

Abstract
Background
Multiple myeloma is an incurable malignancy of bone marrow plasma cells. Progression of multiple myeloma is accompanied by an increase in bone marrow angiogenesis. Studies from our laboratory suggest a role for the CXCL12 chemokine in this process, with circulating levels of CXCL12 correlating with bone marrow angiogenesis in patients with multiple myeloma. While the mechanisms responsible for aberrant plasma cell expression of CXCL12 remain to be determined, studies in other systems suggest a role for hypoxia and hypoxia-inducible transcription factors.
Design and Methods
The expression of hypoxia-inducible factor protein was examined in patients’ bone marrow biopsy specimens using immunohistochemistry. The hypoxic regulation of CXCL12 was examined in multiple myeloma plasma cell lines using polymerase chain reaction and western blotting. The role of hypoxia-inducible factors-1 and -2 in the regulation of CXCL12 expression was examined using over-expression and short hairpin RNA knockdown constructs, electrophoretic mobility shift assays and chromatin immunoprecipitation. The contribution of CXCL12 to hypoxia-induced angiogenesis was examined in vivo using a subcutaneous murine model of neovascularization.
Results
Strong hypoxia-inducible factor-2 protein expression was detected in CD138+ multiple myeloma plasma cells in patients’ biopsy specimens. Prolonged exposure to hypoxia strongly up-regulated CXCL12 expression in multiple myeloma plasma cells and hypoxia-inducible factor-2 was found to play a key role in this response. Promoter analyses revealed increased hypoxia-inducible factor-2 binding to the CXCL12 promoter under hypoxic conditions. Over-expression of hypoxia-inducible factor in multiple myeloma plasma cells strongly induced in vivo angiogenesis, and administration of a CXCL12 antagonist decreased hypoxia-inducible factor-induced angiogenesis.
Conclusions
Hypoxia-inducible factor-2 is a newly identified regulator of CXCL12 expression in multiple myeloma plasma cells and a major contributor to multiple myeloma plasma cell-induced angiogenesis. Targeting the hypoxic niche, and more specifically hypoxia-inducible factor-2, may represent a viable strategy to inhibit angiogenesis in multiple myeloma and progression of this disease.
Keywords: multiple myeloma, CXCL12, hypoxia, HIF-2
Introduction
Multiple myeloma (MM) is an incurable hematologic malignancy characterized by the clonal proliferation of malignant plasma cells in the bone marrow. Accounting for approximately 1% of all cancers, MM is the second most common hematologic malignancy after non-Hodgkin’s lymphoma. As for all tumors, the survival and expansion of MM plasma cells is dependent upon an adequate supply of oxygen and nutrients, and the acquisition of an angiogenic phenotype is a key event in the progression from monoclonal gammopathy of undetermined significance (MGUS) and indolent MM to active MM.1,2
CXCL12 is a constitutively expressed chemokine which binds primarily to the CXCR4 receptor and regulates cell growth, chemotaxis, myelopoiesis, lymphopoiesis, and development of the nervous and cardiovascular systems.3–5 CXCL12 is highly expressed by MM plasma cells,6 and circulating levels of CXCL12 are higher in the peripheral blood of MM patients than in age-matched normal donors and MGUS patients.6,7 CXCL12 is an important mediator of several aspects of MM biology including transendothelial migration,8,9 MM plasma cell migration and retention within the bone marrow,10,11 angiogenesis,7 and osteoclastic bone resorption.6 Recent animal studies involving systemic injection of labeled MM plasma cells have also demonstrated that blocking the CXCL12/CXCR4 axis leads to a 20% reduction in bone marrow tumor burden.11
Unlike most other organs, the bone marrow microenvironment is physiologically hypoxic, a pre-requisite for normal bone marrow hematopoiesis.12 It is well established that hypoxia is an important selective force in the evolution of tumor cells,13 and elevated expression of the hypoxia-inducible transcription factors HIF-1 and HIF-2 has been documented in several human cancers.14,15 HIF-1 and HIF-2 mediate adaptive responses to hypoxia by inducing the transcription of genes associated with erythropoiesis, glycolytic metabolism, cell survival and angiogenesis. While the role of hypoxia in the pathogenesis of hematologic malignancies has yet to be elucidated, recent animal studies have shown that changes in oxygen levels within the bone marrow microenvironment support the survival and expansion of MM plasma cells.16 Furthermore, some drugs active in MM, such as bortezomib and lenalidomide, are believed to exert their effects, in part, by interfering with hypoxia-induced signaling cascades.17,18
HIF-1 and HIF-2 are heterodimers composed of an inducible α-subunit and a constitutively-expressed β-subunit called aryl hydrocarbon receptor nuclear translocator (ARNT). Under normoxic conditions, HIF-α subunits are functionally repressed and undergo rapid proteosomal degradation (t1/2=5 min).19 However under hypoxic conditions, these processes are abrogated and the stabilized HIF-α translocates to the nucleus to dimerize with ARNT. HIF-1 and HIF-2 bind to the same DNA consensus motif; however, they have overlapping but distinct target gene specificities and distinct, non-redundant physiological roles.20,21
In 2002, Hitchon et al. showed for the first time that CXCL12 expression is up-regulated by hypoxia in human synovial fibroblasts.22 While subsequent studies extended this finding to other cell types,23–26 the effect of hypoxia on CXCL12 expression in MM plasma cells has not been investigated. We, therefore, studied the expression of HIF-1, HIF-2 and CXCL12 in relation to hypoxia in MM and the contribution of CXCL12 to hypoxia-induced angiogenesis.
Design and Methods
Cell cultures
LP-1 cells (a human myeloma cell line) were cultured in RPMI medium supplemented with 10% fetal calf serum, 2 mM L-glutamine, 1 mM sodium pyruvate, 15 mM HEPES and 50 IU/mL penicillin-streptomycin in a humidified atmosphere at 37°C. Where specified, hypoxic culture conditions were established (less than 1% oxygen) using an anaerobic sachet (Oxoid, UK).
Immunohistochemical staining
Paraffin-embedded sections (5 μm) of MGUS (n=8) or MM (n=7) trephine specimens collected at the time of diagnosis at the Royal Adelaide Hospital were immunostained with antibodies against CXCL12 (sc-6193, Santa Cruz, CA, USA), HIF-1α (NB100–449, Novus Biologicals, CO, USA), HIF-2α (NB100–132, Novus Biologicals) and CD138 (M7228, Dako, Denmark) as previously described.6 Studies were performed with Institutional Ethics approval following written, informed consent. Images were captured using a NanoZoomer Slide Scanner (Hamamatsu, Japan).
Stable transduction of LP-1 cells
To generate stable over-expressing cell lines, full-length cDNA encoding human CXCL12,27 HIF-1α,28 and HIF-2α29 were cloned into pRUF-IRES-GFP to generate pRUF-IRES-GFP-HIF-1α, pRUF-IRES-GFP-HIF-2α and pRUF-IRES-GFP-CXCL12. Following retroviral infection of LP-1 cells, pooled cell lines were established from the top 30% of GFP-expressing cells as previously described.27 To generate stable knock-downs, RNA duplexes targeting human HIF-1α (CCATGAGGAAATGAGAGAAAT-GCTT), human HIF 2α (GGGGGCTGTGTCTGAGAAGAGT) or a scrambled control (CCAAGGAGTAAGAGATAAAG-GTC)30,31 were cloned into the pFIV-H1-copGFP lentiviral vector (System Biosciences, CA, USA), and clonal cell lines generated from the top 10% of GFP-expressing cells using preparative cell sorting and single-cell deposition. For in vivo studies, cells were co-transduced with the SFG-nes-TGL luciferase vector32 to enable bioluminescent detection of these cells.
Real-time polymerase chain reaction
RNA was reverse transcribed from 1 μg of total RNA using Superscript III according to the manufacturer’s instructions (Invitrogen, CA, USA) and real-time polymerase chain reaction (PCR) was performed on a Rotor-Gene 3000 instrument (Corbett Life Science, Australia) using the following primers: β2-microglobulin Fwd 5′-aggctatccagcgtactcca-3′ and Rev 5′-tcaatgtcggatggatgaaa-3′; human CXCL12 Fwd 5′-atgcccatgccgattcttcg-3′ and Rev 5′-gtctgttgttgttcttcagcc-3′; human HIF-1α Fwd 5′-ccacctatgacctgcttggt-3′ and Rev 5′-tgtcctgtggtgacttgtcc-3′; human HIF-2α Fwd 5′-ctctcctcagtttgctctgaaaa-3′ and Rev 5′-gtcgcagggatgagtgaagt-3′; human vascular endothelial growth factor (VEGF) Fwd 5′-atgccaagtggtcccagg-3′ and Rev 5′-cacacaggatggcttgaaga-3′; human GLUT-1 Fwd 5′-ggccaagagtgtgctaaagaa-3′ and Rev 5′-cagcgttgatgccagaca-3′; human CXCR4 Fwd 5′-cagcaggtagcaaagtgacg-3′ and Rev 5′-gtagatggtgggcaggaaga-3′, as previously described.27 Changes in gene expression were calculated relative to β2-microglobulin using the 2−ΔCt method.33
Western immunoblotting
Whole cell extracts (100 μg) were separated by 8–10% sodium dodecyl sulfate polyacrylamide gel electrophoresis (SDS-PAGE) under reducing conditions and transferred to a polyvinylidene fluoride membrane. Immunoblotting was performed using antibodies to HIF-1α (610959, BD Bioscience, CA, USA), HIF-2α (NB100–122, Novus Biologicals), and α-tubulin (ab6160, Abcam, MA, USA). Following incubation with the appropriate alkaline phosphatase-conjugated secondary antibodies, membranes were developed with ECF (GE Healthcare, UK).
CXCL12 enzyme-linked immunosorbent assay
CXCL12 protein levels in culture media were measured using a commercial immunoassay (R&D Systems, MN, USA) as previously described,6,7 and data were normalized to the total protein content of the cells from which the supernatant was collected.
Luciferase assay
The proximal CXCL12 promoter was amplified from LP-1 genomic DNA (Fwd: 5′-gcgctcgagccatctaacggccaaagtgg-3′ and Rev: 5′-gcgaagcttggctgacggagagtgaaagtg-3′) using Pfu turbo (Stratagene, CA, USA) according to the manufacturer’s instructions, and ligated into the pGL3-basic vector (E1751, Promega, WI, USA) to generate pGL3b-CXCL12. LP-1 cells (4.5×106) were electroporated (Bio-Rad Gene Pulser, 270V/960μF) in 500 μL in RPMI medium (supplemented with 20% fetal calf serum) with 5 μg of reporter plasmid and 10 μg of expression plasmid. Twenty-four hours after transfection, cells were cultured under normoxic or hypoxic conditions for 48 h, and their luciferase activity was assayed as previously described.34
Chromatin immunoprecipitation
Chromatin immunoprecipitation assays were performed using the EZ-Magna ChIP kit (Millipore, MA, USA) according to the manufacturer’s instructions. Briefly, LP-1 cells (1×107/treatment) were cultured under normoxic or hypoxic conditions for 48 h, then cross-linked in 1% formaldehyde for 10 min at room temperature and quenched with 125 mM glycine for 5 min. Cells were washed five times in chilled phosphate-buffered saline, sonicated for 30 min (Diagenode, Belgium, 30 s pulses, 30 s rests) and immunoprecipated with magnetic beads and 5 μg primary antibody (ab199, Abcam) overnight at 4°C with rotation. The next day, complexes were eluted, sequentially washed in low salt buffer, high salt buffer, LiC buffer and TE buffer (5 min each with rotation), then eluted with 100 μg/mL proteinase K at 62°C for 2 h with rotation. Eluted DNA was purified and subjected to PCR analysis using primers directed against the HBS1 region of the CXCL12 promoter: Fwd 5′-tctaacggccaaagtggttt-3′ and Rev 5′-gccacctctctgtgtccttc-3′.
Electrophoretic mobility shift assay
The oligonucleotide for HBS1 of the CXCL12 promoter (5′-gggacagggacgtgtccccaggg-3′) was purchased from Geneworks (Australia) and the full-length product purified from non-denaturing polyacrylamide gels following established protocols.35 Single-stranded DNA probes were prepared by end-labeling 100ng of oligonucleotide with T4 polynucleotide kinase and [γ-32P] ATP (Geneworks) followed by gel purification.
Nuclear extracts were prepared from LP-1 cells following 48 h of normoxic or hypoxic culture as previously described.36 Gel retardations were performed using 0.25 ng double-stranded 32P-labeled oligonucleotide probe in a 10 μL reaction mix containing 10 mM Tris-HCl, pH 7.5, 50 mM NaCl, 1 mM MgCl2, 1 mM EDTA, 5 mM DTT, 5% glycerol, 0.25 μg poly dIdC and 5 μg nuclear extract. Reaction mixes were incubated at 4°C for 30 min and resolved on 4% non-denaturing polyacrylamide gels run in 0.5x TBE buffer. For antibody blocking experiments, protein and antibody (5 μg or 10 μg) were incubated for 5 min at room temperature before adding 32P-labeled probe for 20 min prior to gel loading. Gels were visualized using a Storage Phosphor Screen (GE Healthcare).
Implantation of modified LP-1 cell lines into mice
Cells (5×106/implant) were suspended in 200 μL chilled serum-free RPMI 1640 medium, mixed with 200 μL chilled Matrigel matrix (BD Bioscience) and subcutaneously injected into the right ventral flank of 6-week old female BALB/c nude mice. An equivalent implant containing no cells was injected into the left ventral flank. After 14 days, mice were killed humanely and Matrigel plugs were photographed and removed, and the hemoglobin content determined. Where specified, osmotic pumps (DURECT Corporation, CA, USA) containing 100 μL of the CXCR4 antagonist, T140 (4F-benzoyl-TN14003, 80 mg/mL in phosphate-buffered saline), were subcutaneously implanted in the upper dorsum 2 days prior to cell implantation.
In vivo bioluminescence imaging
Ongoing assessment of tumor growth was performed using bioluminescence imaging as previously described.27
Hemoglobin assessment
Excised implants were sonicated for 15 min (Diagenode, 30 s pulses, 30 s rests) in 300 μL water and centrifuged at 16,000 x g for 90 min at 4°C to remove debris. The hemoglobin content was then assessed according to instructions from Sigma (MO, USA). Briefly, homogenised tumor supernatants (50 μL/well) were added to Drabkin’s solution (200 μL/well), and absorbance was read at 540 nm after 15 min. A standard curve was generated using bovine hemoglobin (Sigma).
The hemoglobin content of each cell-containing implant was initially normalized to that of the corresponding empty implant from each mouse. This value was then normalized to the bioluminescence reading obtained at the time of sacrifice, to relate the angiogenesis assessment to the number of viable cells present in the implant.
Statistical analyses
Experiments were performed in triplicate, and data are presented as mean ± standard error of measurement (SEM). Statistical analyses were performed using a one-way ANOVA with Dunnett’s post-hoc test using SigmaStat® 3.0 software (Systat, IL, USA). In all cases, P values less than 0.05 were considered statistically significant.
Results
Expression of hypoxia-inducible factor and CXCL12 proteins in the bone marrow
Sections of bone marrow trephine specimens from patients with MGUS or MM were co-immunostained with an antibody to CD138 (a marker of MM plasma cells) and antibodies to HIF-1α, HIF-2α or CXCL12. In keeping with the findings of others,15 weak HIF-1α expression was detected in numerous cells throughout the bone marrow (Figures 1A and 1B, panels c and d). In contrast, HIF-2α expression was restricted to CD68+ macrophages (data not shown) and CD138+ MM plasma cells (Figure 1B, panels e and f). Importantly, HIF-2α and CXCL12 proteins were both expressed in CD138+ MM plasma cells (Figure 1B, panels a, b, e and f).
Figure 1.
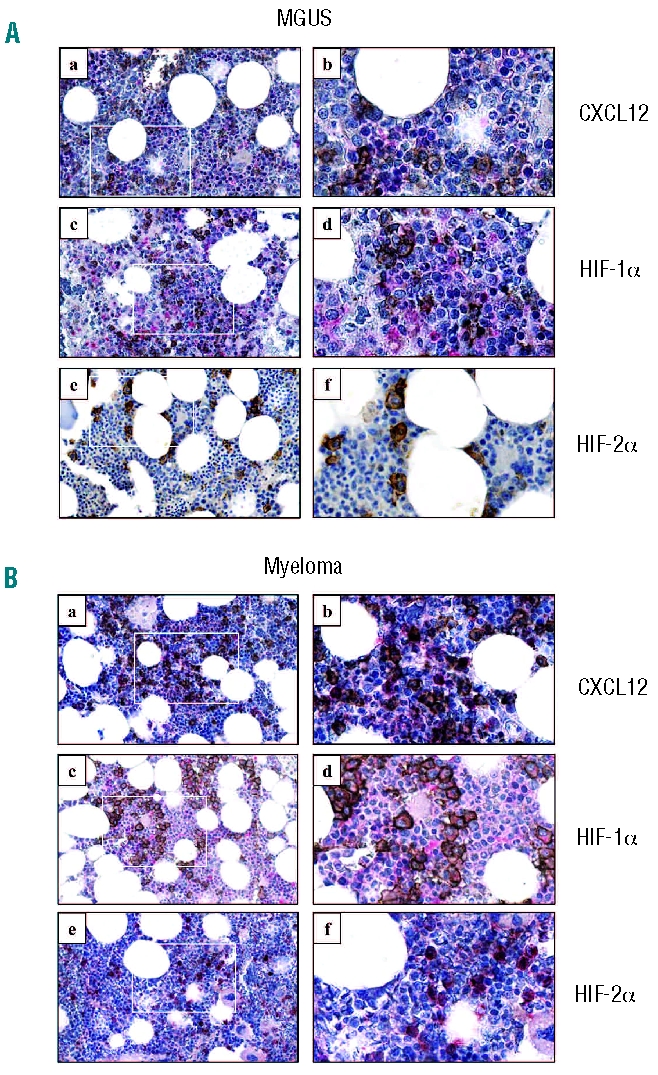
Expression of CXCL12, HIF-1α and HIF-2α in patients’ trephine specimens. Bone marrow trephine sections from (A) MGUS and (B) MM patients at diagnosis were co-stained with CXCL12 (a and b, pink), HIF-1α (c and d, pink) or HIF-2α (e and f, pink) and CD138 (all sections, brown). Original magnifications x40 (a, c and e) and x200 (b, d and f); colors corrected after acquisition with Adobe Photoshop.
The hypoxic regulation of CXCL12 expression in the multiple myeloma cell line, LP-1
To examine the effect of hypoxia on CXCL12 expression in MM plasma cells, levels of CXCL12 mRNA were measured in the MM plasma cell line, LP-1, following 6, 24 and 48 h of normoxic or hypoxic culture. Strong up-regulation of CXCL12 mRNA was observed in response to 24 and 48 h of hypoxia (Figure 2A). This delayed hypoxic induction of CXCL12 expression was observed in three of four MM cell lines tested (U266, JIMI, and LP-1, Online Supplementary Figure S1), with one cell line (RPMI-8226) exhibiting no response. Using enzyme-linked immunosorbent assays, levels of CXCL12 protein were measured in LP-1 culture media following 72 h of normoxic or hypoxic culture (Figure 2B), and higher levels of CXCL12 protein were detected in hypoxic culture media (812±15.52 pg/mL) than in normoxic culture media (364±17.52 pg/mL). While hypoxic up-regulation of CXCL12 protein expression was observed in the U266 and JIMI cell lines (data not shown), the LP-1 cell line was selected for all the subsequent studies outlined below.
Figure 2.
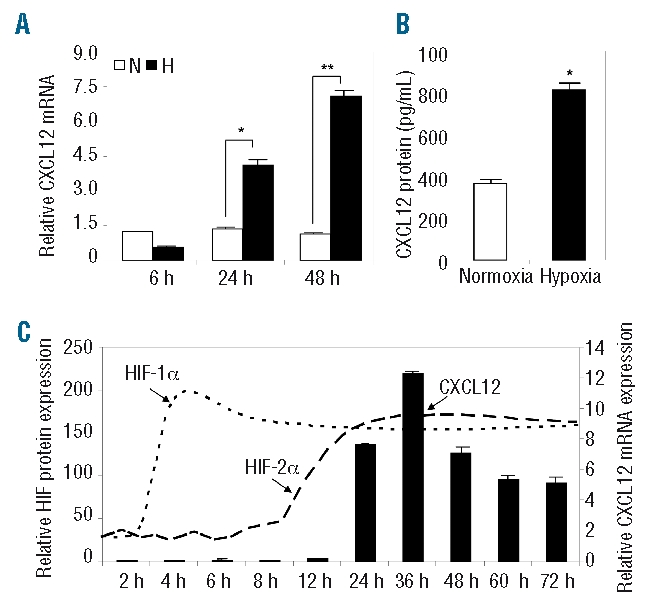
Hypoxic regulation of CXCL12 expression in LP-1 cells. (A) Levels of CXCL12 mRNA expression were assessed in LP-1 cells following 6, 24 and 48 h of normoxic (white bars) or hypoxic (black bars) culture. Columns, mean (n=3); bars, SEM. *P<0.05, **P<0.001, compared to normoxia. (B) Levels of CXCL12 protein were measured in LP-1 conditioned medium following 72 h of normoxic or hypoxic culture. Columns, mean (n=3); bars, SEM. *P<0.001, compared to normoxia. (C) The hypoxic induction of CXCL12 (black bars) mRNA, and HIF-1α (dotted line) and HIF-2α (dashed line) protein expression was examined in LP-1 cells over 72 h. Columns and dashed lines, mean (n=3); bars, SEM.
To examine the kinetics of the hypoxic induction of CXCL12 in MM plasma cells, LP-1 CXCL12 mRNA expression was measured following 2, 4, 6, 8, 12, 24, 36, 48, 60 and 72 h of normoxic or hypoxic culture. A minimum of 24 h of continuous hypoxic exposure was required to up-regulate CXCL12 mRNA (Figure 2C). The hypoxic induction of other HIF target genes (GLUT1, CXCR4 and VEGF) was also examined: unlike CXCL12, their induction occurred in response to 4 – 6 h of hypoxic culture (Online Supplementary Figure S2). To examine the kinetics of HIF-1 and HIF-2 induction in LP-1 cells under hypoxic conditions, levels of HIF-1α and HIF-2α protein expression were measured. While HIF-1α expression was rapidly induced in response to 4–6 h of exposure to hypoxia, the induction of HIF-2α expression was delayed, requiring more than 24 h of continuous exposure to hypoxia (Figure 2C).
Over-expression and knockdown of hypoxia-inducible factors 1α and 2α in LP-1 multiple myeloma plasma cells: the effect on CXCL12
To further examine the contributions of HIF-1 and HIF-2 to the induction of MM plasma cell CXCL12 expression, LP-1 cells were engineered to stably over-express HIF-1α (LP-1-HIF-1α) or HIF-2α (LP-1-HIF-2α), and HIF over-expression confirmed by western immunoblotting (Figure 3A). Levels of CXCL12 mRNA were then measured in LP-1-HIF-1α and LP-1-HIF-2α: it was found that CXCL12 mRNA expression was higher in both cell lines than in the vector control (Figure 3B). Levels of CXCL12 protein were also measured in culture media collected from these transduced cell lines, and increased levels of CXCL12 protein were detected in LP-1-HIF-1α (685.0±17.05 pg/mL) and LP-1-HIF-2α culture media (805.1±20.3 pg/mL), compared to in the vector control culture media (410±28.8 pg/mL) (Figure 3C).
Figure 3.
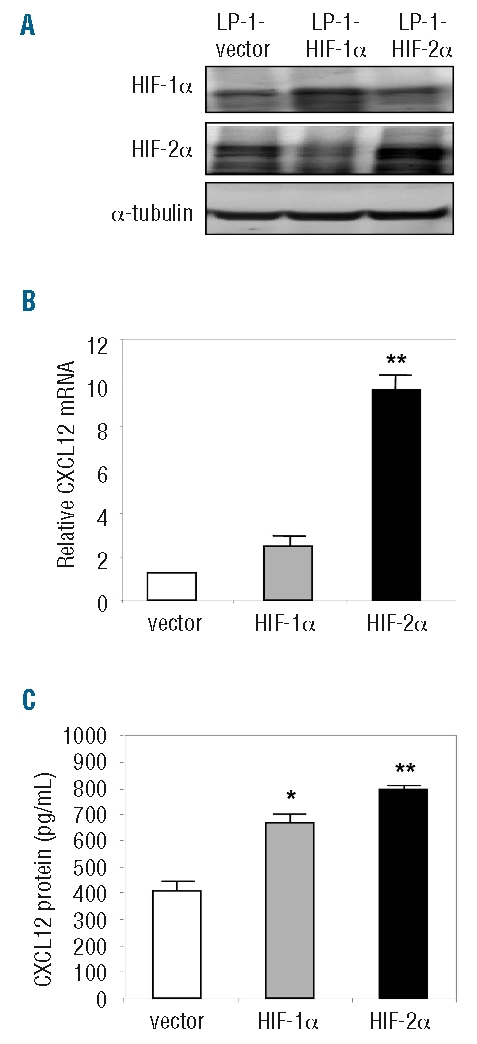
Stable over-expression of HIF-1α and HIF-2α in LP-1 cells. (A) LP-1 cells were engineered to stably over-express HIF-1α or HIF-2α and up-regulation of HIF protein confirmed by western immunoblotting. (B) Levels of CXCL12 mRNA expression were examined in the HIF over-expressing LP-1 cell lines. Columns, mean (n=3); bars, SEM. **P<0.001, compared to vector control. (C) Levels of CXCL12 protein were measured in conditioned media from the HIF over-expressing cell lines. Columns, mean (n=3); bars, SEM. *P<0.05, **P<0.001, compared to vector control.
RNA interference (RNAi) was also used to knockdown HIF-1α or HIF-2α expression in LP-1 cells, and reduced HIF expression was confirmed by western immunoblotting (Figure 4A). Using PCR, levels of CXCL12 mRNA were measured in these transduced cell lines, and a marked reduction in the hypoxic up-regulation of CXCL12 was observed in response to the HIF-1α RNAi, compared to the vector and scrambled controls (Figure 4B). Strikingly, CXCL12 expression was strongly down-regulated under both normoxic and hypoxic conditions in response to the HIF-2α RNAi. CXCL12 protein levels were also measured in culture media from each of these cell lines following normoxic or hypoxic culture (Figure 4C). Importantly, the hypoxic induction of CXCL12 protein was markedly reduced in response to both the HIF-1α RNAi (653.7±53 pg/mL) and HIF-2α RNAi (520.7±43 pg/mL), compared to the vector (1079±59 pg/mL) and scrambled (936.6±21 pg/mL) controls. Considering the degree of down-regulation of CXCL12 mRNA in response to HIF-2α knockdown, the lack of a similar response at the protein level suggests that the hypoxic induction of CXCL12 involves translational and/or post-translational regulation in these cells.
Figure 4.
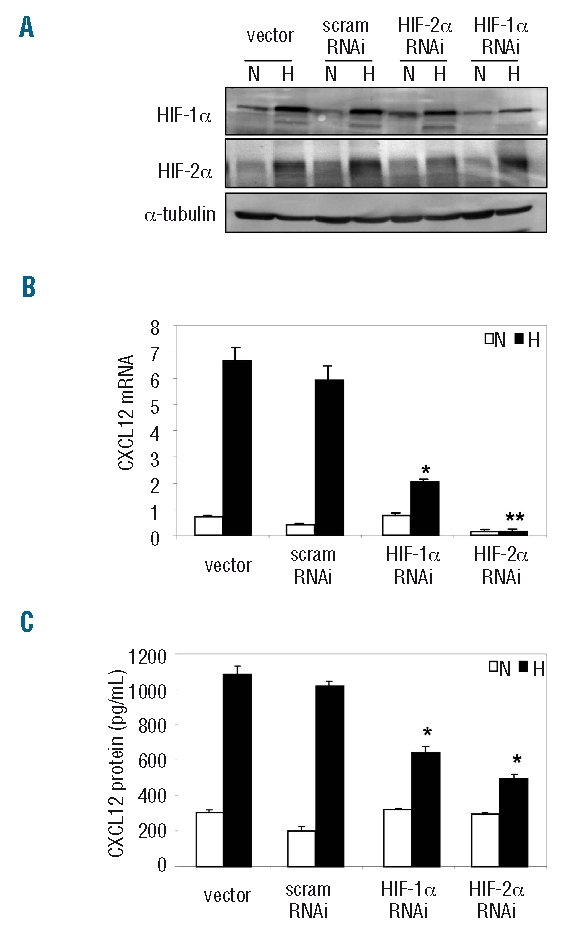
Stable knockdown of HIF-1α and HIF-2α in LP-1 cells. (A) RNA interference was used to knock down endogenous HIF-1α or HIF-2α expression in LP-1 cells, and down-regulation of HIF protein confirmed by western immunoblotting. (B) Levels of CXCL12 mRNA expression were measured in the HIF knockdowns in response to normoxic (white bars) or hypoxic (black bars) culture. Columns, mean (n=3); bars, SEM. *P<0.05, **P<0.001, compared to vector control. (C) Levels of CXCL12 protein were measured in conditioned media from the HIF knockdowns following 72 h of normoxic (white bars) or hypoxic (black bars) culture. Columns, mean (n=3); bars, SEM. *P<0.05, compared to vector control.
Hypoxia-inducible factor-2 binds to the CXCL12 promoter under hypoxic conditions
Previous studies by Ceradini et al. showed that the proximal CXCL12 promoter harbors two putative HIF binding sites (HBS1 and HBS2, situated at nucleotides −1,238 and −783 respectively, Figure 5A) and that the hypoxic induction of CXCL12 expression in human umbilical vein endothelial cells is mediated by the binding of HIF-1 to HBS1.23 While the binding of HIF-1 to the CXCL12 promoter has been demonstrated previously,23 the role of HIF-2 in the regulation of CXCL12 expression has not been examined. In this study, promoter analyses were performed to examine whether HIF-2 binds to and activates the CXCL12 promoter in MM plasma cells.
Figure 5.
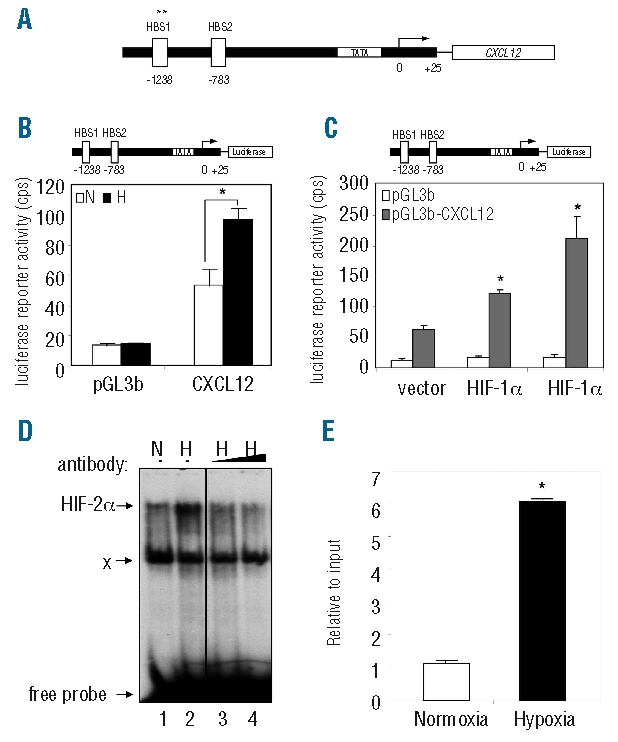
Hypoxia induces HIF-2α binding to the CXCL12 promoter in LP-1 cells. (A) Human CXCL12 locus containing two HIF binding site (HBS) sequences and transcriptional start site. Hypoxic induction of CXCL12 is primarily mediated via HBS1. (B) LP-1 cells were transiently transfected with a luciferase plasmid containing the proximal CXCL12 promoter (top, pGL3b-CXCL12). Twenty-four hours post-transfection, cells were cultured for 48 h under normoxic (white bars) or hypoxic (black bars) conditions, and luciferase assays performed. Columns, mean (n=3); bars, SEM. *P<0.05, compared to normoxia. (C) The pGL3b-CXCL12 construct (top) was transiently transfected into the HIF-over-expressing LP-1 cell lines and luciferase assays performed. Columns, mean (n=3); bars, SEM. *P<0.05, compared to vector control. (D) LP-1 cells were cultured under normoxic (N, lane 1) or hypoxic (H, lane 2) conditions for 48 h and DNA binding activity to HBS1 examined by electromobility shift assay. To determine the contribution of HIF-2 to the hypoxia-inducible complex formation, extracts were pre-incubated with HIF-2α antibody (lanes 3 and 4). (E) Chromatin immunoprecipitation was performed on LP-1 cells cultured under normoxic (white bars) or hypoxic (black bars) conditions for 48 h, and the level of HIF-2α binding to HBS1 of the CXCL12 promoter assessed by PCR. Columns, mean; bars, SEM. *P<0.05, compared to normoxia.
To measure changes in CXCL12 promoter activity in response to hypoxia, LP-1 cells were transiently transfected with a luciferase reporter plasmid containing the proximal CXCL12 promoter (pGL3b-CXCL12), and reporter assays performed (Figure 5B). These studies revealed that luciferase activity was increased (2-fold) under hypoxic conditions (96.8±7.2 cps) compared to normoxic conditions (53.4±10.8 cps). To delineate the role of HIF-1 and HIF-2α in the activation of the CXCL12 promoter, the pGL3b-CXCL12 construct was then transfected into the HIF-over-expressing LP-1 cell lines (Figure 5C), and increased luciferase activity was observed in LP-1-HIF-1α cells (122.4±6.3 cps) and LP-1-HIF-2α cells (211.6±36.5 cps), compared to the vector control (63.5 ±5.9 cps).
To examine the binding of HIF-2 to the CXCL12 promoter in LP-1 cells, electromobility shift assays were performed. These studies revealed strong binding of a hypoxia-inducible complex to the CXCL12 promoter (Figure 5D, lane 2). The presence of HIF-2 within this complex was confirmed using a HIF-2α antibody (Figure 5D, lanes 3 and 4). Chromatin immunoprecipitation assays were then performed to assess the level of HIF-2α binding to the CXCL12 promoter under normoxic and hypoxic conditions. Importantly, a 6-fold increase in the level of HIF-2α binding to the CXCL12 promoter was observed under hypoxic conditions (Figure 5E).
Hypoxia-inducible factor-induced CXCL12 stimulates angiogenesis in vivo
Hypoxia is a major physiological cue for triggering angiogenesis and initiates the transcription of angiogenic genes, such as those coding for VEGF,37,38 basic fibroblast growth factor (bFGF),39 and platelet-derived growth factor (PDGF)40 to activate the “angiogenic switch”. To examine the effect of HIF-1α and HIF-2α over-expression on MM-induced angiogenesis and the contribution of CXCL12 to this process, LP-1-HIF-1α and LP-1-HIF-2α cells were implanted subcutaneously (ventral groin region) into nude mice and half of the mice were administered T140, a potent CXCR4 antagonist41,42 (Figure 6A). In these studies, LP-1 cells stably over-expressing CXCL12 (LP-1-CXCL12) were used as a positive control. Tumor growth was monitored using bioluminescence imaging and a progressive increase in signal intensity and diameter was observed in all mice not receiving T140 (Online Supplementary Figure S3A). While progressive increases in signal intensity and diameter were also observed in mice receiving LP-1-pRUF cells + T140, mice implanted with LP-1-CXCL12, LP-1-HIF-1α and LP-1-HIF-2α and receiving T140 exhibited a marked decrease in signal intensity. The amount of vessel infiltration induced by each cell line after 2 weeks was determined by measuring implant hemoglobin content; vessel infiltration was significantly increased in implants containing LP-1-CXCL12, LP-1-HIF-1α and LP-1-HIF-2α cells, compared to vector control cells (Figure 6B). Furthermore, a significant reduction in vessel infiltration was observed in implants containing LP-1-CXCL12 cells in mice which had been systemically administered T140, compared to the vessel infiltration in mice that had not received T140 (Figure 6B). While decreased implant vascularization was also observed in T140-treated mice harboring LP-1-HIF-1α and LP-1-HIF-2α cell implants compared to that in mice that did not received T140, these differences were not statistically significant. Importantly, the observed differences in in vivo growth and angiogenesis were not attributable to differences in the rate of proliferation of the cell lines (Online Supplementary Figure S3B).
Figure 6.
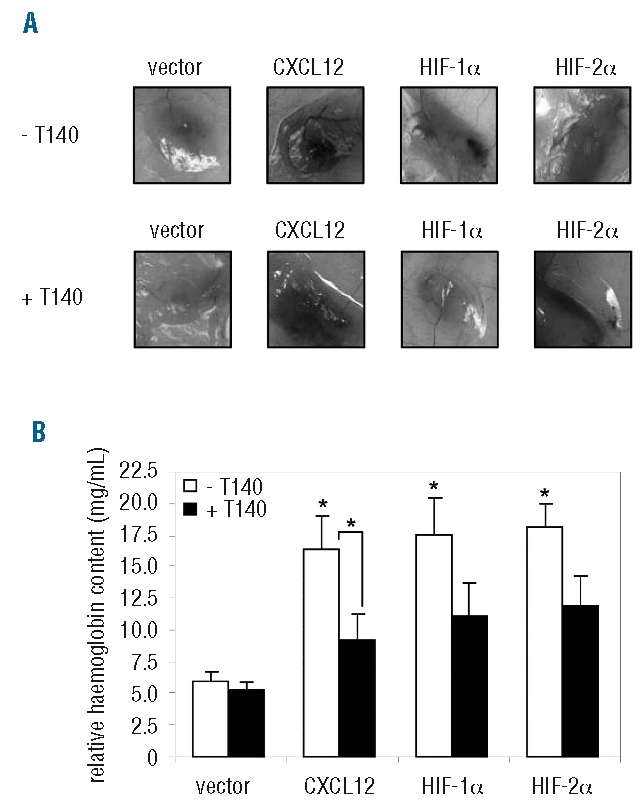
HIF-induced CXCL12 stimulates in vivo angiogenesis. (A) CXCL12-, HIF-1α- or HIF-2α-over-expressing LP-1 cells were injected subcutaneously in a Matrigel plug into mice (n=12/group), and half of the mice were administered the CXCR4 antagonist, T140. After 2 weeks, mice were euthanized and macroscopic photographs were taken of the implants. (B) The hemoglobin content of implants from untreated (white bars) and T140-treated (black bars) mice were measured and normalized to cell burden as described in the Design and Methods sections. Columns, mean (n=6/group); bars, SEM. *P<0.05, compared to vector control.
Discussion
Hypoxia is an important selective force in the evolution of tumor cells13 and aberrant HIF expression is associated with a highly aggressive disease phenotype.43,44 While hypoxia is known to be involved in the pathogenesis of solid tumors, there is a paucity of published data regarding the role of hypoxia and the HIF transcription factors in the pathogenesis of hematologic malignancies such as MM. In this study, we examined HIF-1α and HIF-2α expression in bone marrow trephine specimens from patients with MGUS or MM. While HIF-1α was found to be widely expressed throughout the bone marrow, HIF-2α expression was restricted to macrophages and CD138+ MM plasma cells. These findings suggest that the induction of aberrant HIF-2α expression is associated with the malignant transformation of MM cells, and are in keeping with previous studies showing that aberrant HIF-2α expression in tumor cells is associated with a poorer prognosis than that conferred by HIF-1α.45,46
Of note, our immunohistochemical analyses also revealed strong CXCL12 protein expression in CD138+ MM plasma cells. CXCL12 is involved in several aspects of MM pathogenesis, including transendothelial migration, bone marrow retention of plasma cells, angiogenesis and osteoclastic bone resorption.6–11 Based on previous studies showing that hypoxia regulates CXCL12 expression in other cell systems,22–26 we examined the role of hypoxia in inducing aberrant CXCL12 expression in MM plasma cells. Given the limited number of primary MM plasma cells that can be recovered from MM patients and the difficulty associated with their in vitro culture, MM plasma cell lines were used in this study. We showed, for the first time, that CXCL12 expression is up-regulated in MM plasma cells in response to prolonged hypoxia and that the HIF-2 transcription factor is a key mediator of this response. These findings contrast with those of previous studies which showed that CXCL12 is rapidly up-regulated in response to brief exposure to hypoxia (3 – 4 h) and that this is mediated by HIF-1.22–24 Given the distinct induction kinetics of HIF-1 versus HIF-2 in response to acute and chronic hypoxia, respectively, these findings likely reflect the varying requirements for CXCL12 in adapting to hypoxia - in MM plasma cells, the induction of CXCL12 expression is required for more “long-term” adaptive responses to hypoxia such as angiogenesis rather than immediate, “acute responses” such as cellular conversion to glycolytic metabolism. These data are in keeping with the notion that progressive tumor cell proliferation creates a sustained hypoxic environment and results in the induction of HIF-2α.
As for all tumor cells, MM plasma cell growth is dependent upon an adequate supply of oxygen and nutrients, and in response to increased metabolic demand, MM plasma cells induce the formation of an additional blood supply. In this study, we examined the contribution of HIF-induced CXCL12 to in vivo angiogenesis using an established subcutaneous Matrigel xenograft model of human MM.47–49 As an extension of our previous studies showing that MM-derived CXCL12 is a potent inducer of angiogenesis in vitro,7 we now show that CXCL12 is a potent pro-angiogenic agent in this in vivo model. These findings complement those of previous studies showing that tumor-derived CXCL12 stimulates angiogenesis in other human cancers.24,50,51 However, in the context of MM, our data directly contradict findings published by Menu et al.,11 who showed that the administration of T140 had no effect on bone marrow microvessel density in myelomatous 5T33MM mice. This may, in part, be explained by the fact that 5T33MM cells do not express CXCL12 and as such, the ability of 5T33MM-derived CXCL12 to stimulate angiogenesis was not under direct investigation.
The implantation of HIF over-expressing MM cells in this in vivo model stimulated a marked increase in MM-induced angiogenesis, and using the CXCR4 inhibitor, T140, we showed that CXCL12 is an important contributor to HIF-induced angiogenesis. Given that hypoxia and HIF are known to up-regulate the expression of other angiogenic growth factors such as VEGF,37,38 bFGF,39 and PDGF,40 the fact that T140 was unable to reduce HIF-induced vascularization significantly most likely reflects the important contribution of other angiogenic factors to this response.
The molecular mechanisms underlying angiogenesis in MM are complex and involve angiogenic factors secreted by both MM plasma cells themselves and microenvironmental cells such as stromal cells, osteoclasts and endothelial cells.52,53 The subcutaneous Matrigel xenograft model is a surrogate in vivo assay which is used to examine the pro-vascular properties of MM plasma cells, independently of bone marrow microenvironmental influences.47–49,54 As a result, further studies are required to examine the role of hypoxia and the HIF transcription factors on angiogenesis in the bone marrow setting and to examine the contribution of microenvironmental factors to angiogenesis in MM.
With respect to the wider implications of hypoxia on MM biology, previous studies by Asosingh et al. showed that MM plasma cell survival and expansion are supported by the hypoxic bone marrow environment.16 Using transduced HIF over-expressing MM cell lines, we now show that aberrant HIF expression stimulates a marked increase in MM-induced angiogenesis. Collectively, these findings suggest that hypoxia may play an important role in the transition from indolent MM to active, aggressive MM. This notion is supported by the immunohistochemical staining presented in this study showing that HIF-2α is aberrantly expressed by MM plasma cells in patients’ bone marrow biopsies. Further studies are required to examine the effect of hypoxia on the expression of other cytokines and growth factors known to be important in the pathogenesis of MM.
In summary, we have shown that hypoxic regulation of aberrant CXCL12 expression in MM plasma cells is mediated by the HIF-2 transcription factor. To the best of our knowledge, this is the first study to delineate the role of HIF-1 and HIF-2 in MM, and highlights the importance of assessing whether current and future anti-MM strategies modulate HIF-2 expression and/or activity. These findings also add to our understanding of the CXCL12/CXCR4 axis in MM and will, it is to be hoped, aid the development of future therapeutic strategies directed at blocking the actions of CXCL12 and/or hypoxia in MM.
Footnotes
Funding: this study was supported by grants from the National Health and Medical Research Council of Australia (ACWZ, PD, LBT, DJP, SG), the Cancer Council of South Australia (ACWZ and SG) and an Australian Postgraduate Award (SKM).
The Online version of this article has a Supplementary Appendix.
Authorship and Disclosures
SKM, PD, SAW and ACWZ contributed to the conception and design of the study, data analysis and interpretation, and manuscript writing; LBT contributed to the final approval of the manuscript; DJP, NF and ALH provided study material; SG contributed to the conception and design of the study, data analysis and interpretation, and final approval of manuscript. All authors were involved in the discussion and interpretation of data and all approved the final version.
The authors reported no potential conflicts of interest.
References
- 1.Rajkumar SV, Leong T, Roche PC, Fonseca R, Dispenzieri A, Lacy MQ, et al. Prognostic value of bone marrow angiogenesis in multiple myeloma. Clin Cancer Res. 2000;6(8):3111–6. [PubMed] [Google Scholar]
- 2.Vacca A, Ribatti D, Roncali L, Ranieri G, Serio G, Silvestris F, et al. Bone marrow angiogenesis and progression in multiple myeloma. Br J Haematol. 1994;87(3):503–8. doi: 10.1111/j.1365-2141.1994.tb08304.x. [DOI] [PubMed] [Google Scholar]
- 3.Koshiba T, Hosotani R, Miyamoto Y, Ida J, Tsuji S, Nakajima S, et al. Expression of stromal cell-derived factor 1 and CXCR4 ligand receptor system in pancreatic cancer: a possible role for tumor progression. Clin Cancer Res. 2000;6(9):3530–5. [PubMed] [Google Scholar]
- 4.Salvucci O, Yao L, Villalba S, Sajewicz A, Pittaluga S, Tosato G. Regulation of endothelial cell branching morphogenesis by endogenous chemokine stromal-derived factor-1. Blood. 2002;99(8):2703–11. doi: 10.1182/blood.v99.8.2703. [DOI] [PubMed] [Google Scholar]
- 5.Moller C, Stromberg T, Juremalm M, Nilsson K, Nilsson G. Expression and function of chemokine receptors in human multiple myeloma. Leukemia. 2003;17(1):203–10. doi: 10.1038/sj.leu.2402717. [DOI] [PubMed] [Google Scholar]
- 6.Zannettino AC, Farrugia AN, Kortesidis A, Manavis J, To LB, Martin SK, et al. Elevated serum levels of stromal-derived factor-1alpha are associated with increased osteoclast activity and osteolytic bone disease in multiple myeloma patients. Cancer Res. 2005;65(5):1700–9. doi: 10.1158/0008-5472.CAN-04-1687. [DOI] [PubMed] [Google Scholar]
- 7.Martin SK, Dewar AL, Farrugia AN, Horvath N, Gronthos S, To LB, et al. Tumor angiogenesis is associated with plasma levels of stromal-derived factor-1alpha in patients with multiple myeloma. Clin Cancer Res. 2006;12(23):6973–7. doi: 10.1158/1078-0432.CCR-06-0323. [DOI] [PubMed] [Google Scholar]
- 8.Sanz-Rodriguez F, Hidalgo A, Teixido J. Chemokine stromal cell-derived factor-1alpha modulates VLA-4 integrin-mediated multiple myeloma cell adhesion to CS-1/fibronectin and VCAM-1. Blood. 2001;97(2):346–51. doi: 10.1182/blood.v97.2.346. [DOI] [PubMed] [Google Scholar]
- 9.Parmo-Cabanas M, Bartolome RA, Wright N, Hidalgo A, Drager AM, Teixido J. Integrin alpha4beta1 involvement in stromal cell-derived factor-1alpha-promoted myeloma cell transendothelial migration and adhesion: role of cAMP and the actin cytoskeleton in adhesion. Exp Cell Res. 2004;294(2):571–80. doi: 10.1016/j.yexcr.2003.12.003. [DOI] [PubMed] [Google Scholar]
- 10.Gazitt Y, Akay C. Mobilization of myeloma cells involves SDF-1/CXCR4 signaling and downregulation of VLA-4. Stem Cells. 2004;22(1):65–73. doi: 10.1634/stemcells.22-1-65. [DOI] [PubMed] [Google Scholar]
- 11.Menu E, Asosingh K, Indraccolo S, De Raeve H, Van Riet I, Van Valckenborgh E, et al. The involvement of stromal derived factor 1α in homing and progression of multiple myeloma in the 5TMM model. Haematologica. 2006;91(5):605–12. [PubMed] [Google Scholar]
- 12.Pennathur-Das R, Levitt L. Augmentation of in vitro human marrow erythropoiesis under physiological oxygen tensions is mediated by monocytes and T lymphocytes. Blood. 1987;69(3):899–907. [PubMed] [Google Scholar]
- 13.Graeber TG, Osmanian C, Jacks T, Housman DE, Koch CJ, Lowe SW, et al. Hypoxia-mediated selection of cells with diminished apoptotic potential in solid tumours. Nature. 1996;379(6560):88–91. doi: 10.1038/379088a0. [DOI] [PubMed] [Google Scholar]
- 14.Zhong H, De Marzo AM, Laughner E, Lim M, Hilton DA, Zagzag D, et al. Overexpression of hypoxia-inducible factor 1α in common human cancers and their metastases. Cancer Res. 1999;59(22):5830–5. [PubMed] [Google Scholar]
- 15.Talks KL, Turley H, Gatter KC, Maxwell PH, Pugh CW, Ratcliffe PJ, et al. The expression and distribution of the hypoxia-inducible factors HIF-1alpha and HIF-2alpha in normal human tissues, cancers, and tumor-associated macrophages. Am J Pathol. 2000;157(2):411–21. doi: 10.1016/s0002-9440(10)64554-3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Asosingh K, De Raeve H, de Ridder M, Storme GA, Willems A, Van Riet I, et al. Role of the hypoxic bone marrow microenvironment in 5T2MM murine myeloma tumor progression. Haematologica. 2005;90(6):810–7. [PubMed] [Google Scholar]
- 17.Lu L, Payvandi F, Wu L, Zhang LH, Hariri RJ, Man HW, et al. The anti-cancer drug lenalidomide inhibits angiogenesis and metastasis via multiple inhibitory effects on endothelial cell function in normoxic and hypoxic conditions. Microvasc Res. 2009;77(2):78–86. doi: 10.1016/j.mvr.2008.08.003. [DOI] [PubMed] [Google Scholar]
- 18.Shin DH, Chun YS, Lee DS, Huang LE, Park JW. Bortezomib inhibits tumor adaptation to hypoxia by stimulating the FIH-mediated repression of hypoxia-inducible factor-1. Blood. 2008;111(6):3131–6. doi: 10.1182/blood-2007-11-120576. [DOI] [PubMed] [Google Scholar]
- 19.Salceda SaC J. Hypoxia-inducible factor 1α (HIF-1α) protein is rapidly degraded by the ubiquitin-proteasome system under normoxic conditions. Its stabilization by hypoxia depends on redox-induced changes. J Biol Chem. 1997;272(36):22642–7. doi: 10.1074/jbc.272.36.22642. [DOI] [PubMed] [Google Scholar]
- 20.Hu CJ, Wang LY, Chodosh LA, Keith B, Simon MC. Differential roles of hypoxia-inducible factor 1α (HIF-1α) and HIF-2α in hypoxic gene regulation. Mol Cell Biol. 2003;23(24):9361–74. doi: 10.1128/MCB.23.24.9361-9374.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Sowter HM, Raval RR, Moore JW, Ratcliffe PJ, Harris AL. Predominant role of hypoxia-inducible transcription factor (Hif)-1α versus Hif-2α in regulation of the transcriptional response to hypoxia. Cancer Res. 2003;63(19):6130–4. [PubMed] [Google Scholar]
- 22.Hitchon C, Wong K, Ma G, Reed J, Lyttle D, El-Gabalawy H. Hypoxia-induced production of stromal cell-derived factor 1 (CXCL12) and vascular endothelial growth factor by synovial fibroblasts. Arthritis Rheum. 2002;46(10):2587–97. doi: 10.1002/art.10520. [DOI] [PubMed] [Google Scholar]
- 23.Ceradini DJ, Kulkarni AR, Callaghan MJ, Tepper OM, Bastidas N, Kleinman ME, et al. Progenitor cell trafficking is regulated by hypoxic gradients through HIF-1 induction of SDF-1. Nat Med. 2004;10(8):858–64. doi: 10.1038/nm1075. [DOI] [PubMed] [Google Scholar]
- 24.Kryczek I, Lange A, Mottram P, Alvarez X, Cheng P, Hogan M, et al. CXCL12 and vascular endothelial growth factor synergistically induce neoangiogenesis in human ovarian cancers. Cancer Res. 2005;65(2):465–72. [PubMed] [Google Scholar]
- 25.Zagzag D, Krishnamachary B, Yee H, Okuyama H, Chiriboga L, Ali MA, et al. Stromal cell-derived factor-1α and CXCR4 expression in hemangioblastoma and clear cell-renal cell carcinoma: von Hippel-Lindau loss-of-function induces expression of a ligand and its receptor. Cancer Res. 2005;65(14):6178–88. doi: 10.1158/0008-5472.CAN-04-4406. [DOI] [PubMed] [Google Scholar]
- 26.Tabatabai G, Frank B, Mohle R, Weller M, Wick W. Irradiation and hypoxia promote homing of haematopoietic progenitor cells towards gliomas by TGF-beta-dependent HIF-1alpha-mediated induction of CXCL12. Brain. 2006;129(Pt 9):2426–35. doi: 10.1093/brain/awl173. [DOI] [PubMed] [Google Scholar]
- 27.Diamond P, Labrinidis A, Martin SK, Farrugia AN, Gronthos S, To LB, et al. Targeted disruption of the CXCL12/CXCR4 axis inhibits osteolysis in a murine model of myeloma-associated bone loss. J Bone Miner Res. 2009;24(7):1150–61. doi: 10.1359/jbmr.090210. [DOI] [PubMed] [Google Scholar]
- 28.Wang GL, Jiang BH, Rue EA, Semenza GL. Hypoxia-inducible factor 1 is a basic-helix-loop-helix-PAS heterodimer regulated by cellular O2 tension. Proc Natl Acad Sci USA. 1995;92(12):5510–4. doi: 10.1073/pnas.92.12.5510. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Lando D, Peet DJ, Whelan DA, Gorman JJ, Whitelaw ML. Asparagine hydroxylation of the HIF transactivation domain a hypoxic switch. Science. 2002;295(5556):858–61. doi: 10.1126/science.1068592. [DOI] [PubMed] [Google Scholar]
- 30.Solban N, Selbo PK, Sinha AK, Chang SK, Hasan T. Mechanistic investigation and implications of photodynamic therapy induction of vascular endothelial growth factor in prostate cancer. Cancer Res. 2006;66(11):5633–40. doi: 10.1158/0008-5472.CAN-06-0604. [DOI] [PubMed] [Google Scholar]
- 31.Zimmer M, Doucette D, Siddiqui N, Iliopoulos O. Inhibition of hypoxia-inducible factor is sufficient for growth suppression of VHL−/− tumors. Mol Cancer Res. 2004;2(2):89–95. [PubMed] [Google Scholar]
- 32.Ponomarev V, Doubrovin M, Serganova I, Vider J, Shavrin A, Beresten T, et al. A novel triple-modality reporter gene for whole-body fluorescent, bioluminescent, and nuclear noninvasive imaging. Eur J Nucl Med Mol Imaging. 2004;31(5):740–51. doi: 10.1007/s00259-003-1441-5. [DOI] [PubMed] [Google Scholar]
- 33.Livak KJ, Schmittgen TD. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) method. Methods. 2001;25(4):402–8. doi: 10.1006/meth.2001.1262. [DOI] [PubMed] [Google Scholar]
- 34.Osborne CS, Vadas MA, Cockerill PN. Transcriptional regulation of mouse granulocyte-macrophage colony-stimulating factor/IL-3 locus. J Immunol. 1995;155(1):226–35. [PubMed] [Google Scholar]
- 35.Maniatis T, Fritsch EF, Sambrook J. 1982. Molecular Cloning: A Laboratory Manual. [Google Scholar]
- 36.Chavez JC, Baranova O, Lin J, Pichiule P. The transcriptional activator hypoxia inducible factor 2 (HIF-2/EPAS-1) regulates the oxygen-dependent expression of erythropoietin in cortical astrocytes. J Neurosci. 2006;26(37):9471–81. doi: 10.1523/JNEUROSCI.2838-06.2006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Levy AP, Levy NS, Wegner S, Goldberg MA. Transcriptional regulation of the rat vascular endothelial growth factor gene by hypoxia. J Biol Chem. 1995;270(22):13333–40. doi: 10.1074/jbc.270.22.13333. [DOI] [PubMed] [Google Scholar]
- 38.Liu Y, Cox SR, Morita T, Kourembanas S. Hypoxia regulates vascular endothelial growth factor gene expression in endothelial cells. Identification of a 5′ enhancer. Circ Res. 1995;77(3):638–43. doi: 10.1161/01.res.77.3.638. [DOI] [PubMed] [Google Scholar]
- 39.Kuwabara K, Ogawa S, Matsumoto M, Koga S, Clauss M, Pinsky DJ, et al. Hypoxia-mediated induction of acidic/basic fibroblast growth factor and platelet-derived growth factor in mononuclear phagocytes stimulates growth of hypoxic endothelial cells. Proc Natl Acad Sci USA. 1995;92(10):4606–10. doi: 10.1073/pnas.92.10.4606. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Kourembanas S, Hannan RL, Faller DV. Oxygen tension regulates the expression of the platelet-derived growth factor-B chain gene in human endothelial cells. J Clin Invest. 1990;86(2):670–4. doi: 10.1172/JCI114759. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Tamamura H, Xu Y, Hattori T, Zhang X, Arakaki R, Kanbara K, et al. A low-molecular-weight inhibitor against the chemokine receptor CXCR4: a strong anti-HIV peptide T140. Biochem Biophys Res Commun. 1998;253(3):877–82. doi: 10.1006/bbrc.1998.9871. [DOI] [PubMed] [Google Scholar]
- 42.Takenaga M, Tamamura H, Hiramatsu K, Nakamura N, Yamaguchi Y, Kitagawa A, et al. A single treatment with microcapsules containing a CXCR4 antagonist suppresses pulmonary metastasis of murine melanoma. Biochem Biophys Res Commun. 2004;320(1):226–32. doi: 10.1016/j.bbrc.2004.05.155. [DOI] [PubMed] [Google Scholar]
- 43.Bos R, van der Groep P, Greijer AE, Shvarts A, Meijer S, Pinedo HM, et al. Levels of hypoxia-inducible factor-1alpha independently predict prognosis in patients with lymph node negative breast carcinoma. Cancer. 2003;97(6):1573–81. doi: 10.1002/cncr.11246. [DOI] [PubMed] [Google Scholar]
- 44.Shibaji T, Nagao M, Ikeda N, Kanehiro H, Hisanaga M, Ko S, et al. Prognostic significance of HIF-1 alpha overexpression in human pancreatic cancer. Anticancer Res. 2003;23(6C):4721–7. [PubMed] [Google Scholar]
- 45.Lofstedt T, Fredlund E, Holmquist-Mengelbier L, Pietras A, Ovenberger M, Poellinger L, et al. Hypoxia inducible factor-2alpha in cancer. Cell Cycle. 2007;6(8):919–26. doi: 10.4161/cc.6.8.4133. [DOI] [PubMed] [Google Scholar]
- 46.Patel SA, Simon MC. Biology of hypoxia-inducible factor-2alpha in development and disease. Cell Death Differ. 2008;15(4):628–34. doi: 10.1038/cdd.2008.17. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Frost P, Moatamed F, Hoang B, Shi Y, Gera J, Yan H, et al. In vivo antitumor effects of the mTOR inhibitor CCI-779 against human multiple myeloma cells in a xenograft model. Blood. 2004;104(13):4181–7. doi: 10.1182/blood-2004-03-1153. [DOI] [PubMed] [Google Scholar]
- 48.Podar K, Tonon G, Sattler M, Tai YT, Legouill S, Yasui H, et al. The small-molecule VEGF receptor inhibitor pazopanib (GW786034B) targets both tumor and endothelial cells in multiple myeloma. Proc Natl Acad Sci USA. 2006;103(51):19478–83. doi: 10.1073/pnas.0609329103. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Podar K, Raab MS, Zhang J, McMillin D, Breitkreutz I, Tai YT, et al. Targeting PKC in multiple myeloma: in vitro and in vivo effects of the novel, orally available small-molecule inhibitor enzastaurin (LY317615.HCl) Blood. 2007;109(4):1669–77. doi: 10.1182/blood-2006-08-042747. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Orimo A, Gupta PB, Sgroi DC, Arenzana-Seisdedos F, Delaunay T, Naeem R, et al. Stromal fibroblasts present in invasive human breast carcinomas promote tumor growth and angiogenesis through elevated SDF-1/CXCL12 secretion. Cell. 2005;121(3):335–48. doi: 10.1016/j.cell.2005.02.034. [DOI] [PubMed] [Google Scholar]
- 51.Chu CY, Cha ST, Lin WC, Lu PH, Tan CT, Chang CC, et al. Stromal-cell-derived factor-1{α} (SDF-1{α}/CXCL12)-enhanced angiogenesis of human basal cell carcinoma cells involves ERK1/2-NF-κ B/interleukin-6 pathway. Carcinogenesis. 2009;30(2):205–13. doi: 10.1093/carcin/bgn228. [DOI] [PubMed] [Google Scholar]
- 52.Dankbar B, Padro T, Leo R, Feldmann B, Kropff M, Mesters RM, et al. Vascular endothelial growth factor and interleukin-6 in paracrine tumor-stromal cell interactions in multiple myeloma. Blood. 2000;95(8):2630–6. [PubMed] [Google Scholar]
- 53.Vacca A, Ribatti D. Bone marrow angiogenesis in multiple myeloma. Leukemia. 2006;20(2):193–9. doi: 10.1038/sj.leu.2404067. [DOI] [PubMed] [Google Scholar]
- 54.Zhang J, Sattler M, Tonon G, Grabher C, Lababidi S, Zimmerhackl A, et al. Targeting angiogenesis via a c-Myc/hypoxia-inducible factor-1alpha-dependent pathway in multiple myeloma. Cancer Res. 2009;69(12):5082–90. doi: 10.1158/0008-5472.CAN-08-4603. [DOI] [PubMed] [Google Scholar]