

Abstract
Leukotriene B4 (LTB4) is a pro-inflammatory lipid mediator generated by the enzymes 5-lipoxygenase (5-LO) and LTA4-hydrolase. LTB4 signals primarily through its G protein-coupled receptor BLT1, which is highly expressed on specific leukocyte subsets. Recent genetic studies in humans as well as knockout studies in mice have implicated the leukotriene synthesis pathway in several vascular pathologies. Here we tested the hypothesis that pharmacological inhibition of BLT1 diminishes abdominal aortic aneurysm (AAA) formation, a major complication associated with atherosclerotic vascular disease. Chow-fed Apoe-/- mice were treated with a 4 week infusion of Angiotensin II (AngII, 1000 ng/kg/min) beginning at 10 weeks of age, in a well-established murine AAA model. Administration of the selective BLT1 antagonist CP-105,696 beginning simultaneously with AngII infusion reduced the incidence of AAA formation from 82% to 40% (p<0.05). There was a concordant reduction in maximal aortic diameter from 2.35 mm to 1.56 mm (p<0.05). While administration of the antagonist on day 14 after the onset of AngII infusion diminished lesional macrophage accumulation, it did not significantly alter the size of AAA by day 42. Thus, pharmacological inhibition of BLT1 may ultimately hold clinical promise, but early intervention may be critical.
Introduction
A mounting body of correlative evidence has linked the leukotriene synthesis pathway to several chronic inflammatory conditions, including atherosclerosis. 5-LO and its derivatives are highly expressed within human carotid, aortic, and coronary artery plaques1-3. Furthermore, genetic studies have associated particular variants of 5-LO, its accessory protein FLAP, as well as the enzyme LTA4-hydrolase with stroke and myocardial infarction in humans4-6.
LTB4 is an eicosanoid lipid mediator generated downstream of 5-LO and LTA4-hydrolase. It is known to activate multiple leukocyte subsets leading to cell recruitment, production of reactive oxygen species, and induction of gene expression. LTB4 signals primarily through its high-affinity G protein-coupled receptor (GPCR) BLT1, which is highly expressed in specific subsets of circulating peripheral blood leukocytes, as well as on non-leukocytes including endothelial cells and smooth muscle cells7-9. A low-affinity receptor, BLT2, is also ubiquitously expressed at low levels in many human tissues10, 11.
Functional studies in murine models have begun to address the role of bioactive lipid products in atherogenesis and related vascular complications. Two groups have demonstrated as much as a two-fold reduction in atherosclerotic plaques in Apoe-/-/Blt1-/- compound-deficient mice as compared to Apoe-/- controls7, 12. Animal studies have also addressed the role of the leukotriene pathway in abdominal aortic aneurysm (AAA) formation13. In contrast to luminal plaques, AAAs are characterized by pathological changes in the aortic wall that lead to segmental weakening, progressive dilation, and spontaneous rupture14. Chronic inflammation of the aortic wall and proteinase-mediated degradation of structural matrix proteins are believed to contribute to the development of AAA15, though the precise molecular mechanisms that underlie vessel wall changes remain poorly understood. Zhao et al. demonstrated that 5-LO plays a striking role in aortic aneurysm formation in a cholate-induced model16, though a recent study by the same group using a pharmacological inhibitor of 5-LO had a non-significant effect on AngII-induced AAA17. Our group has demonstrated that Apoe-/-/Blt1-/- mice have decreased aneurysm formation in an Angiotensin II-triggered model of murine AAA formation13. Thus, interventions downstream of 5-LO, specifically isolating the LTB4-BLT1 axis, may have more consistent effects on this pathology.
However, studies performed in mice in which there is complete BLT1 deletion may be limited by developmental confounders. Pharmacological blockade of the receptor represents an important tool to test the role of BLT1-mediated signaling in the adult animal. Such studies are of potential clinical importance, as there are currently no pharmacological interventions available for AAA. Here we used a specific BLT1 antagonist to test whether blockade of the BLT1 receptor in adult animals can modulate the development of AAA, and whether the disease can be reversed once it is established.
Materials and methods
Mice
Male Apoe-/- mice were purchased from Jackson Laboratory, Bar Harbor, ME and fed a chow diet (Prolab). All procedures were approved by Massachusetts General Hospital Center for Comparative Medicine.
Ang II Infusion
Male Apoe-/- mice fed a standard chow diet were treated with AngII (1000 ng/kg/min, Sigma Chemical Co., St. Louis, MO) via 4 week Alzet osmotic pumps (Model 2004; Durect Corporation, Cupertino, CA) beginning at 10 weeks of age as previously described14. Implanted AngII pumps remained in place through subsequent treatment with the inhibitor or vehicle, as indicated.
CP-105,696 compound treatment
The BLT1 inhibitor CP-105,696 was provided by Pfizer18, 19. 100 mg/kg/day of the BLT1 inhibitor was dissolved in 10% ethanol, 0.5% methylcellulose, and 0.5% Tween 80. The CP-105,696 compound was administered daily via gavage concomitant with AngII infusion for 4 weeks in the inhibition study. In the reversal study, the BLT1 inhibitor was also co-administered with the AngII infusion, but began at the end of week 2/start of week 3.
Blood and Plasma Analysis
A total of 1 mL of blood was obtained from experimental mice by right ventricular puncture upon euthanasia. A small aliquot of blood was analyzed for complete blood count (Drew Scientific Group, Oxford, CT), and the remainder was used for enzymatic colorimetric analysis of total cholesterol (Roche Diagnostics, Indianapolis, ID).
Morphometric Analysis
Aortas were dissected from mice, pinned on a black wax board and images were obtained with a Nikon Coolpix camera (Nikon Inc, Melville, NY) attached to an inverted microscope as previously described13. Aortic diameter and area were determined by a Java-based image processing program (ImageJ software, US National Institutes of Health). Measurements were conducted by a single trained observer blinded to the genotype and treatment of the mice.
Chemotaxis assays
For splenocyte isolation, spleens from vehicle- and inhibitor-treated mice were dissected out and passed through a 70 μm filter. 25,000 cells were placed on the top of a 96-well modified Boyden chemotaxis chamber (Neuro-Probe). 32 μL of 10-fold serial dilutions of LTB4 (Cayman Chemical) and SDF-1 (Peprotech) were placed in the bottom wells, separated from the cells by an 8-μm-pore, polyvinylpyrrolidone-free, polycarbonate filter. Following incubation at 37°C in 5% CO2 for 90 minutes, cells that had migrated into the lower chamber were manually counted using a Nikon microscope with a 10× objective.
Ultrasound analysis of mouse aortas
Mice were anesthetized with ketamine/xylazine and laid supine. Aortic imaging was performed using a RMV 704 scanhead with a center frequency of 40MHz (Vevo 770, VisualSonics, Toronto, Canada) as previously described20, 21. The transducer was first oriented parallel to the aorta and the vessel was scanned from the femoral bifurcation to the aortic arch. The renal arteries were identified and long axis images of the infra- and suprarenal aorta were acquired. The transducer was then placed perpendicularly to the aorta and multiple short axis images were acquired at all levels of the aorta. Maximal aortic suprarenal diameter was measured on the short axis view two weeks after AngII infusion, and at 2 and 6 weeks after the start of BLT1 drug inhibitor.
Immunohistochemistry
Suprarenal aortas were embedded in OCT and serial 10 μm sections were cut through the aneurysmal aorta. Immunohistochemistry was performed with an antibody to identify macrophages (Mac-3, BD Biosciences) as previously described13, 22. Gelatinase-A (MMP-2) antibody was kindly provided by Alan Daugherty, University of Kentucky. Negative controls were prepared by substitution with an isotype-matched control antibody or non-immune sera. Quantitative analysis was performed with NIS-Elements AR 2.30 software (Melville, NY). Area of positive staining for Mac-3 and MMP-2 antibody was divided by total lesion area to account for variability in AAA size.
Statistical analysis
Data were analyzed with SigmaStat (Systat software) using Student's t-test and one- or two-way ANOVA as indicated. AAA incidence was analyzed by Fisher's exact test. A probability value of <0.05 was considered statistically significant. All data are reported as mean ± SEM as indicated.
Results
Functional inhibition of BLT1 by CP-105,696
We first established the lowest dose of the drug necessary to functionally inhibit the BLT1 receptor. Of note, prior studies have suggested that doses of CP-105,696 compound greater than 10 mg/kg/day in mammals would ensure plasma levels of at least 5-6 ug/ml, which are necessary to inhibit leukocyte activation19, 23. Following the establishment of dose-response curves of splenocyte chemotaxis to SDF-1 and LTB4 (Fig. 1A, B), we performed chemotaxis experiments on splenocytes isolated from mice that had been treated with the well-described BLT1 inhibitor, CP-105,69618, 19 across a spectrum of doses (0, 10, 100, and 200 mg/kg/day) for 7 days (Fig. 1 C, D, E, F). As expected, LTB4 induced robust chemotaxis of splenocytes from untreated mice in a dose-dependent manner, with the maximal effect reached at 100 nM (Fig. 1B). Splenocyte chemotaxis was relatively unaffected by treatment of mice with CP-105,696 at 10 mg/kg/day (Fig. 1D). By contrast, splenocytes from mice treated with 100 or 200 mg/kg/day CP-105,696 had significantly inhibited chemotaxis to LTB4 (Fig. 1 E, F). We verified the specificity of the antagonist by documenting that chemotaxis to SDF-1, which activates a distinct chemoattractant receptor (CXCR4), was unchanged by administration of CP-105,696. We thus determined that the lowest therapeutic concentration of CP-105,696 to inhibit the LTB4-BLT1 axis was 100 mg/kg/day (Fig. 1E).
Figure 1. Inhibition of BLT1-mediated chemotaxis.

(A, B) Chemotactic response curves of mouse splenocytes to SDF-1 and LTB4.
(C, D, E, F) Chemotaxis assays of isolated splenocytes from mice treated with CP-105,696 for 7 days at the indicated dose. Isolated splenocytes were placed in modified Boyden chambers and chemotaxis was assessed in response to SDF-1 (10 nM) and LTB4 (100 nM). Data shown are from two experiments and are presented as cell number or mean chemotactic index (cells per four-high power fields in duplicate wells moving in response to SDF-1 or LTB4 relative to media alone). *p<0.05 versus media alone.
Aneurysm inhibition by CP-105,696
To assess the effect of early inhibition of BLT1 in aneurysm formation, we infused AngII into 10 week-old Apoe-/- mice receiving CP-105,696 daily at 100 mg/kg/day (n=10 mice) or vehicle (n=11 mice) concomitant with the onset of AngII infusion. Mice were sacrificed after 4 weeks. After Ang II infusion, drug and vehicle treated mice had similar body weights, cholesterol concentrations, and white blood cell counts, thus excluding several potential confounding variables (Table I, Online only Supplementary Material).
Several metrics were used to quantify aneurysm formation in Apoe-/- mice. We first assessed the incidence of aneurysms in both groups, defining an aneurysm as a dilation of the suprarenal aorta greater than 150% of the diameter of the infrarenal aorta, which is entirely spared from enlargement in this model14. Consistent with previously reported findings in the Apoe-/- and Apoe-/-/Blt1-/- mice13, the AAA incidence in the CP-105,696-treated group was 40% as compared to 82% in vehicle-treated mice (p<0.05). We also assessed the extent of aortic dilation in Apoe-/- mice by measuring the maximal suprarenal diameter (Fig. 2B), and found a significant reduction in drug-treated mice compared to the vehicle controls. Representative images from vehicle- and inhibitor-treated mice are shown in Figure 2C. Taken together, these data suggest that pharmacological inhibition of BLT1 abrogates AAA formation in this murine model.
Figure 2. AAA formation in AngII-infused mice treated concomitantly with vehicle or CP-105,696.
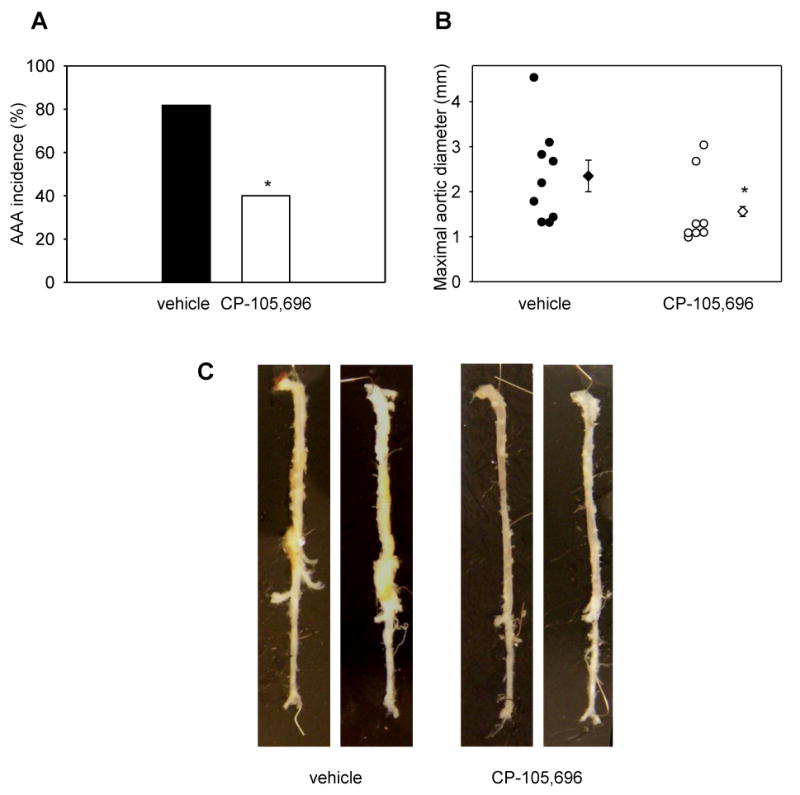
Apoe-/- mice were treated with AngII concomitant with vehicle or CP-105,696 for 4 weeks. (A) AAA incidence (*p<0.05), (B) maximal aortic diameter (*p<0.05), (C) Representative images of aortas from mice treated with vehicle or CP-105,696 from the prevention study. Circles represent individual mice, diamonds represent means and bars represent SEMs. The experiment included n=11 mice in the vehicle group, and n=10 mice in the CP-105,696 group. Two mice from the vehicle-control and two mice from the inhibitor-treated group died of AAA rupture.
The effect of CP-105,696 in the progression of aortic dilation
To assess the extent to which BLT1 inhibition might ameliorate the course of aortic dilation once established, we infused mice with AngII and followed them with serial examinations using ultrasound (Fig. 3), which can be used to accurately monitor AAA formation in this model20, 21. The transducer was oriented in the short axis and sequential images were taken from the infrarenal segment to the thoracic aorta, and maximal aortic suprarenal diameter was measured from the acquired images. After 2 weeks of AngII infusion, mice were then randomly split in two groups: n=10 mice received the BLT1 inhibitor, while n=12 mice received the vehicle daily for 6 weeks. All of the mice developed AAA as assessed by ultrasound. Ultrasound was performed again after 2 and 6 weeks of daily CP-105,696 treatment, at which time the mice were sacrificed. As in the prevention trial, both groups of mice had similar body weights, cholesterol concentrations, and white blood cell counts (Table II Online only Supplementary Material). Both groups of mice had similar aortic diameters at the start of the BLT1 inhibitor treatment. We observed a modest increase in aortic expansion on top of the previously established AAA in both groups of mice after 2 weeks of drug or vehicle treatment. After 6 weeks, we documented a highly significant reduction in aortic macrophage infiltration and MMP-2 expression in the animals receiving the BLT1 inhibitor (Fig. 4 A, B, C, D). We did not detect any positive stain in the isotype-matched IgG control-stained AAA tissues (Fig. 4 A, B, right panels). However, the reduction in inflammation did not translate into modulation of aortic size, as aortic diameters were the same in both the vehicle- and drug-treated groups at the 6 week time point (Fig. 5A). Of note, in vivo (ultrasound) and ex vivo (pathology) measurements taken at 6 weeks were highly correlated (R=0.918, Fig. 5B). Representative ultrasound images of aortas acquired at the beginning of treatment, and at 2 and 6 weeks of treatment are shown in Fig. I Online only Supplementary Material.
Figure 3. Experimental design of AAA reversal study.

Mice were infused with AngII for 2 weeks and then evaluated by ultrasound. Mice that had developed AAA at 2 weeks were randomly assigned to CP-105,696 (n=10) or vehicle control (n=12). All mice were evaluated again by ultrasound at weeks 4 and 8 after the onset of AngII infusion, and sacrificed at week 8 and evaluated for CBC, total cholesterol and aortic quantitation. The dashed line represents the AngII pumps that remained in place throughout the treatment with CP-105,696 for a total of 8 weeks.
Figure 4. Diminished macrophage accumulation and MMP-2 expression in aortas of mice treated with CP-105,696.
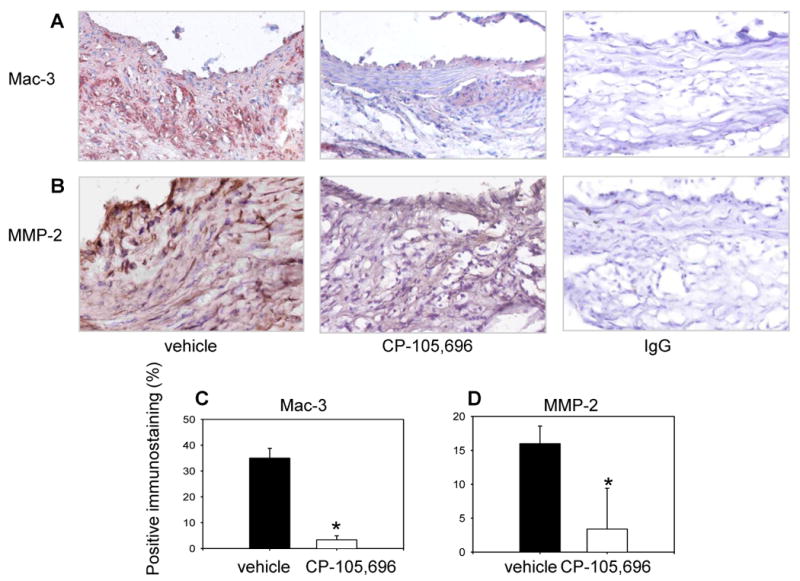
(A) Representative images of macrophage (Mac-3) and (B) MMP-2 stain show intense macrophage infiltration and MMP-2 expression in the vehicle control and less macrophage accumulation and MMP-2 expression in the inhibitor-treated aorta (middle panels). No positive stain is detected in the isotype-matched control IgG for Mac-3 and MMP-2 stains (right panels). Magnification 20×. (C, D) Quantification of Mac-3 and MMP-2 immunostaining from vehicle- and inhibitor-treated AAA (*p<0.001).
Figure 5. Ultrasound measurements of aortas from mice treated with vehicle or CP-105,696 beginning 2 weeks after AngII infusion.

N=12 mice were treated with vehicle and n=10 mice were treated with CP-105,696 at 2 weeks after AngII pumps were placed. (A) Maximal aortic diameters were measured periodically in the suprarenal aorta throughout treatment with vehicle or CP-105,696. Circles represent means and bars represent SEMs. (B) Correlation of maximal aortic diameter measurements by ultrasound versus histological measurements after dissection at 8 weeks (R=0.918).
Discussion
Animal and human studies have increasingly implicated the leukotriene synthesis pathway in chronic inflammatory diseases, including atherosclerosis and its related complications. Here we have shown that BLT1 blockade in the adult animal confers a decrease in aneurysm incidence as well as a concordant reduction in aortic dilation. While BLT1 inhibition diminished aortic macrophage content in established AAA, it did not reverse AAAs at 6 weeks after treatment.
Our findings are consistent with recent studies linking leukotrienes specifically with AAA formation. In a cholate diet-triggered model of AAA, 5-LO deficiency strikingly diminished aneurysmal dilation in a hyperlipidemic mouse background16. However, emerging evidence suggests that atherogenesis and aneurysm formation may be inherently different processes, and thus modulation of the same signal in two disease models may not necessarily demonstrate concordant results22, 24. While 5-LO deficiency markedly attenuated aneurysm formation, there was no significant effect on the formation of lipid-rich atherosclerotic plaques in a comprehensive analysis16. Recent studies have demonstrated that 5-LO generates both pro-inflammatory as well as anti-inflammatory products [e.g., LXA(4) metabolites].25 Such work underscores the need for precise interventions in the leukotriene pathway to best target inflammatory processes. Further downstream of 5-LO, genetic deficiency of the BLT1 receptor decreased both hyperlipidemia-induced atherosclerosis7 and AngII-induced AAA13. The abrogation of BLT1 signaling had similar effects on both overall plaque development and AAA formation—in contrast to results seen with “upstream” 5-LO inhibition. Thus, pharmaceutical intervention aimed at this receptor may have multiple salutary effects on the vasculature.
The use of knockout mice in the prior studies limited the scope of our prior investigation to alterations that precede the onset of the disease model. Furthermore, developmental confounders and aberrant compensatory pathways can also affect studies in knockout mice. The present study thus extends prior work, by demonstrating that BLT1 blockade in the adult animal also modified AAA formation. We were interested to find that the effects of the drug on AAA incidence and size were extremely similar to the genetic modulation of the LTB4-BLT1 axis. Thus, the effects seen in the Blt1-/- mice in previous work were likely due to modulation of the response to injury in the adult animal, and not secondary to effects on developmental pathways that preceded the onset of the stressor.
While institution of the inhibitor with the onset of the AngII infusion blunted the AAA response, treatment with the inhibitor beginning two weeks after the start of AngII treatment failed to engender AAA reversal. By contrast, administration of a JNK inhibitor caused regression of aneurysmal diameter in two models of murine AAA26, 27. After 6 weeks of therapy in our study, there was a reduction in macrophage recruitment and MMP-2 expression in aortas of mice treated with the BLT1 inhibitor, though aortic size did not change. We hypothesize that the anti-inflammatory effects of the BLT1 inhibition occurred after the initiation of AngII-induced elastin degradation and as such were not sufficient to engender aortic aneurysm regression. Our findings are consistent with the notion that the pathogenesis of aneurysmal disease is complex and multifactorial. For example, it has been shown that macrophages are present in the early stages of AngII-induced aneurysms and precede the medial degradation suggesting a contributing role24. However other studies have demonstrated that macrophage depletion actually predisposes mice to aortic intra-mural hematomas28. Thus, the timing of interventions in complex inflammatory diseases is critical, and further work is clearly necessary.
Future studies must also determine whether longer treatment can ultimately cause regression of aortic dilation. Whether a potent anti-inflammatory response might ultimately translate into diminished aortic rupture must ultimately be examined as well. In conclusion, our data have extended the role of the LTB4-BLT1 axis as critical to aneurysm formation in mice. As with classic peptide chemokines, the pleiotropic effects of lipid chemoattractants in disease contexts are increasingly appreciated. These findings are clinically relevant because small-molecule antagonists to BLT1 are under development and would thus target multiple vessel wall cell types and multiple vascular pathologies.
Supplementary Material
Footnotes
Publisher's Disclaimer: This is a PDF file of an unedited manuscript that has been accepted for publication. As a service to our customers we are providing this early version of the manuscript. The manuscript will undergo copyediting, typesetting, and review of the resulting proof before it is published in its final citable form. Please note that during the production process errors may be discovered which could affect the content, and all legal disclaimers that apply to the journal pertain.
References
- 1.Spanbroek R, Grabner R, Lotzer K, et al. Expanding expression of the 5-lipoxygenase pathway within the arterial wall during human atherogenesis. Proc Natl Acad Sci U S A. 2003;100:1238–43. doi: 10.1073/pnas.242716099. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.De Caterina R, Mazzone A, Giannessi D, et al. Leukotriene B4 production in human atherosclerotic plaques. Biomed Biochim Acta. 1988;47:S182–5. [PubMed] [Google Scholar]
- 3.Qiu H, Gabrielsen A, Agardh HE, et al. Expression of 5-lipoxygenase and leukotriene A4 hydrolase in human atherosclerotic lesions correlates with symptoms of plaque instability. Proc Natl Acad Sci U S A. 2006;103:8161–6. doi: 10.1073/pnas.0602414103. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Dwyer JH, Allayee H, Dwyer KM, et al. Arachidonate 5-lipoxygenase promoter genotype, dietary arachidonic acid, and atherosclerosis. N Engl J Med. 2004;350:29–37. doi: 10.1056/NEJMoa025079. [DOI] [PubMed] [Google Scholar]
- 5.Helgadottir A, Manolescu A, Helgason A, et al. A variant of the gene encoding leukotriene A4 hydrolase confers ethnicity-specific risk of myocardial infarction. Nat Genet. 2006;38:68–74. doi: 10.1038/ng1692. [DOI] [PubMed] [Google Scholar]
- 6.Helgadottir A, Manolescu A, Thorleifsson G, et al. The gene encoding 5-lipoxygenase activating protein confers risk of myocardial infarction and stroke. Nat Genet. 2004;36:233–9. doi: 10.1038/ng1311. [DOI] [PubMed] [Google Scholar]
- 7.Heller EA, Liu E, Tager AM, et al. Inhibition of atherogenesis in BLT1-deficient mice reveals a role for LTB4 and BLT1 in smooth muscle cell recruitment. Circulation. 2005;112:578–86. doi: 10.1161/CIRCULATIONAHA.105.545616. [DOI] [PubMed] [Google Scholar]
- 8.Back M, Bu DX, Branstrom R, Sheikine Y, Yan ZQ, Hansson GK. Leukotriene B4 signaling through NF-kappaB-dependent BLT1 receptors on vascular smooth muscle cells in atherosclerosis and intimal hyperplasia. Proc Natl Acad Sci U S A. 2005;102:17501–6. doi: 10.1073/pnas.0505845102. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Watanabe S, Yamasaki A, Hashimoto K, et al. Expression of functional leukotriene B(4) receptors on human airway smooth muscle cells. J Allergy Clin Immunol. 2009;124:59–65. e1–3. doi: 10.1016/j.jaci.2009.03.024. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Yokomizo T, Kato K, Terawaki K, Izumi T, Shimizu T. A second leukotriene B(4) receptor, BLT2. A new therapeutic target in inflammation and immunological disorders. J Exp Med. 2000;192:421–32. doi: 10.1084/jem.192.3.421. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Tager AM, Luster AD. BLT1 and BLT2: the leukotriene B(4) receptors. Prostaglandins, leukotrienes, and essential fatty acids. 2003;69:123–34. doi: 10.1016/s0952-3278(03)00073-5. [DOI] [PubMed] [Google Scholar]
- 12.Subbarao K, Jala VR, Mathis S, et al. Role of leukotriene B4 receptors in the development of atherosclerosis: potential mechanisms. Arteriosclerosis, thrombosis, and vascular biology. 2004;24:369–75. doi: 10.1161/01.ATV.0000110503.16605.15. [DOI] [PubMed] [Google Scholar]
- 13.Ahluwalia N, Lin AY, Tager AM, et al. Inhibited aortic aneurysm formation in BLT1-deficient mice. J Immunol. 2007;179:691–7. doi: 10.4049/jimmunol.179.1.691. [DOI] [PubMed] [Google Scholar]
- 14.Daugherty A, Manning MW, Cassis LA. Angiotensin II promotes atherosclerotic lesions and aneurysms in apolipoprotein E-deficient mice. The Journal of clinical investigation. 2000;105:1605–12. doi: 10.1172/JCI7818. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Thompson RW, Geraghty PJ, Lee JK. Abdominal aortic aneurysms: basic mechanisms and clinical implications. Current problems in surgery. 2002;39:110–230. doi: 10.1067/msg.2002.121421. [DOI] [PubMed] [Google Scholar]
- 16.Zhao L, Moos MP, Grabner R, et al. The 5-lipoxygenase pathway promotes pathogenesis of hyperlipidemia-dependent aortic aneurysm. Nature medicine. 2004;10:966–73. doi: 10.1038/nm1099. [DOI] [PubMed] [Google Scholar]
- 17.Cao RY, Adams MA, Habenicht AJ, Funk CD. Angiotensin II-induced abdominal aortic aneurysm occurs independently of the 5-lipoxygenase pathway in apolipoprotein E-deficient mice. Prostaglandins & other lipid mediators. 2007;84:34–42. doi: 10.1016/j.prostaglandins.2007.03.005. [DOI] [PubMed] [Google Scholar]
- 18.Showell HJ, Breslow R, Conklyn MJ, Hingorani GP, Koch K. Characterization of the pharmacological profile of the potent LTB4 antagonist CP-105,696 on murine LTB4 receptors in vitro. British journal of pharmacology. 1996;117:1127–32. doi: 10.1111/j.1476-5381.1996.tb16706.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Aiello RJ, Bourassa PA, Lindsey S, Weng W, Freeman A, Showell HJ. Leukotriene B4 receptor antagonism reduces monocytic foam cells in mice. Arteriosclerosis, thrombosis, and vascular biology. 2002;22:443–9. doi: 10.1161/hq0302.105593. [DOI] [PubMed] [Google Scholar]
- 20.Barisione C, Charnigo R, Howatt DA, Moorleghen JJ, Rateri DL, Daugherty A. Rapid dilation of the abdominal aorta during infusion of angiotensin II detected by noninvasive high-frequency ultrasonography. J Vasc Surg. 2006;44:372–6. doi: 10.1016/j.jvs.2006.04.047. [DOI] [PubMed] [Google Scholar]
- 21.Martin-McNulty B, Vincelette J, Vergona R, Sullivan ME, Wang YX. Noninvasive measurement of abdominal aortic aneurysms in intact mice by a high-frequency ultrasound imaging system. Ultrasound in medicine & biology. 2005;31:745–9. doi: 10.1016/j.ultrasmedbio.2005.02.012. [DOI] [PubMed] [Google Scholar]
- 22.King VL, Lin AY, Kristo F, et al. Interferon-gamma and the interferon-inducible chemokine CXCL10 protect against aneurysm formation and rupture. Circulation. 2009;119:426–35. doi: 10.1161/CIRCULATIONAHA.108.785949. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Liston TE, Conklyn MJ, Houser J, et al. Pharmacokinetics and pharmacodynamics of the leukotriene B4 receptor antagonist CP-105,696 in man following single oral administration. British journal of clinical pharmacology. 1998;45:115–21. doi: 10.1046/j.1365-2125.1998.00646.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Saraff K, Babamusta F, Cassis LA, Daugherty A. Aortic dissection precedes formation of aneurysms and atherosclerosis in angiotensin II-infused, apolipoprotein E-deficient mice. Arteriosclerosis, thrombosis, and vascular biology. 2003;23:1621–6. doi: 10.1161/01.ATV.0000085631.76095.64. [DOI] [PubMed] [Google Scholar]
- 25.Levy BD, De Sanctis GT, Devchand PR, et al. Multi-pronged inhibition of airway hyper-responsiveness and inflammation by lipoxin A(4) Nature medicine. 2002;8:1018–23. doi: 10.1038/nm748. [DOI] [PubMed] [Google Scholar]
- 26.Yoshimura K, Aoki H, Ikeda Y, Furutani A, Hamano K, Matsuzaki M. Regression of abdominal aortic aneurysm by inhibition of c-Jun N-terminal kinase in mice. Annals of the New York Academy of Sciences. 2006;1085:74–81. doi: 10.1196/annals.1383.031. [DOI] [PubMed] [Google Scholar]
- 27.Yoshimura K, Aoki H, Ikeda Y, et al. Regression of abdominal aortic aneurysm by inhibition of c-Jun N-terminal kinase. Nature medicine. 2005;11:1330–8. doi: 10.1038/nm1335. [DOI] [PubMed] [Google Scholar]
- 28.Babamusta F, Rateri DL, Moorleghen JJ, Howatt DA, Li XA, Daugherty A. Angiotensin II infusion induces site-specific intra-laminar hemorrhage in macrophage colony-stimulating factor-deficient mice. Atherosclerosis. 2006;186:282–90. doi: 10.1016/j.atherosclerosis.2005.08.006. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.