

Abstract
Background and aim
Chronic idiopathic intestinal pseudo-obstruction (CIIP) is characterised by severe impairment of intestinal propulsive motility that mimics bowel obstruction. JC virus (JCV) is a polyomavirus that can infect brain glial cells causing a fatal disease, but may also be found throughout the normal gastrointestinal tract. The hypothesis that JCV infects the myenteric plexuses of patients with CIIP was tested.
Methods
10 patients with CIIP and 61 normal specimens (30 ascending colon and 31 ileum) from patients with uncomplicated colon cancer were studied. DNA was extracted from the myenteric plexuses, and JCV T antigen (TAg) DNA and the viral regulatory region were detected by PCR and sequencing. Immunohistochemistry was performed to detect JCV viral protein expression, neuronal and glial markers. Fluorescence in situ hybridisation was performed for cellular localisation of the JCV infection.
Results
Clinical studies demonstrated neurogenic impairment, and pathological analyses showed neuropathy in each patient with CIIP. JCV TAg DNA was found in the myenteric plexuses of 8/10 (80%) of the patients with CIIP and 3/31 (9.7%) of the control patients (p<0.001). All samples were JCV Mad-1 strains. Seven of the 10 CIIP specimens expressed both JCV TAg and the JCV viral protein VP1, while none of the controls expressed either. JCV infection co-localised with glial fibrillary acidic protein expression, a marker of enteric glial cells.
Conclusion
JCV infection occurs in the myenteric plexuses of patients with CIIP. The JCV localisation in enteroglial cells suggests a possible pathological role for this virus in enteric neuropathy.
Chronic idiopathic intestinal pseudo-obstruction (CIIP) is a rare, progressive and life-threatening syndrome characterised by severely impaired gastrointestinal motility. The clinical presentation of CIIP is dominated by signs and symptoms of intestinal obstruction, including recurrent episodes of abdominal pain and distension, accompanied by bloating, nausea, vomiting, etc., without evidence of mechanical obstruction.1 2 CIIP may occur throughout the gastrointestinal tract, but usually involves the small bowel. The accompanying malabsorption and malnutrition often lead to severe weight loss, requiring parenteral nutritional support. Various infectious, neurological, paraneoplastic and metabolic conditions have been reported to affect the neuromuscular layer of the gut and cause this syndrome; these have been called secondary CIIP. The primary idiopathic form of CIIP has no known aetiology, and the pathogenesis is poorly understood.1 3 4 Familial cases of CIIP with autosomal dominant or recessive modes of inheritance have been described, but the majority of CIIP remains sporadic.5
CIIP can be placed into three major histopathological categories: neuropathies (either with purely degenerative or inflammatory changes to the intrinsic enervation of the gut); mesenchymopathies (abnormalities of the interstitial cells of Cajal (ICCs)); or pure myopathies. However, overlap among these groups is common.1 5 6 Several neurotropic viruses have the ability to infect the central nervous system (CNS), and could possibly infect the enteric nervous system (ENS). An active viral infection affecting enteric neurons would be a plausible explanation for the pathogenesis of CIIP. In fact, several prior reports have proposed that certain DNA viruses may be involved in chronic gastrointestinal motility disorders, including CIIP and achalasia.6–12
JC virus (JCV) is a member of the polyomavirus family, infects humans worldwide, and 90% of the population carry antibodies to the virus by adult life.13 The initial infection is asymptomatic, but it may become persistent. JCV DNA is frequently present in the upper and lower gastrointestinal tract of healthy adults.14 15 The virus encodes a T-antigen (TAg), which is a potent transforming protein both in vivo and in vitro.16–18 In patients with impaired cell-mediated immunity such as AIDS, this neurotropic virus may reactivate, and can cause a lytic infection of brain glial cells, leading to the lethal demyelinating disease, progressive multifocal leucoencephalopathy (PML).19
Because of the known neuropathic capability of this virus, and its frequent presence in the gut, we proposed that JCV might be detectable in tissues of patients with CIIP, and possibly be involved in the pathogenesis of this disease. In this report, we have demonstrated that the virus actively infects the enteroglial cells of the myenteric plexuses of most of the patients with CIIP we studied.
Methods
Inclusion criteria and patients
The study was performed on 10 patients (8/10 females; mean age (SD) at entry in the study 33.2 (8.11) years; range 21–47 years) with the diagnosis of neurogenic CIIP, seen in the Laboratory of Functional Gastrointestinal Disorders at the S. Orsola-Malpighi Hospital of the University of Bologna, between 2004 and 2006. All patients had a long history of severe gastrointestinal symptoms and their diagnosis of CIIP was already established years before based on fulfilment of previous criteria.4 20 21 All patients with CIIP included in this study had undergone multiple surgeries (table 1). Consequently, full-thickness samples of the small and/or large bowel were available. None of the patients was subjected to treatment with immunosuppressors and, among the patients undergoing intestinal transplantation, samples were obtained from the resected specimens. A series of 31 patients who had undergone colonic resection for sporadic colorectal cancer were collected from the pathology archives at Baylor University Medical Center in Dallas, and the Pathology Department of the University of Bologna; macroscopically and histologically normal tissues were selected for study. Institutional Review Board approval was granted for this study from Baylor University Medical Center and from the University of Bologna.
Table 1.
Clinical findings in the patients with chronic idiopathic intestinal pseudo-obstruction
| Patentno. | Sex | Age | Number of obstructive episodes/year | Severe digestive symptoms | BMI | Surgery | Feeding | Outcome |
|---|---|---|---|---|---|---|---|---|
| 1 | Female | 21 | 0.40 | N, V, F, AD, P, Dia | 19.26 | Fundoplication, cholecystectomy, pyloroplasty, lysis of adhesions, SB transplantation | EN | Alive |
| 2 | Female | 26 | 3 | N, F, AD, P, Dia | 14.34 | Rectopexy, left hemicolectomy, SB transplantation | Semi-liquid/formula | Death |
| 3 | Female | 25 | 72 | N, F, AD, P, C | 20.07 | Appendectomy, SB transplantation | HPN | Alive |
| 4 | Female | 30 | 1.5 | AD, Dia | 21.30 | Appendectomy, lysis of adhesions, total colectomy | Oral feeding | Alive |
| 5 | Female | 41 | 0.67 | AD, Dia | 21.87 | Appendectomy, cholecystectomy, ileocolonic resection and ileocolonic anastomosis (twice) | Oral feeding | Alive |
| 6 | Female | 38 | 5 | N, V, F, AD, P, C | 18.42 | Appendectomy, left hemicolectomy, lysis of adhesions, hysterectomy | HPN | Alive |
| 7 | Female | 47 | 36 | AD, P, D | 15.78 | Colostomy, ileostomy | HPN | Death |
| 8 | Female | 30 | 3 | N, V, AD, P, C | 19.81 | Appendectomy, left hemicolectomy, lysis of adhesions | Oral feeding | Alive |
| 9 | Male | 37 | 12 | N, V, AD, P, C | 21.78 | SB transplantation | HPN | Alive |
| 10 | Male | 37 | 10 | N, V, AD, P, C | 22.65 | SB transplantation | HPN | Alive |
AD, abdominal distension; BMI, body mass index; C, constipation; Dia, diarrhoea; EN, enteral nutrition; F, fullness; HPN, home parenteral nutrition; N, nausea; P, abdominal pain; SB, small bowel; V, vomiting.
Histopathology and immunohistochemistry (IHC)
Full-thickness biopsies were obtained from all of the patients with CIIP, including the ileum from 7 patients (5 females and 2 males, age range 21–47 years) or the descending colon in 4 patients (4 females, age range: 30–41 years). One patient provided both small bowel and colonic samples. Full-thickness specimens were obtained from a site distant from the colon cancer in the 31 controls; these came from the caecum or ascending colon in 30 subjects, and from the terminal ileum in 31.
To analyse the ENS, IHC was performed for expression of neuron-specific enolase (NSE), enteroglial protein S100-β, vasoactive intestinal polypeptide (VIP), substance P/related tachykinins (SP), neuropeptide Y (NPY), calcitonin gene-related peptide (CGRP) and neuronal nitric oxide synthase (nNOS) (Chemicon International, Temecula, California, USA). Antibodies against c-Kit and α-actin (DakoCytomation, Milan, Italy) were used to identify the ICCs and smooth muscle cells, respectively, while an antibody directed against the glial fibrillary acidic protein (GFAP) was used to evaluate the possible localisation of JCV in enteric glial cells by dual-labelling immunofluorescence. Control samples were used to establish the normal distribution according to immunoreactive density and intensity of the several markers used. The density and intensity of immunoreactive patterns were compared by two independent investigators (RDeG and LR), according to staining intensity and density of nerve distribution. The sections were scored using a 4-point ordinate scale, as follows: +++, 70–100% of labelled perikarya and nerve fibres with intense immunoreaction; ++−, 30 to <70% of labelled perikarya and nerve fibres with intense/moderate immunoreaction; +−−, <30% of labelled nerve fibres with moderate/weak immunoreaction; and −−−, no detectable immunoreaction.20–22
Antibodies to detect JCV proteins included a mouse monoclonal antibody against the SV40 large T antigen, which cross-reacts with JCV TAg (clone PAb416, 1:100 dilution; Oncogene Research Products, San Diego, California, USA), a mouse monoclonal antibody against JCV TAg (clone PAb2003, 1:100 dilution) and a rabbit polyclonal antibody against JCV viral protein 1 (VP1) (1:1000 and 1:5000 dilutions for IHC and for immunofluorescence, respectively). IHC was performed with the Dako En Vision-labelled polymer (Dako Cytomation). Appropriate positive controls were used in each experiment, including hamster brain expressing JCV TAg, and cells expressing the JCV VP1.
Dual immunofluorescence for VP1 and GFAP proteins was performed using the Alexa Fluor 488 goat anti-rabbit, and the Alexa Fluor 594 goat anti-mouse second antibodies (Invitrogen, Carlsbad, California, USA). Slides were counterstained with 4′,6-diamidino-2-phenylindole (DAPI), observed with an AxioSkop2 multichannel epi-fluorescence microscope and processed by AxioVision software (Carl Zeiss, Thornwood, New York, USA).
Small bowel manometry
Small bowel manometry was performed in seven of the patients with CIIP (all females, age range: 21–47 years) according to a previously described technique.21 23–25 Manometric recordings were analysed by two investigators (VS and RFC) who independently recorded the type and frequency of small bowel abnormal motor patterns, as previously defined.21 25 Briefly, the following abnormalities were considered: (1) abnormal propagation and/or configuration of activity fronts; (2) bursts or periods with continuous high-amplitude and high-frequency phasic pressure activity that were not propagated and not followed by motor quiescence; (3) sustained contractions that occurred in a segment of intestine while normal or reduced contractility was simultaneously recorded at other levels; (4) inability of an adequate meal to abolish migrating motor complexes (MMCs) for at least 180 min; (5) hypomotility or absence of any detectable intestinal contractions; and (6) clustered contractions or 3–10 regular contractions preceded and followed by ≥1 min of absent motor activity lasting for ≥20 min. Pathological tracings required the presence of at least two bursts in 6 h and/or at least an abnormal activity front.
DNA extraction and detection of viral genomic sequences
To isolate the DNA from the myenteric plexuses, we stained the samples of each CIIP and control patient for NSE, followed by brief counterstaining with haematoxylin (fig 2A), and tissue microdissection was performed using a scalpel and a dissecting microscope (fig 2B). DNA isolation and PCR for JCV TAg were performed in a dedicated area isolated from prior PCR products, as previously described.26 PCR products were sequenced with an ABI PRISM 3100 Avant Genetic Analyzer (Applied Biosystems, Foster City, California, USA). The sequences were aligned using GenBank.
Figure 2.
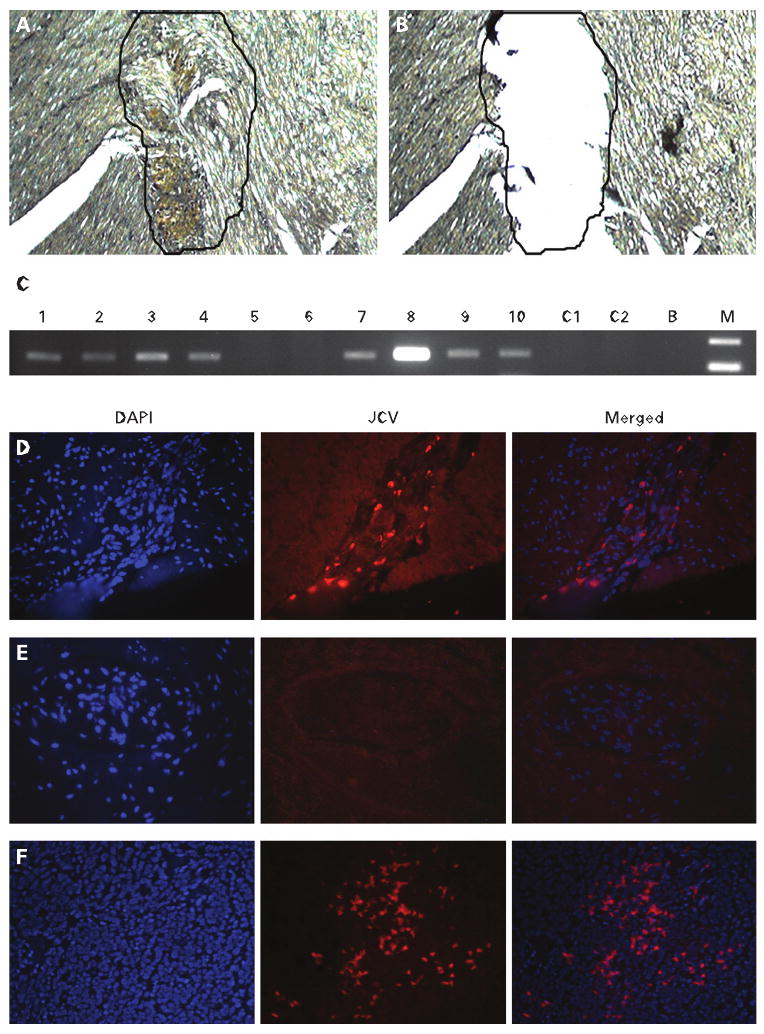
Upper panel (A–C): microdissection of the myenteric plexus. (A) A myenteric plexus is stained with neuron-specific enolase-specific antibody before DNA extraction. (B) Microdissection of the selected area was performed before DNA extraction. (C) Agarose gel electrophoresis of JC virus (JCV)-positive and -negative samples. Samples are numbered from 1 to 10; C1 and C2 are two samples extracted from control patients. B, water lane; M, the molecular size standard. Lower panel (D–F): fluoresce in situ hybridisation (FISH) analysis for JCV DNA. The left column shows nuclear (4′,6-diamidino-2-phenylindole (DAPI)) staining, the middle column shows the same tissue hybridised with FISH markers for JCV DNA, and the right column shows the merged images. (D) The myenteric plexus from a patient with chronic idiopathic intestinal pseudo-obstruction (CIIP) with positive PCR for JCV DNA shows multiple areas of hybridisation. (E) The myenteric plexus from a control sample does not show hybridisation. (F) A section of hamster brain previously transfected with JCV T antigen was used as positive control. Multiple areas of hybridisation were observed.
Samples were also subjected to PCR amplification for the following viruses: BK virus (BKV), simian virus 40 (SV40), cytomegalovirus (CMV), herpes simplex virus (HSV), varicella zoster virus (VZV) and Epstein–Barr virus (EBV) (Supplementary methods).
Fluorescence in situ hybridisation (FISH)
The JCV-Mad1 genome inserted into pBR322 (pM1TC) was used to generate the probes using the BioPrime DNA Labeling System (Invitrogen). Two probes were synthesised from the early and late regions of transcription. Pre-hybridisation and hybridisation were performed with the DNA Detector Kit (KPL, Gaithersburg, Maryland, USA). Hybridised probes were bound to streptavidin-conjugated Cy3, and slides were counterstained with DAPI. Slides were visualised under an AxioSkop2 multichannel epifluorescence microscope and processed by AxioVision software (Carl Zeiss Inc., Thornwood, New York, USA).
Statistical analysis
We used Fisher's exact test to compare the CIIP and control groups with respect to JCV DNA amplification. Two-tailed p values were considered significant when <0.05.
Results
Clinical, manometric and histopathological features of the patients with CIIP
Clinical features of the patients investigated in this study are summarised in table 1. All seven patients who underwent small bowel manometry showed unequivocal motor abnormalities (table 2). As previously reported,25 the dysmotility patterns observed were suggestive of an underlying derangement of intrinsic neural control mechanisms and involved bursts of uncoordinated, non-propagated, frequent contractions in five patients and interdigestive activity fronts which were abnormal for propagation and/or configuration in five patients. In three patients, the manometric analyses showed frequent motor abnormalities, including a generalised paucity of contractions during both fasting and postprandial periods (1 patient) and periods of regular clustered contractions (2 patients).
Table 2.
Manometric results in seven patients with chronic idiopathic intestinal pseudo-obstruction
| Patient no | No. of abnormal activity fronts/h | No. of bursts/h | Inadequate response to meal | Hypomotility | Clustered contractions |
|---|---|---|---|---|---|
| 1 | 0.12 | 0 | Absent | Absent | Absent |
| 2 | 0.36 | 0.14 | Absent | Absent | Absent |
| 3 | 0.25 | 0.11 | Absent | Absent | Absent |
| 4 | 0 | 0.33 | Absent | Absent | Present |
| 6 | 0 | 0.45 | Absent | Absent | Present |
| 7 | 0.08 | 0 | Absent | Present | Absent |
| 8 | 0.22 | 1.64 | Absent | Absent | Absent |
IHC analysis confirmed the presence of an underlying enteric (visceral) neuropathy in all of the patients with CIIP. No evident neuropathic abnormalities were observed in any of the control specimens. Neuronal abnormalities identified at IHC included changes in the structural markers of enteric neurons and glia—that is, reduced expression of NSE and S100-β protein, respectively, associated with swelling, vacuolisation and loss of submucosal and myenteric neurons (table 3). Importantly, we found marked reductions of CGRP, NPY, VIP, nNOS and SP immunoreactivity in the submucosal and myenteric neuronal cell bodies and nerve fibres (supplying the circular muscle layer) compared with controls (fig 1). The ICC network revealed by c-Kit immunolabelling was found to be markedly decreased in 8 of the 10 cases of CIIP compared with controls.
Table 3.
Pathological and immunohistochemical findings in patients with chronic idiopathic intestinal pseudo-obstruction, and in the three controls found positive for JC virus DNA sequences
| Patient no | NSE | S100-β | Vacuolisation and cell loss | CGRP | NPY | VIP | nNOS | SP | c-Kit |
|---|---|---|---|---|---|---|---|---|---|
| 1 | Decreased | Decreased | Increased | Decreased | Decreased | Decreased | Decreased | Decreased | Decreased |
| 2 | Decreased | Decreased | Increased | Decreased | Decreased | Decreased | Decreased | Decreased | Decreased |
| 3 | Decreased | Decreased | Increased | Decreased | Decreased | Decreased | Decreased | Decreased | Decreased |
| 4 | Decreased | Decreased | Increased | Decreased | Decreased | Decreased | Decreased | Decreased | Decreased |
| 5 | Decreased | Decreased | Increased | Decreased | Decreased | Decreased | Decreased | Decreased | Decreased |
| 6 | Decreased | Decreased | Increased | Decreased | Decreased | Decreased | Decreased | Decreased | Decreased |
| 7 | Decreased | Decreased | Increased | Decreased | Decreased | Decreased | Decreased | Decreased | Decreased |
| 8 | Decreased | Decreased | Increased | Decreased | Decreased | Decreased | Decreased | Decreased | Decreased |
| 9 | Decreased | Decreased | Increased | Decreased | Decreased | Decreased | Decreased | Decreased | No changes |
| 10 | Decreased | Decreased | Increased | Decreased | Decreased | Decreased | Decreased | Decreased | No changes |
| Control 1 | No changes | No changes | No changes | No changes | No changes | No changes | No changes | No changes | No changes |
| Control 2 | No changes | No changes | No changes | No changes | No changes | No changes | No changes | No changes | No changes |
| Control 3 | No changes | No changes | No changes | No changes | No changes | No changes | No changes | No changes | No changes |
CGRP, calcitonin gene-related peptide; nNOS, neuronal nitric oxide synthase; NPY, neuropeptide Y; NSE, neuron-specific enolase; S100-β, enteroglial protein S100-β; SP, substance P/related tachykinins; VIP, vasoactive intestinal polypeptide.
Figure 1.
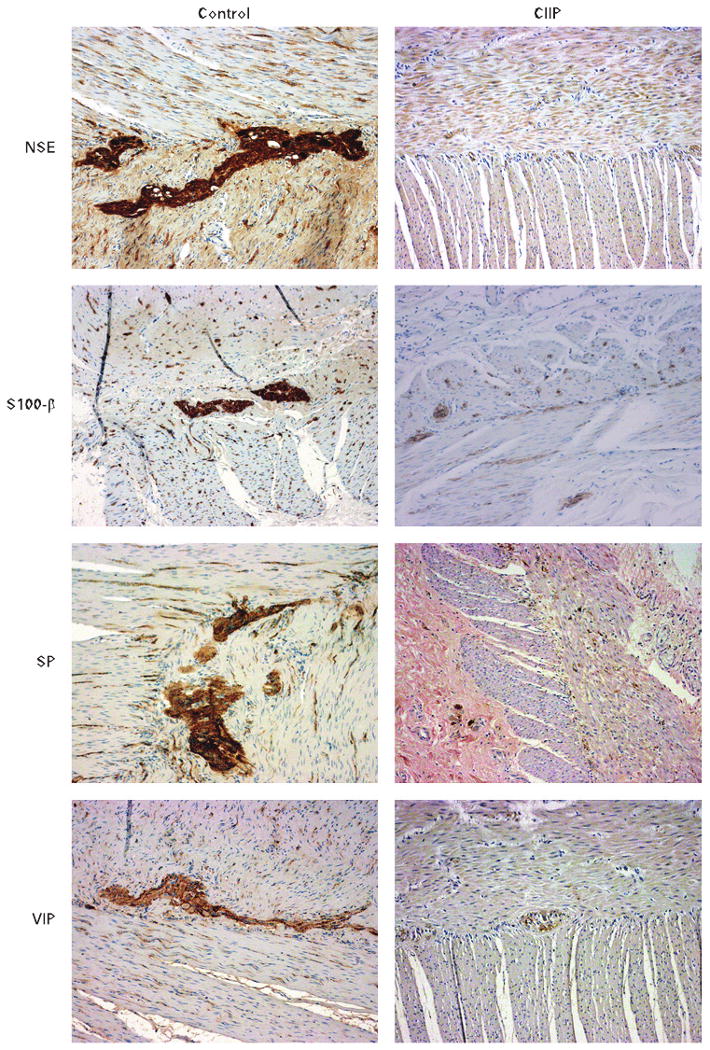
Immunohistochemical staining of the enteric neural network in patients with chronic idiopathic intestinal pseudo-obstruction (CIIP) compared with controls. Representative examples of immunolabelling experiments on cross-sections of patients with CIIP investigated in the present work. Note the marked reduction in the density and intensity of non-specific enolase- (NSE), enteroglial protein S100-β-, substance P/related tachykinins- (SP) and vasoactive intestinal polypeptide (VIP)-immunoreactive perikarya and nerve processes (right panels) compared with controls (left panels).
JCV TAg DNA sequences are frequently present in the myenteric plexus of patients with CIIP
JCV TAg DNA sequences were found in the microdissected myenteric plexuses of 8/10 (80%) of the patients with CIIP (table 4) and in 3/31 (9.7%) of the control patients by PCR (fig 2A–C) (p<0.001). Each of the positive PCR amplicons was confirmed by DNA sequencing. All samples and controls showed no evidence of BKV, SV40, HSV, CMV, VZV and EBV.
Table 4.
JC virus infection in chronic idiopathic intestinal pseudo-obstruction (CIIP) patients and controls
| Patient no | TAg DNA | TAg IHC | VP1 IHC |
|---|---|---|---|
| 1 | Positive | Positive | Positive |
| 2 | Positive | Positive | Positive |
| 3 | Positive | Positive | Positive |
| 4 | Positive | Positive | Positive |
| 5 | Negative | Negative | Negative |
| 6 | Negative | Negative | Negative |
| 7 | Positive | Negative | Negative |
| 8 | Positive | Positive | Positive |
| 9 | Positive | Positive | Positive |
| 10 | Positive | Positive | Positive |
| Control samples: | |||
| Ileal samples31 | 2/31 positive | 31 negative | 31 negative |
| Colon samples30 | 1/30 positive | 30 negative | 30 negative |
All samples and controls showed no evidence of BK virus, simian virus 40, cytomegalovirus, herpes simplex virus, varicella zoster virus and Epstein–Barr virus. IHC, immunohistochemistry; TAg, T antigen; VP1, viral protein 1.
FISH analysis was performed to verify the localisation of JCV DNA in the myenteric plexuses of patients with CIIP. All eight PCR-positive CIIP samples hybridised with JCV probes in the myenteric plexuses, while no signal was found in any of the three normal control samples from which JCV DNA was amplified (fig 2D–F).
JCV actively infects glial cells of patients with CIIP
Figure 3A shows positive and negative immunohistochemical staining for TAg in the myenteric plexuses of patients with CIIP and controls, respectively. In total, 7/10 (70%) patients with CIIP showed TAg expression, including 5 ileal and 2 colonic samples respectively. All 7 TAg-positive CIIP samples harboured JCV DNA sequences; however, one CIIP sample that carried JCV TAg sequences did not express TAg (patient 7 in Fig. 3A). No TAg expression was present in any of the 61 samples from 31 normal control patients.
Figure 3.
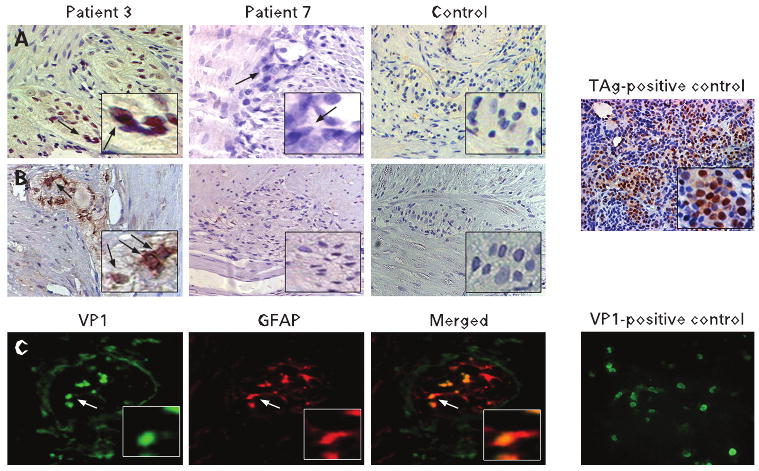
Immunohistochemical staining of myenteric plexuses for JC virus (JCV) T antigen (TAg), JCV viral protein 1 (VP1) and glial fibrillary acidic protein (GFAP). Upper panel. Immunohistochemistry shows expression of TAg (Pab2003) (A) and VP1 (B) in a patient affected by CIIP (patient 3). This patient was found to be positive for JCV DNA by both PCR and fluorescene is situ hybridisation. Patient 7, who was found positive for JCV DNA, tested negative for both TAg and VP1 expression. For a negative control we employed myenteric plexuses from patients affected by colon cancer. For a positive control for TAg we employed hamster brain tissues expressing the protein. Lower panel: (C) dual immunofluorescence shows co-localisation (merged panel) between VP1 expression and GFAP, indicating that the virus actively replicates in enteric glial cells. As positive controls for VP1 expression we employed human glial cells expressing the protein.
The IHC staining for the JCV capsid protein VP1 revealed the presence of the protein in the TAg protein-positive CIIP samples. Figure 3B demonstrates VP1 expression in the cells within the myenteric plexus of CIIP patient 3, suggesting an active lytic JCV infection. As expected, all TAg protein-negative CIIP samples and controls did not express VP1 in the myenteric plexuses.
We performed double-labelling immunofluorescence for JCV VP1 and the human GFAP. As shown in fig 3C, we found co-localisation of JCV VP1 and GFAP, indicating that the virus specifically infects the glial cells of CIIP myenteric plexuses.
Discussion
In our study, we demonstrate that JCV actively infects the enteroglial cells of the myenteric plexus of patients with CIIP. Although this study cannot establish a definitive causative role, our results provide novel evidence for an association between JCV infection and CIIP, which is consistent with the interpretation that this neurotropic virus may contribute to the pathogenesis of neurogenic CIIP by destroying the ENS through a lytic glial cell infection.
The patients included in this study suffered from a wide range of gastrointestinal symptoms, resulting in challenging clinical management. Although the diagnosis of CIIP is essentially clinical, mechanical causes must be excluded by endoscopic and radiological investigations.1 3 4 21 In this study, the patients underwent extensive diagnostic procedures, including small bowel manometry, to provide a definitive diagnostic characterisation of the disease. Furthermore, we established the diagnosis on the basis of a careful histopathological analysis. The diagnosis of an enteric (visceral) neuropathy was made in all the patients with CIIP.
Small bowel motor activities are strictly controlled by the ENS, and derangements of this mechanism result in well defined motor abnormalities that can be recorded in humans by manometric techniques.23–25 All the patients presented obvious motor abnormalities characterised by uncoordinated contractile patterns suggestive of intrinsic neuropathy.27 28 High pressure contractions were recorded in all but one patient whose manometric recording showed a retrograde phase III of the interdigestive MMC, with extremely low contractile pressures. This unique combination of motor abnormalities could be attributed to an intrinsic neuropathy associated with a concomitant involvement of smooth muscle cells or simply to overdistended bowel loops that prevent contractions from being occlusive, and, therefore, properly recordable by manometry. None of the patients had an inability to convert fasting into fed motility, which is considered to be suggestive of extrinsic autonomic neuropathy.29 30
Structural changes in the neurons and glia, as identified by NSE and S100-β, respectively, in association with swollen cell bodies and loss of myenteric neurons, can differentiate degenerative from inflammatory neuropathies. Alterations in the ICC network were found in 80% of the patients with CIIP. These findings illustrate the critical role played by these non-neuronal cells in the regulation of gut motility.31–34 The mechanisms for ICC depletion, as indicated by loss of c-Kit, remain unknown. Reduced c-Kit immunoreactivity may also reflect downregulation of this receptor and not necessarily a reduced number of ICCs. JCV may infect ICCs and therefore contribute either to ICC loss or to re-differentiation to smooth muscle cells, another event which has been proposed in pathological conditions.34 Both these mechanisms will be addressed in future research.
Moreover, we also investigated several neuropeptides, neurotransmitters and related receptors, such as CGRP, NPY, VIP, nNOS and SP, in the CIIP samples. We demonstrated a marked reduction of these markers, indicating an impairment of neurochemical transmitters or modulators, and damage to enteric neuron subclasses.
In seven patients with CIIP, both JCV TAg and the VP1 proteins were selectively expressed in the myenteric plexuses. Expression of both JCV TAg and VP1 indicates an active lytic infection, which would be expected to destroy the host cell. The lytic cycle begins by expression of the viral early protein TAg, followed by expression of the late genes, which are essential for a productive infection.35
Eight of 10 patients with CIIP were positive for JCV by PCR, whereas 3 of 31 control patients were positive. The explanation for these results might be that low level JCV infection is nearly universal among adults,15 and that the PCR can detect low levels of latent virus. We believe the higher prevalence of JCV DNA found in the CIIP specimens is attributable to the specific DNA extraction from the myenteric plexuses. Importantly, FISH analyses were positive only in the myenteric plexuses of patients with CIIP, and were negative in the samples from the unaffected subjects. This was supported by quantitative PCR data indicating that the DNA copy numbers in the three controls were at the lower limits of detection (data not shown). However, the most compelling argument for the speculation that JCV is actively involved in the pathogenesis of CIIP is that we never saw viral gene expression in any specimen except in the myenteric plexuses of patients with CIIP.
None of our samples or controls was positive for other known neurotropic viruses such as CMV, VZV, HSV and EBV, or other polyomaviruses such as BKV and SV40.
The most intriguing aspect of this study is that we found JCV infection in enteric glial cells. VP1 expression co-localised with the expression of GFAP, an intermediate filament found in glial cells, such as astrocytes. In the peripheral nervous system, GFAP has been demonstrated in Schwann cells, enteric glial cells, and satellite cells of human sensory ganglia. Increasing evidence suggests that damage to intestinal glial cells may be involved in the development of certain inflammatory and dysmotility diseases of the gut, including those induced by neurotropic viruses.36 37 A presumptive lytic infection produced by JCV, and the consequent damage to the plexuses, is a plausible explanation for the alterations found in CIIP. JCV causes the fatal demyelinating disease PML in glial cells of the CNS in severely immunocompromised individuals.19 To enter into a lytic phase, the virus has a narrow range of cell-specific tropism, as demonstrated by several in vitro models showing that the virus can replicate only in cells of glial origin. Similar mechanisms may occur in the ENS, which shares structural and neurochemical features with the CNS. The enteroglial cells (closely resembling the astrocytes of the CNS) targeted by the JCV infection may develop abnormalities which prevent their supportive and trophic mechanisms to neurons. As a consequence, the neuronal component would be impaired in its maintenance and survival. Unfortunately, there is no suitable in vivo model in which viral particles induce pathological changes in the brains of infected animals.38 However, a transgenic model has demonstrated specific viral replication in cells of glial origins.39 The well recognised pathogenetic activity of JCV in the CNS seen in PML, the presence of JCV infection of the ENS of patients with CIIP, and the evidence for a lytic infection in these patients provides an opportunity for novel insights into this disabling disease.
The samples obtained from the patients with CIIP had increased levels of apoptosis in the myenteric plexuses. This is of particular interest since oligodendrocytes of patients affected by PML also show high levels of apoptosis,40 and JCV agnoprotein has been shown to induce apoptosis in transfected oligodendrocytes.41 Enteric glial cells protect the ENS, and there is increasing evidence that enteric glia may regulate neuronal activity, similarly to glia in the CNS. Studies on ENS-associated neuropathies have mainly focused on patterns of neuropeptide expression and the involvement of the enteric neurons.36 However, there is increasing evidence which supports a central role for the enteric glia in intestinal dysmotility disorders.
Mouse models of glia disruption have gathered conflicting results. In our patients with CIIP, we could not find prominent inflammation either in the plexuses or in the mucosa. This is in contrast to a previously published transgenic mouse model in which the ablation of the enteric glia led to fulminant jejunoileitis,42 but in keeping with the model proposed by Aubè and colleagues43 in which the authors do not describe prominent inflammation either in the plexuses or in the mucosa, but altered motility patterns.
We explored JCV as a possible candidate in the pathogenesis of CIIP because of observations made in the CNS. The expression of both TAg and viral capsid proteins in enteroglial cells clearly demonstrates that the virus actively infects the enteric glia in CIIP. We had three patients in whom we could not find this, suggesting that other factors may be involved in CIIP.
We cannot exclude the possibility that the role of JCV might be secondary to the underlying disease, meaning an opportunistic infection rather than a causative agent. However, our results show that in healthy controls the virus is rarely present at the DNA level in the myenteric plexuses (3/31), while in the patients with CIIP the virus is in a productive lytic cycle in 70% of the cases. This result supports the hypothesis that the virus might have an active role in the development of this disease.
The association between JCV infection and CIIP leads to the hypothesis that this neurotropic virus may undergo activation from a latent state in the gut to an active lytic infection, with serious clinical consequences in affected patients. While further evidence is needed to demonstrate a causative role for JCV in CIIP, these findings open new perspectives in the pathophysiology and treatment of CIIP, and possibly other idiopathic motility disorders of the gut.
Supplementary Material
Acknowledgments
The authors wish to thank Drs Alessandra Gori and Gema Vera Pasamontes, Department of Internal Medicine and Gastroenterology, University of Bologna, Italy for their skilful technical expertise in processing the full-thickness biopsies and in the histopathological and immunohistochemical techiques. We wish to express gratitude to Dr Catia Sternini, Center for Ulcer Research and Education/ Digestive Diseases Center, UCLA School of Medicine, Los Angeles, California for kind donation of the following antibodies: vasoactive intestinal polypeptide (VIP) (antibody code: 7913), substance P/related tachykinins (SP/TK) (8701), neuropeptide Y (NPY) (8711) and calcitonin gene-related peptide (CGRP) (4901). The authors gratefully thank Dr Richard Frisque (Department of Biochemistry and Molecular Biology, Penn State University, University Park, Pennsylvania) for providing the mouse monoclonal antibody against JCV TAg, Dr Kamel Khalili (Department of Biology, Center for Neurovirology and Cancer Biology, Temple University Philadelphia, Pennsylvania) for providing hamster brain expressing JCV TAg, and Dr Walter Atwood (Department of Molecular Biology, Cell Biology & Biochemistry, Brown University, Providence, Rhode Island) for providing human glial cells expressing JCV VP1 and the anti-VP1 antibody.
Funding: This work was funded by: Grant R01 CA98572 from the National Cancer Institute of the NIH (to CRB), and funds from the Baylor Research Institute; by grants from the Italian Ministry of University, Research, Science and Technology (COFIN Projects number 2002068514_002, 2003064378_003, 2004062155_003) and RFO funds from the University of Bologna to VS, RDeG, GB and RC. RDeG is a recipient of a grant from the “Fondazione Del Monte di Bologna e Ravenna”.
Footnotes
Competing interests: None.
Ethics approval: Institutional Review Board approval was granted for this study from Baylor University Medical Center and from the University of Bologna.
Additional figures and tables are published online only at http://gut.bmj.com/content/vol58/issue1
References
- 1.Coulie B, Camilleri M. Intestinal pseudo-obstruction. Annu Rev Med. 1999;50:37–55. doi: 10.1146/annurev.med.50.1.37. [DOI] [PubMed] [Google Scholar]
- 2.De Giorgio R, Guerrini S, Barbara G, et al. Inflammatory neuropathies of the enteric nervous system. Gastroenterology. 2004;126:1872–83. doi: 10.1053/j.gastro.2004.02.024. [DOI] [PubMed] [Google Scholar]
- 3.Di Lorenzo C. Pseudo-obstruction: current approaches. Gastroenterology. 1999;116:980–7. doi: 10.1016/s0016-5085(99)70082-x. [DOI] [PubMed] [Google Scholar]
- 4.Stanghellini V, Corinaldesi R, Barbara L. Pseudo-obstruction syndromes. Baillieres Clin Gastroenterol. 1988;2:225–54. doi: 10.1016/0950-3528(88)90029-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.De Giorgio R, Guerrini S, Barbara G, et al. New insights into human enteric neuropathies. Neurogastroenterol Motil. 2004;16(Suppl 1):143–7. doi: 10.1111/j.1743-3150.2004.00491.x. [DOI] [PubMed] [Google Scholar]
- 6.De Giorgio R, Camilleri M. Human enteric neuropathies: morphology and molecular pathology. Neurogastroenterol Motil. 2004;16:515–31. doi: 10.1111/j.1365-2982.2004.00538.x. [DOI] [PubMed] [Google Scholar]
- 7.Chang AE, Young NA, Reddick RL, et al. Small bowel obstruction as a complication of disseminated varicella-zoster infection. Surgery. 1978;83:371–4. [PubMed] [Google Scholar]
- 8.Debinski HS, Kamm MA, Talbot IC, et al. DNA viruses in the pathogenesis of sporadic chronic idiopathic intestinal pseudo-obstruction. Gut. 1997;41:100–6. doi: 10.1136/gut.41.1.100. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Press MF, Riddell RH, Ringus J. Cytomegalovirus inclusion disease. Its occurrence in the myenteric plexus of a renal transplant patient. Arch Pathol Lab Med. 1980;104:580–3. [PubMed] [Google Scholar]
- 10.Robertson CS, Martin BA, Atkinson M. Varicella-zoster virus DNA in the oesophageal myenteric plexus in achalasia. Gut. 1993;34:299–302. doi: 10.1136/gut.34.3.299. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Sonsino E, Mouy R, Foucaud P, et al. Intestinal pseudoobstruction related to cytomegalovirus infection of myenteric plexus. N Engl J Med. 1984;311:196–7. doi: 10.1056/NEJM198407193110319. [DOI] [PubMed] [Google Scholar]
- 12.Vassallo M, Camilleri M, Caron BL, et al. Gastrointestinal motor dysfunction in acquired selective cholinergic dysautonomia associated with infectious mononucleosis. Gastroenterology. 1991;100:252–8. doi: 10.1016/0016-5085(91)90609-o. [DOI] [PubMed] [Google Scholar]
- 13.Knowles WA. Discovery and epidemiology of the human polyomaviruses BK virus (BKV) and JC virus (JCV) Adv Exp Med Biol. 2006;577:19–45. doi: 10.1007/0-387-32957-9_2. [DOI] [PubMed] [Google Scholar]
- 14.Laghi L, Randolph AE, Chauhan DP, et al. JC virus DNA is present in the mucosa of the human colon and in colorectal cancers. Proc Natl Acad Sci USA. 1999;96:7484–9. doi: 10.1073/pnas.96.13.7484. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Ricciardiello L, Laghi L, Ramamirtham P, et al. JC virus DNA sequences are frequently present in the human upper and lower gastrointestinal tract. Gastroenterology. 2000;119:1228–35. doi: 10.1053/gast.2000.19269. [DOI] [PubMed] [Google Scholar]
- 16.Barbanti-Brodano G, Sabbioni S, Martini F, et al. BK virus, JC virus and simian virus 40 infection in humans, and association with human tumors. Adv Exp Med Biol. 2006;577:319–41. doi: 10.1007/0-387-32957-9_23. [DOI] [PubMed] [Google Scholar]
- 17.Frisque RJ, Hofstetter C, Tyagarajan SK. Transforming activities of JC virus early proteins. Adv Exp Med Biol. 2006;577:288–309. doi: 10.1007/0-387-32957-9_21. [DOI] [PubMed] [Google Scholar]
- 18.Khalili K, Gordon J, White MK. The polyomavirus, JCV and its involvement in human disease. Adv Exp Med Biol. 2006;577:274–87. doi: 10.1007/0-387-32957-9_20. [DOI] [PubMed] [Google Scholar]
- 19.Padgett BL, Walker DL, ZuRhein GM, et al. Cultivation of papova-like virus from human brain with progressive multifocal leucoencephalopathy. Lancet. 1971;1:1257–60. doi: 10.1016/s0140-6736(71)91777-6. [DOI] [PubMed] [Google Scholar]
- 20.De Giorgio R, Barbara G, Stanghellini V, et al. Clinical and morphofunctional features of idiopathic myenteric ganglionitis underlying severe intestinal motor dysfunction: a study of three cases. Am J Gastroenterol. 2002;97:2454–9. doi: 10.1111/j.1572-0241.2002.06002.x. [DOI] [PubMed] [Google Scholar]
- 21.Stanghellini V, Cogliandro RF, De Giorgio R, et al. Natural history of chronic idiopathic intestinal pseudo-obstruction in adults: a single center study. Clin Gastroenterol Hepatol. 2005;3:449–58. doi: 10.1016/s1542-3565(04)00675-5. [DOI] [PubMed] [Google Scholar]
- 22.Busacchi P, Perri T, Paradisi R, et al. Abnormalities of somatic peptide-containing nerves supplying the pelvic floor of women with genitourinary prolapse and stress urinary incontinence. Urology. 2004;63:591–5. doi: 10.1016/j.urology.2003.09.017. [DOI] [PubMed] [Google Scholar]
- 23.Malagelada JR, Stanghellini V. Manometric evaluation of functional upper gut symptoms. Gastroenterology. 1985;88:1223–31. doi: 10.1016/s0016-5085(85)80083-4. [DOI] [PubMed] [Google Scholar]
- 24.Quigley EM, Deprez PH, Hellstrom P, et al. Ambulatory intestinal manometry: a consensus report on its clinical role. Dig Dis Sci. 1997;42:2395–400. doi: 10.1023/a:1018803819455. [DOI] [PubMed] [Google Scholar]
- 25.Stanghellini V, Camilleri M, Malagelada JR. Chronic idiopathic intestinal pseudo-obstruction: clinical and intestinal manometric findings. Gut. 1987;28:5–12. doi: 10.1136/gut.28.1.5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Goel A, Li MS, Nagasaka T, et al. Association of JC virus T-antigen expression with the methylator phenotype in sporadic colorectal cancers. Gastroenterology. 2006;130:1950–61. doi: 10.1053/j.gastro.2006.02.061. [DOI] [PubMed] [Google Scholar]
- 27.Husebye E. The patterns of small bowel motility: physiology and implications in organic disease and functional disorders. Neurogastroenterol Motil. 1999;11:141–61. doi: 10.1046/j.1365-2982.1999.00147.x. [DOI] [PubMed] [Google Scholar]
- 28.Kellow JE. Schuster atlas of gastrointestinal motility in health and disease. Small intestine: normal function and clinical disorders. In: Schuster MM, Crowell MD, Koch KL, editors. Manometry. London: BC Decker; 2002. pp. 219–236. [Google Scholar]
- 29.Hall KE, el-Sharkawy TY, Diamant NE. Vagal control of canine postprandial upper gastrointestinal motility. Am J Physiol. 1986;250:G501–10. doi: 10.1152/ajpgi.1986.250.4.G501. [DOI] [PubMed] [Google Scholar]
- 30.Thompson DG, Ritchie HD, Wingate DL. Patterns of small intestinal motility in duodenal ulcer patients before and after vagotomy. Gut. 1982;23:517–23. doi: 10.1136/gut.23.6.517. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Boeckxstaens GE, Rumessen JJ, de WL, et al. Abnormal distribution of the interstitial cells of cajal in an adult patient with pseudo-obstruction and megaduodenum. Am J Gastroenterol. 2002;97:2120–6. doi: 10.1111/j.1572-0241.2002.05932.x. [DOI] [PubMed] [Google Scholar]
- 32.Feldstein AE, Miller SM, El-Youssef M, et al. Chronic intestinal pseudoobstruction associated with altered interstitial cells of cajal networks. J Pediatr Gastroenterol Nutr. 2003;36:492–7. doi: 10.1097/00005176-200304000-00016. [DOI] [PubMed] [Google Scholar]
- 33.Isozaki K, Hirota S, Miyagawa J, et al. Deficiency of c-kit+ cells in patients with a myopathic form of chronic idiopathic intestinal pseudo-obstruction. Am J Gastroenterol. 1997;92:332–4. [PubMed] [Google Scholar]
- 34.Sanders KM, Ordog T, Ward SM. Physiology and pathophysiology of the interstitial cells of Cajal: from bench to bedside. IV. Genetic and animal models of GI motility disorders caused by loss of interstitial cells of Cajal. Am J Physiol Gastrointest Liver Physiol. 2002;282:G747–56. doi: 10.1152/ajpgi.00362.2001. [DOI] [PubMed] [Google Scholar]
- 35.Lynch KJ, Frisque RJ. Factors contributing to the restricted DNA replicating activity of JC virus. Virology. 1991;180:306–17. doi: 10.1016/0042-6822(91)90035-a. [DOI] [PubMed] [Google Scholar]
- 36.Savidge TC, Sofroniew MV, Neunlist M. Starring roles for astroglia in barrier pathologies of gut and brain. Lab Invest. 2007;87:731–6. doi: 10.1038/labinvest.3700600. [DOI] [PubMed] [Google Scholar]
- 37.Savidge TC, Newman P, Pothoulakis C, et al. Enteric glia regulate intestinal barrier function and inflammation via release of S-nitrosoglutathione. Gastroenterology. 2007;132:1344–58. doi: 10.1053/j.gastro.2007.01.051. [DOI] [PubMed] [Google Scholar]
- 38.Tooze J. The molecular biology of tumor viruses. New York: Cold Spring Harbor; 1981. [Google Scholar]
- 39.Trapp BD, Small JA, Pulley M, et al. Dysmyelination in transgenic mice containing JC virus early region. Ann Neurol. 1988;23:38–48. doi: 10.1002/ana.410230108. [DOI] [PubMed] [Google Scholar]
- 40.Richardson-Burns SM, Kleinschmidt-DeMasters BK, DeBiasi RL, et al. Progressive multifocal leukoencephalopathy and apoptosis of infected oligodendrocytes in the central nervous system of patients with and without AIDS. Arch Neurol. 2002;59:1930–6. doi: 10.1001/archneur.59.12.1930. [DOI] [PubMed] [Google Scholar]
- 41.Merabova N, Kaniowska D, Kaminski R, et al. JC Virus agnoprotein inhibits in vitro differentiation of oligodendrocytes and promotes apoptosis. J Virol. 2008;82:1558–69. doi: 10.1128/JVI.01680-07. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Bush TG, Savidge TC, Freeman TC, et al. Fulminant jejuno-ileitis following ablation of enteric glia in adult transgenic mice. Cell. 1998;93:189–201. doi: 10.1016/s0092-8674(00)81571-8. [DOI] [PubMed] [Google Scholar]
- 43.Aube AC, Cabarrocas J, Bauer J, et al. Changes in enteric neurone phenotype and intestinal functions in a transgenic mouse model of enteric glia disruption. Gut. 2006;55:630–7. doi: 10.1136/gut.2005.067595. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.