

SUMMARY
Previous serological studies on the Arara do Laranjal Indian group revealed extensive HTLV-2 infections. A collection of 97 new samples from the Arara were found repeatedly negative using three different commercial enzyme immunoassays. Eight samples that exhibited optical density readings close to the cut-off value were re-evaluated using Western blot (GeneLab 2.4, Singapore) assay. One sample was found to be non-reactive, five exhibited indeterminate patterns, one was classified as HTLV, and one was confirmed as HTLV-2. Peripheral blood mononuclear cell DNA of the eight samples were subjected to nested PCR and restriction fragment length polymorphism (RFLP) analysis of the pX and env regions, and nucleotide sequencing of the 5′-LTR region. All produced amplification products of pX, but env could be amplified in only one sample with the commonly used primers. RFLP analysis of the pX region using TaqI confirmed HTLV-2 infection. Nucleotide sequencing of the 5′-LTR region was performed in three samples (HTLV-2, HTLV and indeterminate based on Western blot pattern). Phylogenetic analysis of a 449-nt fragment using the Neighbour-Joining method clearly demonstrated that the three samples clustered within the HTLV-2c molecular subtype. The present study confirms the wide dissemination of the HTLV-2c subtype among linguistically and culturally distinct Amazonian Indian groups, and emphasizes the unique occurrence of infection by this subtype in Brazil. Moreover, it emphasizes the limitation of employing the present serological screening assays in blood banks, epidemiological studies, and the importance of molecular assays in the confirmatory procedures for the primary detection of HTLV-2 infections.
INTRODUCTION
Human T-cell lymphotropic virus 1 and 2 (HTLV-1 and HTLV-2) are members of the family Retroviridae and share several molecular and biological properties [1]. Molecular characterization of HTLV-2 strains originated from endemic regions shows four molecular subtypes, named HTLV-2a, HTLV-2b, HTLV-2c and HTLV-2d, with a nucleotide divergence ranging from 4% to 6%, according to the genomic region investigated [2–5]. Restriction maps from the LTR region shows the occurrence of at least five haplotypes of subtype 2a and six of subtype 2b [2, 6], but there is no evidence so far, of distinct biological properties or differences in their pathogenesis.
HTLV-2a, HTLV-2b and HTLV-2c are largely distributed among native Indian population groups in the Americas. In North America, 2a and 2b are present among the Navajo and Pueblo from New Mexico [7] and Seminole, in Florida [6, 8]. HTLV-2b is endemic among the Guaymi in Panama [9–11] and is present in Mayan descendants from Mexico [12]. In South America, HTLV-2b shows a widespread distribution among the Guahibo and Wayu, Colombia [13–15], the Toba and Mataco, Argentina [16] and in small groups from Chile. Subtype HTLV-2c is a unique molecular subtype occurring among several native Indians from the Amazon Region of Brazil as well as in urban areas [2, 4, 17–20].
The present work expands the molecular characterization of HTLV-2 strains originated from previously visited Arara do Laranjal Indian group of the Brazilian Amazon Region, providing further evidences for the molecular epidemiology of HTLV-2c in the region, and the limitation of sensitivity of commercial serological tests presently available to detect antibodies to HTLV.
MATERIALS AND METHODS
Population examined and samples
Blood samples were collected from 97 subjects residing in the Arara do Laranjal Indian tribe (a Karib-speaking linguistic group) from the Amazon Region of Brazil (Fig. 1). All the subjects had a sample of blood drawn and placed in tubes without anticoagulant to obtain serum and in tubes containing Hespan (DuPont, Wilmington, DE, USA) in order to separate peripheral blood mononuclear cells (PBMC). All samples were stored at −20°C before use.
Fig. 1.

Geographical location of the Arara do Laranjal Indian tribe in the State of Pará (PA).
Serological assays
Serum samples were assayed for the presence of antibodies to HTLV-1/2 using three different enzyme immunoassays – EIA (Ortho Diagnostic, Raritan, NJ, USA), which uses four recombinant antigens from core and envelope of HTLV-1 and -2; Murex (Dartford, UK), which employs microwells coated with synthetic peptides representing immunodominant antigens from HTLV-1 and -2 envelope proteins and a recombinant transmembrane protein from HTLV-2; and a third one from Cambridge Biotech Corporation (Worcester, MA, USA), which employs a combination of viral lysate and a recombinant source for HTLV-1 as antigen. The positive samples were tested by a Western blot (Genelab 2.4, Singapore) that permits confirmation of HTLV-1 and HTLV-2 seroreactivity. The discriminatory criteria of the Western blot followed the manufacturer's recommendations for reactivity to p19, p24 and to the synthetic peptides added to the kit, that specifically react with HTLV-1 (MTA, rgp46-I) and HTLV-2 (K55, rgp46-II).
Polymerase chain reaction (PCR) and restriction fragment length polymorphism (RFLP) analysis
DNA extraction was performed on PBMC from HTLV-2 seropositive samples. The extracted product was used for the detection of the provirus genome using a nested PCR to amplify the pX region. The external primers sequences were 5′-TTCCCAGGGTTTGGACAGAG-3′ (nucleotides 7219–7238) and 5′-GGGTAAGGACCTTGAGGGTC-3′ (nucleotides 7483–7464). The internal primer sequences were 5′-CGGATACCCAGTCTACGTGTT-3′ (nucleotides 7248–7268) and 5′-GAGCCGATAACGCGTCCATCG-3′ (nucleotides 7406–7386). The two amplified products were detected after electrophoresis in 2% agarose gel (200 V, 120 A/45 min), revealed by staining with ethidium bromide and visualized under UV light.
RFLP typing of this product (159 bp for both HTLV-1 and HTLV-2) was performed by mixing it (10 μl) with 0·4 μl of restriction endonuclease enzyme TaqI and incubating at 65°C for at least 5 h. The restriction site for the enzyme (T/CGA) is present in HTLV-2, generating two bands (85 bp and 53 bp) that are visualized following electrophoresis in 3% agarose gel. HTLV-1 lacks this particular site and runs as a single band [21].
The subtyping of HTLV-2 was attempted using a nested PCR, with two sets of primers that match a conservative nucleotide sequence flanking the env region, and coding for the envelope protein, as previously described [3].
All samples characterized as HTLV-2 were submitted to a nested PCR to the 5′-LTR region, with two sets of primers, 5′-CCTTACCCACTTCCCCTAGCACTGA′ and 5′-GGGAAAGCCCGTGGATTTGCCCCAT-3′, corresponding to nucleotides 194–218 and 807–831 to the first step and 5′-AAAAGCGCAAGGACAGTTCAGGAGG-3′ and 5′-ATCCCGGACGAGCCCCCACTTGTTT-3′ in a second step, corresponding to nucleotides 290–314 and 759–783 from HTLV-2Mo strain. Following the amplification, the 5′-LTR fragment was electrophoresed on 1·5% agarose gel and purified by QIA Quick Purification kit (Qiagen Inc., Valencia, CA, USA) prior to direct sequencing of the product.
DNA sequencing
The amplified fragments were submitted to a direct sequencing assay according to a protocol described on the ABI Prism Dye Terminator Cycle Sequencing Ready kit (PerkinElmer Cetus, Norwalk, CA, USA), and the reaction products were loaded on the ABI Prism 373 DNA Sequencer (PerkinElmer).
Phylogenetic analysis
The nucleotide sequences originated from the 5′-LTR region (449 nt), obtained in the present study (Genbank accession nos: Ara8813, AY334275; Ara8845, AY334276; Ara8855, AY334277), were used to establish the phylogenetic relationship together with the 31 following HTLV-2 strains described in the Genbank: MO, M10060; NRA, L20734; G12, L11456; PYGCAM-1, Z46888; PH230PCAM, Z46838; ATL18, U10252; BRAZ.A21, U10253; LA8A, U10256; NAV.DS, U10257; PUEG.AG, U10261; PUEB.RB, U10262; ITA50A, U10255; NY185, U10259; PENN7A, U10260; SEM1051, U10264; SPAN129, U10266; WYU1, U12792; GHKT, L42507; KAY73, L42509; KAY139, L42508; RJ01, U32886; SP01, U32888; SP02, U32889; SP03, U328890; SP04, U32891; SP05, U32892; SP06, U32893; SP07, U32894; Mexy17, L42510; DP, L77237; Pygmy-2, Y14365 and HTLV-2-GAB, Y13051. The sequence alignments were performed using the Eyeball Sequence Editor [22]. The phylogenetic relationship among the samples was performed by the Phylogenetic Inference Package – phylip 3.5 v [23]. A phylogenetic tree was infered by the Neighbour-Joining (NJ) method, using the HKY substitution model. The statistical reliability of the NJ tree was evaluated using 2000 bootstrap replicates.
RESULTS
Serological analysis
All the Arara do Laranjal subjects were repeatedly negative using three different commercial enzyme immunoassays. Eight samples that showed optical density (OD) closest to the cut-off value were selected for further analysis using Western blot assay, and one was found to be negative, five exhibited an indeterminate pattern, one was classified as HTLV and one was confirmed as HTLV-2. OD values and Western blot results are listed in the Table.
Table.
Serological and molecular evidence of HTLV-2 infection in the Arara do Laranjal village from the Amazon region of Brazil
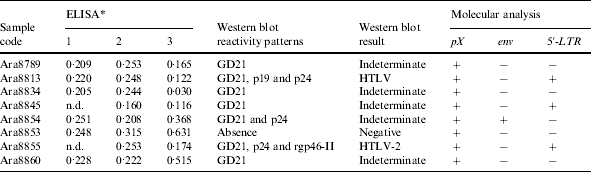
n.d., Not done.
ELISA: 1, Murex (OD/cut-off); 2, Ortho Diagnostic (OD/cut-off); 3, Cambridge (OD/cut-off).
Molecular characterization and phylogenetic analysis
The amplification of the pX genomic region (Fig. 2), followed by enzymatic digestion using TaqI endonuclease confirmed the HTLV-2 infection among the eight Amerindian samples, producing a restriction pattern with the two bands (85 bp and 53 bp). However, the amplification of the env and subsequent restriction analysis with XhoI was possible in only one of the samples (Table).
Fig. 2.

The amplification of the pX genomic region detected after electrophoresis in 3% agarose gel (200 V, 120 A/45 min), revealed by staining with ethidium bromide and visualized under UV light. CN1 and C2, negative control; CP1, positive control; ladder 100 bp.
The direct confirmation was performed by nucleotide sequence analysis of 449 bp amplified from the 5′-LTR segment of three samples followed by phylogenetic analysis. This method confirmed that the three samples clustered within the HTLV-2c molecular subtype found in other native Indian and urban populations from Brazil (Fig. 3).
Fig. 3.

Unrooted phylogenetic tree showing the evolutionary relationship of HTLV-2 reported so far including the three newly sequenced isolates from the Arara do Laranjal village. The tree was constructed by NJ method after alignment of 449 nt of the 5′-LTR region. The statistical support was applied using 2000 bootstrap replicates.
DISCUSSION
The present study clearly confirms both the wide dissemination of the HTLV-2c as a unique molecular subtype throughout the Amazon Region of Brazil and its presence in different linguistic and culturally distinct Amazonian Indian groups. So far, two different groups (Karib and Jê) have been shown to be infected with HTLV-2c [4, 18, 20].
The occurrence of HTLV-2c in the North and several other areas of Brazil is intriguing and raises some possible explanations for its origin. The two different migratory routes paralleling the Andean Cordillera [11] along the Pacific Coast and towards the Amazon Region could have independently resulted in the emergence of the HTLV-2c strain (perhaps from a prototype 2a) or in the introduction of 2c exclusively into the Amazon Region [20].
Within the Indian communities, sexual and vertical transmission play an important role in the maintenance of the virus because of the apparent absence of intravenous drug use or blood transfusions [4, 18, 24].
The present study highlights the limitations of employing the present commercial serological assays both for diagnostic and epidemiological purposes in different geographical areas of the country. The Amazon Region of Brazil comprises an area of more than 3 million km2 and HTLV-2c has been further described in the Southern area of the country [2, 25]. It is an extensive epidemiological area for the occurrence of HTLV-2c which requires proper diagnosis.
The initial serological results (non-reactive) obtained were unusual and unexpected. Three of the subjects tested (Ara8789, Ara8834 and Ara8854, formerly registered as Ara7655, Ara7657 and Ara7676, respectively) yielded seroreactive samples to HTLV-2 from a previous visit to the group [4]. One case of serological reversion has been described in the Xicrin, a Kayapo village [26]. This result raises some possible explanations: (i) the long persistence of the virus in latency associated to low viral load could lead to a gradually diminished concentration of specific antibodies, consequently reducing the sensitivity of the present serological assays, or (ii) the commercial enzyme immunoassays presently available are not fully prepared to detect antibodies to HTLV-2c and particularly to what seems to be a novel variant strain within the Arara do Laranjal. Few commercial and technological ventures have been implemented towards the improvement of serological assays for the detection of antibodies, despite the amount of financial resources which are spent annually to test more than two million samples for the presence of antibodies to HTLV.
Presently, it is highly recommended to use molecular methods to confirm diagnostic and epidemiological results, which are not conclusive. Confirmatory test such as indirect immunofluorescence or Western blot are either not available or too expensive to adapt for routine use [27]. Furthermore, the same problems of sensitivity and specificity detected for the screening assays are expected to occur as commercial companies are not releasing new products for the testing of human samples.
ACKNOWLEDGEMENTS
We thank the Arara do Laranjal Indians and send them our best wishes. We also thank V. Monteiro and E. S. Abrahim for their help in the collection of the samples. This work was partially supported by grants from the CNPq, CAPES, SECTAM and Universidade Federal do Para.
DECLARATION OF INTEREST
None.
REFERENCES
- 1.Hall WW et al. Human T-cell leukemia virus type II (HTLV-2): emergence of an important newly recognized pathogen. Seminars in Virology. 1994;5:165–178. [Google Scholar]
- 2.Eiraku N et al. Identification and characterization of a new and distinct molecular subtype of human T-cell lymphotropic virus type 2. Journal of Virology. 1996;70:1481–1492. doi: 10.1128/jvi.70.3.1481-1492.1996. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Hall WW et al. Multiple isolates and characteristics of human T-cell leukemia virus type II. Journal of Virology. 1992;66:2456–2463. doi: 10.1128/jvi.66.4.2456-2463.1992. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Ishak R et al. Identification of human T-cell lymphotropic virus type IIa infection in the Kayapo, an indigenous population of Brazil. AIDS Research and Human Retroviruses. 1995;11:813–821. doi: 10.1089/aid.1995.11.813. [DOI] [PubMed] [Google Scholar]
- 5.Vandamme A-M et al. African origin of Human T-Lymphotropic Virus Type 2 (HTLV-2) supported by a potential new HTLV-2d subtype in Congolese Bambuti Efe Pygmies. Journal of Virology. 1998;72:4327–4340. doi: 10.1128/jvi.72.5.4327-4340.1998. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Switzer WM et al. Phylogenetic relationship and geographic distribution of multiple human T-cell lymphotropic virus type II subtypes. Journal of Virology. 1995;69:621–632. doi: 10.1128/jvi.69.2.621-632.1995. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Hjelle B et al. Endemic human T cell leukemia virus type II infection in Southwestern US Indians involves two prototypes variants of virus. Journal of Infectious Diseases. 1993;168:737–740. doi: 10.1093/infdis/168.3.737. [DOI] [PubMed] [Google Scholar]
- 8.Levine PH et al. HTLV-2 infection in Florida Indians. AIDS Research and Human Retroviruses. 1993;9:123–127. doi: 10.1089/aid.1993.9.123. [DOI] [PubMed] [Google Scholar]
- 9.Heneine W et al. HTLV-II endemicity among Guaymi Indians in Panama. New England Journal of Medicine. 1991;324:565. doi: 10.1056/NEJM199102213240815. [DOI] [PubMed] [Google Scholar]
- 10.Pardi D et al. Complete nucleotide sequence of an Amerindian human T-cell lymphotropic virus type II (HTLV-II) isolate: identification of a variant HTLV-IIb subtype from a Guaymi Indian. Journal of Virology. 1993;67:4659–4664. doi: 10.1128/jvi.67.8.4659-4664.1993. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Rothhamer F, Silva C. Peopling of Andean South America. American Journal of Physical Anthropology. 1989;78:403–410. doi: 10.1002/ajpa.1330780308. [DOI] [PubMed] [Google Scholar]
- 12.Góngora-Biachi RAet al. VIII International Conference on Human Retrovirology: HTLV1997Rio de Janeiro, Brasil [Google Scholar]
- 13.Duenas-Barajas E V International Conference on Human Retrovirology, HTLV. Kumamoto, Japan: 1992. Co-existence of HTLV-I and HTLV-II infection among the Wayu Indians of Colombia. [Google Scholar]
- 14.Switzer WM et al. Molecular analysis of human T-cell lymphotropic virus subtype II from Wayu indians of Colombia demonstrates two subtypes of HTLV-IIb. Virus Gene. 1995;10:153–162. doi: 10.1007/BF01702596. [DOI] [PubMed] [Google Scholar]
- 15.Zaninovic V, Tajima K, Kajiwara M.Epidemiology of HTLV-I and II in Amerindians from Andes, Amazon, and Caribbean coast of Colombia (South America)V International Conference on Human Retrovirology: HTLV 1992Kumamoto, Japan [Google Scholar]
- 16.Biglione M et al. The G. Endemic HTLV-II infection among Tobas and Matacos Amerindians from North Argentina. AIDS. 1993;6:631–633. [PubMed] [Google Scholar]
- 17.Ishak R et al. Detection of HTLV-IIa in blood donors in an urban area of the Amazon Region of Brazil (Belem, Para) Revista da Sociedade Brasileira de Medicina Tropical. 1998;31:193–197. doi: 10.1590/s0037-86821998000200005. [DOI] [PubMed] [Google Scholar]
- 18.Ishak R et al. Molecular evidence of mother-to-child transmission of HTLV-IIc in the Kararaô Village (Kayapo) in the Amazon region of Brazil. Revista da Sociedade Brasileira de Medicina Tropical. 2001;34:519–525. doi: 10.1590/s0037-86822001000600004. [DOI] [PubMed] [Google Scholar]
- 19.Vallinoto ACR et al. Serological evidence of HTLV-I and HTLV-II co-infections in HIV-1 positive patients in Belem, State of Para, Brazil. Memórias do Instituto Oswaldo Cruz. 1998;93:407–409. doi: 10.1590/s0074-02761998000300026. [DOI] [PubMed] [Google Scholar]
- 20.Vallinoto ACR et al. Molecular Epidemiology of Human T-Lymphotropic Vírus Type II Infection in Amerindian and Urban Populations of the Amazon region of Brazil. Human Biology. 2002;74:633–644. doi: 10.1353/hub.2002.0059. [DOI] [PubMed] [Google Scholar]
- 21.Tuke PW, Luton P, Garson JA. Differential diagnosis of HTLV-I and HTLV-II infections by restriction enzyme analysis of nested PCR products. Journal of Virological Methods. 1992;40:163–174. doi: 10.1016/0166-0934(92)90065-l. [DOI] [PubMed] [Google Scholar]
- 22.Cabot EL, Beckenbach AT. Simultaneous editing of multiplic nucleic acid and protein sequences with ESEE. Computer Applications in the Biosciences. 1989;5:233–234. doi: 10.1093/bioinformatics/5.3.233. [DOI] [PubMed] [Google Scholar]
- 23.Felsenstein J. PHYLIP – Phylogeny Inference Package. Seattle: 1993. [Google Scholar]
- 24.Hjelle B, Cyrus S, Swenson SG. Evidence for sexual transmission of human T lymphotropic virus type II. Annals of Internal Medicine. 1992;116:90–91. doi: 10.7326/0003-4819-116-1-90. [DOI] [PubMed] [Google Scholar]
- 25.Vicente ACPet al. Human T-Cell Lymphotropic Virus Type II in Brazil: Epidemiological Aspects and EvolutionVII International Symposium on HTLV in Brasil 2002Belém, Pará, Brasil [Google Scholar]
- 26.Black FL et al. Twenty-five years of HTLV type II follow-up with a possible case of tropical spastic paraparesis in the Kayapo, a Brazilian Indian Tribe. AIDS Research and Human Retroviruses. 1996;12:1623–1627. doi: 10.1089/aid.1996.12.1623. [DOI] [PubMed] [Google Scholar]
- 27.Ministério da Saúde. Brasilia: 2004. [Google Scholar]