

Abstract
The discovery that delta-9-tetrahydrocannabinol (Δ9-THC) is the primary psychoactive ingredient in marijuana prompted research that helped elucidate the endogenous cannabinoid system of the brain. Δ9-THC and other cannabinoid ligands with agonist action (CP 55,940, HU210, and WIN 55,212-2) increase firing of dopamine neurons and increase synaptic dopamine in brain regions associated with reward and drug addiction. Such changes in cellular processes have prompted investigators to examine the conditioned rewarding effects of the cannabinoid ligands using the place conditioning task with rats and mice. As reviewed here, these cannabinoid ligands can condition place preferences (evidence for rewarding effects) and place aversions (evidence for aversive qualities). Notably, the procedural details used in these place conditioning studies have varied across laboratories. Such variation includes differences in apparatus type, existence of procedural biases, dose, number of conditioning trials, injection-to-placement intervals, and pretraining drug exposure. Some differences in outcome across studies can be explained by these procedural variables. For example, low doses of Δ9-THC appear to have conditioned rewarding effects, whereas higher doses have aversive effects that either mask these rewarding effects or condition a place aversion. Throughout this review, we highlight key areas that need further research.
Keywords: Drug addiction, choice behavior, conditioned place preference, marijuana, mesolimbic dopamine, reward
1. Introductory remarks
Isolation of brain cannabinoid (CB) receptors and the endogenous CB compounds, arachidonylethanolamide (anandamide) and 2-arachidonylglycerol (2-AG), as well as the development of exogenous ligands, has enabled a growing body of research into the actions of cannabinoids in the brain and their effects on behavior. Because the primary active ingredient of marijuana is cannabinoidergic and because the prevalence of marijuana use is a global concern, an area of particular interest is how the CB system functions within the brain reward system. Place conditioning is a common and potentially useful task for evaluating the conditioned motivational effects of a drug (1,2). In this task, the animal (usually a rat or mouse) has a distinct environment (context) repeatedly paired with the drug of interest. There is an alternate environment that differs along some stimulus dimension(s) that is equally experienced, but not paired with the drug. Using this method of Pavlovian conditioning, the conditioned appetitive (rewarding) or aversive effects of a drug can be assessed. A conditioned place preference (CPP) is inferred when in a choice test animals spend more time in an environment that had been previously paired with a drug stimulus compared to an alternate environment. Such an outcome suggests that the drug has some rewarding effects that entered into an association with the paired environment. A conditioned place aversion (CPA) is inferred when animals spend less time in the drug-paired environment; this outcome is taken to indicate an aversive effect of the drug (see later for more detailed discussion).
Similar to other behavioral research, the parameters used in place conditioning studies with CB ligands vary widely across laboratories. With some drugs, these variations in procedural details across laboratories seem to make little difference in the overall conclusion regarding the motivational impact of the drug. As detailed in Tzschentke’s (3) excellent review of the place conditioning literature, studies with drugs such as cocaine or amphetamine consistently report CPP. The opiate drug heroin produces CPP, whereas the opioid antagonist naloxone consistently produces CPA. Alternatively, place conditioning literature involving the cannabinoid system seems to parallel the place conditioning literature with nicotine. That is, reports of no effect, CPA, and CPP with no clear answer yet as to the relevant conditions under which conditioned appetitive or aversive effects will be expressed. With this in mind, the purpose of the present review was to discuss the role of cannabinoids within the reward (motivation) system and to coalesce into one paper the published research on place conditioning with cannabinoid agonists. In doing so, we hoped to identify some critical variables that predict when a cannabinoid agonist may have conditioned appetitive versus conditioned aversive effect. Such information would be important for guiding future research attempting to identify the behavioral and neurochemical processes underlying the conditioned motivational effects of cannabinoid agonists.
2. Endogenous Cannabinoid System
2.1. Receptors
Cannabis has been used for thousands of years for its mood-altering, hallucinogenic, and anesthetic properties. The neurological effects of the drug suggested a central mechanism of action. Delta-9-tetrahydrocannabinol (Δ9-THC) was isolated as the primary psychoactive component of cannabis (4). This compound was then used to help elucidate the CB receptors (5). CB receptors have been divided into two groups, CB1 and CB2 receptors, on the basis of functionality and distribution. CB1 receptors are found widely throughout the brain and perform a variety of modulatory functions, whereas the CB2 receptors have generally been associated with the peripheral and central regulation of the immune system (6,7,8). Furthermore, recent evidence suggests non-CB receptor binding of endogenous (internally produced) and exogenous (externally produced) cannabinoid compounds (9,10). The current review will primarily focus on the CB1 receptor because of its purported role involving the rewarding and reinforcing effects of drugs.
The distribution of CB1 receptors on brain neurons in the striatum was first described using in vitro receptor autoradiography with the radioligand [3H]CP 55,940 (11). The receptors were initially found in high densities (quantities) in various striatal areas including the caudate putamen and the globus pallidus, as well as in the substantia nigra. The subsequent development of CB1 antibodies allowed for more specific cellular localization throughout the entire rat brain (12). Briefly, a high density of CB1 receptors was found in the hippocampus, cerebellum, striatum, and substantia nigra. Receptors in the olfactory bulb, piriform cortex, anterior part of the medial forebrain bundle, the cingulate cortex, amygdala, claustrum, and nucleus accumbens were found in moderate density. Finally, CB1 receptors were found in low density in the thalamus, hypothalamus, periaqueductal gray, pons, medulla, and the area postrema.
Importantly, receptor density does not necessarily indicate signaling strength. CB receptors are G-protein coupled (13), meaning the effects of receptor binding are mediated by the second messenger pathways activated by the G-protein. As such, binding can have a different impact depending on localization. For instance, using autoradiography and membrane saturation analyses in male Sprague−Dawley rats, the average number of G-proteins activated per bound CB1 receptor (i.e., amplification factor) was lowest in the frontal cortex, cerebellum, hippocampus, and striatum (14). These are regions with generally high numbers of receptors. Moderate amplification factors were found in the thalamus, brainstem, amygdala, and sensorimotor cortex. Finally, the hypothalamus had the highest amplification factor, an area with low receptor density. These data suggest that areas with low receptor density may enhance signal strength by increasing the impact of G-proteins on subsequent intracellular processes.
The location of these receptors on neurons is important for understanding receptor function. CB1 receptors have been found to be primarily localized presynaptically on GABAergic neurons as determined by electrophysiological analyses (15,16). This presynaptic localization suggests a modulatory role of endocannabinoids. Notably, G-protein activation by bound CB1 receptors reduces Ca2+ conductance (17) and increases K+ conductance (18). Both of these actions have been linked to a process known as depolarization-induced suppression of inhibition (19). As diagramed in Figure 1, activation of presynaptic CB1 receptors functions to inhibit subsequent neurotransmitter release from that presynaptic terminal (20). Postsynaptic endogenous cannabinoid release increases with increased postsynaptic excitation [21 see later]. The subsequent retrograde cannabinoid signaling can attenuate the release of GABA, resulting in less inhibitory input into the synapse and therefore further postsynaptic excitation.
Figure 1.
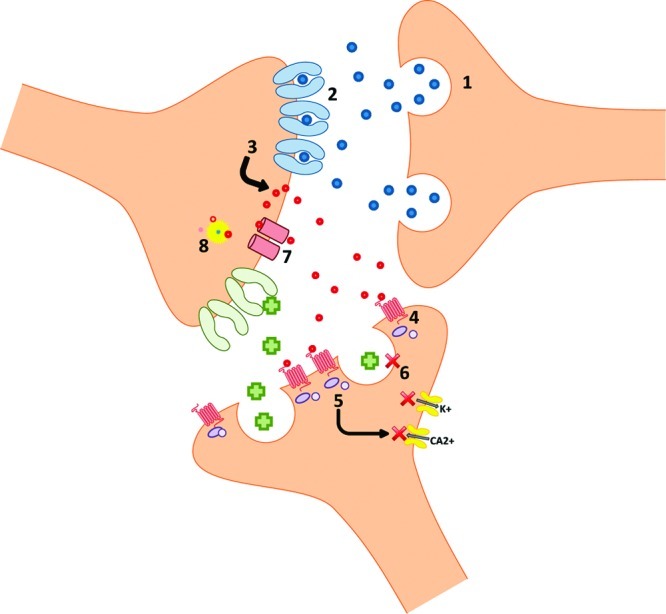
Cellular excitation causes the release of neurotransmitters (1). Neurotransmitters bind to receptors causing postsynaptic excitation (2) that triggers synthesis and nonvesicular release of endocannabinoids (3). Activation of presynaptically localized G-protein coupled cannabinoid receptors typically found on GABAergic neurons (4) signals a reduction in K+ and Ca2+ conductance (5), thereby inhibiting presynaptic GABA release (6). Inhibition of GABA release allows excitatory neurotransmitters to have a greater impact on the postsynaptic membrane. Cannabinoid transporters are responsible for reuptake of the cannabinoids in the synapse (7) where they are broken down by enzymes into their constituent parts (8). More details can be found in the text.
2.2. Ligands
Cannabinoidergic ligands found naturally in the body, including anandamide and 2-AG, are derived from arachidonic acid (22−25). As displayed in Figure 1, these molecules are synthesized and released in a Ca2+ dependent manner following membrane excitation (21,26). They are then taken back into the cell by anandamide or 2-AG transporters (26,27) and broken down by anandamide amidohydrolase (28) or fatty acid amide hydrolase (29) into constituent parts. These constituent parts can be synthesized back into the signaling molecules again when needed.
Experimentation with chemical analogues of Δ9-THC yielded a number of potent exogenous CB agonists, including CP 55,940, HU-210, and WIN 55,212-2 (30−35). Each of these agonists has since been used in behavioral and neurochemical research to help elucidate the localization and role of CB receptors in the brain. For instance, since Δ9-THC made a poor candidate for radiolabeling, the synthesis of a more potent, more stable ligand was required. CP 55,940 was just such a molecule, and radiolabeled CP 55,940 was instrumental in providing evidence of the CB receptor (5). CP 55,940 has also been used in behavioral assays such as drug discrimination (36−38). HU-210 has been useful in examining the stress response to drug withdrawal (39,40). In addition to a number of place conditioning studies, WIN 55,212-2 has been used to examine the role of cannabinoids in memory (41,42). Furthermore, an anandamide transport inhibitor, AM404, has also been used to examine the effect of increased synaptic availability of anandamide on behaviors ranging from place conditioning to cocaine self-administration (43−45).
3. Drug Reward Circuitry
3.1. Mesolimbic System
There are a number of excellent reviews of the reward system and addiction (46−49). As such, we will only provide a brief introduction of this system and refer the interested reader to these reviews for more detail. Although the mesolimbic dopamine pathway of the ventral striatum is now perceived as being a small portion of a much larger circuitry responsible for the transition into compulsive drug-taking behavior (50), the pathway is generally considered to be associated with the rewarding properties of drugs and the initiation of drug use (51). Drug administration and cues predictive of drug administration activate that system (52−57). The ventral tegmental area (VTA) has dopaminergic projections to the nucleus accumbens (NAcc) and to the prefrontal cortex. Increase in dopamine in the NAcc in particular has been associated with the rewarding effects of abused drugs because prevention of dopamine transmission in this region typically reduces drug self-administration [e.g. ref (58), but see ref (59)] and conditioned place preference (60,61). The increased dopamine can come about in a number of ways. For instance, stimulants such as methamphetamine, amphetamine, and cocaine block the reuptake of dopamine in the synapse, resulting in prolonged availability of the neurotransmitter (62). Nicotine functions at pre- and postsynaptic sites, modulating activation of the VTA both directly on dopaminergic neurons and by potentiating excitatory glutamate release (63,64). Additionally, dopaminergic neurons of the VTA receive inhibitory GABAergic input from interneurons in the VTA and from medium spiny neurons of the nucleus accumbens (65). Heroin and morphine activate VTA dopamine release into the NAcc via disinhibition of GABA transmission in the VTA (66). These GABA neurons are also the purported source for the effects of cannabinoid compounds on the mesolimbic dopamine system (see Figure 1).
3.2. Cannabinoid Modulation
Administration of Δ9-THC, CP 55,940, HU210, and WIN 55,212-2 dose-dependently enhanced the firing of dopamine neurons in the VTA (67−69) and increased dopamine concentration in the NAcc shell (70) in rat mesolimbic slices and in vivo. Further, administration of the CB1 antagonist SR 141716 (rimonabant) prevented these effects, indicating direct contribution of CB receptor activation in the dopamine enhancement (67,69−71). Similar to the effects of opiates, the mechanism of this increased dopamine release in the NAcc is considered to be modulatory rather than a direct effect on VTA dopamine neurons (72). As described earlier, CB1 receptors are localized presynaptically, typically on GABAergic neurons (16). Within the mesolimbic system, there are GABAergic inputs onto VTA dopaminergic neurons that project to the NAcc (73). Using a patch-clamp technique, administration of WIN 55,212-2 or CP 55,940 was shown to depress GABA-mediated inhibitory postsynaptic currents via a presynaptic inhibition of GABA release (74,75). This modulatory effect was due to the inhibition of Ca2+ and activation of K+ channels at presynaptic terminals attenuating further GABA release (17,18). The reduction of GABA-mediated inhibitory postsynaptic currents on VTA projections to the NAcc resulted in increased transmission of dopamine in midbrain slices that was also blocked by the administration of SR 141716 (75). For further discussion of the role of endocannabinoid modulation of neurotransmission with the mesolimbic system, we refer the reader to the following reviews in refs (76−79).
4. Place Conditioning
4.1. Typical Protocol
Environmental or situational cues that reliably co-occur with a drug can acquire the control of drug-related behaviors through Pavlovian conditioning processes (1,80). Researchers can capitalize on this associative learning to study the appetitive (rewarding) or aversive effects of a drug. As noted in the Introductory Remarks, one widely used task to do so is referred to as place conditioning (1−3,81). Although there are many variations of this method, in a typical place conditioning experiment, animals are exposed to two distinct contexts (e.g., variations in floor texture, wall color, odor, etc.); however, the drug is experienced only in one of the two contexts. When the animal is given a subsequent choice test with unrestricted access to both contexts in a drug-free state, the drug-paired context (conditioned stimulus; CS) now evokes either an approach (conditioned place preference; CPP) or avoidance (conditioned place aversion; CPA) conditioned response (CR) depending on the nature of the previously experienced drug effects (unconditioned stimulus; US).
4.2. Measurement Considerations
Determination of conditioning is typically based on an increase (i.e., CPP) or decrease (i.e., CPA) in the time spent in the paired context relative to the unpaired context at test, the paired context before conditioning (i.e., familiarization session), or to an untreated control. In an apparatus constructed with two compartments, not reporting time spent in the unpaired context is acceptable practice because time spent in the paired context necessarily subtracts from time spent in the unpaired context (82). However, many laboratories use three-compartment chambers, in which a center compartment distinct from the other two is used as a discrete starting place for choice tests (83). For some researchers, a benefit of having this third compartment is that it provides a novel context, thus detracting from a potential novelty-seeking account of increased time spent in the drug-paired compartment while in the nondrug state (1). However, there is some argument against the influence of this effect (2). In laboratories using three-compartment chambers, not reporting time spent in the unpaired context or a ratio measure that includes this time is problematic for interpretation. That is, an increase in time spent in the paired context may reflect less time spent in the center compartment rather than a shift from the unpaired to the paired compartment (84). If this occurred, there may still be greater time spent in the unpaired than in the paired compartment. Clearly, this is not a conditioned place preference, and no conclusion regarding the conditioned rewarding effects of the drug under study may be made.
4.3. Apparatus and Procedure Bias
When reading a paper on place conditioning, one should be aware of whether the apparatus was constructed in such a way as to bias the behavior of the animal. This bias in rats and mice can be accomplished by using stimuli such as a dark chamber that evokes approach behaviors. Alternatively, using bright illumination or widely space rod bars for flooring can evoke avoidance behaviors (85). An important issue with apparatus bias is that the unconditioned bias may shift with repeated exposures which can complicate interpretation of place conditioning results. That is, it becomes unclear whether the drug treatment, the apparatus exposure, or some combination of the two evoked the change in choice behavior. One solution several investigators have tried to solve this problem is to include a vehicle−vehicle control (86,87). This control allows one to determine how mere apparatus exposure shifts the unconditioned apparatus bias. Unfortunately, this control does not provide an assessment of how drug exposure might interact with these unconditioned biases still leaving some questions regarding the interpretation of any purported place conditioning effect (1,2).
Another important consideration is whether a biased conditioning procedure was used in the study. This bias refers to how and which compartment was assign to be paired with the drug. A biased procedure is being used if all subjects are assigned to receive the exact same compartment (all drug pairing in white side) or if subjects were assigned based on their behavior in a preconditioning choice test (all drug parings in the initially nonpreferred side). Using the initially preferred compartment could result in a ceiling effect, preventing detection of the development of the conditioned reward. Using the nonpreferred compartment as the paired context allows for detection of a change in compartment choice; however, the mechanism of that response becomes unclear. Is time spent in the paired compartment increasing because that compartment now had conditioned rewarding value due to the pairing of the drug effects or because the aversive qualities of the context were decreased by further familiarization? The attenuated aversion could be a result of unconditioned drug effects, but they could also have developed without the influence of the drug, thereby confounding the interpretation of the results. Another bias of the conditioning procedure often occurs when animals are given two conditioning sessions per day. In order to prevent lingering effects of (or possibly withdrawal from) the drug treatment, all animals receive vehicle for the first conditioning session and drug treatment for the second conditioning session of the day. This conditioning protocol confounds drug exposure and training order.
A complete discussion of the implications of procedure and apparatus bias in place conditioning studies is outside the scope of this review. Thus, we refer the reader to some articles that directly examine and/or review the issue in more detail (1−3,88,89). However, we would like to echo their general conclusion and ask the reader to keep these issues in mind for the following section that discusses the place conditioning literature with cannabinoids. That is, to obviate any interpretative issues, place conditioning research should use unbiased procedures and balanced apparatus construction.
5. Place Conditioning and Cannabinoid Compounds
Unlike classic psychomotor stimulants such as amphetamine or cocaine that readily condition a place preference at a number of doses and under a variety of conditions, cannabinoid agonists show more mixed results [3,81 see earlier]. The extant variation in parameters used by different laboratories to assess place conditioning with cannabinoidergic compounds complicates interpretation and hence conclusions regarding this system. Despite this difficulty, there are a number of experimental variables that appear to impact the outcome of some studies. These variables include the specific compound and its dose, number of conditioning trials, session length, injection-to-placement interval, and pretreatment. Table 1 provides details about Δ9-THC studies, and Table 2 provides details about other cannabinoid compounds. Figure 2 provides the chemical structures of each of these compounds. The remainder of this review will discuss each of these variables and the evidence for conditioned appetitive or aversive effects of these ligands.
Table 1. Δ9-THC Place Conditioning Study Details.
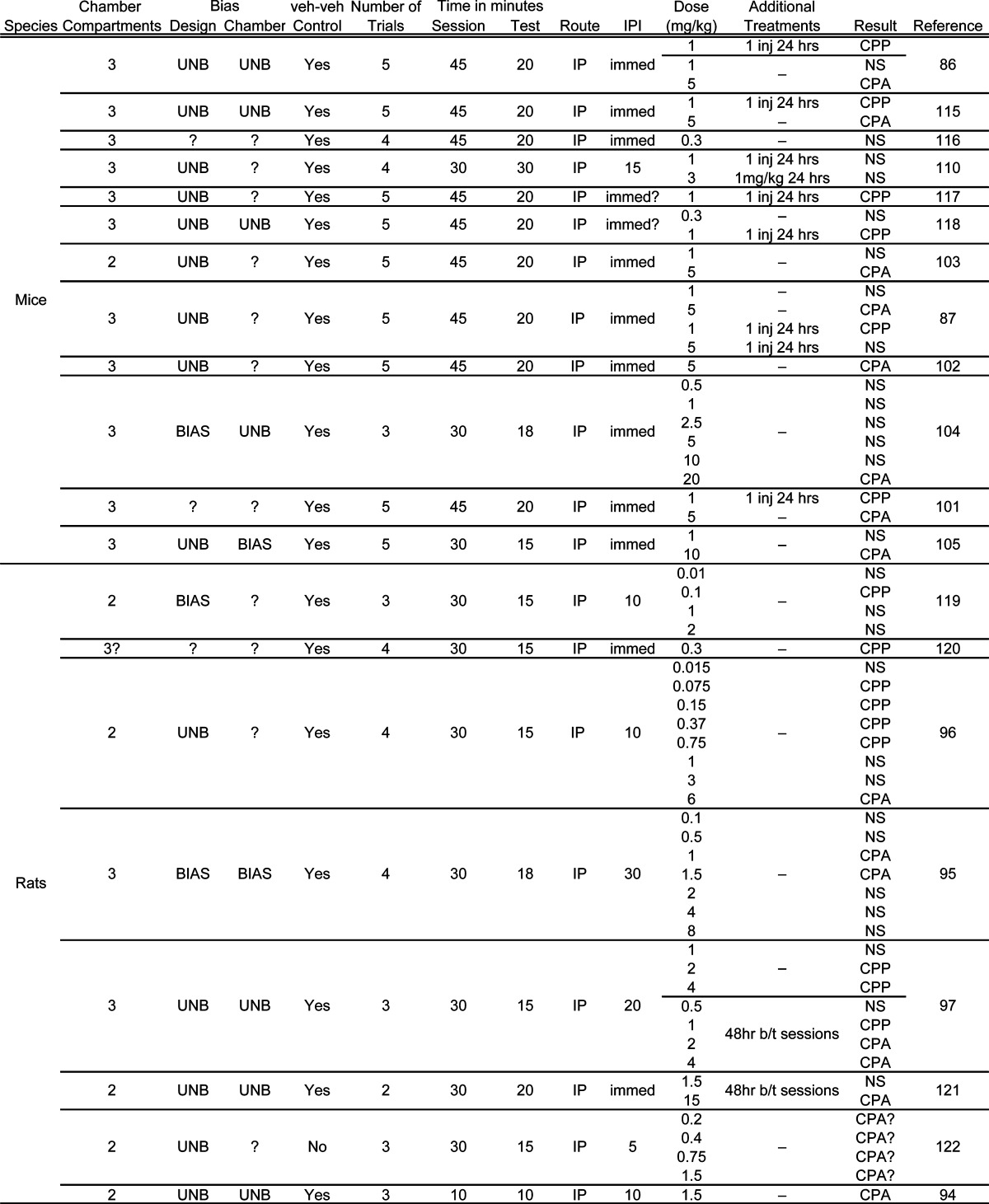 |
Table 2. CB Agonists and Transporter Inhibitor Place Conditioning Study Details.
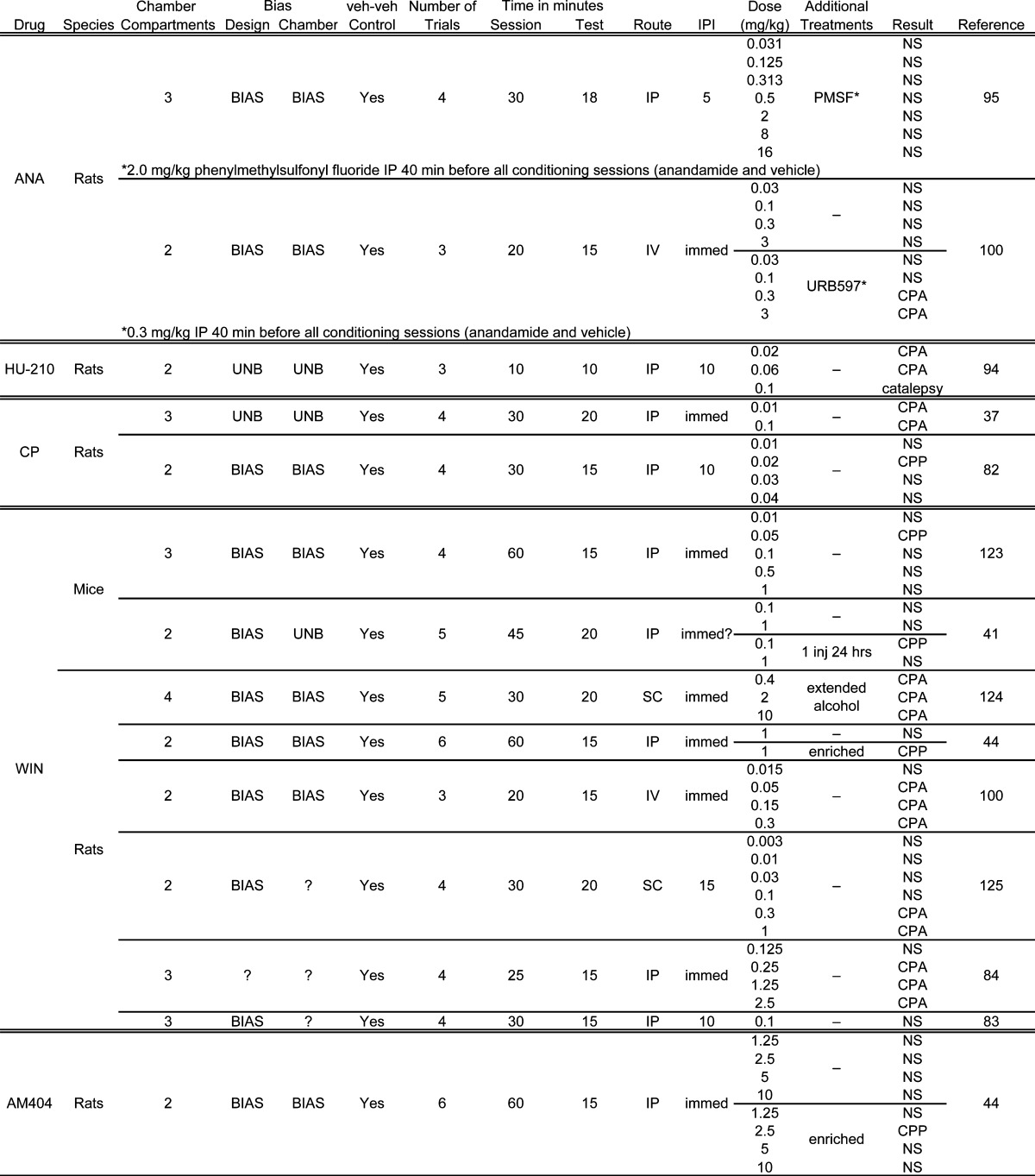 |
Figure 2.
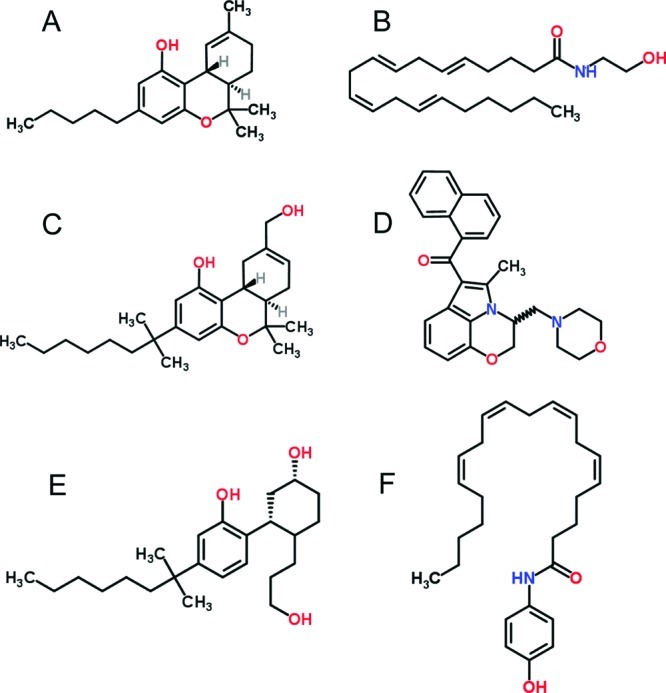
Structures of Δ9-THC (A), anandamide (B), HU-210 (C), WIN 55,212-2 (D), CP 55,940 (E), and AM404 (F).
5.1. Drug Specificity
Cannabinoid ligands have distinct specificity and potency. Initial interest in the cannabinoid system was prompted by the psychotropic effects of Δ9-THC. Subsequently developed synthetic cannabinoids are more efficacious at CB receptors than Δ9-THC, which is a partial agonist (90,91). The two primary endogenous cannabinoids in the brain, anandamide and 2-AG, have different efficacies at CB1 receptors. Anandamide is a partial agonist, whereas 2-AG is a full agonist (92,93). The synthetic cannabinoids HU-210 and WIN 55,212-2 are full agonists (90,92). CP 55,940 has been described as a full agonist (90) and as a high efficacy partial agonist (92) . Finally, the anandamide transport inhibitor, AM 404 selectively blocks the reuptake of anandamide, allowing the signal to last longer in the synapse and effectively serving as an agonist (43). Because of these differences in action, behavioral differences in their effects likely exist. As such, generalization of effects across compounds should not be made. Rather, it will be important to test these compounds within a given set of behavioral parameters if firm conclusions are to be made regarding the mechanism. For instance, Cheer et al. (94) found CPA in rats with both Δ9-THC and HU-210 using an unbiased apparatus and procedure. Bortolato et al. (44) found similar CPP effects using AM404 and WIN 55,212-2. Mallet and Beninger (95), however, found no effect of anandamide, yet Δ9-THC conditioned a place aversion.
5.2. Drug Dose
One of the primary considerations for behavioral research is the dose of the drug. Very low doses of Δ9-THC generally have limited rewarding effects and therefore often do not condition an approach response relative to the vehicle. Conversely, high doses appear to include an aversive quality often sufficient to compete with or overshadow any rewarding effects and thus condition an avoidance response. For instance, rats conditioned with 0.015 mg/kg Δ9-THC display choice behavior during testing indistinguishable from that of the vehicle (96). Doses from 0.075 to 0.75 mg/kg Δ9-THC, however, appear to condition a place preference; 1 and 3 mg/kg show no difference from vehicle, yet 6 mg/kg Δ9-THC conditions an aversion (96). Time between training sessions appears to interact with drug dose and has an impact on the conditioned effects of Δ9-THC. When rats were trained with 24 h between sessions, 1 mg/kg Δ9-THC had no effect, and 2 and 4 mg/kg Δ9-THC conditioned a place preference (97). However, when there were 48 h between sessions, 1 mg/kg conditioned a place preference, and 2 and 4 mg/kg Δ9-THC conditioned a place aversion. Thus, allowing a longer wash-out period between training sessions shifted the dose−effect curve to the left. The importance of this wash-out period to the motivational effects of Δ9-THC and the underlying processes deserve more empirical attention. Another consideration with regard to drug dose is the species of the subject. Mice generally have a higher rate of drug metabolism (98). As such, mice are typically treated with higher doses than rats. For instance, in place conditioning studies using Δ9-THC, mice were treated with a range of 0.3 to 20 mg/kg, whereas rats were treated with 0.01 to 8 mg/kg (see Table 1). The lowest effective dose (producing significant CPP or CPA) in mice was 1 mg/kg (86); in rats, that dose was more than ten times lower, 0.075 mg/kg (96).
5.3. Temporal Considerations
The temporal relationship between the drug effects and exposure to the to-be-paired compartment during the conditioning phase appears to be an important variable in the cannabinoid place conditioning literature. Variation across studies in this relationship between the CS (paired compartment) and the US (i.e., relevant drug effects) might help explain some of the discrepant results. In all conditioning tasks, there is a temporal window in which the US must occur for learning (i.e., CS−US association) to occur. Relationships in which the CS onset occurs before the US onset but with some overlap tends to be more conducive to conditioning than a relationship where the US occurs before or well after the CS (80). In the case of place conditioning research with drug USs, drugs have many physiological effects, and the duration and peak of these effects may differ. Because drug effects are variable due to pharmacokinetic and pharmacodynamic reasons, careful consideration should be given to such factors in the design of the experiment. For instance, if a compound is metabolized quite quickly (e.g., anandamide), then conditioning sessions should be shorter so that compartment exposure does not extend well beyond that of the drug effects (99). Allowing this may weaken conditioning through a process called extinction (80). Alternatively, an enzyme inhibitor can be used to slow ligand breakdown to extend the length of drug effects (95,100).
In addition, the more an US is paired with the CS, the stronger the conditioned association (80). The Maldonado group used this idea to their advantage by extending the time of typical conditioning. In their studies that examined 5 mg/kg Δ9-THC, mice acquired a CPA [86,87,101,102 see also (103)]. Each of these studies used 5 conditioning trials and 45-min sessions. However, when three 30-min sessions had been used by the group instead, there was no conditioned effect (104). This latter study had less time in the compartment on each placement, as well as fewer conditioning trials; both variables are important for learning. Although comparing across studies is fraught with difficulties, there are some basic ideas that can be extrapolated and suggest an important avenue for future parametric research.
Similar to the effects of session length, with repeated pairings the strength of the conditioned effect increases. Therefore, conditioning with only a few trials may not be sufficient to observe conditioning that would be expressed with a greater number of trials (i.e., environment−drug pairings). Again, taking the necessary caution of comparisons across studies, mice conditioned with Δ9-THC in five 30-min sessions developed a CPA at 10 mg/kg as shown in a 15-min test (105). However, mice only conditioned for three 30-min sessions did not show a conditioned effect at 10 mg/kg but did show a CPA at 20 mg/kg (104). This pattern suggests that the increased number of trials was required for conditioning to be sufficiently strong before being expressed as conditioned avoidance on the test day. With repeated drug exposure, there is also the potential for tolerance and sensitization to the drug effects. Tolerance refers to a gradual decrease in the effects of the drug; sensitization refers to an increase in the drug effects with repeated exposure. Sensitization and tolerance to the effects of Δ9-THC have been reported (106−109). Since increased pairings were required for the development of CPA, an alternative account suggests that sensitization of the aversive properties may have developed.
As the discussion in this section implies, the injection-to-placement interval (IPI) would have an impact on the extent of place conditioning. Thus, allowing time to pass following the injection of the drug establishes a different temporal relationship between the compartment CS and the drug US than immediate placement. Along these lines, CPA in rats developed at two doses of CP 55,940 using an immediate IPI (37), but an intermediate dose of CP 55,940 conditioned a place preference when a 10 min IPI was used (82). Furthermore, when rats had a longer IPI using Δ9-THC (95), the aversive properties of the drug were conditioned at lower doses than when rats had a shorter IPI (96). Perhaps the early effects of CP 55,940 are aversive and lengthening the IPI allows those effects to diminish before the start of conditioning. Conversely, perhaps the longer IPI with Δ9-THC allows the early rewarding effects to diminish, allowing conditioning to occur with the aversive effects of the drug. Of course, there were other variables that differed between these studies (i.e., chamber/placement bias and length of test session); therefore, a final conclusion will need to await the conduction of the appropriate parametric studies.
5.4. Pretreatment Effects
Drug pre-exposure purportedly attenuates the unconditioned aversive effects of the compound and hence reveals CPP upon subsequent training. For instance, mice given 1 mg/kg Δ9-THC in the home cage 24 h before beginning conditioning developed CPP, whereas mice not pretreated with Δ9-THC did not develop CPP or CPA (86,87,110). Furthermore, pretreatment can attenuate the aversive effects of Δ9-THC. Mice given 5 mg/kg Δ9-THC in the home cage before conditioning did not develop the CPA shown by mice not given pretreatment (87). These findings may be due to the development of some tolerance to initial aversive drug effects that are experienced in the home cage rather than in the conditioning chamber. Considering the consistent results of the Maldonado group, there is certainly clear support for the efficacy of pre-exposure attenuating some of the aversive properties of Δ9-THC in place conditioning studies.
6. Concluding Comments
Place conditioning is useful in determining the conditioned rewarding and/or aversive effects of cannabinergic compounds. However, as stated previously, there is much variability in the place conditioning apparatus and protocol across laboratories studying the conditioned appetitive or aversive effects of cannabinoids. As detailed in this review, such variability likely affects the outcome of the study and indicates a real need for careful parametric research on key factors that might affect conditioning with a particular ligand (dose, session length, number of trials, injection-to-placement interval, etc.). As recommended here and elsewhere (2,88,89), place conditioning studies should avoid procedural or apparatus biases. If achieved, then the field will be better able to advance and have a more coherent picture of the neurochemical system mediating the conditioned motivational effects of cannabinoids. Although the current review was focused on the rewarding and aversive properties of cannabinoids, this system has a much broader applicability. Cannabinoids have been shown to have a role in a variety of behavioral processes; these include pain (111), appetite, energy, nausea (112,113), and stress and mood (114). A better understanding of each of these effects and how they may interact with each other will no doubt benefit therapeutic outcomes across these areas.
Abbreviations
2-AG, 2-arachidonylglycerol; ?, uncertain/not stated; Δ9-THC, delta-9-tetrahydrocannabinol; ANA, anandamide; CB, cannabinoid; CP, CP 55,940; CPA, conditioned place aversion; CPP, conditioned place preference; CR, conditioned response; CS, conditional stimulus; GABA, gamma-aminobutyric acid; hrs, hours; immed, immediate; inj, injection; IP, intraperitoneal; IPI, injection-to-placement interval; IV, intravenous; NAcc, nucleus accumbens; NS, not significant; SC, subcutaneous; UNB, unbiased; veh, vehicle; VTA, ventral tegmental area; WIN, WIN 55,212-2.
While preparing the manuscript for publication, J.E.M. was supported by NIDA F31 DA025399 and MRC 9536855 (awarded to B. J. Everitt), and R.A.B. was partially supported by DA018114 and DA023591.
Funding Statement
National Institutes of Health, United States
References
- Bardo M. T.; Bevins R. A. (2000) Conditioned place preference: what does it add to our preclinical understanding of drug reward?. Psychopharmacology 153, 31–43. [DOI] [PubMed] [Google Scholar]
- Bevins R. A., and Cunningham C. L. (2006) Place Conditioning: A Methodological Analysis, in Tasks and Techniques: A Sampling of the Methodologies for the Investigation of Animal Learning, Behavior and Cognition (Anderson M. J., Ed.) pp 99−110, Nova Science Publishers, Inc., New York. [Google Scholar]
- Tzschentke T. M. (2007) Measuring reward with the conditioned place preference (CPP) paradigm: update of the last decade. Addict. Biol. 12, 227–462. [DOI] [PubMed] [Google Scholar]
- Gaoni Y.; Mechoulam R. (1964) Isolation, structure, and partial synthesis of an active constituent of hashish. J. Am. Chem. Soc. 86, 1646–1647. [Google Scholar]
- Devane W. A.; Dysarz F. A. III; Johnson M. R.; Melvin L. S.; Howlett A. C. (1988) Determination and characterization of a cannabinoid receptor in rat brain. Mol. Pharmacol. 34, 605–613. [PubMed] [Google Scholar]
- Van Sickle M. D.; Duncan M.; Kingsley P. J.; Mouihate A.; Urbani P.; Mackie K.; Stella N.; Makriyannis A.; Piomelli D.; Davison J. S.; Marnett L. J.; Di Marzo V.; Pittman Q. J.; Patel K. D.; Sharkey K. A. (2005) Identification and functional characterization of brainstem cannabinoid CB2 receptors. Science 310, 329–332. [DOI] [PubMed] [Google Scholar]
- Cabral G. A.; Dove Pettit D. A. (1998) Drugs and immunity: cannabinoids and their role in decreased resistance to infectious disease. J. Immunol. 83, 116–123. [DOI] [PubMed] [Google Scholar]
- Howlett A. C.; Breivogel C. S.; Childers S. R.; Deadwyler S. A.; Hampson R. E.; Porrino L. J. (2004) Cannabinoid physiology and pharmacology: 30 years of progress. Neuropharmacology 47(Suppl 1), 345–358. [DOI] [PubMed] [Google Scholar]
- De Petrocellis L., Di Marzo V. (2009) Non-CB1, non-CB2 receptors for endocannabinoids, plant cannabinoids, and synthetic cannabimimetics: Focus on G-protein-coupled receptors and transient receptor potential channels. J. Neuroimmune Pharmacol. 5, 103−121. [DOI] [PubMed] [Google Scholar]
- De Petrocellis L.; Cascio M. G.; Di Marzo V. (2004) The endocannabinoid systerm: a general view and latest additions. Br. J. Pharmacol. 141, 765–774. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Herkenham M.; Lynn A. B.; de Costa B. R.; Richfield E. K. (1991) Neuronal localization of cannabinoid receptors in the basal ganglia of the rat. Brain Res. 547, 267–274. [DOI] [PubMed] [Google Scholar]
- Moldrich G.; Wenger T. (2000) Localization of the CB1 cannabinoid receptor in the rat brain. An immunohistochemical study. Peptides 21, 1735–1742. [DOI] [PubMed] [Google Scholar]
- Howlett A. C. (1984) Inhibition of neuroblastoma adenylate cyclase by cannabinoid and nantradol compounds. Life Sci. 35, 1803–1810. [DOI] [PubMed] [Google Scholar]
- Breivogel C. S.; Sim L. J.; Childers S. R. (1997) Regional differences in cannabinoid receptor/G-protein coupling in rat brain. J. Pharmacol. Exp. Ther. 282, 1632–1642. [PubMed] [Google Scholar]
- Katona I.; Sperlagh B.; Sik A.; Kafalvi A.; Vizi E. S.; Mackie K.; Freund T. F. (1999) Presynaptically located CB1 cannabinoid receptors regulate GABA release from axon terminals of specific hippocampal interneurons. J. Neurosci. 19, 4544–4558. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Katona I.; Rancz E. A.; Acsady L.; Ledent C.; Mackie K.; Hajos N.; Freund T. F. (2001) Distribution of CB1 cannabinoid receptors in the amygdala and their role in the control of GABAergic transmission. J. Neurosci. 21, 9506–9518. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mackie K.; Hille B. (1992) Cannabinoids inhibit N-type calcium channels in neurobalstoma-glioma cells. Proc. Natl. Acad. Sci. U.S.A. 89, 3825–3829. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mackie K.; Lai Y.; Westenbroek R.; Mitchell R. (1995) Cannabinoids activate an inwardly rectifying potassium conductance and inhibit Q-type calcium currents in AtT20 cells transfected with rat brain cannabinoid receptor. J. Neurosci. 15, 6552–6561. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Alger B. E. (2002) Retrograde signaling in the regulation of synaptic transmission: focus on endocannabinoids. Prog. Neurobiol. 68, 247–286. [DOI] [PubMed] [Google Scholar]
- Hashimotodani Y.; Ohno-Shosaku T.; Maejima T.; Fukami K.; Kano M. (2008) Pharmacological evidence for the involvement of diacylglycerol lipase in depolarization-induced endocannabinoids release. Neuropharmacology 54, 58–67. [DOI] [PubMed] [Google Scholar]
- Adermark L.; Lovinger D. M. (2007) Retrograde endocannabinoids signaling at striatal synapses requires a regulated postsynaptic release step. Proc. Natl. Acad. Sci. U.S.A. 104, 20564–20569. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cadas H.; Gaillet S.; Beltramo M.; Venance L.; Piomelli D. (1996) Biosynthesis of an endogenous cannabinoid precursor in neurons and its control by calcium and cAMP. J. Neurosci. 16, 3934–3942. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Devane W. A.; Hanus L.; Breuer A.; Pertwee R. G.; Stevenson L. A.; Griffin G.; Gibson D.; Mandelbaum A.; Etinger A.; Mechaulam R. (1992) Isolation and structure of a brain constitutent that binds to the cannabinoid receptor. Science 258, 1946–1949. [DOI] [PubMed] [Google Scholar]
- Stella N.; Schweitzer P.; Piomelli D. (1997) A second endogenous cannabinoid that modulates long-term potentiation. Nature 388, 773–778. [DOI] [PubMed] [Google Scholar]
- Sugiura T.; Kondo S.; Sukagawa A.; Nakane S.; Shinoda A.; Itoh K.; Yamashita A.; Waku K. (1995) 2-Arachidonoylglycerol: a possible endogenous cannabinoid receptor ligand in brain. Biochem. Biophys. Res. Commun. 215, 89–97. [DOI] [PubMed] [Google Scholar]
- Di Marzo V.; Fontana A.; Cadas H.; Schinelli S.; Cimino G.; Schwartz J. C.; Piomelli D. (1994) Formation and inactivation of endogenous cannabinoid anandamide in central neurons. Nature 372, 686–691. [DOI] [PubMed] [Google Scholar]
- Bisogno T.; Maccarrone M.; De Petrocellis L.; Jarrahian A.; Finazzi-Agrò A.; Hillard C.; Di Marzo V. (2001) The uptake by cells of 2-arachidonoylglycerol, an endogenous agonist of cannabinoid receptors. Eur. J. Biochem. 268, 1982–1989. [DOI] [PubMed] [Google Scholar]
- Desarnaud F.; Cadas H.; Piomelli D. (1995) Anandamide amidohydrolase activity in rat brain microsomes. Identification and partial characterization. J. Biol. Chem. 270, 6030–6035. [DOI] [PubMed] [Google Scholar]
- Egertová M.; Giang D. K.; Cravatt B. F.; Elphick M. R. (1998) A new perspective on cannabinoid signaling: complementary localization of fatty acid amide hydrolase and the CB1 receptor in rat brain. Proc. R. Soc. London Biol. Sci. 265, 2081–2085. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Howlett A. C.; Champion T. M.; Wilken G. H.; Mechoulam R. (1990) Stereochemical effects of 11-OH-Δ8-tetrahydrocannabinoldimethylheptyl to inhibit aenylate cyclase and bind to the cannabinoid receptor. Neuropharmacology 29, 161–165. [DOI] [PubMed] [Google Scholar]
- Johnson M. R.; Melvin L. S.; Althuis T. H.; Bindra J. S.; Harbert C. A.; Milne G. M.; Weissman A. (1981) Selective and potent analgetics derived from cannabinoids. J. Clin. Pharmacol. 21, 271S–282S. [DOI] [PubMed] [Google Scholar]
- Little P. J.; Compton D. R.; Johnson M. R.; Melvin L. S.; Martin B. R. (1988) Pharmacology and stereoselectivity of structurally novel cannabinoids in mice. J. Pharmacol. Exp. Ther. 247, 1046–1051. [PubMed] [Google Scholar]
- Luk T.; Jin W.; Zvonok A.; Lu D.; Lin X.-Z.; Chavkin C.; Makriyannis A.; Mackie K. (2004) Identification of a potent and highly efficacious, yet slowly desensitizing CB1 cannabinoid receptor agonist. Br. J. Pharmacol. 142, 495–500. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pacheco M.; Childers S. R.; Arnold R.; Casiano F.; Ward S. J. (1991) Aminoalkylindoles: actions on specific G-protein-linked receptors. J. Pharmacol. Exp. Ther. 257, 170–183. [PubMed] [Google Scholar]
- Thomas B. F.; Gilliam A. F.; Burch D. F.; Roche M. J.; Seltzman H. H. (1998) Comparative receptor binding analyses of cannabinoid agonists and antagonists. J. Pharmacol. Exp. Ther. 285, 285–292. [PubMed] [Google Scholar]
- De Vry J.; Jentzsch K. R. (2003) Intrinsic activity estimation of cannabinoid CB1 receptor ligands in a drug discrimination paradigm. Behav. Pharmacol. 14, 471–476. [DOI] [PubMed] [Google Scholar]
- McGregor I. S.; Issakidis C. N.; Prior G. (1996) Aversive effects of the synthetic cannabinoid CP 55,940 in rats. Pharm. Biochem. Behav. 53, 657–664. [DOI] [PubMed] [Google Scholar]
- Murray J. E.; Wells N. R.; Lyford G. D.; Bevins R. A. (2009) Investigation of endocannabinoid modulation of conditioned responding evoked by a nicotine CS and the Pavlovian stimulus effects of CP 55,940 in adult male rats. Psychopharmacology 205, 655–665. [DOI] [PubMed] [Google Scholar]
- Mechoulam R.; Feigenbaum J. J.; Lander N.; Segal M.; Järbe T. U. C.; Hiltunen A. J.; Consroe P. (1988) Enantiomeric cannabinoids: stereospecificity of psychotropic activity. Experientia 44, 762–764. [DOI] [PubMed] [Google Scholar]
- Rodríguez de Fonseca F.; Carrera M. R. A.; Navarro M.; Koob G. F.; Weiss F. (1997) Activation of corticotropin-releasing factor in the limbic system during cannabinoid withdrawal. Science 276, 2050–2054. [DOI] [PubMed] [Google Scholar]
- Castañé A.; Maldonado R.; Valverde O. (2004) Role of different brain structures in the behavioural expression of WIN 55,212-2 withdrawal in mice . Br. J. Pharmacol. 142, 1309–1317. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hampson R. E.; Deadwyler S. A. (2000) Cannabinoids reveal the necessity of hippocampal neural encoding for short-term memory in rats. J. Neurosci. 20, 8932–8942. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Beltramo M.; Stella N.; Calignano A.; Lin S. Y.; Makriyannis A.; Piomelli D. (1997) Functional role of high-affinity anandamide transport, as revealed by selective inhibition. Science 277, 1094–1097. [DOI] [PubMed] [Google Scholar]
- Bortolato M.; Campolongo P.; Mangieri R. A.; Scattoni M. L.; Frau R.; Trezza V.; La Rana G.; Russo R.; Calignano A.; Gessa G. L.; Cuomo V.; Piomelli D. (2006) Anxiolytic-like properties of the anandamide transport inhibitor AM404. Neuropsychopharmacology 31, 2652–2659. [DOI] [PubMed] [Google Scholar]
- Vlachou S.; Stamatopoulou F.; Nomikos G. G.; Panagis G. (2008) Enhancement of endocannabinoid neurotransmission through CB1 cannabinoid receptors counteracts the reinforcing and psychostimulant effects of cocaine. Int. J. Neuropsychopharmacol. 11, 905–923. [DOI] [PubMed] [Google Scholar]
- Dalley J. W.; Everitt B. J. (2009) Dopamine receptors in the learning, memory and drug reward circuitry. Semin. Cell Dev. Biol. 20, 403–410. [DOI] [PubMed] [Google Scholar]
- Di Chiara G.; Bassareo V.; Fenu S.; De Luca M. A.; Spina L.; Cadoni C.; Acquas E.; Carboni E.; Valentini V.; Lecca D. (2004) Dopamine and drug addiction: the nucleus accumbens shell connection. Neuropharmacology 47, 227–241. [DOI] [PubMed] [Google Scholar]
- Hyman S. E.; Malenka R. C.; Nestler E. J. (2006) Neural mechanisms of addiction: the role of reward-related learning and memory. Annu. Rev. Neurosci. 29, 565–598. [DOI] [PubMed] [Google Scholar]
- Wise R. A. (2005) Forebrain substrates of reward and motivation. J. Comp. Neurol. 493, 115–121. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Everitt B. J.; Robbins T. W. (2005) Neural systems of reinforcement for drug addiction: from actions to habits to compulsion. Nat. Neurosci. 8, 1481–1487. [DOI] [PubMed] [Google Scholar]
- Wise R. A.; Rompre P.-P. (1989) Brain dopamine and reward. Annu. Rev. Psychol. 40, 191–225. [DOI] [PubMed] [Google Scholar]
- Carelli R. M.; Ijames S. G. (2000) Nucleus accumbens cell firing during maintenance, extinction, and reinstatement of cocaine self-administration behavior in rats. Brain Res. 866, 44–54. [DOI] [PubMed] [Google Scholar]
- Di Chiara G.; Imperato A. (1988) Drugs abused by humans preferentially increase synaptic dopamine concentrations in the mesolimbic system of freely moving rats. Proc. Natl. Acad. Sci. U.S.A. 85, 5274–5278. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Duvauchelle C. L.; Ikegami A.; Castaneda E. (2000) Conditioned increases in behavioral activity and accumbens dopamine levels produced by intravenous cocaine. Behav. Neurosci. 114, 1156–1166. [DOI] [PubMed] [Google Scholar]
- Schultz W.; Dayan P.; Montague P. R. (1997) A neural substrate of prediction and reward. Science 275, 1593–1599. [DOI] [PubMed] [Google Scholar]
- Sombers L. A.; Beyene M.; Carelli R. M.; Wightman R. M. (2009) Synaptic overflow of dopamine in the nucleus accumbens arises from neuronal activity in the ventral tegmental area. J. Neurosci. 11, 1735–1742. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Weiss F.; Lorang M. T.; Bloom F. E.; Koob G. F. (1993) Oral alcohol self-administration stimulates dopamine release in the rat nucleus accumbens: genetic and motivational determinants. J. Pharmacol. Exp. Ther. 267, 250–258. [PubMed] [Google Scholar]
- Caine S. B.; Koob G. F. (1994) Effects of mesolimbic dopamine depletion on responding maintained by cocaine and food. J. Exp. Anal. Behav. 61, 213–221. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pettit H. O.; Ettenberg A.; Bloom F. E.; Koob G. F. (1984) Destruction of dopamine in the nucleus accumbens selectively attenuates cocaine but not heroin self-administration in rats. Psychopharmacology 84, 167–173. [DOI] [PubMed] [Google Scholar]
- Gremel C. M.; Cunningham C. L. (2008) Roles of the nucleus accumbens and amygdale in the acquisition and expression of ethanol-conditioned behavior in mice. J. Neurosci. 28, 1076–1084. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sellings L. H. L.; Clarke P. B. S. (2003) Segregation of amphetamine reward and locomotor stimulation between nucleus accumbens medial shell and core. J. Neurosci. 23, 6295–6303. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fleckenstein A. E.; Gibb J. W.; Hanson G. R. (2000) Differential effects of stimulants on monoaminergic transporters: pharmacological consequences and implications for neurotoxicity. Eur. J. Pharmacol. 406, 1–13. [DOI] [PubMed] [Google Scholar]
- Dani J. A.; De Biasi M. (2001) Cellular mechanisms of nicotine addiction. Pharmacol., Biochem. Behav. 70, 439–446. [DOI] [PubMed] [Google Scholar]
- Placzek A. N., and Dani J. A. (2008) Synaptic Plasticity within Midbrain Dopamine Centers Contributes to Nicotine Addiction, in Nebraska Symposium on Motivation, Volume 55: The Motivational Impact of Nicotine and its Role in Tobacco Use (Bevins R. A, and Caggiula A. R, Eds.), pp 5−15, Springer, New York. [DOI] [PubMed] [Google Scholar]
- Spanagel R.; Weiss F. (1999) The dopamine hypothesis of reward: past and current status. Trends Neurosci. 22, 521–527. [DOI] [PubMed] [Google Scholar]
- Shippenberg T. S.; LeFevour A.; Chefer V. I. (2008) Targeting endogenous mu- and delta-opioid receptor systems for the treatment of drug addiction. CNS Neurol, Disord.: Drug Targets 7, 442–453. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cheer J. F.; Marsden C. A.; Kendall D. A.; Mason R. (2000) Lack of response suppression follows repeated ventral tegmental cannabinoid administration: an in vitro electrophysiological study. Neuroscience 99, 661–667. [DOI] [PubMed] [Google Scholar]
- French E. D.; Dillon K.; Wu X. (1997) Cannabinoids excite dopamine neurons in the ventral tegmentum and substantia nigra. NeuroReport 8, 649–652. [DOI] [PubMed] [Google Scholar]
- Gessa G. L.; Melis M.; Muntoni A. L.; Diana M. (1998) Cannabinoids activate mesolimbic dopamine neurons by an action on cannabinoid CB1 receptors. Eur. J. Pharmacol. 341, 39–44. [DOI] [PubMed] [Google Scholar]
- Tanda G.; Pontieri F. E.; Di Chiara G. (1997) Cannabinoid and heroin activation of mesolimbic dopamine transmission by a common μ1 opioid receptor mechanism. Science 276, 2048–2050. [DOI] [PubMed] [Google Scholar]
- French E. D. (1997) Δ9-Tetrahydrocannabinol excites rat VTA dopamine neurons through activation of cannabinoid CB1 but not opioid receptors. Neurosci. Lett. 226, 159–162. [DOI] [PubMed] [Google Scholar]
- Schlicker E.; Kathmann M. (2001) Modulation of transmitter release via presynaptic cannabinoid receptors. Trends Pharmacol. Sci. 22, 565–572. [DOI] [PubMed] [Google Scholar]
- Matsuda L. A.; Bonner T. I.; Lolait S. J. (1993) Localization of cannabinoid receptor mRNA in rat brain. J. Comp. Neurol. 327, 535–550. [DOI] [PubMed] [Google Scholar]
- Manzoni O. J.; Bockaert J. (2001) Cannabinoids inhibit GABAergic synaptic transmission in mice nucleus accumbens. Eur. J. Pharmacol. 412, R3–R5. [DOI] [PubMed] [Google Scholar]
- Szabo B.; Siemes S.; Wallmichrath I. (2002) Inhibition of GABAergic neurotransmission in the ventral tegmental area by cannabinoids. Eur. J. Neurosci. 15, 2057–2061. [DOI] [PubMed] [Google Scholar]
- Gardner E. L. (2002) Addictive potential of cannabinoids: the underlying neurobiology. Chem. Phys. Lipids 121, 267–290. [DOI] [PubMed] [Google Scholar]
- Gardner E. L.; Vorel S. R. (1998) Cannabinoid transmission and reward-related events. Neurobiol. Dis. 5, 502–533. [DOI] [PubMed] [Google Scholar]
- Lupica C. R.; Riegel A. C. (2005) Endocannabinoid release from midbrain dopamine neurons: a potential substrate for cannabinoid receptor antagonist treatment of addiction. Neuropharmacology 48, 1105–1116. [DOI] [PubMed] [Google Scholar]
- Maldonado R.; Valverde O.; Berrendero F. (2006) Involvement of the endocannabinoid system in drug addiction. Trends Neurosci. 29, 225–232. [DOI] [PubMed] [Google Scholar]
- Pavlov I. P. (1927) Conditioned Reflexes, Oxford University Press, London. [Google Scholar]
- Tzschentke T. M. (1998) Measuring reward with the conditioned place preference paradigm: a comprehensive review of drug effects, recent progress and new issues. Prog. Neurobiol. 56, 613–672. [DOI] [PubMed] [Google Scholar]
- Braida D.; Pozzi M.; Cavallini R.; Sala M. (2001) Conditioned place preference induced by the cannabinoid agonist CP 55,940: interaction with the opioid system. Neuroscience 104, 923–926. [DOI] [PubMed] [Google Scholar]
- Polissidis A.; Chouliara O.; Galanopoulos A.; Marselos M.; Papadopoulou-Daifoti Z.; Antoniou K. (2009) Behavioural and dopaminergic alterations induced by a low dose of WIN 55,212-2 in a conditioned place preference procedure. Life Sci. 85, 248–254. [DOI] [PubMed] [Google Scholar]
- Pandolfo P.; Vendruscolo L. F.; Sordi R.; Takahashi R. N. (2009) Cannabinoid-induced conditioned place preference in the spontaneously hypertensive rat: an animal model of attention deficit hyperactivity disorder. Psychopharmacology 205, 319–326. [DOI] [PubMed] [Google Scholar]
- Allison J.; Larson D.; Jensen D. D. (1967) Acquired fear, brightness preference, and one-way shuttlebox performance. Psychonomic Sci. 8, 269–270. [Google Scholar]
- Ghozland S.; Matthews H. W. D.; Simonin F.; Filliol D.; Kieffer B. L.; Maldonado R. (2002) Motivational effects of cannabinoids are mediated by μ-opioid and κ-opioid receptors. J. Neurosci. 22, 1146–1154. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Valjent E.; Maldonado R. (2000) A behavioural model to reveal place preference to Δ9-tetrahydrocannabinol in mice. Psychopharmacology 147, 436–438. [DOI] [PubMed] [Google Scholar]
- Cunningham C. L.; Ferree N. K.; Howard M. A. (2003) Apparatus bias and place conditioning with ethanol in mice. Psychopharmacology 170, 409–422. [DOI] [PubMed] [Google Scholar]
- Roma P. G.; Riley A. L. (2005) Apparatus bias and the use of light and texture in place conditioning. Pharmacol., Biochem. Behav. 82, 163–169. [DOI] [PubMed] [Google Scholar]
- Burkey T. H.; Quock R. M.; Consroe P.; Ehlert F. J.; Hosohata Y.; Roeske W. R.; Yamamura H. I. (1997) Relative efficacies of cannabinoid CB1 receptor agonists in the mouse brain. Eur. J. Pharmacol. 336, 295–298. [DOI] [PubMed] [Google Scholar]
- Sim L. J.; Hampson R. E.; Deadwyler S. A.; Childers S. R. (1996) Effects of chronic treatment with Δ9-tetrahydrocannabinol on cannabinoid-stimulated [35S]GTPγS autoradiography in rat brain. J. Neurosci. 16, 8057–8066. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Breivogel C. S.; Selley D. E.; Childers S. R. (1998) Cannabinoid receptor agonist efficacy for stimulating [35S]GTPγS binding to rat cerebellar membranes correlates with agonist-induced decreases in GDP affinity. J. Biol. Chem. 273, 16865–16873. [DOI] [PubMed] [Google Scholar]
- Hillard C. J. (2000) Biochemistry and pharmacology of the endocannabinoids arachidonylethanolamide and 2-arachidonylglycerol. Prostaglandins Other Lipid Mediators 61, 3–18. [DOI] [PubMed] [Google Scholar]
- Cheer J. F.; Kendall D. A.; Mardsen C. A. (2000) Cannabinoid receptors and reward in the rat: a conditioned place preference study. Psychopharmacology 151, 25–30. [DOI] [PubMed] [Google Scholar]
- Mallet P. E.; Beninger R. J. (1998) Δ9-tetrahydrocannabinol, but not the endogenous cannabinoid receptor ligand anandamide, produces conditioned place avoidance. Life Sci. 62, 2431–2439. [DOI] [PubMed] [Google Scholar]
- Braida D.; Iosuè S.; Pegorini S.; Sala M (2004) Δ9-tetrahydrocannabinol-induced conditioned place preference and intracerebroventricular self-administration in rats. Eur. J. Pharmacol. 506, 63–69. [DOI] [PubMed] [Google Scholar]
- Lepore M.; Vorel S. R.; Lowinson J.; Gardner E. L. (1995) Conditioned place preference induced by Δ9-tetrahydrocannabinol: comparison with cocaine, morphine, and food reward. Life Sci. 56, 2073–2080. [DOI] [PubMed] [Google Scholar]
- Lemberger L.; McMahon R. E.; Archer R. A.; Matsumoto K.; Rowe H. (1973) The in vitro and in vivo metabolism of Δ6a, 10a dimethyl heptyl tetrahydrocannabinol (DMHP). J. Pharmacol. Exp. Ther. 187, 169–175. [PubMed] [Google Scholar]
- Romero J.; García-Palomero E.; Lin S. Y.; Ramos J. A.; Makriyannis A.; Fernández-Ruiz J. J. (1996) Extrapyramidal effects of methanandamide, an analog of anandamide, the endogenous CB1 receptor ligand. Life Sci. 58, 1249–1257. [DOI] [PubMed] [Google Scholar]
- Scherma M.; Medalie J.; Fratta W.; Vadivel S. K.; Makriyannis A.; Piomelli D.; Mikics E.; Haller J.; Yasar S.; Tanda G.; Goldberg S. R. (2008) The endogenous cannabinoid anandamide has effects on motivation and anxiety that are revealed by fatty acid amide hydrolase (FAAH) inhibition. Neuropharmacology 54, 129–140. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Soria G.; Castañé A.; Berrendero F.; Ledent C.; Parmentier M.; Maldonado R.; Valverde O. (2004) Adenosine A2A receptors are involved in physical dependence and place conditioning induced by Δ9-THC. Eur. J. Neurosci. 20, 2203–2213. [DOI] [PubMed] [Google Scholar]
- Zimmer A.; Valjent E.; König M.; Zimmer A. M.; Robledo P.; Hahn H.; Valverde O.; Maldonado R. (2001) Absence of Δ-9-tetrahydrocannabinol dysphoric effects on dynorphin-deficient mice. J. Neurosci. 21, 9499–9505. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cheng H.-Y. M.; Laviolette S. R.; van der Kooy D.; Penninger J. M. (2004) DREAM ablation selectively alters Δ9-THC place aversion and analgesia but leaves intact the motivational and analgesic effects of morphine. Eur. J. Neurosci. 19, 3033–3041. [DOI] [PubMed] [Google Scholar]
- Hutcheson D. M.; Tzavara E. T.; Smadja C.; Valjent E.; Roques B. P.; Hanoune J.; Maldonado R. (1998) Behavioural and biochemical evidence for signs of abstinence in mice chronically treated with Δ-9-tetrahydrocannabinol. Br. J. Pharmacol. 125, 1567–1577. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Vann R. E.; Gamage T. F.; Warner J. A.; Marshall E. M.; Taylor N. L.; Martin B. R.; Wiley J. L. (2008) Divergent effects of cannabidiol on the discriminative stimulus and place conditioning effects of Δ9-tetrahydrocannabinol. Drug Alcohol Depend. 94, 191–198. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Barnes C.; Fried P. A. (1974) Tolerance to Δ9-THC in adult rats with differential Δ9-THC exposure when immature or during early adulthood. Psychopharmacologia 34, 181–190. [DOI] [PubMed] [Google Scholar]
- Lamb R. J.; Järbe T. U. C.; Makriyannis A.; Lin S.; Goutopoulos A. (2000) Effects of Δ9-tetrahydrocannabinol, (R)-methanandamide, SR 141716, and d-amphetamine before and during daily Δ9-tetrahydrocannabinol dosing. Eur. J. Pharmacol. 398, 251–258. [DOI] [PubMed] [Google Scholar]
- Rubino T.; Viganò D.; Massi P.; Parolaro D. (2003) Cellular mechanisms of Δ9-tetrahydrocannabinol behavioural sensitization. Eur. J. Neurosci. 17, 325–330. [DOI] [PubMed] [Google Scholar]
- Wiley J. L.; Evans R. L.; Grainger D. B.; Nicholson K. L. (2008) Age-dependent differences in sensitivity and sensitization to cannabinoids and ‘club drugs’ in male adolescent and adult rats. Addict. Biol. 13, 277–286. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Vlachou S.; Nomikos G. G.; Stephens D. N.; Panagis G. (2007) Lack of evidence for appetitive effects of Δ9-tetrahydrocannabinol in the intracranial self-stimulation and conditioned place preference procedures in rodents. Behav. Pharmacol. 18, 311–319. [DOI] [PubMed] [Google Scholar]
- Sagar D. R., Gaw A. G., Okine B. N., Woodhams S. G., Wong A., Kendall D. A., Chapman V. (2009) Dynamic regulation of the endocannabinoid system: implications for analgesia. Mol. Pain [Online eraly access] DOI: 10.1186/1744-8069-5-59. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cavuoto P.; Wittert G. A. (2009) The role of the endocannabinoid system in the regulation of energy expenditure. Best Pract. Res. Clin. Endocrinol. Metab. 23, 79–86. [DOI] [PubMed] [Google Scholar]
- Slatkin N. E. (2007) Cannabinoids in the treatment of chemotherapy-induced nausea and vomiting: beyond prevention of acute emesis. J. Support. Oncol. 5, 1–9. [PubMed] [Google Scholar]
- Hill M. N.; Hillard C. J.; Bambico F. R.; Patel S.; Gorzalka B. B.; Gobbi G. (2009) The therapeutic potential of the endocannabinoid system for the development of a novel class of antidepressants. Trends Pharmacol. Sci. 30, 484–493. [DOI] [PubMed] [Google Scholar]
- Célérier E.; Ahdepil T.; Wikander H.; Berrendero F.; Nyberg F.; Maldonado R. (2006) Influence of the anabolic-androgenic steroid nandrolone on cannabinoid dependence. Neuropharmacology 50, 788–806. [DOI] [PubMed] [Google Scholar]
- Robledo P.; Trigo J. M.; Panayi F.; de la Torre R.; Maldonado R. (2007) Behavioural and neurochemical effects of combined MDMA and Δ9-THC administration in mice. Psychopharmacology 195, 255–264. [DOI] [PubMed] [Google Scholar]
- Valjent E.; Pagès C.; Rogard M.; Besson M.-J.; Maldonado R.; Caboche J. (2001) Δ9-tetrahydrocannabinol-induced MAPK/ERK and Elk-1 activation in vivo depends on dopaminergic transmission. Eur. J. Neurosci. 14, 342–352. [DOI] [PubMed] [Google Scholar]
- Valjent E.; Mitchell J. M.; Besson M.-J.; Caboche J.; Maldonado R. (2002) Behavioural and biochemical evidence for interactions between Δ9-tetrahydrocannabinol and nicotine. Br. J. Pharmacol. 135, 564–578. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Le Foll B.; Wiggins M.; Goldberg S. R. (2006) Nicotine pre-exposure does not potentiate the locomotor or rewarding effects of Δ-9-tetrahydrocannabinol in rats. Behav. Pharm. 17, 195–199. [DOI] [PubMed] [Google Scholar]
- Ji S.-P.; Zhang Y.; Cleemput J. V.; Jiang W.; Liao M.; Li L.; Wan Q.; Backstrom J. R.; Zhang X. (2006) Disruption of PTEN coupling with 5-HT2C receptors suppresses behavioral responses induced by drugs of abuse. Nat. Med. 12, 324–329. [DOI] [PubMed] [Google Scholar]
- Sañudo-Peña M. C.; Tsou K.; Delay E. R.; Hohman A. G.; Force M.; Walker J. M. (1997) Endogenous cannabinoids as an aversive or counter-rewarding system in the rat. Neurosci. Lett. 223, 125–128. [DOI] [PubMed] [Google Scholar]
- Parker L. A.; Gillies T. (1995) Δ9-THC-induced place and taste aversions in Lewis and Sprague-Dawley rats. Behav. Neurosci. 109, 71–78. [DOI] [PubMed] [Google Scholar]
- Manzanedo C.; Aguilar M. A.; Rodriguez-Arias M.; Navarro M.; Miñarro J. (2004) Cannabinoid agonist-induced sensitization to morphine place preference in mice. NeuroReport 15, 1373–1377. [DOI] [PubMed] [Google Scholar]
- López-Moreno J. A.; González-Cuevas G.; Rodríguez de Fonseca F.; Navarro M. (2004) Long-lasting increase of alcohol relapse by the cannabinoid receptor agonist WIN 55,212-2 during alcohol deprivation. J. Neurosci. 24, 8245–8252. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chaperon F.; Soubrié P.; Puech A. J.; Thiébot M.-H. (1998) Involvement of central cannabinoid (CB1) receptors in the establishment of place conditioning in rats. Psychopharmacology 135, 324–332. [DOI] [PubMed] [Google Scholar]