

Abstract
We study the handicap principle in terms of the Sir Philip Sidney game. The handicap principle asserts that cost is required to allow for honest signalling in the face of conflicts of interest. We show that the significance of the handicap principle can be challenged from two new directions. Firstly, both the costly signalling equilibrium and certain states of no communication are stable under the replicator dynamics (i.e. standard evolutionary dynamics); however, the latter states are more likely in cases where honest signalling should apply. Secondly, we prove the existence and stability of polymorphisms where players mix between being honest and being deceptive and where signalling costs can be very low. Neither the polymorphisms nor the states of no communication are evolutionarily stable, but they turn out to be more important for standard evolutionary dynamics than the costly signalling equilibrium.
Keywords: costly signalling, evolutionary dynamics, handicap principle, polymorphic equilibrium, Sir Philip Sidney game
1. Introduction
The Sir Philip Sidney game (Maynard Smith 1991) has been a locus of attention for the analysis of costly signalling between relatives. It represents perhaps the simplest game that captures the central notion of handicaps owing to Zahavi (1975); i.e. that cost is required to maintain honest signalling in the presence of partial conflict of interest. The canonical example for the Sir Philip Sidney game is the interaction between feeding chicks and their parents. Maynard Smith introduced the discrete Sir Philip Sidney game as a simplified version of Grafen's (1990) notable costly signalling model (see also Johnstone & Grafen 1992, 1993; Bergstrom & Lachmann 1997, 1998; Lachmann & Bergstrom 1998; Godfray & Johnstone 2000; Brilot & Johnstone 2003; Hamblin & Hurd 2009). Many variations of the initial game have been considered, and there is an ongoing debate about how widespread this phenomenon is (Maynard Smith & Harper 2003; Searcy & Nowicki 2005). However, much of this debate retains the central methodology of calculating evolutionarily stable states (ESS) (Maynard Smith & Price 1973; Maynard Smith 1982).
In this paper, we conduct a dynamic analysis of the original Sir Philip Sidney game, which identifies gaps in our understanding of honest signalling caused by the strong focus on ESS. We concentrate on two issues. Firstly, we contrast the evolutionary significance of states of perfect communication (the signalling ESS) and states of no communication (pooling equilibria). In most of the cases relevant for costly signalling, the pooling equilibrium appears to be the most probable evolutionary outcome in terms of having the larger basin of attraction under standard evolutionary dynamics (§4). Secondly, while the existence of the signalling ESS and of pooling equilibria is a well-established result in the literature, we prove that, in addition, a family of polymorphic equilibria exists which allow for partial communication (§2). These polymorphisms can establish partly honest signalling even in the face of low signalling costs. We prove dynamic stability of the polymorphic equilibrium (§3), and show that this outcome is in many cases more likely than the signalling ESS (§4). These results have some important biological implications. We suggest in particular that one should not place too much weight on perfect communication in situations involving conflicting interests; instead, one may look for alternative equilibria such as the polymorphisms described below. We shall discuss this issue more thoroughly in the concluding section of this paper.
2. The Sir Philip Sidney game
The Sir Philip Sidney game proceeds in two stages. A sender can be either healthy (with probability 1 − m) or needy (with probability m). In both states, the sender can send a signal bearing a cost c > 0, or decline sending a signal at all. In the second stage, after having (or not having) received the signal, a donor may respond by transferring a resource d > 0 to the sender and thus reducing her survival probability to 1 − d, or may abstain from doing so. The donor does not know the sender's true state. The extensive form of this game is depicted in figure 1. Without receiving the resource, a sender's probability of surviving is 1 − a if she is needy and 1 − b if she is healthy; we assume that a > b throughout this paper (i.e. a needy individual profits more from receiving the resource).
Figure 1.

An extensive form representation of the Sir Philip Sidney game. The dotted lines represent the donor's information sets, i.e. the donor is unable to distinguish between the decision nodes connected by the dotted lines. The terminal nodes show the sender's and the donor's payoffs, respectively.
The extensive form game of figure 1 does not allow for communication as the donor's strategy of never transferring the resource is strictly dominant. Introducing a relatedness parameter k ∈ [0,1] permits a higher degree of common interest between the sender and the donor. At each outcome, a player receives her own payoff plus k times the payoff of the other player. This gives rise to a four-by-four strategic-form game. A player's strategies are given by a rule of behaviour which tells her what to do at each of her information sets. These strategies are enumerated in table 1.
Table 1.
Sender and donor strategies in the Sir Philip Sidney Game.
| sender strategies | donor strategies |
|---|---|
| S1: signal only if healthy | R1: donate only if no signal |
| S2: signal only if needy | R2: donate only if signal |
| S3: never signal | R3: never donate |
| S4: always signal | R4: always donate |
Investigators often reduce the four-by-four strategic-form structure by ignoring the strategies ‘signal only if healthy’ and ‘donate only if no signal’ (like Bergstrom & Lachmann 1997; Maynard Smith & Harper 2003). In this truncated game, two kinds of equilibria are usually considered. The first one is the most important Nash equilibrium in the literature on handicap signals. It is given by the strategy pair ‘signal only if needy’ and ‘donate only if signal’, and it is a Nash equilibrium if
| 2.1a |
and
| 2.1b |
(Bergstrom & Lachmann 1997). At this equilibrium, cost can be said to maintain signal reliability as a needy sender is willing to pay the cost c in order to get the resource, while a healthy sender does not pretend to be needy by sending the signal. If the inequalities are strict, then the equilibrium is a strict Nash equilibrium and, therefore, also an ESS in the symmetrized game (cf. Cressman 2003). This equilibrium is usually called a signalling ESS or a separating equilibrium.
There always exists a pooling equilibrium where no information is transferred. Bergstrom & Lachmann (1997) show that the profile ‘never donate’ and ‘never signal’ is Nash equilibrium if
| 2.2 |
if this inequality is reversed, then the profile ‘always donate’ and ‘never signal’ is Nash equilibrium. As the signal is not sent at both equilibria, these profiles can be called pooling equilibria.
In appendix B, we prove that ‘signal only if healthy’ and ‘donate only if no signal’ is Nash equilibrium if equation (2.1b) holds together with
| 2.3 |
This equilibrium is often ignored by other investigators on grounds of requiring too much common interest between sender and donor to be relevant for costly signalling theory (Maynard Smith 1991; Bergstrom & Lachmann 1997). We think that this exclusion is a mistake. Firstly, the existence conditions of this equilibrium play a role in interpreting dynamical properties of the Sir Philip Sidney game as parameters are varied (§§3 and 4). Moreover, R1 is important for the equilibrium structure of one of the pooling equilibria. Once we allow all sender and donor strategies, a pooling equilibrium will never be a strict Nash equilibrium. Given that the sender chooses ‘never signal’, R2 and R3 as well as R1 and R4 are behaviourally equivalent. In appendix A, we show that pooling equilibria are elements of larger sets of Nash equilibria. In the case of (S3, R3), the donor can play a mixture (1 − λ)R2 + λ R3. As long as
| 2.4 |
the strategy profile (S3, (1 − λ)R2 + λR3) is a Nash equilibrium if equation (2.2) holds; otherwise, (S3, (1 − μ)R1 + μR4) is a Nash equilibrium as long as
| 2.5 |
As pooling equilibria are elements of a line of equilibria, they cannot be evolutionarily stable. One can show, however, that they correspond to neutrally stable strategies (Maynard Smith 1982). We will come back to this point shortly in the broader context of dynamic stability (§3).
To the best of our knowledge, another kind of equilibrium has been completely overlooked in the biological literature. In the corresponding economics literature (starting with Spence 1973), similar equilibria—called hybrid equilibria—are known to exist but were considered unimportant. Recently, Wagner (2009) has shown that, contrary to received wisdom, a hybrid equilibrium can be very significant in Spence's game from the point of view of game dynamics. In the Sir Philip Sidney game, there exists a family of polymorphisms which corresponds to hybrid equilibria. Each polymorphism is a mixed Nash equilibrium where the sender mixes between ‘signal only when healthy’ and ‘always signal’, while the donor mixes between ‘donate if signal’ and ‘never donate’. The family of polymorphic equilibria is given by λS2 + (1 − λ)S4 and μR2 + (1 − μ)R3, where
| 2.6 |
As λ, μ must be well defined, the polymorphism (2.6) exists if
| 2.7 |
and if equation (2.2) is met. (The proof can be found in appendix B.) The conditions (2.7) show that, under generic conditions, the mixed Nash equilibrium (2.6) exists if and only if the signalling ESS (2.1) does not exist. The smallest cost which allows for the existence of a seperating equilibrium is given by c* = b − kd. This is, in a sense, the minimum cost of a believable signal (Bergstrom & Lachmann 1997). Condition (2.7) implies that at the polymorphism, the cost of the signal will be lower than c*. However, the polymorphic equilibrium sustains some level of meaningful communication. Thus, information transfer is possible in a wider range of cases than was previously believed. (Notice that partly honest communication here does not arise because of the introduction of different sender types as in Johnstone & Grafen (1993).)
3. Dynamic stability of equilibria
The multiplicity of equilibria in the Sir Philip Sidney game makes a priori conclusions about its evolutionary outcomes difficult. In order to assess the evolutionary significance of the different equilibria, we shall investigate their dynamic stability properties in terms of the two-population replicator dynamics (Hofbauer & Sigmund 1998). If xi is the relative frequency of sender type i and yj is the relative frequency of donor type j, i, j = 1, … ,4, then this dynamics is given by
| 3.1a |
and
| 3.1b |
Here, 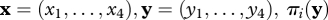 is the payoff of i against y and π(x, y) is the average payoff in the sender population; πj(x) is the payoff of j against x and π(y, x) is the average payoff in the donor population. Many of our results also apply to the one-population replicator dynamics if we consider the symmetrized Sir Philip Sidney game (for details, see Cressman 2003).
is the payoff of i against y and π(x, y) is the average payoff in the sender population; πj(x) is the payoff of j against x and π(y, x) is the average payoff in the donor population. Many of our results also apply to the one-population replicator dynamics if we consider the symmetrized Sir Philip Sidney game (for details, see Cressman 2003).
Under the relevant conditions ((2.1a,b) and (2.3) with strict inequalities, respectively) (S2, R2) and (S1, R1) are strict Nash equilibria and therefore asymptotically stable population states for equations (3.1). In appendix A, we show that if the inequalities in equations (2.4) and (2.5) are strict, then the equilibria in the components (2.4) or (2.5) are quasi-strict. (An equilibrium is quasi-strict if there is no best repsonse to any of its components outside of its support.) In the symmetrized version of the game, these profiles are neutrally stable. Quasi-strictness, in turn, implies the following result (for details on why quasi-strictness implies dynamic stability in this case cf. Cressman 2003).
Theorem 3.1. —
The pooling equilibria given by equations (2.4) and (2.5) attract an open set of nearby population states under the appropriate conditions.
Theorem 3.1 tells us that pooling equilibria are meaningful for the replicator dynamics as a non-negligible portion of initial populations will end up in the set of pooling equilibria.
The dynamic stability properties of the polymorphic equilibrium (2.6) are particularly interesting. In appendix B, we prove two results: (i) all transversal eigenvalues of the Jacobian matrix J of equations (3.1) evaluated at the points given by (2.6) are negative and (ii) the two remaining eigenvalues of J are purely imaginary. Both results hold if we assume that conditions (2.2) and (2.7) are met. (i) and (ii) imply that the polymorphism (2.6) is a spiralling centre; i.e. initial population states close to the polymorphism in the interior of the state space converge towards K = span(S2, S4) × span(R2, R3) and, once they get close enough, spiral around the polymorphism forever. The following theorem summarizes the stability properties of the polymorphic equilibrium.
Theorem 3.2. —
If equations (2.2) and (2.7) hold, then the polymorphic equilibrium (2.6) is Liapunov stable under the replicator dynamics (3.1).
As all transversal eigenvalues are negative, the interesting dynamical behaviour is confined to K. The phase portrait of this face for one set of parameter values is depicted in figure 2. We should note that this dynamical behaviour is structurally unstable (Guckenheimer & Holmes 1983); perturbations of the dynamics (3.1) will either result in an asymptotically stable or an unstable polymorphism. The issue of structural instability is a subtle one and needs more space; see Hofbauer & Huttegger (2008) for a case study. Suffice it to say here that many perturbations will result in an asymptotically stable polymorphism.
Figure 2.

A phase portrait of one boundary face of the space of population states (face K) of the Sir Philip Sidney game illustrating the motion around the polymorphic equilibrium. Close to the unique rest point, interior trajectories (of the full state space) converge to K. Notice the resemblance between this phase portrait and the phase portrait of the well-known game of matching pennies under the replicator dynamics.
4. Basins of attraction
The results of the previous section suggest that one cannot immediately use the ESS analysis of the Sir Philip Sidney game to conclude that signalling is likely to evolve. Although for many parameter configurations the separating equilibrium (S2, R2) is an ESS, there are several other outcomes which have a non-negligible basin of attraction. Both pooling equilibria and also the other separating equilibrium can be outcomes of evolution. If we are interested in determining the likelihood that signalling will evolve, we have to estimate the relative sizes of the basins of attraction for the various outcomes. In this section, we will use numerical simulations to resolve this issue.
Investigators generally regard the region where k is high to be uninteresting because signalling is stable for all values of c, including c = 0. We nevertheless start with the limiting case k = 1 as an important benchmark case. Figure 3 shows the size of the basins of attraction for both separating equilibria. The initial population states not converging to them are of non-negligible size. In fact, cost appears to hinder the evolution of successful signalling. Understanding this result is not difficult. Suppose that we have a population of donors who adopt either the strategy ‘donate only if signal’ or ‘never donate’. Let the proportion of the former be represented by α. The sender prefers the strategy ‘signal only if needy’ to the strategy ‘never signal’ only when (15/32)α > c. So, as c grows, there must be a proportionally larger percentage of the donor population who will respond to a signal in order for signalling to be beneficial for the sender.
Figure 3.

Simulation results showing the basin of attraction for the boundary face containing the hybrid equilibrium for different values of m and c. Here, k = 0.54, a = 31/32, b = 9/32 and d = 1/2. Plus symbol, c = 0; multiplication symbol, c = 0.05; asterisk, c = 0.1; unfilled square, c = 0.15; filled square, c = 0.2.
While costs generally hurt the evolution of signalling for these parameter values, it does not do so in every case. For extreme values of m, moderate cost appears to assist the evolution of signalling. The mechanism by which this result is produced is relatively complicated, but looking at a slightly simplified situation is insightful. Consider the initial population state illustrated in table 2 for m = 0.1 (i.e. the sender is rarely needy). If c = 0, there is no selection acting on the sender population at all. However, there is selection in the donor population in favour of the strategy ‘never donate’. So without cost, the population converges to a pooling equilibrium. This occurs because there is little information in the sender's signal. When c > 0, there is selection acting on the sender population, primarily in favour of the strategy ‘never signal’. Although the separating strategy ‘signal only if needy’ does worse than ‘never signal’, it does better than average and is initially selected for. (This is illustrated in the ‘selective force’ column, which is the payoff of that type minus the average payoff in the population.) As it increases in proportion, it changes the selective force exercised on the donor population sufficiently that ‘donate only if signal’ becomes superior and grows. Once a sufficiently large percentage of the donor population is playing ‘donate only if signal’, the selective force exercised on the sender population shifts in favour of ‘signal only if needy’ over ‘never signal’, and the system evolves to a state with perfect information transfer. A similar situation obtains for the case of m being sufficiently high.
Table 2.
An initial population and its associated payoffs that illustrate how a smaller c might retard the evolution of signalling for extreme values of m.
| strategy | proportion | payoff | selective force |
|---|---|---|---|
| S1, signal if healthy | 0.05 | 1.626 − 0.9c | −0.254c |
| S2, signal if needy | 0.01 | 1.626 − 0.1c | 0.546c |
| S3, never signal | 0.34 | 1.626 | 0.646c |
| S4, signal always | 0.6 | 1.626 − c | −0.354c |
| R1, donate if no signal | 0.09 | 2.22713 − 0.646c | |
| R2, donate if signal | 0.09 | 2.21388 − 0.646c | |
| R3, never donate | 0.75 | 2.3265 − 0.646c | |
| R4, always donate | 0.07 | 2.115 − 0.646c |
This considers only the case of k = 1. Figure 4 illustrates what happens as k is reduced. One will notice from figure 3 that the basins of attraction for separating equilibria are maximized when m = 0.3. This represents the situation where, if the sender provides no information (by playing either S3 or S4), the donor is indifferent between donating or not. This indifference point changes as k is reduced. For each value of k in figure 4, we set m equal to this point of indifference so as to maximize the basins of attraction for signalling. This provides the most benign scenario for the evolution of signalling. In figure 4, we see that so long as cost free signalling is an ESS, it always evolves. However, similar to what happens if k = 1, the presence of cost hinders the evolution of signalling and the higher costs result in smaller basins of attraction. One will notice that, as k is reduced, there are sudden drops in the basins of attraction of costly signalling. This occurs as one crosses the boundaries in equation (2.3), when the equilibrium where the signal is used to indicate health no longer exists.
Figure 4.

Simulation results showing the cumulative size of the basins of attraction for the two signalling equilibria as k and m vary. m is set so as to maximize the basins of attraction of signalling given the specified k. As before, the simulation uses the discrete time replicator dynamics when a = 31/32, b = 9/32 and d = 1/2. Plus symbol, c = 0.20; multiplication symbol, c = 0.15; asterisk, c = 0.10; unfilled square, c = 0.05; filled square, c = 0.
If 0.51 < k < 0.56, then k is sufficiently low that (S2, R2) is only stable with some signal cost. Here, one will notice a few interesting features. In the first place, although it is the only ESS, the separating equilibrium has a relatively small basin of attraction—it is always less than half the state space. This suggests that an analysis, based on the ESS concept, which ignores aspects of evolutionary processes can lead to misleading implications. Moreover, we see that the lower costs usually produce larger basins of attraction for signalling, similar to what happens in the common interest case. For brevity's sake, we cannot report results for other values of k and m; but we have found that, like in the k = 1 case, the relationship between cost and the evolution of signalling can be very complex.
Finally, we turn to the evolutionary significance of the polymorphic equilibrium. Figure 5 illustrates the basin of attraction of the face K (described in §3). These basins of attraction are for parameters where signal cost is required in order to sustain full communication. For these settings, the minimum believable cost c* ≈ 0.011. However, the corresponding polymorphic equilibrium has a significant basin of attraction for costs of an order of magnitude smaller. When comparing the results here with figure 4, one sees that, by considering the best cases, the polymorphic equilibrium has a larger basin of attraction (for fixed k), than does the signalling ESS with higher signal costs. Because of this result, we believe that the polymorphic equilibrium may be more relevant than the signalling ESS to the study of signalling in existing populations.
Figure 5.

Simulation results showing the cumulative size of the basins of attraction for the two signalling equilibria when k = 1 for several differing values of c. The simulation uses the discrete time replicator dynamics when a = 31/32, b = 9/32 and d = 1/2. Plus symbol, c = 0.001; multiplication symbol, c = 0.005; asterisk, c = 0.01.
5. Discussion
Our results point out certain limitations of the ESS methodology insofar as one identifies the long-run evolutionary outcomes with ESS. By focusing on ESS, traditional investigations of the Sir Philip Sidney game suggest that the evolution of costly signalling is far more likely than it appears to be for standard evolutionary dynamics. While the signalling equilibrium is the only ESS of the game for conflicting interests between the players, our dynamic analysis indicates that there are other states, namely pooling equilibria, which attract a significant portion of the initial populations. From the point of view of standard evolutionary dynamics, pooling equilibria also have a larger basin of attraction for many specifications of the parameters, indicating that they are the most probable evolutionary outcome in these cases. This suggests that the pooling equilibrium will be observed more often than the signalling equilibrium as there are more initial populations converging to the former than to the latter. We would like to emphasize that this holds for those parameters where the handicap principle should apply because of conflicts of interest between sender and donor.
The existence and stability of the polymorphic equilibria (2.6) illustrates another problem of the ESS methodology. By exclusively investigating the conditions under which a signalling ESS exists, one loses sight of other evolutionarily significant outcomes. This also poses problems for empirical studies of signalling, as some theoretically well-founded evolutionary outcomes may not be known; consequently, the corresponding predictions of the existence of, e.g. polymorphisms such as given in equation (2.6) are not tested in the field.
We believe that the existence of partial information transfer even in situations where communication was believed to be impossible, as exemplified by the polymorphic equilibrium, has broad biological implications. For the relevant parameters, the polymorphic equilibrium appears to be a more likely evolutionary outcome than the traditional costly signalling ESS. As an example, one may consider the paradigmatic signalling interaction between relatives, the begging of chicks for food from their parents. Parents would like to know the state of the chick while the chick would prefer food regardless of its state. If signalling need is associated with a sufficiently high cost c, then the Sir Philip Sidney game (and many other signalling models) predicts that the state where only needy chicks send the signal and where parents react to the signal is a possible evolutionary outcome. Our new results predict that even if c is very low, an alternative equilibrium is possible which is weakly stable and allows some information transfer between chicks and their parents. More specifically, the population dynamics may lead to a state where some, but not all chicks will signal need honestly; and some parents will respond to the signal by transferring food. The other chicks will always use the signal regardless of their state, and the other parents will never transfer food.
Our results also suggest that, for the relevant parameters, the polymorphic equilibrium is more likely to be observed than the signalling ESS. This resonates one of the issues found in many experiments which seek to determine the cost in signalling interactions such as solicitation (Searcy & Nowicki 2005). Metabolic costs often do not seem to be high enough to accord with the existence of a signalling ESS (Bachmann & Chapell 1998). Cost by risk of predation has also not been shown to be consistently high (Haskell 1994). Even if costs are not sufficiently high to sustain full communication, one may find communication in a polymorphic state. This opens a new avenue for empirical research which might prove insightful.
Acknowledgments
We would like to thank Elliott Wagner for teaching us the importance of hybrid equilibria in Spence's signalling game. We also thank Carl Bergstrom, Josef Hofbauer and an anonymous referee for helpful comments.
Appendix A. Geometry and dynamic stability of pooling equilibria
Suppose that the donor mixes between R2 and R3. Then, S2 and S3 earn the same payoff against (1 − λ)R2 + λR3 if
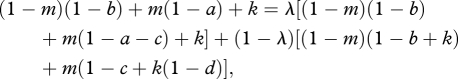 |
or if
If λ > λ*, then S3 gets a higher payoff than S2; the reverse relation obtains if the inequality is reversed. Concerning S4, we have to know when 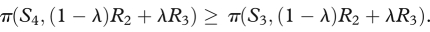 . This equation is equivalent to
. This equation is equivalent to
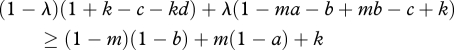 |
or
As a ≥ b, it is clear that the right-hand side of this inequality is less than or equal to λ*. Hence, if λ > λ*, then S4 will also earn less payoff than S3. As to S1,  if
if
 |
which is clearly always the case as c, kd ≥ 0. We already know that equation (2.2) implies that there is no donor strategy which does better against S3 than either R2 or R3. The relation (2.5) can be proved similarly. These arguments also show that both kinds of lines of Nash equilibria are quasi-strict (and neutrally stable in the symmetrized game) under the appropriate existence conditions, as donor strategies not in the support of those equilibria earn less payoff than the strategies in their support.
Let us consider the line of strategy profiles given by (S3, (1 − λ)R2 + λ R3). Rest points in the relative interior of this line attract an open set of nearby initial population states. This follows from the centre-manifold theorem (Carr 1981), together with the fact that all transversal eigenvalues of the Jacobian matrix of equations (3.1) evaluated at those rest points are negative. The latter fact is shown by the calculations above.
Appendix B. Alternative equilibria
Under certain conditions, (S1, R1) is a strict Nash equilibrium. Note that the condition for donors to transfer the resource must be the same as for the equilibrium (2.1), i.e. a ≥ d/k ≥ b. The reason for this is that the decision problem of the donor is the same in both cases. As to the sender strategies, we have to find the conditions under which Si, i = 2,3,4, earn less payoff against R1 than S1 does. Concerning S3, π(S1, R1) ≥ π(S3, R1), if
This is the case if and only if kd − c ≥ b. Similarly, π(S1, R1) ≥ π(S4, R1), if
 |
which is equivalent to a ≥ kd − c. The relation b ≤ kd − c implies d ≥ (b + c)/k, and thus
Furthermore, a ≥ kd − c implies
From these two inequalities, it follows that
 |
which is the same as π(S1, R1) ≥ π(S2, R1).
We next prove the location of (p, q) = ((λS2 + (1 − λ)S4), (μR2 + (1 − μ)R3)), 0 < λ,μ < 1. In order to do this, we follow Hofbauer & Sigmund (1998, §10.4). The relevant payoff information for the game restricted to K = span(S2, S4)× span(R2, R3) is contained in the two payoff matrices
where
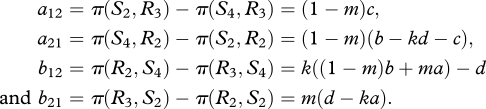 |
As shown in Hofbauer & Sigmund (1998, §10.4), a unique interior equilibrium exists if a12a21 > 0 and b12b21 > 0. As c > 0, we must have b − kd > c. If b12 > 0, then (p, q) is a saddle (provided that b12b21 > 0). We will instead focus on the more interesting case where b12b21 < 0. This means that condition 2.2 is met and that ka > d. The rest point (p, q) is given by
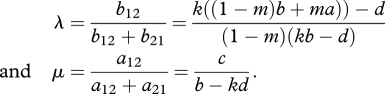 |
Now a12b12 < 0, and hence the Jacobian matrix of equations (3.1) evaluated at (p, q) has purely imaginary eigenvalues (Hofbauer & Sigmund 1998, §10.4).
We next show that all transversal eigenvalues of the Jacobian matrix of equations (3.1) at (p, q) are negative. For the sender, the transversal eigenvalues are given by πi(q) − π(p, q) for i = 1,3. It is easy to show that π1(q) < π(p, q) and π3(q) < π(p, q) if μ > c/(a − kd). The transversal eigenvalue π1(p) − π(q, p) is negative if and only if d > bk. Both conditions follow straightforwardly from our assumptions. Similarly, π4(p) < π(q, p) if equation (2.2) holds. That the polymorphism (2.6) is a spiralling centre follows from the centre-manifold theorem (Carr 1981). In our case, this manifold coincides with an open neighbourhood in K around the polymorphism (2.6). Nearby solution trajectories approach the centre manifold exponentially. But on K, solution trajectories spiral around equation (2.6) forever. We also note that the polymorphic equilibrium is a Nash–Pareto pair (Hofbauer & Sigmund 1998, §11.4). This means that the polymorphic equilibrium is almost as stable as a strict Nash equilibrium.
References
- Bachmann G. C., Chapell M. A.1998The energetic cost of begging behaviour in nestling house wrens. Anim. Behav. 55, 1607–1618 (doi:10.1006/anbe.1997.0719) [DOI] [PubMed] [Google Scholar]
- Bergstrom C. T., Lachmann M.1997Signalling among relatives I. Is costly signalling too costly? Phil. Trans. R. Soc. Lond. B 352, 609–617 (doi:10.1098/rstb.1997.0041) [Google Scholar]
- Bergstrom C. T., Lachmann M.1998Signalling among relatives III. Talk is cheap. Proc. Natl Acad. Sci. USA 95, 5100–5105 (doi:10.1073/pnas.95.9.5100) [DOI] [PMC free article] [PubMed] [Google Scholar]
- Brilot B. O., Johnstone R. A.2003The limits to cost-free signalling of need between relatives. Proc. R. Soc. Lond. B 270, 1055–1060 (doi:10.1098/rspb.2003.2329) [DOI] [PMC free article] [PubMed] [Google Scholar]
- Carr J.1981Applications of centre manifold theory New York, NY: Springer [Google Scholar]
- Cressman R.2003Evolutionary dynamics and extensive form games Cambridge, MA: MIT Press [Google Scholar]
- Godfray H. C. J., Johnstone R. A.2000Begging and bleating: the evolution of parent–offspring signalling. Phil. Trans. R. Soc. Lond. B 355, 1581–1591 (doi:10.1098/rstb.2000.0719) [DOI] [PMC free article] [PubMed] [Google Scholar]
- Grafen A.1990Biological signals as handicaps. J. Theor. Biol. 144, 517–546 (doi:10.1016/S0022-5193(05)80088-8) [DOI] [PubMed] [Google Scholar]
- Guckenheimer J., Holmes P.1983Nonlinear oscillations, dynamical systems, and bifurcations of vector fields New York, NY: Springer [Google Scholar]
- Hamblin S., Hurd P. L.2009When will evolution lead to deceptive signalling in the Sir Philip Sidney game? Theor. Popul. Biol. 75, 176–182 (doi:10.1016/j.tpb.2009.02.002) [DOI] [PubMed] [Google Scholar]
- Haskell D.1994Experimental evidence that nestling begging behaviour incurs a cost due to nest predation. Proc. R. Soc. Lond. B 257, 161–164 (doi:10.1098/rspb.1994.0110) [Google Scholar]
- Hofbauer J., Huttegger S. M.2008Feasibility of communication in binary signalling games. J. Theor. Biol. 254, 843–849 (doi:10.1016/j.jtbi.2008.07.010) [DOI] [PubMed] [Google Scholar]
- Hofbauer J., Sigmund K.1998Evolutionary games and population dynamics Cambridge, UK: Cambridge University Press [Google Scholar]
- Johnstone R. A., Grafen A.1992The continuous Sir Philip Sidney game: a simple model of biological signalling. J. Theor. Biol. 156, 215–234 (doi:10.1016/S0022-5193(05)80674-5) [DOI] [PubMed] [Google Scholar]
- Johnstone R. A., Grafen A.1993Dishonesty and the handicap principle. Anim. Behav. 46, 759–764 (doi:10.1006/anbe.1993.1253) [Google Scholar]
- Lachmann M., Bergstrom C. T.1998Signalling among relatives II. Beyond the Tower of Babel. Theor. Popul. Biol. 54, 146–160 (doi:10.1006/tpbi.1997.1372) [DOI] [PubMed] [Google Scholar]
- Maynard Smith J.1982. Evolution and the theory of games Cambridge, UK: Cambridge University Press [Google Scholar]
- Maynard Smith J.1991. Honest signalling: the Philip Sidney game. Anim. Behav. 42, 1034–1035 [Google Scholar]
- Maynard Smith J., Harper D.2003Animal signals Oxford, UK: Oxford University Press [Google Scholar]
- Maynard Smith J., Price G.1973The logic of animal conflict. Nature 146, 15–18 (doi:10.1038/246015a0) [Google Scholar]
- Searcy W. A., Nowicki S.2005The evolution of animal communication: reliability and deception in signalling systems Princeton, NJ: Princeton University Press [Google Scholar]
- Spence M.1973Job market signalling. Quart. J. Econ. 87, 355–374 (doi:10.2307/1882010) [Google Scholar]
- Wagner E.2009Dynamics of costly signalling. Working Paper, University of California at Irvine, USA [Google Scholar]
- Zahavi A.1975Mate selection—a selection for a handicap. J. Theor. Biol. 53, 205–214 (doi:10.1016/0022-5193(75)90111-3) [DOI] [PubMed] [Google Scholar]