

SUMMARY
The weekly number of dengue cases in Peru, South America, stratified by province for the period 1994–2006 were analysed in conjunction with associated demographic, geographic and climatological data. Estimates of the reproduction number, moderately correlated with population size (Spearman ρ=0·28, P=0·03), had a median of 1·76 (IQR 0·83–4·46). The distributions of dengue attack rates and epidemic durations follow power-law (Pareto) distributions (coefficient of determination >85%, P<0·004). Spatial heterogeneity of attack rates was highest in coastal areas followed by mountain and jungle areas. Our findings suggest a hierarchy of transmission events during the large 2000–2001 epidemic from large to small population areas when serotypes DEN-3 and DEN-4 were first identified (Spearman ρ=−0·43, P=0·03). The need for spatial and temporal dengue epidemic data with a high degree of resolution not only increases our understanding of the dynamics of dengue but will also generate new hypotheses and provide a platform for testing innovative control policies.
INTRODUCTION
Dengue fever is a mosquito-borne disease that affects between 50 and 100 million people each year [1]. The disease is transmitted primarily via mosquitoes of the species Aedes aegypti and Aedes albopictus, carriers of the virus serotypes (DEN-1, DEN-2, DEN-3, and DEN-4), of the genus Flavivirus [1]. The severity of the disease ranges from asymptomatic, clinically non-specific with flu-like symptoms, dengue fever, dengue haemorrhagic fever, and dengue shock syndrome [1]. Dengue attack rates are around 40–50% but may be as high as 80–90% [2]. Efforts to eradicate A. aegypti in the Americas began in the 1950s with some success albeit sporadic outbreaks took place. The cancellation of mosquito eradication programmes in the 1970s throughout the Americas facilitated dengue re-emergence in various regions of Central and South America with A. aegypti as the primary vector [3]. The situation of dengue in Latin American countries has evolved from non-endemic (no virus present), to hypo-endemic (one virus serotype present) to hyper-endemic (multiple virus serotypes co-circulating) [4]. The factors associated with these co-evolutionary outcomes include climatological and environmental changes as well as the increased migration and travel of humans [5].
In Peru, dengue serotype DEN-1 was first identified in 1990 in the department of Iquitos in the Amazon region [6]. The establishment of DEN-1 was followed by the successful invasion of American genotype DEN-2, the driver of the large dengue epidemic of 1995–1996 [6]. The dengue epidemic of 2000–2001 in Peru was the first in which all four dengue serotypes co-circulated (Asian DEN-2, DEN-3, and DEN-4 were identified for the first time) [7]. The new serotypes probably travelled from Ecuador where they had been isolated 6 months before the Peruvian outbreak [7]. In this paper, we carry out spatial–temporal analyses based on Peru (1994–2006) weekly dengue incidence data stratified by province that also includes demographic, geographic and climate data. In this article, we estimate transmissibility of dengue outbreaks using the local reproduction numbers; assess the outbreak dependence on community size as a function of geographic region; characterize the underlying distributions of dengue attack rates and epidemic duration across Peru; quantify the spatial heterogeneity of dengue attack rates using the Lorenz curve and corresponding Gini index; assess the correlation between dengue incidence and climatological variables using regression analysis; and evaluate the level of association between local epidemic timing and demographics.
MATERIALS AND METHODS
Peru, a South American country, is located on the Pacific coast between the latitudes: −3° S to −18° S. Peru shares borders with Bolivia, Brazil, Chile, Colombia, and Ecuador (Fig. 1). The total population of Peru is about 29 million, and it is heterogeneously distributed in a surface area of 1 285 220 km2. Peru's geographic composition includes a western coastal plain, the Andes mountains in the centre, and the eastern jungle of the Amazon (Fig. 1). Peru's weather varies from dry by its coast, tropical by the Amazon, and temperate and frigid in the Andes mountain range. Peru is divided into 25 administrative regions composed of 195 provinces [8]. Spatial dengue epidemic, demographic, geographic, and climate data were gathered from multiple sources in our analyses.
Fig. 1.

Map of Peru highlighting boundaries of 195 provinces and 25 regions. The geography of Peru covers a range of features, from a western coastal plain (yellow), the Andes Mountains in the centre (brown), and the eastern jungle of the Amazon (green). The total population of Peru is about 29 million heterogeneously distributed in an area of 1 285 220 km2.
Dengue epidemic data
The Department of Epidemiology of Peru's Health Ministry is in charge of epidemiological surveillance, which is carried out from a network of over 6000 geographically distributed notifying units. Peru's epidemiological surveillance system has collected weekly disease data reports since 1994 [9]. The case definitions for probable and confirmed dengue cases in use are those from the World Health Organization (WHO) guidelines [10]. The weekly number of probable and confirmed cases at symptoms onset, as recorded by the Health Ministry's General Office of Epidemiology, are stratified by province, and we use these data from 1994 to 2006. A dengue case is classified as probable whenever fever or chills were present in addition to at least two symptoms among myalgia, arthralgia, retro-orbtial pain, headache, rash, or some haemorrhagic manifestation (e.g. petechiae, haematuria, haematemesis, melena). Seventy-three provinces reported dengue cases some time during the period of interest (1994–2006). Eighteen percent of the probable dengue cases were confirmed via virus isolation or anti-dengue IgM antibody tests by the regional laboratories under the supervision of Peru's Health Ministry. For the purpose of our analyses, we define a dengue outbreak as the occurrence of five or more recorded dengue cases within three or more consecutive weeks. Furthermore, cases considered had to be recorded in a window of time bounded above and below by the absence of dengue cases for at least two consecutive weeks. These definitions and assumptions were put into place to limit the confusion that results from imported cases (e.g. cases generated by humans visiting other provinces). We identified 315 dengue outbreaks in 1994–2006 as defined by this conservative definition. Furthermore, the total number of outbreaks identified turned out to be insensitive to the size of the threshold outbreak size. In other words, our results hardly varied when either five, six, or seven reported cases were used as the definition of outbreak. For each dengue outbreak we computed the final epidemic size, the total number of dengue cases occurring during the outbreak, the dengue attack rate as the ratio of the final epidemic size to the population size of the province where the outbreak took place, and the epidemic duration (as the number of weeks of the outbreak).
Population, geographic, and climate data
The population size of the Peruvian provinces during the years 1994–2006 was obtained from the National Institute of Statistics and Informatics of Peru [11] (Fig. S1, available in the online version of the paper). The population density of each province (people/km2) is estimated by dividing the province population size by the surface area (km2) [12]. These averages ranged from a mean of 22·3 people/km2 in the mountain range, 12·38 in the jungle areas, and 172 in the coastal areas (Fig. S2, online).
Each province is classified as a coastal area, mountain range, or jungle area (Fig. 1). The provinces are also differentiated by their latitude, longitude, and elevation (in metres) [13]. The mean elevation of the provinces ranged from 207·38 m in the coastal areas, to 454·96 m in the jungle areas, to 3112·5 m in the mountain range (Fig. S3, online) [13].
Weekly climate data are available for most of the departments comprising Peru. We obtained the weekly mean, minimum, and maximum temperature (Fahrenheit) and precipitation (inches) from meteorological stations located in 15 out of the 18 departments in Peru that reported a dengue outbreak, some time during the period 1994–2006 [14]. Climatological records from Peru suffer from underreporting problems. Hence, we were able to analyse the potential correlation between an outbreak and climatological variables only when sufficient data was available.
Estimation of transmissibility
The basic reproduction number (R0) gives the average number of secondary cases generated by a primary infectious case through the vectors in an entirely susceptible host population [15]. Recurrent infectious diseases alter the susceptibility structure of a population (herd immunity [16]) by letting p denote the fraction of the population that is effectively protected from infection due to prior exposures to the infectious agent. Hence, the reproduction number (R) is a function of R0 and p. The reproduction number is modelled as R=(1 – p)R0, a reasonable model when the population is well mixed and in situations where herd immunity data are not available.
We estimate the transmissibility (R) of the dengue outbreaks at the level of provinces in Peru during the period 1994–2006 using a previously published approach [16]. As in our earlier work, we use the mean and variance of the distributions for the incubation period in the human host (mean=1/kh, variance=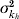 ) and vector (
) and vector (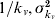 ) and the mean and variance of the host's infectious period (
) and the mean and variance of the host's infectious period (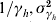 ). The distributions are approximated through estimates of the number of progressive stages needed to fit a linear-chain model [17]. The number of compartments necessary to model the incubation periods are given by
). The distributions are approximated through estimates of the number of progressive stages needed to fit a linear-chain model [17]. The number of compartments necessary to model the incubation periods are given by 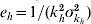 and
and 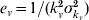 , and
, and 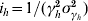 for the infectious period in humans. For example, the number of compartments necessary to model the dengue infectious period in humans turned out to be 25 if one assumes a mean infectious period of 5 days with a standard deviation of 1 day [16].
for the infectious period in humans. For example, the number of compartments necessary to model the dengue infectious period in humans turned out to be 25 if one assumes a mean infectious period of 5 days with a standard deviation of 1 day [16].
The formula of the reproduction number used to obtain our estimates is given by [16]:
where C denotes the mean rate of mosquito bites per mosquito; βhv is a constant transmission probability per bite from an infectious mosquito to a human; βvh is a constant transmission probability per bite from an infectious human to a susceptible mosquito; m is the mean ratio of female mosquitoes per host; and μv is the mean mosquito mortality rate. R0 is therefore the product of the number of infectious mosquitoes generated during the infectious period of a primary infectious human (mCβvh)/γh and
the number of infectious humans generated by the proportion of infectious mosquitoes surviving the extrinsic incubation period.
There are few estimates of mosquito infestation in Peru (e.g. densities of pupae and positive containers) [18]. We were not able to obtain reliable estimates of the number of female mosquitoes per person (parameter m). On the other hand, a study in Thailand reported an average of 20 female mosquitoes per room [19] while a similar study in Puerto Rico found 5–10 female mosquitoes per house [20]. Here, we set the average number of female mosquitoes per person to 4 and perform a sensitivity analysis on the impact of this parameter (see [16]). Estimates of the reproduction number for dengue have been found to be insensitive to this parameter, from 3 to even 5 had no impact on R [e.g. pairwise correlation coefficient (>0·90); analysis of variance (P=0·94)].
The mean adult mosquito lifespan is taken as 10·5 days (95% CI 6–15) [21]; the mean intrinsic incubation period is taken to be 5·5 days (95% CI 4–7) [22]; and the mean infectious period is assumed to be 5 days (95% CI 3–7) [22]. Infected mosquitoes must survive longer than their extrinsic incubation period to be able to infect another human [23]. A typical infected mosquito never becomes infectious. Further, the extrinsic incubation period is sensitive to ambient temperature. We use the thermodynamic relation from Focks et al. [24] to model the temperature dependence of the extrinsic incubation time. As the temperature increases, the incubation period decreases while the mosquito lifespan is relatively unchanged (Fig. S4, online). The proportion of mosquitoes that survive the extrinsic incubation period is given by
For parameter values of 1/kv=5 days, ev=25 (for a mean ambient temperature of 35°C and 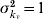 ) and μv=1/10·5 days, the fraction of mosquitoes surviving the extrinsic incubation period is 62·4%.
) and μv=1/10·5 days, the fraction of mosquitoes surviving the extrinsic incubation period is 62·4%.
We estimate the transmission rate from mosquitoes to humans (Cβhv), the transmission rate from humans to mosquitoes (Cβvh), the initial number of infectious hosts, and the initial number of infectious vectors through our least squares fitting of the initial phase of the epidemic curve of dengue cases using as a marker the onset of symptoms (see [16]). We are only able to estimate the reproduction number for outbreaks whose initial phase was comprised of at least five epidemic weeks of data as four parameters had to be estimated. The best parameter estimates are obtained from model fits bounded when the χ2 goodness-of-fit statistic reaches a minimum [16].
Critical community size
Several studies have addressed the problem of persistence of measles as a function of community size in island and non-island populations (see [15]). It is therefore of interest to provide rough estimates of a ‘critical’ community size by geographic region above which dengue epidemics would typically take off. The determination of the critical community size, above which infectious diseases persist, is of central importance [15, 25]. Determining the effective or critical community size for a particular invasion is a complex matter because of variations in herd immunity, immigration rates, the possibility of disease re-introductions in the population, and the nature of human interactions. Following the approach of Wearing & Rohani [26], we assess whether or not the critical community size for the case of dengue in Peru could be estimated from the proportion of weeks with no dengue reports for each of the provinces in the time series (1994–2006) is a good indicator.
Scaling laws in the distributions of attack rates and duration of epidemics
Here we estimate the type of distributions underlying epidemic attack rates and epidemic durations at a particular spatial resolution (e.g. province) over a long window of time. We use these estimates to characterize dengue attack rates and epidemic duration across Peru's provinces during 1994–2006. We observed that if Y denotes the dengue epidemic outbreak duration then Y does not have a typical distribution (like a normal) but rather a power-law distribution (known as a Pareto distribution in statistics) of the form Y−b where b is a positive constant. Similarly, when X denotes the dengue attack rate over this long window in time it turns out to also follow a power-law distribution. That is, the data follows a straight line when the points are plotted in a double-logarithmic diagram. Final epidemic size and duration distributions have been estimated from multi-annual measles epidemic data generated from outbreaks in island populations [27]. They have also shown to be best described by power laws [27].
Spatial heterogeneity
Spatial variations in attack rates have not been extensively studied. Here, using the Lorenz curve and associated summary Gini index, an approach derived from econometrics, is used to quantify spatial heterogeneity of infectious diseases (see [28–31]). The Lorenz curve is a graphical representation of the cumulative distribution function of a probability distribution, representing in our case the proportion of cases associated with the bottom y% of a population. Equal attack rates (no heterogeneity) result in a first diagonal Lorenz curve. On the other hand, perfectly unbalanced distributions give rise to a vertical Lorenz line (maximum heterogeneity). Most empirical attack rate distributions lie somewhere in between.
The Gini index summarizes the statistics of the Lorenz curve (ranging between 0 and 1). It is calculated as the area between the Lorenz curve and the diagonal representing no heterogeneity. A large Gini index indicates high heterogeneous attack rates, i.e. a situation where the highest attack rates are concentrated in a small proportion of the population. A Gini index of zero indicates that attack rates are directly proportional to population size (no heterogeneity).
Time-series analysis with climatological variables
Different aspects of the transmission dynamics of dengue depend on climatological conditions including the survival, development, and maturation of the vector A. aegypti [32–34]. Moreover, the extrinsic (mosquito) incubation period and susceptibility of the mosquito depends on temperature [34]. We carry out lagged cross-correlation analyses to assess the role of climatological variables on dengue incidence lagged effects. The time it takes for mosquito larva to develop to adult stages; the time it takes infected mosquitoes to become infectious; and the time from infection in host to developing clinical symptoms are critical to the transmission process [35, 36].
Timing of dengue epidemics and demographics
We evaluate the degree of association between the timing of epidemic onset with demographic and geographic variables. Epidemic onset is defined here as the first week with dengue reports for a given epidemic period.
RESULTS
The aggregated dengue epidemic curve in Peru seems to support extremely high attack rates in 1996 when the serotype DEN-2 first appeared in Peru, and in 2001 when serotypes DEN-3 and DEN-4 were first identified (Fig. S5, online). Since 2001, recurrent annual dengue outbreaks have been reported in Peru (Fig. 2).
Fig. 2.

(a) Weekly dengue incidence per 100 000 people in each of the 73 provinces [8] reporting dengue in Peru during the period 1994–2006. For visualization purposes, we took the log-transformation of the dengue incidence. Data were sorted by latitude coordinate from south to north. (b) The aggregated dengue epidemic curve of the new number of dengue cases by symptom onset during the period 1994–2006.
We conservatively estimate (using the definition in this paper) an overall average dengue attack rate of 1·53 dengue cases/1000 people (s.d.=4·5). The average attack rates vary spatially from 0·0012 to 38·70 dengue cases/1000 people. Dengue attack rates over the 12-year window in Peru were negatively correlated with population size (Spearman ρ=−0·38, P<0·0001). No significant correlations with population density, latitude, longitude, and altitude are found. Final epidemic size is strongly correlated to the timing of the epidemic peak (Spearman ρ=0·66, P<0·0001) and its peak size (Spearman ρ=0·89, P<0·0001). The final epidemic size is found to be weakly correlated with longitude. That is, the final epidemic size increases as outbreaks ‘move’ from the jungle areas (−69° W) to the coastal areas (−81° W) (Spearman ρ=−0·14, P=0·01). The epidemic peak size followed a similar pattern (Spearman ρ=−0·16, P=0·004). The possibility of a dengue epidemic ‘wave’ cannot be discarded.
Transmissibility estimates
Estimates of the reproduction number (R) showed high variability. Their interquartile ranges (IQR) were from 0·83 to 4·46 with a median of 1·76. The estimated R ranged from 0·1 to 112·8. We estimated 59 reproduction numbers (43 dengue outbreaks had R>1) using dengue outbreaks with an initial epidemic phase comprising of at least five epidemic weeks. We found a moderate positive correlation between reproduction number and population size (Spearman ρ=0·28, P=0·03). We found that R is negatively correlated with the timing of the epidemic peak (Spearman ρ=−0·28, P=0·03) and positively correlated with the epidemic peak size (Spearman ρ=0·32, P=0·01). The estimates for R are not correlated with latitude or longitude coordinates, elevation, or type of geography (one-way ANOVA for comparison of means, P=0·20).
Critical community size
We found that the proportions of weeks with no dengue reports during the entire period (1994–2006) are negatively correlated with population size in jungle areas (Spearman ρ=−0·72, P<0·0001). Less than 30% of the weekly records had zero dengue incidences whenever the population was >500 000 people (Fig. 3). We did not find a correlation between these variables in coastal or mountain range areas.
Fig. 3.

The proportion of weeks with no dengue reports as a function of population size of the Peruvian provinces classified in coastal (▹), mountain (□), and jungle (○) areas. The proportion of weeks with no dengue reports during the entire dengue time were negatively correlated with population size in jungle areas (Spearman ρ=−0·72, P<0·0001) with <30% of weekly records with zero dengue incidence in jungle areas with a population >500 000 people (dashed line is a linear fit to the jungle data), a pattern not observed in coastal or mountain range areas.
Scaling laws in the distributions of attack rates and duration of epidemics
The distribution of dengue attack rates and epidemic (outbreak) durations (weeks) both follow power-law distributions (coefficient of determination >85%, P<0·004). Both distributions have approximately the same power-law exponent of 1·7 when data for the 1994–2006 outbreaks were used (Fig. 4).
Fig. 4.

The distributions of dengue attack rates and epidemic durations across Peruvian provinces during the period 1994–2006. Both distributions follow a power law with remarkably similar mean scaling exponents of about 1·7. The dashed lines are the log-log linear fits to the data (○).
Regression models for dengue attack rates and reproduction numbers
Regression models are used to explain the correlation structure between dengue attack rates and three significant predictor variables, population size (P=0·0004), epidemic duration (P<0·0001), and longitude (P=0·0023). In fact, simple models were obtained via stepwise regression. Population size and epidemic duration are log-transformed to stabilize their corresponding variances. This regression model explains 26% of the variability in the data with an overall P<0·0001. Moreover, population size (log transformed, P=0·0475) and epidemic duration (log transformed, P<0·0001) are found to be significant predictor variables for a model of the final epidemic size. This regression model explains 22% of the observed variance with an overall P<0·0001. No model with more than one significant predictor variable was found to explain the reproduction number estimates.
Spatial heterogeneity
The spatial heterogeneity of dengue incidence, as measured by the Gini index, decreases from the coastal areas (0·59), to the mountain range (0·36), to the jungle areas (0·27) (Fig. 5). The elevations of Peruvian provinces range from 0 m to 4942 m with high variability across coastal, mountain and jungle areas (one-way ANOVA for comparison of means, F=46·62, P<0·0001) (Fig. S3, online). The dengue attack rates (n=315) in the coastal areas are significantly higher than in the mountain range and the jungle areas (one-way ANOVA for comparison of means, P=0·0012). Dengue outbreaks are reported in provinces with a median elevation of 304 m (IQR 173-518). The outbreaks that occurred at the highest elevations were at altitudes of 1236 m (163 cases), 1277 m (10 cases), 1536 m (16 cases), and 4041 m (seven cases). The last outbreak was probably ‘imported’ from other locations (26/195 provinces are at an elevation >4041 m, and did not report dengue outbreaks). We do not find a significant correlation between elevation and final epidemic size (P=0·21), attack rates (P=0·55), or transmissibility as measured by the reproduction number (P=0·4).
Fig. 5.

The Lorenz curves (– – –) of the distribution of the total number of dengue case notifications as a function of population size at the level of provinces in Peru. The black line (—) represents a constant distribution of dengue case notifications (no heterogeneity).
Time-series analysis with climatological variables
We apply stepwise regression models using the weekly number of dengue cases as the response variable to four climatological variables as predictors (mean, minimum, and maximum temperature and precipitation). These models explain on average from 39·4% to 47·3% of the observed variance when the climatological variables are lagged from 0 to 20 weeks (Fig. 6). The best regression model (explaining a mean of 47·3% of the variability) is obtained when the climatological variables are lagged by 5 weeks. The regression model without lag periods in the climatological variables explains a mean of 42% of the observed variance. We found minimum temperature to be the most significant climatological variable, i.e. the one with the highest frequency in regression models. The maximum temperature ranked second (Fig. 6).
Fig. 6.

The correlation between dengue incidence and climatological variables for each of the dengue outbreaks occurring across the Peruvian provinces during the period 1994–2006. (a) Stepwise regression models of the weekly number of dengue cases using initially four climatological variables as predictors (mean, minimum, and maximum temperature and precipitation) explain on average between 39·4% and 47·3% of the observed variance when the climatological variables are lagged from 0 to 20 weeks. The best regression model (explaining a mean of 47·3% of the variability) was obtained when the climatological variables were lagged by 5 weeks. (b) The minimum temperature was found to be the most significant predictor variable in most of the regression models analysed for each dengue outbreak, followed by maximum temperature.
Timing of epidemic onset
Our analysis shows that the 2000–2001 epidemics were most likely to start in larger population areas from where the disease spreads to lower population areas (Spearman ρ=−0·43, P=0·03). This happens during a time when serotypes DEN-3 and DEN-4 are identified for the first time. Similar trends have been observed in other years, but the pattern for those repeated instances is not statistically significant. Hence, the possibility of epidemic outbreaks as the result of emergent and re-emergent diseases seems the most likely explanation. In other words, the absence of herd immunity, homogenizes the host population and increases the likelihood of epidemic outbreak waves.
DISCUSSION
We have analysed the transmission dynamics of dengue fever in Peru during the period 1994–2006 using weekly dengue incidence data stratified by provinces and a diverse set of statistical and mathematical tools. To the best of our knowledge this is the first population study of dengue fever at such spatial resolution. Our analyses included the estimation of the local reproduction number for each of the dengue outbreaks occurring across Peru during the period 1996–2004, the evaluation of the notion of critical community size previously studied in the context of other infectious diseases [15, 25], the characterization of the distributions of attack rates and epidemic durations, and the quantification of spatial heterogeneity using standard measures in economics (Lorenz curve and Gini index).
Estimates of the reproduction number (R) for dengue fever have varied significantly across locations because of variations in several factors including serotype circulation, available data, immunological history of the population, mosquito infestation levels, and climate variability. Our R estimates lie in the range from 0·1 to 112·8 (IQR 0·83–4·46). These are comparable with estimates for R (between 1·3 and 2·4) in the 1986 national dengue serosurvey in Mexico [37]; 1·1–3·3 in the state of Colima, Mexico in 2002 [16]; 4–6 in Thailand [38]; 1·6–2·5 in Sao Paulo, Brazil in 1991 [39]; 3·6–12·9 in Sao Paulo, Brazil in 2000 [40]; and 2–103 in nine Brazilian regions during the period 1996–2003 [41].
The data in jungle areas suggest that a critical community size of about half a million individuals is needed to sustain an epidemic. We did not observe a clear pattern in the role of community size in coastal or mountain areas where dengue seems to be reaching endemic levels (Fig. 3). Kuno [23] estimated that the critical community size lies between 150 000 and 1 000 000 to sustain a dengue epidemic in Puerto Rico and Trinidad. Wearing & Rohani [26] report that local dengue extinctions in Thai provinces become rare when the population size is higher than one million.
Heterogeneity in dengue attack rates was higher in coastal areas followed by mountain range areas and jungle areas. Dengue outbreaks occurred at elevations of up to ~1500 m, and a small possibly ‘imported’ outbreak of seven cases occurred at an elevation of 4041 m. In fact, mosquitoes are able to survive a life-cycle indoors at altitudes as high as 2200 m above sea level [42]. A dengue outbreak has been documented at an altitude of 1700 m in Mexico [43].
We found that most dengue outbreaks are associated with a small attack rate, although a small number of epidemics are associated with high attack rates. In fact, we found that the distribution of dengue attack rates and epidemic durations at the level of provinces in Peru follow power-law (Pareto) distributions with remarkably similar mean power-law exponents.
Minimum temperature was the most significant climatological variable in the stepwise regression models. This is in agreement with Yasuno & Tonn [19] who found that the lowest daily temperature, rather than the average temperature, was more influential on the extrinsic (mosquito) incubation period.
In our study, climatological variables alone were able to explain up to an average of about 50% in the variability in dengue incidence when these were temporarily lagged by 5 weeks. Other factors such as mosquito infestation levels that were not considered in this study might be contributing substantially to the dengue incidence levels in Peru. Such infestation levels will be directly associated with the intensity and timing of control interventions. In fact, a positive correlation between A. Aegypti indices and seroprevalence of dengue antibody levels has been reported, which suggests the presence of a critical vector density in order for dengue outbreaks to take place [44]. In some regions in Peru, highly heterogeneous mosquito infestation levels and dengue seroprevalence have been reported (e.g. [45–47]). Regarding the effects of vector control interventions, the fraction of houses infested by vector mosquitoes has been found to be negatively correlated with the intensity of anti-mosquito interventions in a Brazilian city [48]. Unfortunately, spatially resolved data on vector infestation levels across Peru was not available, and therefore, the interaction of climatological information, geography and demographics with infestation levels and the intensity and timing of control interventions remains an open question for future research.
The highest correlation of climatological variables with dengue incidence data at different temporal lags has been found to vary significantly across studies. For instance, an analysis using monthly data of the 2002 dengue epidemic in Colima, Mexico [49] indicated that precipitation, mean temperature, and minimum temperature were most correlated with dengue incidence without a lag period whereas maximum temperature and evaporation were most correlated to dengue incidence at lags of 1 and 3 months, respectively [49]. On the other hand, Keating [36] found the highest correlation between mean temperature and dengue incidence at a lag period of 3 months in Puerto Rico [36]. Furthermore, Depradine & Lovell [35] reported different lag periods with highest correlation with dengue incidence in the small Caribbean island of Barbados. They found a 6-week lag for vapour pressure, 7-week lag for precipitation, 12-week lag for minimum temperature, and a 16-week lag for maximum temperature.
Large population areas were associated with an earlier epidemic onset than low population areas during the 2000–2001 epidemic suggesting a hierarchical transmission network. This result is in accord with Cummings et al. [50] who identified a travelling wave of dengue infection in Thailand emanating from Bangkok and disseminating to less populous areas. Travelling waves have also been reported in the spread of other infectious diseases including measles [51] and seasonal influenza [52, 53].
Overall our findings indicate that highly refined spatial and temporal epidemic data of dengue fever are critically needed not only to increase our understanding of the dynamics of dengue fever around the world but also to generate new hypothesis and provide a platform for testing innovative control policies.
NOTE
Supplementary information accompanies this paper on the Journal's website (http://journals.cambridge.org).
DECLARATION OF INTEREST
None.
REFERENCES
- 1.Halstead SB, Cohen J, Powderly WG. Infectious Diseases. 2nd edn Vol. 2. Spain: Mosby; 2004. Dengue fever/dengue hemorrhagic fever; pp. 1681–1689. , vol. , pp. [Google Scholar]
- 2.World Health Organization http://www.who.int/mediacentre/factsheets/fs117/en/ http://www.who.int/mediacentre/factsheets/fs117/en/ . Dengue and dengue hemorrhagic fever ( ). Accessed 4 June 2007. [PubMed]
- 3.Center for Disease Control and Prevention http://www.cdc.gov/NCIDOD/DVBID/DENGUE/ http://www.cdc.gov/NCIDOD/DVBID/DENGUE/ . Dengue fever ( ). Accessed 16 July 2007.
- 4.Gubler DJ, Kuno G. Dengue and Dengue Hemorrhagic Fever. Wallingford, Oxford, UK: CABI Publishing; 1997. [Google Scholar]
- 5.Gubler DJ. Epidemic dengue/dengue hemorrhagic fever as a public health, social and economic problem in the 21st century. Trends in Microbiology. 2002;10:100–103. doi: 10.1016/s0966-842x(01)02288-0. [DOI] [PubMed] [Google Scholar]
- 6.Kochel T et al. Effect of dengue-1 antibodies on American dengue-2 viral infection and dengue haemorrhagic fever. Lancet. 2002;360:310–312. doi: 10.1016/S0140-6736(02)09522-3. [DOI] [PubMed] [Google Scholar]
- 7.Montoya Y et al. Circulation of dengue viruses in north-western Peru, 2000–2001. Dengue Bulletin. 2003;27:52–62. [Google Scholar]
- 8.Wikipedia http://en.wikipedia.org/wiki/Provinces_of_Peru. http://en.wikipedia.org/wiki/Provinces_of_Peru . Provinces of Peru ( ). Accessed 16 July 2007.
- 9.Dirección General de Epidemiología http://www.oge.sld.pe/ve_etapas.php. http://www.oge.sld.pe/ve_etapas.php . Epidemiological surveillance system. Stages of epidemiological surveillance ( ). Accessed 9 August 2007.
- 10.World Health Organization http://www.who.int/csr/resources/publications/dengue/012-23.pdf. http://www.who.int/csr/resources/publications/dengue/012-23.pdf . Clinical diagnosis of dengue ( ). Accessed 4 June 2007.
- 11.National Institute of Statistics and Informatics http://www.inei.gob.pe/ http://www.inei.gob.pe/ . Projection from 1993 census ( ). Accessed 13 July 2007.
- 12.Wikipedia http://es.wikipedia.org/wiki/Peru. http://es.wikipedia.org/wiki/Peru . Surface areas of provinces in Peru ( ). Accessed 13 July 2007.
- 13.MeteoRed http://tierra.meteored.com/tierra-America+del+Sur-Peru-esp.html. http://tierra.meteored.com/tierra-America+del+Sur-Peru-esp.html . Latitude and longitude coordinates, and elevation of provinces in Peru ( ). Accessed 19 July 2007.
- 14.MeteoRed http://clima.meteored.com/clima-en-America%20del%20Sur-Peru-PR.html. http://clima.meteored.com/clima-en-America%20del%20Sur-Peru-PR.html . Climate data for Peru ( ). Accessed 19 July 2007.
- 15.Anderson RM, May RM. Infectious Diseases of Humans. New York: Oxford University Press; 1991. [Google Scholar]
- 16.Chowell G et al. Estimation of the reproduction number of dengue fever from spatial epidemic data. Mathematical Biosciences. 2007;208:571–589. doi: 10.1016/j.mbs.2006.11.011. [DOI] [PubMed] [Google Scholar]
- 17.Jacquez JA. Compartmental Analysis in Biology and Medicine. Ann Arbor, MI: Thompson-Shore Inc.; 1996. [Google Scholar]
- 18.Morrison A et al. Temporal and geographic patterns of Aedes aegypti (Diptera: Culicidae) production in Iquitos, Peru. Journal of Medical Entomology. 2004;41:1123–1142. doi: 10.1603/0022-2585-41.6.1123. [DOI] [PubMed] [Google Scholar]
- 19.Yasuno M, Tonn RJ. A study of biting habits of Aedes aegypti in Bangkok, Thailand. Bulletin of the World Health Organization. 1970;43:319–325. [PMC free article] [PubMed] [Google Scholar]
- 20.Newton EAC, Reiter P. A model of the transmission of dengue fever with an evaluation of the impact of ultra-low volume (UVL) insecticide applications on dengue epidemics. American Journal of Tropical Medicine and Hygiene. 1992;47:709–720. doi: 10.4269/ajtmh.1992.47.709. [DOI] [PubMed] [Google Scholar]
- 21.Muir LE, Kay BH. Aedes aegypti survival and dispersal estimated by mark-release-recapture in northern Australia. American Journal of Tropical Medicine and Hygiene. 1998;8:277–282. doi: 10.4269/ajtmh.1998.58.277. [DOI] [PubMed] [Google Scholar]
- 22.Gubler DJ. Dengue and dengue hemorrhagic fever. Clinical Microbiology Reviews. 1998;11:480–496. doi: 10.1128/cmr.11.3.480. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Kuno G. Review of the factors modulating dengue transmission. Epidemiological Reviews. 1995;17:321–335. doi: 10.1093/oxfordjournals.epirev.a036196. [DOI] [PubMed] [Google Scholar]
- 24.Focks DA et al. A simulation model of the epidemiology of urban dengue fever: literature analysis, model development, preliminary validation, and samples of simulation results. American Journal of Tropical Medicine and Hygiene. 1995;53:489–506. doi: 10.4269/ajtmh.1995.53.489. [DOI] [PubMed] [Google Scholar]
- 25.Keeling MJ, Grenfell BT. Disease extinction and community size: modeling the persistence of measles. Science. 1997;275:65–67. doi: 10.1126/science.275.5296.65. [DOI] [PubMed] [Google Scholar]
- 26.Wearing HJ, Rohani P. Ecological and immunological determinants of dengue epidemics. Proceedings of the National Academy of Sciences USA. 2006;103:11802–11807. doi: 10.1073/pnas.0602960103. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Rhodes CJ, Anderson RM. Power laws governing epidemics in isolated populations. Nature. 1996;381:600–602. doi: 10.1038/381600a0. [DOI] [PubMed] [Google Scholar]
- 28.Lee WC. Characterizing exposure-disease association in human populations using the Lorenz curve and Gini index. Statistics in Medicine. 1997;16:729–739. doi: 10.1002/(sici)1097-0258(19970415)16:7<729::aid-sim491>3.0.co;2-a. [DOI] [PubMed] [Google Scholar]
- 29.Woolhouse ME et al. Heterogeneities in the transmission of infectious agents: implications for the design of control programs. Proceedings of the National Academy of Sciences USA. 1997;94:338–342. doi: 10.1073/pnas.94.1.338. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Kerani RP et al. Comparative geographic concentrations of 4 sexually transmitted infections. American Journal of Public Health. 2005;95:324–330. doi: 10.2105/AJPH.2003.029413. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Green CG, Krause D, Wylie J. Spatial analysis of campylobacter infection in the Canadian province of Manitoba. International Journal of Health Geographics. 2006;5:2. doi: 10.1186/1476-072X-5-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Jetten TH, Focks DA. Potential changes in the distribution of dengue transmission under climate warming. American Journal of Tropical Medicine and Hygiene. 1997;57:285–297. doi: 10.4269/ajtmh.1997.57.285. [DOI] [PubMed] [Google Scholar]
- 33.Li CF et al. Rainfall, abundance of Aedes aegypti and dengue infection in Selangor, Malaysia. Southeast Asian Journal of Tropical Medicine and Public Health. 1985;16:560–568. [PubMed] [Google Scholar]
- 34.Mourya DT, Yadav P, Mishra AC. Effect of temperature stress on immature stages and susceptibility of Aedes aegypti mosquitoes to chikungunya virus. American Journal of Tropical Medicine and Hygiene. 2004;70:346–350. [PubMed] [Google Scholar]
- 35.Depradine CA, Lovell E. Climatological variables and the incidence of dengue fever in Barbados. International Journal of Environmental Health Research. 2004;4:429–441. doi: 10.1080/09603120400012868. [DOI] [PubMed] [Google Scholar]
- 36.Keating J. An investigation into the cyclical incidence of dengue fever. Social Science and Medicine. 2001;53:1587–1597. doi: 10.1016/s0277-9536(00)00443-3. [DOI] [PubMed] [Google Scholar]
- 37.Koopman JS et al. Determinants and predictors of dengue infection in Mexico. American Journal of Epidemiology. 1991;133:1168–1178. doi: 10.1093/oxfordjournals.aje.a115829. [DOI] [PubMed] [Google Scholar]
- 38.Ferguson NM, Donnelly CA, Anderson RM. Transmission dynamics and epidemiology of dengue: insights from age-stratified sero-prevalence surveys. Proceedings of the Royal Society of London, Series B. 1999;354:757–768. doi: 10.1098/rstb.1999.0428. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Marques CA, Forattini OP, Massad E. The basic reproduction number for dengue fever in Sao Paulo state, Brazil: 1990–1991 epidemic. Transactions of the Royal Society of Tropical Medicine and Hygiene. 1994;88:58–59. doi: 10.1016/0035-9203(94)90498-7. [DOI] [PubMed] [Google Scholar]
- 40.Massad E et al. The risk of yellow fever in a dengue-infested area. Transactions of the Royal Society of Tropical Medicine and Hygiene. 2001;95:370–374. doi: 10.1016/s0035-9203(01)90184-1. [DOI] [PubMed] [Google Scholar]
- 41.Favier C et al. Early determination of the reproduction number for vector-borne diseases: the case of dengue in Brazil. Tropical Medicine and International Health. 2006;11:332–340. doi: 10.1111/j.1365-3156.2006.01560.x. [DOI] [PubMed] [Google Scholar]
- 42.Suarez MF, Nelson MJ. Records of altitude for Aedes aegypti in Colombia [in Spanish] Biomedica (Colombia) 1981;1:225. [Google Scholar]
- 43.Herrera-Basto E et al. First reported outbreak of classical dengue fever at 1,700 meters above sea level in Guerrero State, Mexico, June 1988. American Journal of Tropical Medicine and Hygiene. 1992;46:649–653. doi: 10.4269/ajtmh.1992.46.649. [DOI] [PubMed] [Google Scholar]
- 44.Figueroa M et al. Dengue epidemic in Honduras, 1978–1980. Bulletin Pan American Health Organization. 1982;16:130–137. [PubMed] [Google Scholar]
- 45.Morrison AC et al. Temporal and geographic patterns of Aedes aegypti (Diptera: Culicidae) production in Iquitos, Peru. Journal of Medical Entomology. 2004;41:1123–1142. doi: 10.1603/0022-2585-41.6.1123. [DOI] [PubMed] [Google Scholar]
- 46.Hayes CG et al. The epidemiology of dengue virus infection among urban, jungle, and rural populations in the Amazon region of Peru. American Journal of Tropical Medicine and Hygiene. 1996;55:459–463. doi: 10.4269/ajtmh.1996.55.459. [DOI] [PubMed] [Google Scholar]
- 47.Getis A et al. Characteristics of the spatial pattern of the dengue vector, Aedes aegypti, in Iquitos, Peru. American Journal of Tropical Medicine and Hygiene. 2003;69:494–505. [PubMed] [Google Scholar]
- 48.Pontes RJ et al. Vector densities that potentiate dengue outbreaks in a Brazilian city. American Journal of Tropical Medicine and Hygiene. 2000;62:378–383. doi: 10.4269/ajtmh.2000.62.378. [DOI] [PubMed] [Google Scholar]
- 49.Chowell G, Sanchez F. Climate-based descriptive models of dengue fever. Journal of Environmental Health. 2006;68:60–63. [PubMed] [Google Scholar]
- 50.Cummings DAT et al. Traveling waves in the occurrence of dengue hemorrhagic fever in Thailand. Nature. 2004;427:344–347. doi: 10.1038/nature02225. [DOI] [PubMed] [Google Scholar]
- 51.Grenfell BT, Bjornstad ON, Kappey J. Travelling waves and spatial hierarchies in measles epidemics. Nature. 2001;414:716–723. doi: 10.1038/414716a. [DOI] [PubMed] [Google Scholar]
- 52.Viboud C et al. Synchrony, waves, and spatial hierarchies in the spread of influenza. Science. 2006;312:447–451. doi: 10.1126/science.1125237. [DOI] [PubMed] [Google Scholar]
- 53.Alonso WJ et al. Seasonality of influenza in Brazil: a traveling wave from the Amazon to the subtropics. American Journal of Epidemiology. 2007;165:1434–1442. doi: 10.1093/aje/kwm012. [DOI] [PubMed] [Google Scholar]