

Abstract
Human body temperature is regulated within a very narrow range. When exposed to hyperthermic conditions, via environmental factors and/or increased metabolism, heat dissipation becomes vital for survival. In humans, the primary mechanism of heat dissipation, particularly when ambient temperature is higher than skin temperature, is evaporative heat loss secondary to sweat secretion from eccrine glands. While the primary controller of sweating is the integration between internal and skin temperatures, a number of non-thermal factors modulate the sweating response. In addition to summarizing the current understanding of the neural pathways from the brain to the sweat gland, as well as responses at the sweat gland, this review will highlight findings pertaining to studies of proposed non-thermal modifiers of sweating, namely, exercise, baroreceptor loading state, and body fluid status. Information from these studies not only provides important insight pertaining to the basic mechanisms of sweating, but also perhaps could be useful towards a greater understanding of potential mechanisms and consequences of disease states as well as aging in altering sweating responses and thus temperature regulation.
Keywords: Sweat gland, Non-thermoregulatory factors, Exercise, Baroreflex, Body fluid regulation
2. INTRODUCTION AND HISTORICAL PERSPECTIVE
Evaporative heat loss is critical for human survival in a hot environment, particularly when environmental temperature is higher than skin temperature. Exercise or exposure to a hot environment elevates internal and skin temperatures, and subsequently increases sweat rate and skin blood flow. Historically it was thought that skin temperature was more important than internal temperature in the control of sweating (24, 109). In 1956 Kuno (46) proposed a novel concept that sweating responses were primarily controlled by a central thermoregulatory center, although he did not evaluate sweating as a function of internal temperature in those studies. Later, Benzinger was the first to present a relationship between internal temperature and sweat rate (5, 6) and proposed that ‘under steady state conditions increases in sweat rate during exercise and/or variations in the environmental temperature were very closely correlated to the elevation in tympanic temperature; a finding later supported by Nielsen and Nielsen (69). However Nielsen and Nielsen emphasized an importance of skin temperature given that rapid decreases in mean skin temperature reduced sweat rate in the absence of a change in internal temperature. With the understanding that internal and mean skin temperatures both have the capability to control sweating, researchers began to assess the relationship between contributions of skin to internal temperature in the modulation of sweat rate (25, 59, 64, 77, 78, 93, 108). Early in the 1970’s, Nadel and colleagues (65, 66) performed seminal work in this area during ‘dynamic’ increases in internal temperature in humans. The question of the influence of internal and skin temperatures in governing sweating was further addressed in non-human primates in which direct measures of brain temperature were obtained (91). Those studies concluded that sweating is primarily controlled by brain temperature and secondarily modulated by mean skin temperature, which is generally the current consensus of the scientific community. The concept of mean body temperature originated from these studies, with this variable being a weighed sum of internal and mean skin temperatures (64, 65).
Given these and other findings, sweating responses are now commonly characterized by the internal or mean body temperature threshold for the onset of sweating, as well as the slope of the relationship between the elevation in sweating relative to the elevation in internal or mean body temperature, as eloquently presented by Gisolfi and Wenger (19). They proposed that an increase in the internal or mean body temperature threshold for the onset of sweating and/or an attenuation of the elevation in sweating relative to the elevation in internal or mean body temperature is representative of impaired sweating responsiveness. Conversely, a reduced internal or mean body temperature threshold for the onset of sweating and/or an elevated slope is representative of enhanced sweating responsiveness, as which occurs with heat acclimation.
3. NEURAL PATHWAY FROM THE BRAIN TO SWEAT GLAND
The primary thermoregulatory center, first reported in the late 1800s, is located within the pre-optic hypothalamic regions of the brain (4, 34, 63, 71). Because of the difficulty of precisely identifying neural pathways responsibility for sweating, in humans these pathways are not entirely understood. However based upon evidence from animal studies and human anatomical data (46, 51, 67, 80), the neural pathway from the brain to sweat gland is thought to be as follows; efferent signals from the pre-optic hypothalamus travel via the tegmentum of the pons and the medullary raphe regions to the intermediolateral cell column of the spinal cord. In the spinal cord, neurons emerge from the ventral horn, pass through the white ramus communicans and then synapse in the sympathetic ganglia. Postganglionic non-myelinated C-fibers pass through the gray ramus communicans, combine with peripheral nerves and travel to sweat glands, with these nerve fibers “entwined around” the periglandular tissue of the eccrine sweat gland (99).
Direct recordings of human post-ganglionic skin sympathetic nerve activity (SSNA) are accomplished by the microneurography technique. Much of the original work characterizing SSNA was performed by Wallin and his colleagues (11, 23, 104). The neural signal recorded from this technique contains efferent activity responsible for sweating, cutaneous vasoconstriction, pilomotor responses, and perhaps cutaneous vasodilation (7, 8, 11). Due to this integrated nature of the SSNA recordings, caution must be taken when attempting to link an efferent response (e.g., sweating, cutaneous vasoconstriction, etc.) to specific neural activity. Nevertheless, during heat stress SSNA is partially synchronized with galvanic skin response (an index of sweating) and pulsatile sweat expulsion (7, 94), with approximately 80% of SSNA bursts being reported to be synchronized with pulsatile sweat expulsion (95). These observations suggest a dominance of the recorded SSNA signal in heat stressed subjects to be sudomotor in nature. However in mildly heat stressed individuals, increased SSNA bursts amplitude was related with increases in cutaneous vasodilation as well as sweat expulsion (35), thereby raising the possibility that active cutaneous vasodilator signals may also be contained within the integrated SSNA signal. Because sympathetic axons are clustered within a nerve fascicle, such recordings have been primarily limited to the analysis of multi-unit neural activity; although Macefield and Wallin recorded neural activity from single sympathetic neurons proposed to innervate sweat glands (55, 56). However, confirmation that the activity from a single unit recording specifically innervates sweat glands, at the exclusion of other structures (e.g., cutaneous blood vessels), would be challenging at best.
The sympathetic nerves distributed to sweat glands consist of large numbers of cholinergic terminals and a few adrenergic terminals (99). The effect of these adrenergic terminals in causing sweating is minimal given that exogenous administration of adrenergic agents will cause only minimal sweating relative to acetylcholine administration, the latter of which is the primary neurotransmitter causing sweating (73–75, 79). Local and systemic administration of atropine (a muscarinic receptor antagonist) greatly attenuates or abolishes sweating during a thermal challenge or during exogenous administration of acetylcholine or its analogs (17, 38, 40, 50, 57), further confirming the dominance of the cholinergic system and muscarinic receptors in human sweating.
In addition to a central neural drive for sweating, sweating can also be initiated by an axon reflex (53). Exogenous administration of acetylcholine, or its analogues, not only directly stimulates muscarinic receptors on sweat glands, but also activates an axon reflex via stimulation of axonal nicotinic receptors. The neural impulse due to the activated axon terminal is thought to travel antidromically to a branch-point, and then travel orthodromically to other nerve terminals, culminating in the release of acetylcholine (52). Thus not only the central drive from thermoregulatory center but also local mechanisms (e.g., perhaps endogenous acetylcholine stimulation of the axon reflex) can contribute to the modulation of sweating.
Acetylcholine released from cholinergic nerves is rapidly hydrolyzed by acetylcholinesterase (50). Thus acetylcholinesterase is capable of modulating sweat rate during low to moderate sweating activity but its effectiveness is greatly reduced when sweat rate is substantially increased (85). Consistent with this finding, elevations in sweat rate occur earlier with exogenous methacholine than with acetylcholine administration, given the reduced cholinesterase susceptibility of methacholine (39).
The neurotransmitter(s) responsible for active cutaneous vasodilation has yet to be fully elucidated, although neuropeptides such as calcitonin gene-related peptide (CGRP), vasoactive intestinal polypeptide (VIP), and substance P as well as nitric oxide (NO) have been implicated (37). See the chapter by J.M. Johnson in the current volume for further insight regarding these vasodilators (29). For a number of years researchers have inquired whether these peptides and NO modulate the sweating response. Studies have shown a presence of VIP immunoreactive nerve fibers around the eccrine sweat glands of cat foot pads (54) as well as in human eccrine sweat glands (13, 45). The function of these fibers is not entirely clear, although VIP enhances sweat secretion based upon findings from in vitro and in vivo studies (81, 110). The distribution of immunoreactivities towards atrial natriuretic peptide (ANP), calcitonin gene-related peptide (CGRP), galanin and substance P have been confirmed in human skin; although CGRP, but not substance P, have been specifically identified around eccrine sweat glands (96). Consistent with this observation, exogenous CGRP increases sweat rate, while exogenous substance P suppressed sweat rate (44, 84), during administration of sudorific agents. Finally NO also has been shown to augment sweat rate during exogenous acetylcholine administration as well as during an exercise heat stress (48, 105). Although acetylcholine is the primary neurotransmitter responsible for sweat secretion, enhanced sweating due to local administration of VIP, CGRP, or NO suggest that these peptides as well as NO may contribute to the overall modulation of sweating during a thermal challenge.
Aquaporins (AQPs) are a family of membrane water channel proteins. At least 10 mammalian aquaporins have been identified (1) and some have been implicated in physiological processes. AQP5 has been localized to the apical membrane of multiple secretory glands, including lacrimal glands, salivary glands and submucosal glands of airways (70). These glands facilitate the secretion of large amounts of fluid. Nejsum et al. (68) identified the distribution and function of AQP5 in the apical membranes of sweat glands in rat, mouse and humans. They demonstrated that sweat secretion was markedly diminished in paws of AQP5 null mice. Furthermore, AQP5 immunoreactivity was also observed from the dark cells of the secretory portion of human eccrine sweat glands (32). Given that AQP proteins have been identified in human sweat glands, coupled with findings that botulinum toxin inhibits water permeability via AQP-dependent mechanisms (72), botulinum toxin, which is recognized to abolish sweating (38), may do so via pre- and post- cholinergic synaptic mechanisms; although botulinum toxin is considered primarily as a pre-synaptic inhibitor of neurotransmission. Recently, Shibasaki et al. (86) found that local administration of botulinum toxin in human skin completely blocked the sweating response to exogenous acetylcholine, lending support to a post-synaptic mechanism by which botulinum toxin can abolish sweating. Future studies are warranted to identify the precise mechanism(s) by which botulinum toxin blocks sweating independent of inhibition of cholinergic neurotransmission.
4. NON-THERMAL MODULATORS OF SWEAT RATE
Independent of the aforementioned controllers and modulators of sweat rate, a number of perturbations have been suggested to alter the sweating response, specifically exercise, baroreceptor perturbations, and fluid/osmolality status (Figure 1).
Figure 1.
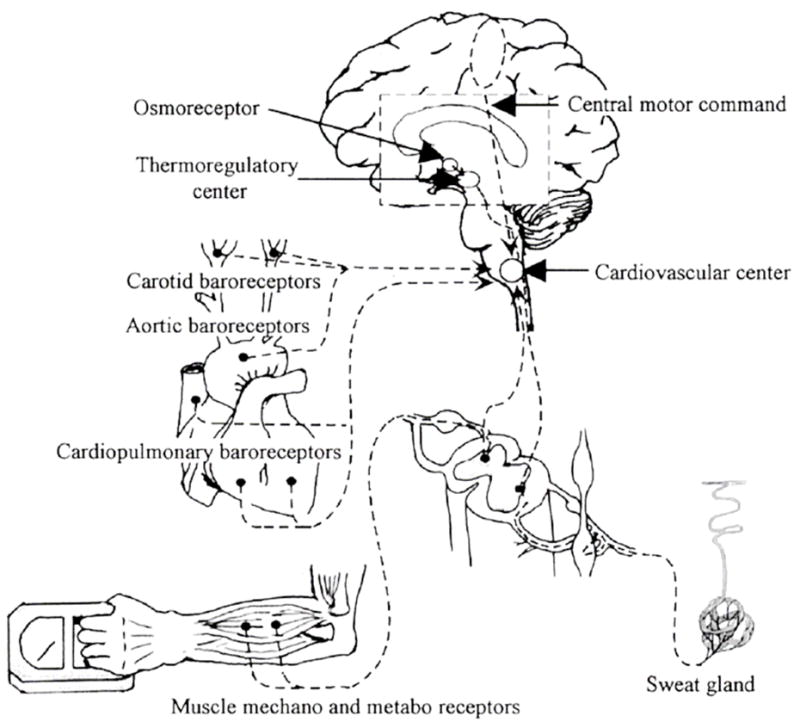
Schematic illustrating possible non-thermal modifiers of sweating. From Shibasaki et al. Exercise Sport Science Review 31(1): 34–39, 2003. Reprinted with permission from Wolters Kluwer/Lippincott, Williams & Wilkins.
4.1. Effect of exercise in modulating sweat rate
The mechanisms associated with human temperature regulation during exercise is complex, resulting in a number of proposed theories and concepts (19). Generation of heat associated with muscular contraction during dynamic exercise rapidly elevates internal temperature, followed by appropriate increases in sweat rate. It is interesting to note that factors unrelated to this elevation in internal temperature, which are engaged during exercise, modulate the sweating response. van Beaumont and Bullard (100, 101) were the first to report this phenomenon upon observing that sweating occurred immediately (within 1.5–2 sec) with the onset of dynamic exercise, as well as during isometric exercise of humans in warm environmental conditions (Figure 2). Importantly, the increase in sweating occurred prior to a measurable change in internal temperature. Later, Gisolfi and Robinson (18) observed rapid changes in sweating during intermittent exercise independent of changes in internal, muscle, or skin temperatures. Consistent with these observations, during sinusoidal dynamic exercise (i.e. workload was changed in sinusoidal manner) sweating response followed the change in workload but not skin or internal temperatures (111, 112). Together, these findings strongly suggest that non-thermal factors related to exercise (i.e. independent of skin and internal temperatures) modulate sweating, perhaps via a feed-forward mechanism.
Figure 2.

The first report that sweating can be induced by non-thermal factors. In heat stressed subjects, that were already sweating, performing “very hard work” resulted in immediate increases in calf and forearm sweat rate despite the absence of an increase in internal temperature. From van Beaumont & Bullard, Science, 141: 643–646, 1963. Reprinted with permission from AAAS.
To address the possible mechanism(s) by which exercise increases sweating independent of temperature, one needs to understand the work of Johansson (28) who postulated that two separate and distinct neural mechanisms control cardiovascular responses during exercise. One mechanism arises from the central nervous system that irradiates impulses from the motor cortex. Krogh and Lindhard (43) termed this central mechanism as “cortical irradiation” and later it was called “central command” (20). The other mechanism, termed the exercise pressor reflex, originates from the stimulation of afferent nerve endings within the skeletal muscle and is engaged during muscle contraction (3). Later it was shown that mechano and metabo-sensitive afferent nerves were responsible for evoking this exercise pressor reflex (60, 61). Since sweating during exercise can occur prior to a change in thermal status, coupled with the aforementioned responses associated with modulating cardiovascular responses during exercise, researchers sought to identify whether similar mechanisms could be responsible for modulating sweat rate during exercise.
Partial neuromuscular blockade (e.g. using curare derivatives) has been used to augment central command during exercise, resulting in greater increases in heart rate and blood pressure at a given workload (27, 49, 60). Shibasaki et al. (90) used this technique to test the hypothesis that central command is capable of modulating the sweating response. Subjects performed isometric handgrip exercise under control conditions (without neuromuscular blockade) and when central command was augmented via partial neuromuscular blockade. Under both conditions, isometric exercise increased sweat rate, however the increase in sweat rate was significantly greater when central command was augmented. This, and a related study assessing SSNA to isometric exercise during partial neuromuscular blockade (102), provide strong evidence that central command is capable of modulating sweating during exercise.
Alam and Smirk (2, 3) showed that blood pressure increases during dynamic and static exercise and remains elevated if blood flow to that limb was occluded just prior to the cessation of exercise. Upon release of the occlusion, blood pressure returns to pre-exercise levels. Their observations led to numerous and ongoing studies investigating the role of muscle metaboreceptors in modulating blood pressure during exercise. A number of studies have been performed to investigate the possible role of metaboreceptors in modulating sweating responses during exercise (9, 41, 87). In general, the cited studies were performed by monitoring sweat rate during isometric exercise and subsequent post-exercise ischemia, to isolate muscle metaboreceptor stimulation. In those studies sweat rate increased during isometric exercise, remained elevated during post-exercise ischemia, and then returned towards pre-exercise levels following release of ischemia. This pattern of response provides evidence that stimulation of muscle metaboreceptors is capable of modulating sweat rate during exercise. Interestingly, if the breakdown of acetylcholine was inhibited via local administration of neostigmine, the aforementioned metaboreceptor-dependent stimulating of sweating occurs even in non-heat stressed subjects (Figure 3; lower panels).
Figure 3.

Influence of isometric exercise on sweat rate in normothermic subjects. In normothermia, isometric exercise increased sweating rate (SR) at the neostigmine-treated site but not at the control site. Neostigmine is a cholinesterase inhibitor and thus inhibits the breakdown of acetylcholine. Sweat rate remained elevated during post-exercise ischaemia (PEI) at the neostigmine treated site regardless of whether mean arterial blood pressure (MAP) remained elevated during PEI (left panel) or was reduced via bolus infusion of sodium nitroprusside (right panel; see arrow). These data provide evidence that stimulation of muscle metaboreceptors can increase sweating. From Shibasaki, et al. Journal of Physiology, 534(Pt2): 605–611, 2001. Reprinted with permission from Blackwell Publishing.
During post-exercise ischemia blood pressure is also elevated and may therefore contribute to the elevation in sweating secondary to the loading of baroreceptors. To test this hypothesis, Shibasaki et al. (87) performed an experiment in which blood pressure during the post-exercise ischemia period was restored to pre-exercise levels via intravenous administration of sodium nitroprusside (Figure 3, right panel). Under these conditions muscle metaboreceptors remained stimulated but blood pressure returned to pre-exercise levels. Despite returning to pre-exercise levels, sweat rate remained elevated throughout the ischemic period (87). Thus, the elevation in sweat rate during post-exercise ischemia occurred through activation of metaboreceptors and was independent of the increase in blood pressure during post-exercise ischemia and presumably during isometric exercise. In addition, sweat rate is enhanced during dynamic exercise when performed in combination with lower body positive pressure (i.e. accumulation of metabolites due to partial ischemia of leg muscle blood flow) relative to exercise without the application of this positive pressure (14, 33). Together, these findings strongly suggest that the muscle metaboreflex is capable of modulating sweat rate.
Another muscle afferent signal that could contribute to sweating responses during exercise is that related to mechanical stimulation that occurs during muscle contraction (31, 42, 76, 89), which has been suggested to contribute to the exercise pressor response (60, 61). The cited studies used protocols involving passive limb movement or passive cycling to stimulate muscle mechanoreceptors, without central command and with little muscle metaboreceptor stimulation, while assessing sweating responses in heat stressed subjects. In general, these findings suggest that stimulation of muscle mechanoreceptors is capable of modulating sweat rate, although responses are appreciably less than that observed during augmentation of central command or muscle metaboreceptor stimulation.
4.2. Effects of baroreceptors in modulating sweat rate
Given that prolonged exposure to hyperthermic conditions and/or exercise reduces blood volume if fluid intake is not adequate, coupled with baroreceptors being sensitive to changes in blood volume through alterations in blood pressures (i.e., arterial and perhaps central venous blood pressures), it seems reasonable to hypothesize that sweating associated with these conditions could be modulated by baroreceptor unloading. However, the effects of baroreceptor unloading on attenuating the elevation in sweat rate are controversial. Johnson and Park (30) assessed the internal temperature threshold for the onset of sweating during exercise and found that this threshold was unaltered regardless of whether the individual exercised in the upright (i.e. baroreceptor unloading) or supine positions. In contrast, Mack et al. (58) observed an increase in the internal temperature threshold for the onset of sweating (i.e., a delayed sweating response) during exercise in combination with lower-body negative pressure (LBNP), which simulates the upright position and unloads baroreceptors.
The effect of baroreceptor unloading on sweat rate was further addressed by applying LBNP in passively (i.e., non-exercising) heat stressed subjects (10, 92, 103). These studies suggested that sweat rate was not affected by baroreceptor unloading. A possible explanation for differences in findings between LBNP studies (10, 58, 92, 103) was proposed by Vissing et al. (103) who suggested that reduced electrodermal response (index of sweating) and SSNA during LBNP resulted from skin cooling that frequently occurs upon application of LBNP, not via baroreceptor unloading. To address this question, Wilson et al. (106) assessed sweat rate and SSNA in heat stressed subjects during bolus and steady-state infusions of pharmacological agents (nitroprusside and phenylephrine) to perturb baroreceptors without causing cooling that accompanies LBNP. Despite pronounced changes in blood pressure, neither SSNA nor sweat rate was significantly affected. However, it should be stressed that pharmacologically-induced decreases in blood pressure will likely perturb baroreceptors differently relative to LBNP or head-up tilt.
Dodt et al. (12) addressed this question differently by exposing subjects to a mild heat stress, followed by 30° head-up tilt. They observed significant reductions in forearm SSNA and an index of sweat rate during tilt, and concluded that baroreceptor unloading could modulate SSNA and sweating. Differences in conclusions between Dodt et al.’s study and the findings of others (10, 92, 103, 106) may be related to the lower level of heat stress employed by Dodt et al. (12). For example, baroreceptors may be capable of modulating sweating under mild to moderate heating conditions but not during more pronounced heat stress. To address this question, Wilson et al. (107) measured SSNA and sweat rate during multiple 30° head-up tilts, with tilting occurring every 10 min throughout the heat stress. Regardless of the level of heating, they did not observe a reduction in sweat rate or SSNA during the same magnitude of tilt used by Dodt et al. (12). Taken together, although findings remain controversial, relatively acute unloading of baroreceptors (i.e. on the order of minutes) is unlikely to modulate sweat rate.
4.3. Effects of body fluid status and osmolality on sweat rate
Prolonged exposure to hyperthermic conditions and/or prolonged exercise in the heat can induce water deficits due to profuse sweating, resulting in hypohydration. This water deficit lowers both intracellular and extracellular volumes and results in plasma hyperosmolality and hypovolemia; both of which impair sweating. For example, Greenleaf and Castle (22) proposed that the excessive rise in internal temperature in dehydrated subjects was due to inadequate sweating secondary to the dehydration. Expanding this concept, Sawka et al. (83) observed that in progressively dehydrated subjects sweat rate was dramatically reduced despite greater elevations in rectal temperature. Later Montain, et al. (62) demonstrated that the threshold for the onset of sweating was elevated while the slope of the relationship between the elevation in sweat rate relative to the elevation in internal temperature was attenuated as a function of the level of dehydration; both of which are strongly suggestive that dehydration impairs sweating responsiveness.
Fortney et al. (16) conducted a study to identify the importance and independence of decreases in fluid volume (hypovolemia) from increases in plasma osmolality (hyperosmotic) on sweat rate. Normovolemic subjects were exposed to heat stresses under hyperosmotic and iso-osmotic conditions while sweat rate was assessed. During the ensuing exercise bout, the internal temperature threshold for the onset of sweating was significantly elevated relative to the response during exercise under iso-osmotic conditions, although the slope of the relationship between the elevation in sweating and the elevation in internal temperature was not affected by increased plasma osmolality. Takamata et al. (97, 98) extended these findings upon assessing sweat rate in heat stressed subjects who received an infusion of 0.9% or 3% saline. They found that the threshold for sweating in the hyperosmotic condition (i.e. 3% saline infusion) was greatly shifted to a higher internal temperature relative to the iso-osmotic condition (Figure 4). This hyperosmolality induced suppression of sweating occurred regardless of heat acclimation status (26).
Figure 4.
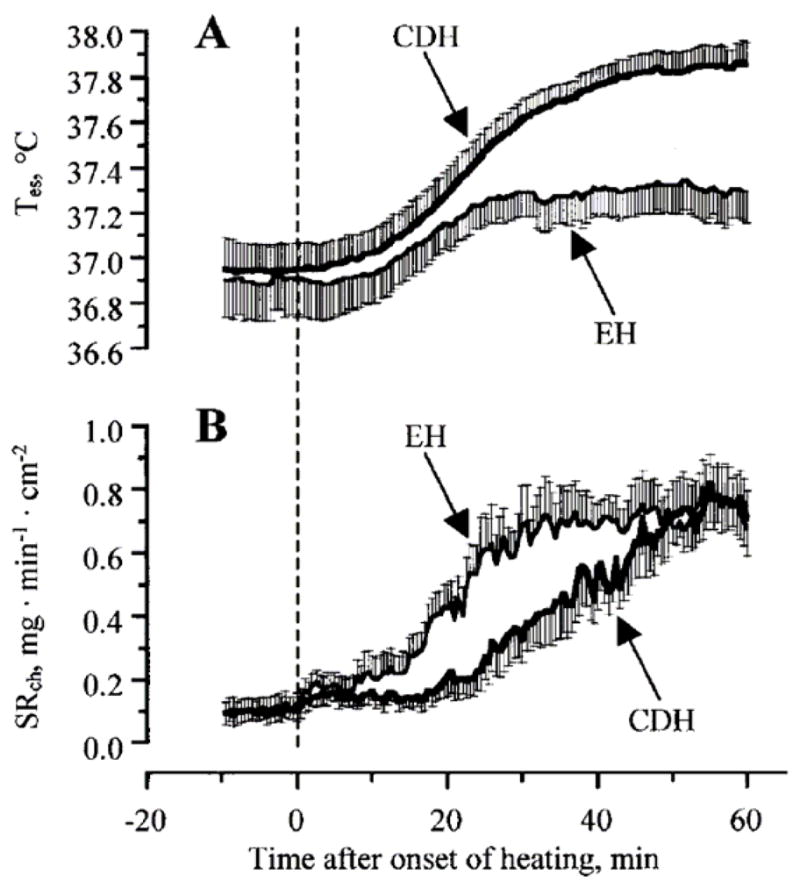
Effects of elevated plasma osmolality (cell dehydration; CDH) on body core temperature (Tes; panel A) and local chest sweat rate (SRch; panel B) in humans. During the heat stress, when plasma osmolality was elevated, the increase in body core temperature was greater, while sweat rate was significantly reduced, when compared with iso-osmotic (EH) conditions. Differences in local sweat rate between osmotic conditions were primarily due to a delay in the onset of sweating during the hyperosmotic heat stress. From Takamata, A. et al., American Journal of Physiology 268:R414 R422, 1995. Reprinted with permission from the American Physiological Society.
It is interesting to note that Takamata et al. (97) found that when hyperosmotic subjects drink deionized water (38 °C) that sweat rate immediately increased, and this occurred in the absence of changes in plasma osmolality. In contrast, drinking deionized water in iso-osmotic subjects did not alter sweat rate. In a follow-up study, Kamijo et al. (36) confirmed such a release of sweating suppression by drinking occurred during exercise. These investigators concluded that stimulation of an oral-pharyngeal reflex, associated with the act of drinking, releases an otherwise inhibition of sweating by dehydration. These findings demonstrate that increased plasma osmolality, independent of plasma volume, impairs sweating responses, and that stimulation of an oral-pharyngeal reflex can modulate the sweating response in hyperosmotic individuals (97).
Fortney et al. (15) addressed the opposite question relative to that presented above, in that they investigated whether changes in blood volume, while keeping plasma osmolality constant, modulates the sweating response. They found that iso-osmotic hypovolemia reduced the slope of the relationship between the change in sweating relative to the change in internal temperature, without altering the internal temperature threshold for the onset of sweating (15). Such a finding suggests that once sweating has begun, for the same elevation in internal temperature there was less of an elevation in sweating when the individuals were hypovolemic but iso-osmotic. Conversely, iso-osmotic hypervolemia did not change the internal temperature threshold for sweating nor the aforementioned slope (15, 47), unless plasma/blood volume expansion occurs via erythrocyte infusion (82). These observations suggest that sweating can be inhibited by iso-osmotic hypovolemia, whereas hypervolemia in the absence of erythrocyte infusion does not alter sweating responses.
5. SUMMARY
Neural control of sweating, primarily regulated by the integration of internal and skin temperatures, is paramount for temperature regulation. However, a variety of other non-thermal factors, such as factors associated with exercise and fluid status, modify the sweating response. Further studies are necessary to identify the precise mechanisms by which these non-thermal factors serve to accentuate or attenuate sweating in the resting and exercising human.
Acknowledgments
The authors acknowledge the many investigators who focus on this field of study. Support for the authors was provided by Grand-in-Aid for the Encouragement of Young Scientists from the Japanese Society for the Promotion of Science Grant 21790225 (to Shibasaki); National Heart Lung, and Blood Institute Grant HL-61388 and 84072 (to Crandall).
References
- 1.Agre P, King LS, Yasui M, Guggino WB, Ottersen OP, Fujiyoshi Y, Engel A, Nielsen S. Aquaporin water channels--from atomic structure to clinical medicine. J Physiol. 2002;542:3–16. doi: 10.1113/jphysiol.2002.020818. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Alam M, Smirk FH. Observations in man on a pulse accelerating reflex from the voluntary muscles of the legs. J Physiol. 1938;92:167–177. doi: 10.1113/jphysiol.1938.sp003592. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Alam M, Smirk FH. Observations in man upon a blood pressure raising reflex arising from the voluntary muscles. J Physiol. 1937;89:372–383. doi: 10.1113/jphysiol.1937.sp003485. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Aronsohn E, Sachs J. Die beziehungen des gehirns zur korperwairme und zum fieber. Pflugers Arch. 1885;37:232–300. [Google Scholar]
- 5.Benzinger TH. On physical heat regulation and the sense of temperature in man. Proc Natl Acad Sci USA. 1959;45:645–659. doi: 10.1073/pnas.45.4.645. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Benzinger TH. The sensory receptor organ and quantitative mechanism of human temperature control in warm environment. Fed Proc. 1960;19:32–43. [Google Scholar]
- 7.Bini G, Hagbarth KE, Hynninen P, Wallin BG. Thermoregulatory and rhythm-generating mechanisms governing the sudomotor and vasoconstrictor outflow in human cutaneous nerves. J Physiol. 1980;306:537–552. doi: 10.1113/jphysiol.1980.sp013413. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Blumberg H, Wallin BG. Direct evidence of neurally mediated vasodilatation in hairy skin of the human foot. J Physiol. 1987;382:105–121. doi: 10.1113/jphysiol.1987.sp016358. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Crandall CG, Stephens DP, Johnson JM. Muscle metaboreceptor modulation of cutaneous active vasodilation. Med Sci Sports Exerc. 1998;30:490–496. doi: 10.1097/00005768-199804000-00004. [DOI] [PubMed] [Google Scholar]
- 10.Cui J, Wilson TE, Crandall CG. Orthostatic challenge does not alter skin sympathetic nerve activity in heat-stressed humans. Auton Neurosci. 2004;116:54–61. doi: 10.1016/j.autneu.2004.08.009. [DOI] [PubMed] [Google Scholar]
- 11.Delius W, Hagbarth KE, Hongell A, Wallin BG. Manoeuvres affecting sympathetic outflow in human skin nerves. Acta Physiol Scand. 1972;84:177–186. doi: 10.1111/j.1748-1716.1972.tb05168.x. [DOI] [PubMed] [Google Scholar]
- 12.Dodt C, Gunnarsson T, Elam M, Karlsson T, Wallin BG. Central blood volume influences sympathetic sudomotor nerve traffic in warm humans. Acta Physiol Scand. 1995;155:41–51. doi: 10.1111/j.1748-1716.1995.tb09946.x. [DOI] [PubMed] [Google Scholar]
- 13.Eedy DJ, Shaw C, Armstrong EP, Johnston CF, Buchanan KD. Vasoactive intestinal peptide (VIP) and peptide histidine methionine (PHM) in human eccrine sweat glands: demonstration of innervation, specific binding sites and presence in secretions. Br J Dermatol. 1990;123:65–76. doi: 10.1111/j.1365-2133.1990.tb01825.x. [DOI] [PubMed] [Google Scholar]
- 14.Eiken O, Mekjavic IB. Ischaemia in working muscles potentiates the exercise-induced sweating response in man. Acta Physiol Scand. 2004;181:305–311. doi: 10.1111/j.1365-201X.2004.01289.x. [DOI] [PubMed] [Google Scholar]
- 15.Fortney SM, Nadel ER, Wenger CB, Bove JR. Effect of blood volume on sweating rate and body fluids in exercising humans. J Appl Physiol. 1981;51:1594–1600. doi: 10.1152/jappl.1981.51.6.1594. [DOI] [PubMed] [Google Scholar]
- 16.Fortney SM, Wenger CB, Bove JR, Nadel ER. Effect of hyperosmolality on control of blood flow and sweating. J Appl Physiol. 1984;57:1688–1695. doi: 10.1152/jappl.1984.57.6.1688. [DOI] [PubMed] [Google Scholar]
- 17.Foster KG, Weiner JS. Effects of cholinergic and adrenergic blocking agents on the activity of the eccrine sweat glands. J Physiol. 1970;210:883–895. doi: 10.1113/jphysiol.1970.sp009247. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Gisolfi CV, Robinson S. Central and peripheral stimuli regulating sweating during intermittent work in men. J Appl Physiol. 1970;29:761–768. doi: 10.1152/jappl.1970.29.6.761. [DOI] [PubMed] [Google Scholar]
- 19.Gisolfi CV, Wenger CB. Temperature regulation during exercise: old concepts, new ideas. Exerc Sport Sci Rev. 1984;12:339–372. [PubMed] [Google Scholar]
- 20.Goodwin GM, McCloskey DI, Mitchell JH. Cardiovascular and respiratory responses to changes in central command during isometric exercise at constant muscle tension. J Physiol. 1972;226:173–190. doi: 10.1113/jphysiol.1972.sp009979. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Greene RM, Winkelmann RK, Opfer-Gehrking TL, Low PA. Sweating patterns in atopic dermatitis patients. Arch Dermatol Res. 1989;281:373–376. doi: 10.1007/BF00455319. [DOI] [PubMed] [Google Scholar]
- 22.Greenleaf JE, Castle BL. Exercise temperature regulation in man during hypohydration and hyperhydration. J Appl Physiol. 1971;30:847–853. doi: 10.1152/jappl.1971.30.6.847. [DOI] [PubMed] [Google Scholar]
- 23.Hagbarth KE, Hallin RG, Hongell A, Torebjork HE, Wallin BG. General characteristics of sympathetic activity in human skin nerves. Acta Physiol Scand. 1972;84:164–176. doi: 10.1111/j.1748-1716.1972.tb05167.x. [DOI] [PubMed] [Google Scholar]
- 24.Hammel HT. Regulation of internal body temperature. Ann Rev Physiol. 1968;30:641–710. doi: 10.1146/annurev.ph.30.030168.003233. [DOI] [PubMed] [Google Scholar]
- 25.Hardy JD, Stolwijk JAJ. Partitional calorimetric studies of man during exposures to thermal transients. J Appl Physiol. 1966;21:1799–1806. doi: 10.1152/jappl.1966.21.6.1799. [DOI] [PubMed] [Google Scholar]
- 26.Ichinose T, Okazaki K, Masuki S, Mitono H, Chen M, Endoh H, Nose H. Ten-day endurance training attenuates the hyperosmotic suppression of cutaneous vasodilation during exercise but not sweating. J Appl Physiol. 2005;99:237–243. doi: 10.1152/japplphysiol.00813.2004. [DOI] [PubMed] [Google Scholar]
- 27.Iwamoto GA, Mitchell JH, Mizuno M, Secher NH. Cardiovascular responses at the onset of exercise with partial neuromuscular blockade in cat and man. J Physiol. 1987;384:39–47. doi: 10.1113/jphysiol.1987.sp016442. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Johansson JE. Uber die einwirkung der muskelthatigkeit auf die athmung und die herzthatigkeit. Skand Arch Phsiol. 1895;5:20–66. [Google Scholar]
- 29.Johnson JM, Kellogg DL., Jr Mechanisms of Control in the Human Cutaneous Circulation. Front Biosci. 2009 doi: 10.2741/s105. in print. [DOI] [PubMed] [Google Scholar]
- 30.Johnson JM, Park MK. Effect of upright exercise on threshold for cutaneous vasodilation and sweating. J Appl Physiol. 1981;50:814–818. doi: 10.1152/jappl.1981.50.4.814. [DOI] [PubMed] [Google Scholar]
- 31.Journeay WS, Reardon FD, Martin CR, Kenny GP. Control of cutaneous vascular conductance and sweating during recovery from dynamic exercise in humans. J Appl Physiol. 2004;96:2207–2212. doi: 10.1152/japplphysiol.01201.2003. [DOI] [PubMed] [Google Scholar]
- 32.Kabashima K, Shimauchi T, Kobayashi M, Fukamachi S, Kawakami C, Ogata M, Kabashima R, Mori T, Ota T, Fukushima S, Hara-Chikuma M, Tokura Y. Aberrant aquaporin 5 expression in the sweat gland in aquagenic wrinkling of the palms. J Am Acad Dermatol. 2008;59:S28–32. doi: 10.1016/j.jaad.2008.04.023. [DOI] [PubMed] [Google Scholar]
- 33.Kacin A, Golja P, Eiken O, Tipton MJ, Gorjanc J, Mekjavic IB. Human temperature regulation during cycling with moderate leg ischaemia. Eur J Appl Physiol. 2005;95:213–220. doi: 10.1007/s00421-005-1387-3. [DOI] [PubMed] [Google Scholar]
- 34.Kahn RH. Uber die erwarmung des carotidenblutes. Arch Anat Physiol. 1904:81–134. [Google Scholar]
- 35.Kamijo Y, Lee K, Mack GW. Active cutaneous vasodilation in resting humans during mild heat stress. J Appl Physiol. 2005;98:829–837. doi: 10.1152/japplphysiol.00235.2004. [DOI] [PubMed] [Google Scholar]
- 36.Kamijo Y, Okumoto T, Takeno Y, Okazaki K, Inaki M, Masuki S, Nose H. Transient cutaneous vasodilatation and hypotension after drinking in dehydrated and exercising men. J Physiol. 2005;568:689–698. doi: 10.1113/jphysiol.2005.090530. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Kellogg DL., Jr In vivo mechanisms of cutaneous vasodilation and vasoconstriction in humans during thermoregulatory challenges. J Appl Physiol. 2006;100:1709–1718. doi: 10.1152/japplphysiol.01071.2005. [DOI] [PubMed] [Google Scholar]
- 38.Kellogg DL, Jr, Pergola PE, Piest KL, Kosiba WA, Crandall CG, Grossmann M, Johnson JM. Cutaneous active vasodilation in humans is mediated by cholinergic nerve cotransmission. Circ Res. 1995;77:1222–1228. doi: 10.1161/01.res.77.6.1222. [DOI] [PubMed] [Google Scholar]
- 39.Kimura K, Low DA, Keller DM, Davis SL, Crandall CG. Cutaneous blood flow and sweat rate responses to exogenous administration of acetylcholine and methacholine. J Appl Physiol. 2007;102:1856–1861. doi: 10.1152/japplphysiol.01069.2006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Kolka MA, Stephenson LA. Cutaneous blood flow and local sweating after systemic atropine administration. Pflugers Arch. 1987;410:524–529. doi: 10.1007/BF00586536. [DOI] [PubMed] [Google Scholar]
- 41.Kondo N, Tominaga H, Shibasaki M, Aoki K, Koga S, Nishiyasu T. Modulation of the thermoregulatory sweating response to mild hyperthermia during activation of the muscle metaboreflex in humans. J Physiol. 1999;515:591–598. doi: 10.1111/j.1469-7793.1999.591ac.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Kondo N, Tominaga H, Shiojiri T, Shibasaki M, Aoki K, Takano S, Koga S, Nishiyasu T. Sweating responses to passive and active limb movements. J Therm Biol. 1997;22:351–356. [Google Scholar]
- 43.Krogh A, Lindhard J. The regulation of respiration and circulation during the initial stages of muscular work. J Physiol. 1913;47:112–136. doi: 10.1113/jphysiol.1913.sp001616. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Kumazawa K, Sobue G, Mitsuma T, Ogawa T. Modulatory effects of calcitonin gene-related peptide and substance P on human cholinergic sweat secretion. Clin Auton Res. 1994;4:319–322. doi: 10.1007/BF01821532. [DOI] [PubMed] [Google Scholar]
- 45.Kummer W, Herbst WM, Heym C. Vasoactive intestinal polypeptide receptor-like immunoreactivity in human sweat glands. Neurosci Lett. 1990;110:239–243. doi: 10.1016/0304-3940(90)90853-2. [DOI] [PubMed] [Google Scholar]
- 46.Kuno Y. Human Perspiration. CC Thomas; Springfield, Illinois: 1956. [Google Scholar]
- 47.Latzka WA, Sawka MN, Montain SJ, Skrinar GS, Fielding RA, Matott RP, Pandolf KB. Hyperhydration: thermoregulatory effects during compensable exercise-heat stress. J Appl Physiol. 1997;83:860–866. doi: 10.1152/jappl.1997.83.3.860. [DOI] [PubMed] [Google Scholar]
- 48.Lee K, Mack GW. Role of nitric oxide in methacholine-induced sweating and vasodilation in human skin. J Appl Physiol. 2006;100:1355–1360. doi: 10.1152/japplphysiol.00122.2005. [DOI] [PubMed] [Google Scholar]
- 49.Leonard B, Mitchell JH, Mizuno M, Rube N, Saltin B, Secher NH. Partial neuromuscular blockade and cardiovascular responses to static exercise in man. J Physiol. 1985;359:365–379. doi: 10.1113/jphysiol.1985.sp015590. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Longmore J, Jani B, Bradshaw CM, Szabadi E. Effect of locally administered anticholinesterase agents on the secretory response of human eccrine sweat glands to acetylcholine and carbachol. Br J Clin Pharmac. 1986;21:131–135. doi: 10.1111/j.1365-2125.1986.tb05166.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Low PA. Evaluation of sudomotor function. Clin Neurophysiol. 2004;115:1506–1513. doi: 10.1016/j.clinph.2004.01.023. [DOI] [PubMed] [Google Scholar]
- 52.Low PA. Testing the autonomic nervous system. Semin Neurol. 2003;23:407–421. doi: 10.1055/s-2004-817725. [DOI] [PubMed] [Google Scholar]
- 53.Low PA, Kennedy WR. Cutaneous effectors as indicators of abnormal autonomic function. In: Gibbins IL, Morris JL, editors. The Autonomic Nervous System. Harwood Academic Publisher; Amsterdam: 1997. pp. 165–212. [Google Scholar]
- 54.Lundberg JM, Anggård A, Fahrenkrug J, Hökfelt T, Mutt V. Vasoactive intestinal polypeptide in cholinergic neurons of exocrine glands: functional significance of coexisting transmitters for vasodilation and secretion. Proc Natl Acad Sci USA. 1980;77:1651–1655. doi: 10.1073/pnas.77.3.1651. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Macefield VG, Elam M, Wallin BG. Firing properties of single postganglionic sympathetic neurones recorded in awake human subjects. Auton Neurosci. 2002;95:146–159. doi: 10.1016/s1566-0702(01)00389-7. [DOI] [PubMed] [Google Scholar]
- 56.Macefield VG, Wallin BG. The discharge behaviour of single sympathetic neurones supplying human sweat glands. J Auton Nerv Syst. 1996;61:277–286. doi: 10.1016/s0165-1838(96)00095-1. [DOI] [PubMed] [Google Scholar]
- 57.MacIntyre BA, Bullard RW, Banerjee M, Elizondo R. Mechanism of enhancement of eccrine sweating by localized heating. J Appl Physiol. 1968;25:255–260. doi: 10.1152/jappl.1968.25.3.255. [DOI] [PubMed] [Google Scholar]
- 58.Mack GW, Cordero D, Peters J. Baroreceptor modulation of active cutaneous vasodilation during dynamic exercise in humans. J Appl Physiol. 2001;90:1464–1473. doi: 10.1152/jappl.2001.90.4.1464. [DOI] [PubMed] [Google Scholar]
- 59.McCook RD, Wurster RD, Randall WC. Sudomotor and vasomotor responses to changing environmental temperature. J Appl Physiol. 1965;20:371–378. doi: 10.1152/jappl.1965.20.3.371. [DOI] [PubMed] [Google Scholar]
- 60.Mitchell JH. Neural control of the circulation during exercise. Med Sci Sports Exerc. 1990;22:141–154. [PubMed] [Google Scholar]
- 61.Mitchell JH, Kaufman MP, Iwamoto GA. The exercise pressor reflex: its cardiovascular effects, afferent mechanisms, and central pathways. Annu Rev Physiol. 1983;45:229–242. doi: 10.1146/annurev.ph.45.030183.001305. [DOI] [PubMed] [Google Scholar]
- 62.Montain SJ, Latzka WA, Sawka MN. Control of thermoregulatory sweating is altered by hydration level and exercise intensity. J Appl Physiol. 1995;79:1434–1439. doi: 10.1152/jappl.1995.79.5.1434. [DOI] [PubMed] [Google Scholar]
- 63.Moorhouse VHK. Effect of increased temperature of the carotid blood. Am J Physiol. 1911;28:223–234. [Google Scholar]
- 64.Nadel ER, Bullard RW, Stolwojk JAJ. Importance of skin temperature in the regulation of sweating. J Appl Physiol. 1971;31:80–87. doi: 10.1152/jappl.1971.31.1.80. [DOI] [PubMed] [Google Scholar]
- 65.Nadel ER, Mitchell JW, Saltin B, Stolwojk JAJ. Peripheral modifications to the central drive for sweating. J Appl Physiol. 1971;31:828–833. doi: 10.1152/jappl.1971.31.6.828. [DOI] [PubMed] [Google Scholar]
- 66.Nadel ER, Pandolf KB, Roberts MF, Stolwojk JAJ. Mechanisms of thermal acclimation to exercise and heat. J Appl Physiol. 1974;37:515–520. doi: 10.1152/jappl.1974.37.4.515. [DOI] [PubMed] [Google Scholar]
- 67.Nakamura K, Matsumura K, Hubschle T, Nakamura Y, Hioki H, Fujiyama F, Boldogkoi Z, Konig M, Thiel HJ, Gerstberger R, Kobayashi S, Kaneko T. Identification of sympathetic premotor neurons in medullary raphe regions mediating fever and other thermoregulatory functions. J Neurosci. 2004;24:5370–5380. doi: 10.1523/JNEUROSCI.1219-04.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 68.Nejsum LN, Kwon TH, Jensen UB, Fumagalli O, Frokiaer J, Krane CM, Menon AG, King LS, Agre PC, Nielsen S. Functional requirement of aquaporin-5 in plasma membranes of sweat glands. Proc Nat Acad Sci USA. 2002;99:511–516. doi: 10.1073/pnas.012588099. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 69.Nielsen B, Nielsen M. On the regulation of sweat secretion in exercise. Acta Physiol Scand. 1965;64:314–322. doi: 10.1111/j.1748-1716.1965.tb04185.x. [DOI] [PubMed] [Google Scholar]
- 70.Nielsen S, King LS, Christensen BM, Agre P. Aquaporins in complex tissues. II. Subcellular distribution in respiratory and glandular tissues of rat. Am J Physiol. 1997;273:C1549–1561. doi: 10.1152/ajpcell.1997.273.5.C1549. [DOI] [PubMed] [Google Scholar]
- 71.Ott I. Heat center in the brain. J Nerv Ment Dis. 1877;14:152. [Google Scholar]
- 72.Quigley R, Chu PY, Huang CL. Botulinum toxins inhibit the antidiuretic hormone (ADH)-stimulated increase in rabbit cortical collecting-tubule water Permeability. J Membr Biol. 2005;204:109–116. doi: 10.1007/s00232-005-0754-7. [DOI] [PubMed] [Google Scholar]
- 73.Quinton PM. Physiology of sweat secretion. Kidney Int Suppl. 1987;21:S102–108. [PubMed] [Google Scholar]
- 74.Randall WC, Kimura KK. The pharmacology of sweating. Pharmacol Rev. 1955;7:365–397. [PubMed] [Google Scholar]
- 75.Robertshaw D. Catecholamines and control of sweat glands. In: Blaschko H, Sayers G, Smith AD, editors. Handbook of Physiology: Endocrinology. Bethesda, Maryland: 1975. [Google Scholar]
- 76.Robinson S. Temperature Regulation in Exercise. Pediatrics. 1963;32(SUPPL):691–702. [PubMed] [Google Scholar]
- 77.Saltin B, Gagge AP. Sweating and body temperatures during exercise. Int J Biometeor. 1971;15:189–194. doi: 10.1007/BF01803896. [DOI] [PubMed] [Google Scholar]
- 78.Saltin B, Gagge AP, Stolwijk JAJ. Body temperatures and sweating during thermal transients caused by exercise. J Appl Physiol. 1970;28:318–327. doi: 10.1152/jappl.1970.28.3.318. [DOI] [PubMed] [Google Scholar]
- 79.Sato K. The physiology, pharmacology, and biochemistry of the eccrine sweat gland. Rev Physiol Biochem Pharmacol. 1977;79:51–131. doi: 10.1007/BFb0037089. [DOI] [PubMed] [Google Scholar]
- 80.Sato K, Kang WH, Saga K, Sato KT. Biology of sweat glands and their disorders. I. Normal sweat gland function. J Am Acad Dermatol. 1989;20:537–563. doi: 10.1016/s0190-9622(89)70063-3. [DOI] [PubMed] [Google Scholar]
- 81.Sato K, Sato F. Effect of VIP on sweat secretion and cAMP accmulation in isolated simian eccrine glands. Am J Physiol. 1987;253:R935–R941. doi: 10.1152/ajpregu.1987.253.6.R935. [DOI] [PubMed] [Google Scholar]
- 82.Sawka MN, Gonzalez RR, Young AJ, Muza SR, Pandolf KB, Latzka WA, Dennis RC, Valeri CR. Polycythemia and hydration: effects on thermoregulation and blood volume during exercise-heat stress. Am J Physiol. 1988;255:R456–463. doi: 10.1152/ajpregu.1988.255.3.R456. [DOI] [PubMed] [Google Scholar]
- 83.Sawka MN, Young AJ, Francesconi RP, Muza SR, Pandolf KB. Thermoregulatory and blood responses during exercise at graded hypohydration levels. J Appl Physiol. 1985;59:1394–1401. doi: 10.1152/jappl.1985.59.5.1394. [DOI] [PubMed] [Google Scholar]
- 84.Schlereth T, Dittmar JO, Seewald B, Birklein F. Peripheral amplification of sweating--a role for calcitonin gene-related peptide. J Physiol. 2006;576:823–832. doi: 10.1113/jphysiol.2006.116111. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 85.Shibasaki M, Crandall CG. Effect of local acetylcholinesterase inhibition on sweat rate in humans. J Appl Physiol. 2001;90:757–762. doi: 10.1152/jappl.2001.90.3.757. [DOI] [PubMed] [Google Scholar]
- 86.Shibasaki M, Davis SL, Cui J, Low DA, Keller DM, Crandall CG. Botulinum toxin abolishes sweating via impaired sweat gland responsiveness to exogenous acetylcholine. Br J Dermatol. 2009 doi: 10.1111/j.1365-2133.2009.09248.x. in print. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 87.Shibasaki M, Kondo N, Crandall CG. Evidence for metaboreceptor stimulation of sweating in normothermic and heat-stressed humans. J Physiol. 2001;534:605–611. doi: 10.1111/j.1469-7793.2001.00605.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 88.Shibasaki M, Kondo N, Crandall CG. Non-thermoregulatory modulation of sweating in humans. Exerc Sport Sci Rev. 2003;31:34–39. doi: 10.1097/00003677-200301000-00007. [DOI] [PubMed] [Google Scholar]
- 89.Shibasaki M, Sakai M, Oda M, Crandall CG. Muscle mechanoreceptor modulation of sweat rate during recovery from moderate exercise. J Appl Physiol. 2004;96:2115–2119. doi: 10.1152/japplphysiol.01370.2003. [DOI] [PubMed] [Google Scholar]
- 90.Shibasaki M, Secher NH, Selmer C, Kondo N, Crandall CG. Central command is capable of modulating sweating from non-glabrous human skin. J Physiol. 2003;553:999–1004. doi: 10.1113/jphysiol.2003.051102. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 91.Smiles KA, Elizondo RS, Barney CC. Sweating responses during changes of hypothalamic temperature in the rhesus monkey. J Appl Physiol. 1976;40:653–657. doi: 10.1152/jappl.1976.40.5.653. [DOI] [PubMed] [Google Scholar]
- 92.Solack SD, Brengelmann GL, Freund PR. Sweat rate vs. forearm blood flow during lower body negative pressure. J Appl Physiol. 1985;58:1546–1552. doi: 10.1152/jappl.1985.58.5.1546. [DOI] [PubMed] [Google Scholar]
- 93.Stolwijk JA, Hardy JD. Partitional calorimetric studies of responses of man to thermal transients. J Appl Physiol. 1966;21:967–977. doi: 10.1152/jappl.1966.21.3.967. [DOI] [PubMed] [Google Scholar]
- 94.Sugenoya J, Iwase S, Mano T, Ogawa T. Identification of sudomotor activity in cutaneous sympathetic nerves using sweat expulsion as the effector response. Eur J Appl Physiol. 1990;61:302–308. doi: 10.1007/BF00357617. [DOI] [PubMed] [Google Scholar]
- 95.Sugenoya J, Iwase S, Mano T, Sugiyama Y, Ogawa T, Nishiyama T, Nishimura N, Kimura T. Vasodilator component in sympathetic nerve activity destined for the skin of the dorsal foot of mildly heated humans. J Physiol. 1998;507:603–610. doi: 10.1111/j.1469-7793.1998.603bt.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 96.Tainio H, Vaalasti A, Rechardt L. The distribution of substance P-, CGRP-, galanin- and ANP-like immunoreactive nerves in human sweat glands. Histochem J. 1987;19:375–380. doi: 10.1007/BF01680455. [DOI] [PubMed] [Google Scholar]
- 97.Takamata A, Mack GW, Gillen CM, Jozsi AC, Nadel ER. Osmoregulatory modulation of thermal sweating in humans: reflex effects of drinking. Am J Physiol. 1995;268:R414–R422. doi: 10.1152/ajpregu.1995.268.2.R414. [DOI] [PubMed] [Google Scholar]
- 98.Takamata A, Nagashima K, Nose H, Morimoto T. Osmoregulatory inhibition of thermally induced cutaneous vasodilation in passively heated humans. Am J Physiol. 1997;273:R197–R204. doi: 10.1152/ajpregu.1997.273.1.R197. [DOI] [PubMed] [Google Scholar]
- 99.Uno H. Sympathetic innervation of the sweat glands and piloarrector muscles of macaques and human beings. J Invest Dermatol. 1977;69:112–120. doi: 10.1111/1523-1747.ep12497915. [DOI] [PubMed] [Google Scholar]
- 100.van Beaumont W, Bullard RW. Sweating exercise stimulation during circulatory arrest. Science. 1966;152:1521–1523. doi: 10.1126/science.152.3728.1521. [DOI] [PubMed] [Google Scholar]
- 101.van Beaumont W, Bullard RW. Sweating: its rapid responses to muscular work. Science. 1963;141:643–646. doi: 10.1126/science.141.3581.643. [DOI] [PubMed] [Google Scholar]
- 102.Vissing SF, Hjortso EM. Central motor command activates sympathetic outflow to the cutaneous circulation in humans. J Physiol. 1996;492:931–939. doi: 10.1113/jphysiol.1996.sp021359. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 103.Vissing SF, Scherrer U, Victor RG. Increase of sympathetic discharge to skeletal muscle but not to skin during mild lower body negative pressure in humans. J Physiol. 1994;481:233–241. doi: 10.1113/jphysiol.1994.sp020434. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 104.Wallin BG, Fagius J. Peripheral sympathetic neural activity in conscious humans. Ann Rev Physiol. 1988;50:565–576. doi: 10.1146/annurev.ph.50.030188.003025. [DOI] [PubMed] [Google Scholar]
- 105.Welch G, Foote KM, Hansen C, Mack GW. Nonselective NOS inhibition blunts the sweat response to exercise in a warm environment. J Appl Physiol. 2009;106:796–803. doi: 10.1152/japplphysiol.90809.2008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 106.Wilson TE, Cui J, Crandall CG. Absence of arterial baroreflex modulation of skin sympathetic activity and sweat rate during whole-body heating in humans. J Physiol. 2001;536:615–623. doi: 10.1111/j.1469-7793.2001.0615c.xd. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 107.Wilson TE, Cui J, Crandall CG. Mean body temperature does not modulate eccrine sweat rate during upright tilt. J Appl Physiol. 2005;98:1207–1212. doi: 10.1152/japplphysiol.00648.2004. [DOI] [PubMed] [Google Scholar]
- 108.Wurster RD, McCook RD. Influence of rate of change in skin temperature on sweating. J Appl Physiol. 1969;27:237–240. doi: 10.1152/jappl.1969.27.2.237. [DOI] [PubMed] [Google Scholar]
- 109.Wyndham CH. The physiology of exercise under heat stress. Ann Rev Physiol. 1973;35:193–220. doi: 10.1146/annurev.ph.35.030173.001205. [DOI] [PubMed] [Google Scholar]
- 110.Yamashita Y, Ogawa T, Ohnishi N, Imamura R, Sugenoya J. Local effect of vasoactive intestinal polypeptide on human sweat-gland function. Jpn J Physiol. 1987;37:929–936. doi: 10.2170/jjphysiol.37.929. [DOI] [PubMed] [Google Scholar]
- 111.Yamazaki F, Fujii N, Sone R, Ikegami H. Responses of sweating and body temperature to sinusoidal exercise in physically trained men. J Appl Physiol. 1996;80:491–495. doi: 10.1152/jappl.1996.80.2.491. [DOI] [PubMed] [Google Scholar]
- 112.Yamazaki F, Sone R, Ikegami H. Responses of sweating and body temperature to sinusoidal exercise. J Appl Physiol. 1994;76:2541–2545. doi: 10.1152/jappl.1994.76.6.2541. [DOI] [PubMed] [Google Scholar]