SFB-P1718 B10).
Footnotes
Conflict of interests
The authors declare there is no conflict of interest.
References and recommended reading
Papers of particular interest, published within the period of review, have been highlighted as:
• of special interest
•• of outstanding interest
- 1.Barski A, Zhao K. Genomic location analysis by ChIP-Seq. J Cell Biochem. 2009;107:11–18. doi: 10.1002/jcb.22077. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Lister RPM, Dowen RH, Hawkins RD, Hon G, Tonti-Filippini J, Nery JR, Lee L, Ye Z, Ngo QM, Edsall L, et al. Human DNA methylomes at base resolution show widespread epigenomic differences. Nature. 2009;462:315–322. doi: 10.1038/nature08514. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Carninci P. Is sequencing enlightenment ending the dark age of the transcriptome? Nat Methods. 2009;6:711–713. doi: 10.1038/nmeth1009-711. [DOI] [PubMed] [Google Scholar]
- 4.Allis CD, Jenuwein T, Reinberg D. In: Epigenetics. edn 1. Marie-Laure Caparros AE, editor. Cold Spring Harbor Laboratory Press; New York: 2007. [Google Scholar]
- 5••.Berger SL, Kouzarides T, Shiekhattar R, Shilatifard A. An operational definition of epigenetics. Genes Dev. 2009;23:781–783. doi: 10.1101/gad.1787609. The results of a meeting hosted by the Banbury Conference Center and Cold Spring Harbor Laboratory that discussed epigenetic control of genomic function reached a consensus definition of “epigenetics” to be considered by the broader community based on multiple mechanistic steps leading to the stable heritance of the epigenetic phenotype.
- 6.Barlow DP, Bartolomei MS. In: Genomic Imprinting in Mammals. Epigenetics. David Allis C, Jenuwein Thomas, Reinberg Danny, editors. Cold Spring Harbor Laboratory Press; New York: 2007. Chapter 19. [Google Scholar]
- 7••.Kawahara M, Kono T. Longevity in mice without a father. Hum Reprod. 2009 doi: 10.1093/humrep/dep400. Epub ahead of print. A new analysis of bi-maternal mice that lack a paternal genomic contribution, which shows longevity may have been the price to pay for sexual reproduction in mammals.
- 8.Hark AT, Schoenherr CJ, Katz DJ, Ingram RS, Levorse JM, Tilghman SM. CTCF mediates methylation-sensitive enhancer-blocking activity at the H19/Igf2 locus. Nature. 2000;405:486–489. doi: 10.1038/35013106. [DOI] [PubMed] [Google Scholar]
- 9••.Parelho V, Hadjur S, Spivakov M, Leleu M, Sauer S, Gregson HC, Jarmuz A, Canzonetta C, Webster Z, Nesterova T, et al. Cohesins functionally associate with CTCF on mammalian chromosome arms. Cell. 2008;132:422–433. doi: 10.1016/j.cell.2008.01.011. See annotations of Ref. [10••].
- 10••.Wendt KS, Yoshida K, Itoh T, Bando M, Koch B, Schirghuber E, Tsutsumi S, Nagae G, Ishihara K, Mishiro T, et al. Cohesin mediates transcriptional insulation by CCCTC-binding factor. Nature. 2008;451:796–801. doi: 10.1038/nature06634. Ref. [9••] along with this paper show in the mouse and human genome that COHESIN is found at most CTCF (CCCTC-binding factor) binding sites and that CTCF is required for COHESIN localization. This explains previous findings that COHESIN, originally identified as mediating chromatid segregation, appeared to regulate gene expression and enhancer–promoter interactions.
- 11.Kurukuti S, Tiwari VK, Tavoosidana G, Pugacheva E, Murrell A, Zhao Z, Lobanenkov V, Reik W, Ohlsson R. CTCF binding at the H19 imprinting control region mediates maternally inherited higher-order chromatin conformation to restrict enhancer access to Igf2. Proc Natl Acad Sci USA. 2006;103:10684–10689. doi: 10.1073/pnas.0600326103. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12••.Nativio R, Wendt KS, Ito Y, Huddleston JE, Uribe-Lewis S, Woodfine K, Krueger C, Reik W, Peters JM, Murrell A. Cohesin is required for higher-order chromatin conformation at the imprinted IGF2-H19 locus. PLoS Genet. 2009;5:e1000739. doi: 10.1371/journal.pgen.1000739. The authors use chromosome conformation capture (3C) assays and RNAi-mediated depletion to show that COHESIN maintains CTCF-mediated chromatin conformation at the imprinted Igf2 cluster locus that is independent of the function of COHESIN in sister chromatid cohesion.
- 13.Mancini-Dinardo D, Steele SJ, Levorse JM, Ingram RS, Tilghman SM. Elongation of the Kcnq1ot1 transcript is required for genomic imprinting of neighboring genes. Genes Dev. 2006;20:1268–1282. doi: 10.1101/gad.1416906. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Sleutels F, Tjon G, Ludwig T, Barlow DP. Imprinted silencing of Slc22a2 and Slc22a3 does not need transcriptional overlap between Igf2r and Air. EMBO J. 2003;22:3696–3704. doi: 10.1093/emboj/cdg341. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15•.Shin JY, Fitzpatrick GV, Higgins MJ. Two distinct mechanisms of silencing by the KvDMR1 imprinting control region. EMBO J. 2008;27:168–178. doi: 10.1038/sj.emboj.7601960. The authors show that truncation of the Kcnq1ot1 macro ncRNA does not affect imprinted expression of Cdkn1c in a subset of embryonic tissues. This indicates that in some tissues, either a short form of the Kcnq1ot1 ncRNA is sufficient for silencing Cdkn1c, or, that two distinct mechanisms operate in this cluster.
- 16••.Wood AJ, Schulz R, Woodfine K, Koltowska K, Beechey CV, Peters J, Bourc’his D, Oakey RJ. Regulation of alternative polyadenylation by genomic imprinting. Genes Dev. 2008;22:1141–1146. doi: 10.1101/gad.473408. The demonstration that imprinted expression can arise through allele-specific differences in cotranscriptional processes, which also shows that epigenetic modifications can influence utilization of alternative polyadenylation sites.
- 17•.Hudson QJ, Kulinski TM, Huetter SP, Barlow DP. Genomic Imprinting Mechanisms In Embryonic And Extra-Embryonic Mouse Tissues. Heredity. 2010 doi: 10.1038/hdy.2010.23. in press. An up-to-date overview of the similarities and differences between genomic imprinting in embryonic and extra-embryonic tissues.
- 18••.Nagano T, Mitchell JA, Sanz LA, Pauler FM, Ferguson-Smith AC, Feil R, Fraser P. The Air noncoding RNA epigenetically silences transcription by targeting G9a to chromatin. Science. 2008;322:1717–1720. doi: 10.1126/science.1163802. The Airn macro ncRNA is shown to act differently in placenta to silence the Slc22a3 gene which shows placental-specific imprinted expression compared to the Igf2r gene which shows imprinted expression in both placenta and embryo.
- 19••.Terranova R, Yokobayashi S, Stadler MB, Otte AP, van Lohuizen M, Orkin SH, Peters AH. Polycomb group proteins Ezh2 and Rnf2 direct genomic contraction and imprinted repression in early mouse embryos. Dev Cell. 2008;15:668–679. doi: 10.1016/j.devcel.2008.08.015. This paper shows that in early extra-embryonic cells of pre-implantation embryos, the paternally expressed Kcnq1ot1 macro ncRNA uses POLYCOMB proteins to direct chromatin contraction to the paternal chromosome and that POLYCOMB mutants fail to repress some imprinted genes in later extra-embryonic tissues but not in embryonic tissues. Both this and reference [18], show that macro ncRNAs may act differently to repress genes in extra-embryonic and embryonic tissues.
- 20.Mager J, Montgomery ND, de Villena FP, Magnuson T. Genome imprinting regulated by the mouse Polycomb group protein Eed. Nat Genet. 2003;33:502–507. doi: 10.1038/ng1125. [DOI] [PubMed] [Google Scholar]
- 21••.Mikkelsen TS, Ku M, Jaffe DB, Issac B, Lieberman E, Giannoukos G, Alvarez P, Brockman W, Kim TK, Koche RP, et al. Genome-wide maps of chromatin state in pluripotent and lineage-committed cells. Nature. 2007 doi: 10.1038/nature06008. High-throughput profiling of histone modifications in mouse embryonic stem (ES) cells which shows that imprinted clusters contain focal H3K9me3 on the ICE or Imprint Control Element.
- 22••.Regha K, Sloane MA, Huang R, Pauler FM, Warczok KE, Melikant B, Radolf M, Martens JH, Schotta G, Jenuwein T, et al. Active and repressive chromatin are interspersed without spreading in an imprinted gene cluster in the mammalian genome. Mol Cell. 2007;27:353–366. doi: 10.1016/j.molcel.2007.06.024. A detailed study of multiple active and repressive histone modifications in the mouse Igf2r imprinted cluster in mouse embryonic fibroblasts, which shows a lack of widespread repressive marks and the presence of focal heterochromatin comprising H3K9me3/H4K20me3/HP1 on the maternally methylated ICE and on the paternally repressed Igf2r promoter. H3K27me3 was shown to be absent from both parental alleles in this cluster.
- 23••.Latos PA, Stricker SH, Steenpass L, Pauler FM, Huang R, Senergin BH, Regha K, Koerner MV, Warczok KE, Unger C, et al. An in vitro ES cell imprinting model shows that imprinted expression of the Igf2r gene arises from an allele-specific expression bias. Development. 2009;136:437–448. doi: 10.1242/dev.032060. The demonstration that differentiating embryonic stem (ES) cells provide an in vitro system for studying the developmental gain of imprinted Igf2r expression that reveals a novel mechanism for Airn-mediated imprinting—through an expression bias rather than silencing.
- 24••.Pauler FM, Sloane MA, Huang R, Regha K, Koerner MV, Tamir I, Sommer A, Aszodi A, Jenuwein T, Barlow DP. H3K27me3 forms BLOCs over silent genes and intergenic regions and specifies a histone banding pattern on a mouse autosomal chromosome. Genome Res. 2009;19:221–233. doi: 10.1101/gr.080861.108. A demonstration that in mouse embryonic fibroblasts the majority of H3K27me3 modifications cover large domains called ‘BLOCs’ rather than focal peaks. H3K27me3 BLOCs were shown to modify tissue-specific silent genes of all types, plus flanking intergenic regions and their distribution indicates a negative correlation between H3K27me3 and transcription.
- 25.Li E, Beard C, Jaenisch R. Role for DNA methylation in genomic imprinting. Nature. 1993;366:362–365. doi: 10.1038/366362a0. [DOI] [PubMed] [Google Scholar]
- 26.Seidl CI, Stricker SH, Barlow DP. The imprinted Air ncRNA is an atypical RNAPII transcript that evades splicing and escapes nuclear export. EMBO J. 2006;25:3565–3575. doi: 10.1038/sj.emboj.7601245. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Lee J, Inoue K, Ono R, Ogonuki N, Kohda T, Kaneko-Ishino T, Ogura A, Ishino F. Erasing genomic imprinting memory in mouse clone embryos produced from day 11.5 primordial germ cells. Development. 2002;129:1807–1817. doi: 10.1242/dev.129.8.1807. [DOI] [PubMed] [Google Scholar]
- 28.Onyango P, Miller W, Lehoczky J, Leung CT, Birren B, Wheelan S, Dewar K, Feinberg AP. Sequence and comparative analysis of the mouse 1-megabase region orthologous to the human 11p15 imprinted domain. Genome Res. 2000;10:1697–1710. doi: 10.1101/gr.161800. [DOI] [PubMed] [Google Scholar]
- 29.Bhogal B, Arnaudo A, Dymkowski A, Best A, Davis TL. Methylation at mouse Cdkn1c is acquired during postimplantation development and functions to maintain imprinted expression. Genomics. 2004;84:961–970. doi: 10.1016/j.ygeno.2004.08.004. [DOI] [PubMed] [Google Scholar]
- 30.Lewis A, Mitsuya K, Umlauf D, Smith P, Dean W, Walter J, Higgins M, Feil R, Reik W. Imprinting on distal chromosome 7 in the placenta involves repressive histone methylation independent of DNA methylation. Nat Genet. 2004;36:1291–1295. doi: 10.1038/ng1468. [DOI] [PubMed] [Google Scholar]
- 31.Green K, Lewis A, Dawson C, Dean W, Reinhart B, Chaillet JR, Reik W. A developmental window of opportunity for imprinted gene silencing mediated by DNA methylation and the Kcnq1ot1 noncoding RNA. Mamm Genome. 2007;18:32–42. doi: 10.1007/s00335-006-0092-9. [DOI] [PubMed] [Google Scholar]
- 32.Kobayashi H, Suda C, Abe T, Kohara Y, Ikemura T, Sasaki H. Bisulfite sequencing and dinucleotide content analysis of 15 imprinted mouse differentially methylated regions (DMRs): paternally methylated DMRs contain less CpGs than maternally methylated DMRs. Cytogenet Genome Res. 2006;113:130–137. doi: 10.1159/000090824. [DOI] [PubMed] [Google Scholar]
- 33•.Koerner MV, Pauler FM, Huang R, Barlow DP. The function of non-coding RNAs in genomic imprinting. Development. 2009;136:1771–1783. doi: 10.1242/dev.030403. An up-to-date overview of the organization and epigenetic regulation of six well-studied mouse imprinted gene clusters.
- 34•.Ooi SK, O’Donnell AH, Bestor TH. Mammalian cytosine methylation at a glance. J Cell Sci. 2009;122:2787–2791. doi: 10.1242/jcs.015123. An admirably succinct up-to-date overview of the known factors involved in the establishment and maintenance of methylation profiles in the mammalian genome.
- 35.Bourc’his D, Xu GL, Lin CS, Bollman B, Bestor TH. Dnmt3L and the establishment of maternal genomic imprints. Science. 2001;294:2536–2539. doi: 10.1126/science.1065848. [DOI] [PubMed] [Google Scholar]
- 36.Kaneda M, Okano M, Hata K, Sado T, Tsujimoto N, Li E, Sasaki H. Essential role for de novo DNA methyltransferase Dnmt3a in paternal and maternal imprinting. Nature. 2004;429:900–903. doi: 10.1038/nature02633. [DOI] [PubMed] [Google Scholar]
- 37••.Ooi SK, Qiu C, Bernstein E, Li K, Jia D, Yang Z, Erdjument-Bromage H, Tempst P, Lin SP, Allis CD, et al. DNMT3L connects unmethylated lysine 4 of histone H3 to de novo methylation of DNA. Nature. 2007;448:714–717. doi: 10.1038/nature05987. DNMT3L was shown to specifically interact with the extreme amino terminus of histone H3; this interaction was strongly inhibited by methylation at lysine 4 of histone H3 but was insensitive to modifications at other positions. This indicates that DNMT3L recognizes histone H3 tails that are unmethylated at lysine 4 and induces de novo DNA methylation by recruitment or activation of DNMT3A2.
- 38••.Ciccone DN, Su H, Hevi S, Gay F, Lei H, Bajko J, Xu G, Li E, Chen T. KDM1B is a histone H3K4 demethylase required to establish maternal genomic imprints. Nature. 2009;461:415–418. doi: 10.1038/nature08315. Oocytes from KDM1B-deficient females showed increased H3K4 methylation and failed to set up the DNA methylation marks at four out of seven imprinted genes examined. This shows demethylation of H3K4 is critical for establishing the DNA methylation imprints during oogenesis.
- 39••.Nakamura T, Arai Y, Umehara H, Masuhara M, Kimura T, Taniguchi H, Sekimoto T, Ikawa M, Yoneda Y, Okabe M, et al. PGC7/Stella protects against DNA demethylation in early embryogenesis. Nat Cell Biol. 2007;9:64–71. doi: 10.1038/ncb1519. See annotations of Ref. [40••].
- 40••.Li X, Ito M, Zhou F, Youngson N, Zuo X, Leder P, Ferguson-Smith AC. A maternal-zygotic effect gene, Zfp57, maintains both maternal and paternal imprints. Dev Cell. 2008;15:547–557. doi: 10.1016/j.devcel.2008.08.014. Along with Ref. [39••] this paper identifies factors that are needed to maintain DNA methylation at a subset of maternally and paternally methylated ICEs.
- 41.Neumann B, Kubicka P, Barlow DP. Characteristics of imprinted genes. Nat Genet. 1995;9:12–13. doi: 10.1038/ng0195-12. [DOI] [PubMed] [Google Scholar]
- 42••.Chotalia M, Smallwood SA, Ruf N, Dawson C, Lucifero D, Frontera M, James K, Dean W, Kelsey G. Transcription is required for establishment of germline methylation marks at imprinted genes. Genes Dev. 2009;23:105–117. doi: 10.1101/gad.495809. The demonstration that transcription of overlapping mRNA genes plays a role in targeting maternal DNA methylation to the ICE in the Gnas cluster. Overlapping transcription was also identified on other maternally methylated ICEs, indicating it may be an essential component of the oocyte de novo methylation system.
- 43.Mercer TR, Dinger ME, Mattick JS. Long non-coding RNAs: insights into functions. Nat Rev Genet. 2009;10:155–159. doi: 10.1038/nrg2521. [DOI] [PubMed] [Google Scholar]
- 44.Gingeras TR. Origin of phenotypes: genes and transcripts. Genome Res. 2007;17:682–690. doi: 10.1101/gr.6525007. [DOI] [PubMed] [Google Scholar]
- 45.Wilusz JE, Sunwoo H, Spector DL. Long noncoding RNAs: functional surprises from the RNA world. Genes Dev. 2009;23:1494–1504. doi: 10.1101/gad.1800909. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46••.Niessen RC, Hofstra RM, Westers H, Ligtenberg MJ, Kooi K, Jager PO, de Groote ML, Dijkhuizen T, Olderode-Berends MJ, Hollema H, et al. Germline hypermethylation of MLH1 and EPCAM deletions are a frequent cause of Lynch syndrome. Genes Chromosomes Cancer. 2009;48:737–744. doi: 10.1002/gcc.20678. Germline deletions of the 3′ region of EPCAM was shown to cause sense transcriptional read-through of the downstream MSH2 tumor suppressor gene, which resulted in silencing by hypermethylation.
- 47.Tufarelli C, Stanley JA, Garrick D, Sharpe JA, Ayyub H, Wood WG, Higgs DR. Transcription of antisense RNA leading to gene silencing and methylation as a novel cause of human genetic disease. Nat Genet. 2003;34:157–165. doi: 10.1038/ng1157. [DOI] [PubMed] [Google Scholar]
- 48••.Yu W, Gius D, Onyango P, Muldoon-Jacobs K, Karp J, Feinberg AP, Cui H. Epigenetic silencing of tumour suppressor gene p15 by its antisense RNA. Nature. 2008;451:202–206. doi: 10.1038/nature06468. A study that shows many tumor suppressor genes have nearby antisense RNAs that demonstrated an inverse relation between p15 antisense (p15AS) and p15 sense expression in leukaemia. References [46••,47,48••] indicate a role for macro ncRNAs in deregulating gene expression in human disease.
- 49••.Williamson CM, Blake A, Thomas S, Beechey CV, Hancock J, Cattanach BM, Peters J. Mouse Imprinting Data and References World Wide Web Site - Mouse Imprinting Data and References MRC Harwell. Oxfordshire; 2009. http://www.har.mrc.ac.uk/research/genomic_imprinting. The renowned imprinting database that comprise information on mouse chromosome regions associated with imprinted phenotypes, imprinted genes within these regions, and imprinted genes in other regions of the genome.
- 50.Brosius J, Tiedge H. RNomenclature. RNA Biol. 2004;1:81–83. doi: 10.4161/rna.1.2.1228. [DOI] [PubMed] [Google Scholar]
- 51.Bartolomei MS. Genomic imprinting: employing and avoiding epigenetic processes. Genes Dev. 2009;23:2124–2133. doi: 10.1101/gad.1841409. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52•.Renfree MB, Hore TA, Shaw G, Graves JA, Pask AJ. Evolution of genomic imprinting: insights from marsupials and monotremes. Annu Rev Genomics Hum Genet. 2009;10:241–262. doi: 10.1146/annurev-genom-082908-150026. An excellent review of the evolution and function of genomic imprinting in mammals.
- 53.Yamasaki Y, Kayashima T, Soejima H, Kinoshita A, Yoshiura K, Matsumoto N, Ohta T, Urano T, Masuzaki H, Ishimaru T, et al. Neuron-specific relaxation of Igf2r imprinting is associated with neuron-specific histone modifications and lack of its antisense transcript Air. Hum Mol Genet. 2005;14:2511–2520. doi: 10.1093/hmg/ddi255. [DOI] [PubMed] [Google Scholar]
- 54.Szabo PE, Mann JR. Allele-specific expression and total expression levels of imprinted genes during early mouse development: implications for imprinting mechanisms. Genes Dev. 1995;9:3097–3108. doi: 10.1101/gad.9.24.3097. [DOI] [PubMed] [Google Scholar]
- 55.Edwards CA, Ferguson-Smith AC. Mechanisms regulating imprinted genes in clusters. Curr Opin Cell Biol. 2007;19:281–289. doi: 10.1016/j.ceb.2007.04.013. [DOI] [PubMed] [Google Scholar]
- 56.Wan LB, Bartolomei MS. Regulation of imprinting in clusters: noncoding RNAs versus insulators. Adv Genet. 2008;61:207–223. doi: 10.1016/S0065-2660(07)00007-7. [DOI] [PubMed] [Google Scholar]