

Abstract
TNF-like protein 1A (TL1A), a TNF superfamily cytokine that binds to death receptor 3 (DR3), is highly expressed in macrophage foam cell-rich regions of atherosclerotic plaques, although its role in foam cell formation has yet to be elucidated. We investigated whether TL1A can directly stimulate macrophage foam cell formation in both THP-1 and primary human monocyte-derived macrophages with the underlying mechanisms involved. We demonstrated that TL1A promotes foam cell formation in human macrophages in vitro by increasing both acetylated and oxidized low-density lipoprotein uptake, by enhancing intracellular total and esterified cholesterol levels and reducing cholesterol efflux. This imbalance in cholesterol homeostasis is orchestrated by TL1A-mediated changes in the mRNA and protein expression of several genes implicated in the uptake and efflux of cholesterol, such as scavenger receptor A and ATP-binding cassette transporter A1. Furthermore, through the use of virally delivered DR3 short-hairpin RNA and bone marrow-derived macrophages from DR3 knockout mice, we demonstrate that DR3 can regulate foam cell formation and contributes significantly to the action of TL1A in this process in vitro. We show, for the first time, a novel proatherogenic role for both TL1A and DR3 that implicates this pathway as a target for the therapeutic intervention of atherosclerosis.
Atherosclerosis is a progressive, chronic inflammatory disease of the vasculature that is characterized by the formation of fibrotic plaques in the major arteries (1, 2). The chronic inflammation that is observed in atherosclerosis is believed to be a direct result of immune cells, such as monocytes and T lymphocytes, being continually recruited to atherosclerotic lesions. Macrophages are capable of ingesting modified forms of low-density lipoprotein (LDL) to form cholesterol-rich foam cells, which become the initial building blocks of atherosclerotic plaques (1, 2). Cytokines, such as IFN-γ and TNF-α, are known to be master regulators of immune cell recruitment to atherosclerotic plaques and can also modulate macrophage foam cell formation (3). The role of TNFR superfamily (TNFRSF) members and their ligands, such as TNF-α/TNFR1, in promoting atherosclerosis has been shown (4) and supports the notion that this cytokine super-family strongly contributes to atherogenesis.
Death receptor 3 (DR3; TNFRSF25) is a death-domain containing TNFRSF member, closely homologous to TNFR1, which can regulate cell apoptosis and survival (5). DR3 is expressed on a range of cell types, including lymphocytes, NK cells, endothelial cells, and macrophages (5-7), which are all involved in atherosclerosis. DR3 has been shown to modulate immune cell function (7-10) and to promote many inflammatory-driven diseases, including inflammatory bowel disease and arthritis (5, 7, 10-14). The only confirmed ligand for DR3 is TNF-like protein 1A (TL1A) (8), which is expressed on immune cells like macrophages and dendritic cells (5, 6, 11) and can also drive inflammatory disease progression (5, 7, 9, 10, 12-14). Both TL1A and DR3 have been implicated as mediators of atherosclerosis since they have been found localized in foam cell rich regions of atherosclerotic plaques (6). However, the role of TL1A–DR3 pathway in macrophage foam cell formation has yet to be reported. In this study, we set out to determine whether TL1A can stimulate macrophage foam cell formation in both THP-1 and primary human monocyte-derived macrophages (HMDMs).
Materials and Methods
Reagents and cell culture
Human THP-1 and mouse bone-marrow macrophage (BMM) cell cultures were maintained in RPMI 1640 or DMEM medium, respectively, supplemented with 10% (v/v) heat-inactivated FCS, 100 U/ml penicillin, 100 μg/ml streptomycin, and 2 mM L-glutamine (all Invitrogen, Paisley, U.K.), at 37°C in a humidified atmosphere containing 5% (v/v) CO2. HMDMs were isolated from buffy coats using Ficoll-Hypaque purification. Blood from the buffy coat was layered over Lymphoprep (Nycomed, Oslo, Norway) and centrifuged at 800 × g for 30 min. The resultant mononuclear cell interface was collected and washed six to eight times in PBS containing 0.4% (v/v) trisodium citrate to remove platelets. Monocytes were allowed to adhere to tissue culture plates in RPMI 1640 medium containing 5% (v/v) FCS for 7 d to enable macrophage differentiation and permit removal of nonadherent lymphocytes. Recombinant human IFN-γ and TL1A were acquired from PeproTech (London, U.K.) and R&D Systems (Abingdon, U.K.), respectively. PMA was acquired from Sigma-Aldrich (Poole, U.K.). Recombinant human sDR3-Fc and human NKG2D-Fc chimeras were also from R&D Systems.
Mice
The DR3 mouse colony was founded from animals supplied by Cancer Research U.K., London. BMMs were isolated from 6–8-wk-old female wild type (WT; DR3WT) and knockout (KO; DR3KO) mice (15) as previously described (16). All procedures were approved by the Local Research Ethics Committee and performed in strict accordance with the Home Office, approved licenses PPL 30/1999 and 30/2580.
DiI-acetylated LDL and DiI-Ox LDL uptake assays
Cells were incubated for 24 h with 10 μg/ml 1,1′-dioctadecyl-3,3,3′,3′-tetramethyllindocarbocyane (DiI)-labeled acetylated LDL (DiI-AcLDL) or oxidized LDL (DiI-OxLDL; Intracel, Frederick, MD) in RPMI 1640 or DMEM (mouse BMMs) containing 10% (v/v) delipidated FCS at 37°C. Next, cells were released from the tissue culture plate by trypsinization and DiI-AcLDL or DiI-OxLDL uptake was then analyzed by flow cytometry on a FACS Canto (BD Biosciences, Oxford, U.K.) flow cytometer. DiI-AcLDL or DiI-OxLDL uptake is represented as a percentage, with the untreated control indicated as 100%.
Cholesterol efflux assay
Macrophages (THP-1, HMDMs, and mouse BMMs) were converted into foam cells by incubation with 50 μg/ml AcLDL (micrograms of AcLDL protein per milliliter; Intracel) in media containing 10% (v/v) delipidated FCS or 0.2% (v/v) fatty-acid free BSA (Sigma-Aldrich) and 0.5 μCi/ml [4-14C]cholesterol (Amersham, Buckinghamshire, U.K.) for 24 h. Subsequently, cells were treated for 24 h in media containing 10% (v/v) delipidated FCS or 0.2% (v/v) fatty-acid free BSA and 10 μg/ml apolipoprotein A-I (apoA-I; Sigma-Aldrich) in the presence or absence of TL1A or IFN-γ. Next, the media were collected and the remaining cells were solubilized in 0.2 M NaOH. Cholesterol efflux was calculated as the percentage of [14C] counts in the medium versus total (cells and medium) as measured using a liquid scintillation counter. Fold change in cholesterol efflux was the fold difference in efflux between control and treated samples.
Quantitation of macrophage total and esterified cholesterol content
Macrophage (THP-1) cholesterol content was quantified using the Amplex Red Cholesterol Assay Kit (Molecular Probes, Eugene, OR). Macrophages were fixed in 2% (v/v) paraformaldehyde for 15 min and washed three times with PBS before incubation with 200 μl absolute ethanol for 30 min at 4°C to extract cellular lipids. Total cholesterol and cholesterol ester content was determined by incubating 50 μl extracted lipids, diluted in 1× reaction buffer supplied with the kit, with 50 μl Amplex Red Assay reagent containing cholesterol esterase (total cholesterol) or with 50 μl Amplex Red Assay reagent lacking cholesterol esterase (free cholesterol) for 30 min at 37°C in the dark and then measuring fluorescence (BMG Labtech [Aylesbury, U.K.] FLUOstar Optima microplate reader; 535 nm excitation, 595 nm emission). Total and free cholesterol content of each sample was calculated using a cholesterol standard curve. Cholesterol ester content was calculated by subtracting levels of free cholesterol from levels of total cholesterol for each sample. Lipid-extracted macrophages were incubated with 0.1% (w/v) SDS/0.2 M NaOH for 30 min at room temperature, and total cellular protein levels were determined using the Pierce BCA assay (Pierce, Cramlington, U.K.). Total cholesterol and cholesterol ester levels were represented as nanograms of total cholesterol or cholesterol ester per microgram of protein.
Real-time quantitative PCR and DR3 RT-PCR
Total RNAwas prepared using the RNeasy Micro kit (Qiagen, Crawley, U.K.) and was reverse transcribed into cDNA using M-MLV reverse transcriptase (Promega, Southampton, U.K.) and random hexamer primers. Real-time quantitative PCR analysis was performed using SYBR Green JumpStart Taq ReadyMix (Sigma-Aldrich) and primers, detailed in Supplemental Table I, specific for human scavenger receptor (SR) A (SR-A), CD36, SR-BI, lipoprotein lipase (LPL), apolipoprotein E (apoE), ATP-binding cassette transporter A1 (ABCA-1), ABCG-1 and mouse SR-A, CD36, SR-BI, apoE, ABCA-1 and ABCG-1. Quantitative PCR was performed using the Opticon 2 real-time PCR detection system (MJ Research, Cambridge, MA) and mRNA levels were determined using the comparative Ct method and normalized to 28S rRNA (human) and β-actin (mouse) mRNA levels. All PCRs were performed in duplicate and cDNAs, cloned into pGEM-T vector, were used as a standard for quantitation and to verify specificity by DNA sequencing. DR3 RT-PCR was performed using the following primers: 5′-TCACCCTTCTACTGCCAACC-3′ and 5′-CCAGCTGTTACCCACCAACT-3′ (30 cycles, 65°C annealing temperature). 28S rRNA was used as an internal control (12 cycles, 62°C annealing temperature). The PCR products were size fractionated by agarose gel electrophoresis and analyzed using a Syngene gel documentation system (GRI, Braintree, U.K.).
Western blotting
Total cell lysates from THP-1 cells and HMDMs were size fractionated on NuPAGE 4–12% SDS-polyacrylamide gels (Invitrogen) and then analyzed by Western blotting using a previously described method (17). Abs against CD36 (sc-9154), SR-A (sc-20660), SR-B1 (sc-32342) and ABCG-1 (sc-11150) were from Santa Cruz Biotechnology (Santa Cruz, CA). Abs to apoE (0650-1904), ABCA-1 (NB400-105) and β-actin were from Biogenesis (Poole, U.K.), Novus Biologicals (Cambridge, U.K.), and Sigma-Alrich respectively. Semiquantitative measurement of Western blots was performed by densitometric analysis using the Gene Tools software (GRI).
Annexin V/propidium iodide staining of macrophages
Apoptosis was measured in THP-1 macrophages and HMDMs by dual-color immunofluorescent flow cytometry with annexin V-FITC and propidium iodide (BD Biosciences) according to the manufacturer’s instructions. Cells were washed twice in cold PBS, resuspended in 1× binding buffer (1 × 106 cells/ml), and incubated in the dark with 5 μl annexin V-FITC and 10 μl propidium iodide (50 μg/ml) for 15 min at room temperature; 400 μl 1× binding buffer was then added to each sample, and cells were immediately analyzed on a FACSCalibur flow cytometer (BD Biosciences).
Generation of adenovirus encoding DR3 shRNA
An adenovirus type 5 (Ad5) vector was engineered to encode DR3 shRNA flanked by sequences from the murine BIC gene (134–161 and 221–265; RAd-DR3 shRNA) using recombineering technology (18, 19). Two oligonucleotides, detailed in Supplemental Table II, that overlap by 25 bp at their 3′ and 5′ ends, respectively, were designed to contain the appropriate DR3 shRNA sequence and arms of homology to facilitate homologous recombination into the Ad5 vector; 100 ng of each oligonucleotide was transformed into induced SW102 Escherichia coli that contained the Ad5 vector genome in a modified bacterial artificial chromosome and appropriate recombinants were identified as previously described (19). Adenovirus was then amplified, purified, and titered as described (19). A recombinant adenovirus expressing a scrambled shRNA sequence (RAd-scrambled shRNA) was also generated for use as a control. THP-1 cells and 7 d differentiated HMDMs were infected with RAd-scrambled shRNA or RAd-DR3 shRNA at a multiplicity of infection (MOI) of 100, as described in the supporting information. An MOI of 100 was sufficient to infect >98% THP-1 cells and >90% HMDMs as measured by flow cytometry following infection with a GFP-expressing recombinant adenovirus (data not shown).
Results
TL1A drives foam cell formation in human macrophages
Macrophage foam cell formation is driven by an imbalance in the uptake of modified LDL into the cell and the efflux of cholesterol out of it (1-3). To investigate the role of TL1A in macrophage foam cell formation, we first measured the impact of TL1A stimulation on the uptake of acetylated LDL, a type of LDL extensively used for invitro foam cell formation assays (20, 21), and oxidized LDL, a key form of modified LDL found in atherosclerotic plaques (1, 2, 20), and on the efflux of [14C]cholesterol by both 24 h PMA-differentiated THP-1 macrophages and 7 d differentiated HMDMs. THP-1 macrophages were PMA-differentiated for 24 h, because this induces a high level of scavenger receptor expression (22). Fig. 1A and 1B demonstrate that 24 h TL1A stimulation produced a statistically significant increase in the uptake of DiI-AcLDL and DiI-OxLDL, which are widely used by such studies (23), by THP-1 macrophages and HMDMs compared with the untreated control. IFN-γ was used as a positive control because it is known to increase AcLDL and OxLDL uptake and promote foam cell formation (3, 24). The enhancement in DiI-AcLDL uptake by TL1Awas ablated by a soluble DR3-Fc fusion protein (sDR3-Fc), which binds to and neutralizes TL1A (Fig. 1C), and ratified the cytokine-specific response. In addition, it was found that TL1A enhanced the levels of total and esterified cholesterol in AcLDL-converted foam cells (Fig. 1D).
FIGURE 1.

TL1A increases acetylated/oxidized LDL uptake and total and esterified cholesterol levels in THP-1 macrophages and HMDMs. Twenty-four-hour 16 nM PMA-differentiated THP-1 macrophages or 7 d differentiated HMDMs were incubated with 10 μg/ml DiI-AcLDL (A), 10 μg/ml DiI-OxLDL (B), or 10 μg/ml DiI-AcLDL and either 1 μg/ml NKG2D-Fc (as control) or 1 μg/ml sDR3-Fc (C) for 24 h in the presence or absence of 100 ng/ml TL1A or 250 IU/ml IFN-γ (A only). DiI-AcLDL and DiI-OxLDL uptake was measured by flow cytometry. D, Twenty-four-hour PMA-differentiated THP-1 macrophages were incubated with 50 μg/ml AcLDL in the presence or absence of TL1A (100 ng/ml) in media containing 0.2% (w/v) fatty-acid free BSA for 24 h. Cellular lipids and protein were extracted and total cholesterol and cholesterol ester levels were calculated using the Amplex Red assay and normalized to total cellular protein levels as nanograms per micrograms of protein. Filled histograms indicate data for THP-1 macrophages and the striped histograms for HMDMs. Data are derived from four separate experiments (n = 4) and represent mean ± SD. The percent DiI-AcLDL or DiI-oxLDL uptake in untreated cells in A–C has been arbitrarily assigned as 100%. One-way ANOVA with Dunnett’s post hoc test (A) and Student t test (B–D) were used to ascertain the statistical significance of all results. *p < 0.05; **p < 0.01; ***p < 0.001.
Fig. 2A and 2B demonstrate that 24 h TL1A stimulation produced a statistically significant reduction in cholesterol efflux from THP-1 macrophages and HMDMs, compared with the untreated control, which was ablated by coincubation with sDR3-Fc (Fig. 2C). IFN-γ was also used as a positive control, because it is known to decrease cholesterol efflux from macrophages (25) and to validate the cholesterol efflux assay. A similar action of TL1A was observed when the cells were loaded with AcLDL in the absence of serum (Supplemental Fig. 1). Collectively, these data indicate that TL1A drives macrophage foam cell formation by both promoting cholesterol uptake and reducing cholesterol efflux and displays phenotypic similarities to the proatherogenic cytokine IFN-γ3.
FIGURE 2.

TL1A decreases cholesterol efflux from THP-1 macrophages and HMDMs. Twenty-four-hour PMA-differentiated THP-1 macrophages (A) or 7 d differentiated HMDMs (B) were converted into foam cells with 50 μg/ml AcLDL and 0.5 μCi/ml [4-14C]cholesterol for 24 h. Next, cells were treated with 10 μg/ml apoA-I alone (A, B) or in addition to either 1 μg/ml NKG2D-Fc (as control) or 1 μg/ml sDR3-Fc (C) in the presence or absence of 100 ng/ml TL1A or 250 IU/ml IFN-γ (A only) for 24 h. Cholesterol efflux was then assayed. Control cells exhibited ~40% efflux and were arbitrarily assigned a value of 1 from which fold change was calculated. Filled histogram indicates data for THP-1 macrophages, and the striped histogram indicates data for HMDMs. Data are derived from three to four separate experiments (C, n =3; A, B, n = 4) and represents mean ± SD. One-way ANOVA with Dunnett post hoc test (A) and Student t test (B, C) was used to ascertain the statistical significance of all results. *p<0.05; **p<0.01; ***p<0.001.
TL1A regulates the expression of key genes implicated in the uptake and efflux of cholesterol
The uptake of AcLDL and OxLDL into macrophages principally involves a family of scavenger receptors, including SR-A, SR-B1, and CD36 (3, 26). The efflux of cholesterol from macrophages primarily involves reverse cholesterol transport by members of the ABC transporter family, such as ABCA-1 and ABCG-1, and also requires lipid-free components of high-density lipoprotein, such as apoE, and apoA-I as cholesterol acceptors (1-3). Cytokines, like IFN-γ, are known to regulate macrophage foam cell formation by triggering changes in the expression of key genes implicated in the uptake and efflux of cholesterol (3, 25). Because TL1A enhanced AcLDL and OxLDL uptake and reduced cholesterol efflux (Figs. 1, 2) in both THP-1 macrophages and HMDMs, we next investigated whether TL1A can modulate changes in the expression of these key genes by real-time quantitative PCR. Fig. 3 demonstrates that TL1A produced a statistically significant increase in expression of all three scavenger receptor genes as well as LPL, which also aids the uptake of modified LDL (27), in both THP-1 macrophages and HMDMs. Although these increases in gene expression were modest in isolation, collectively they contribute to the global phenotype of increased cholesterol uptake by macrophages. TL1A produced a statistically significant decrease in apoE and ABCG-1 gene expression in THP-1 macrophages and HMDMs. TL1A also reduced ABCA-1 expression in these cells, although this inhibition was only statistically significant in THP-1 macrophages. These TL1A-stimulated reductions in apoE, ABCA-1, and ABCG-1 mRNA expression were also similarly seen in AcLDL-converted foam cells (Supplemental Fig. 2). In addition, the reduction in gene expression by TL1A was found not to involve the apoptotic pathway in THP-1 macrophages and HMDMs (Supplemental Fig. 3).
FIGURE 3.

TL1A regulates the expression of key genes implicated in the uptake and efflux of cholesterol in THP-1 macrophages and HMDMs. Real-time quantitative PCR for SR-A, CD36, SR-BI, apoE, ABCA-1, ABCG-1, and LPL was performed on cDNA from 24 h PMA-differentiated THP-1 macrophages or 7 d differentiated HMDMs incubated for 24 h in the presence or absence of 100 ng/ml TL1A. mRNA levels were calculated using comparative Ct method and normalized to 28s rRNA levels, with untreated cells given an arbitrary value of 1. Filled histogram indicates data for THP-1 macrophages, and the striped histogram indicates data for HMDMs. Data is derived from four separate experiments (n = 4) and represents mean ± SD. One-way ANOVA with Dunnett post hoc test was used to ascertain the statistical significance of the results. *p < 0.05; **p < 0.01; ***p < 0.001.
To confirm that the real-time quantitative PCR data correlated with changes in protein expression, Western blot analysis was performed to analyze CD36, SR-A, SR-B1, apoE, ABCA-1, and ABCG-1 protein expression. Fig. 4 shows that TL1A increased the expression of all three scavenger receptor proteins and reduced the expression of apoE, ABCA-1, and ABCG-1 proteins in correlation with the real-time quantitative PCR data. These changes were all found to be statistically significant (Supplemental Fig. 4). Collectively, these data suggest that TL1A promotes macrophage foam cell formation by altering the mRNA and protein expression of key genes implicated in the uptake and efflux of cholesterol.
FIGURE 4.
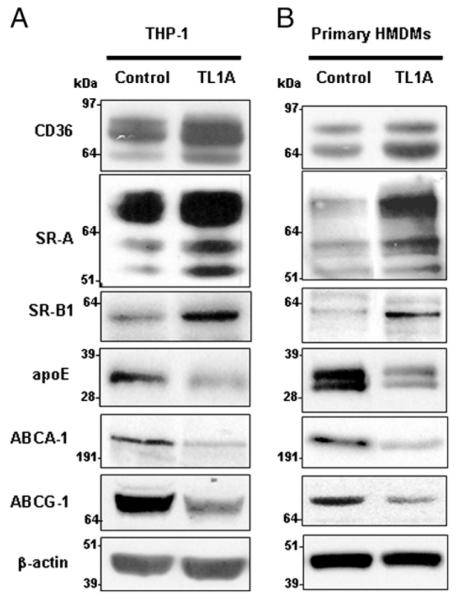
TL1A regulates the expression of key proteins implicated in the uptake and efflux of cholesterol in THP-1 macrophages and HMDMs. Twenty-four-hour PMA-differentiated THP-1 macrophages (A) or 7 d differentiated HMDMs (B) were incubated for 24 h in the presence or absence of 100 ng/ml TL1A. Equal levels of protein from total lysates were analyzed by Western blotting using Abs against CD36, SR-A, SR-B1, apoE, ABCA-1, ABCG-1, and β-actin. The results shown are representative of four separate experiments (n = 4). Semiquantitative analysis of all Western blots shows that all changes were found to be statistically significant (Supplemental Fig. 4)
DR3 regulates foam cell formation in untreated and TL1A-stimulated macrophages
Having shown that TL1A can promote macrophage foam cell formation in both THP-1 macrophages and HMDMs, we next investigated whether its target receptor, DR3, was integral to this action. To do this, we generated a recombinant adenovirus encoding DR3 shRNA to knock down DR3 expression in these cells. The DR3 shRNA adenovirus produced an ~83% knockdown in DR3 mRNA expression in THP-1 macrophages and an ~40% knockdown in HMDMs—compared with a control adenovirus expressing scrambled shRNA (Fig. 5A,5B), which is widely used as a control for RNA interference studies—and thus successfully reduced DR3 expression in human macrophages.
FIGURE 5.

DR3 regulates foam cell formation in THP-1 and HMDMs. THP-1 macrophages (A) or 7 d differentiated HMDMs (B) were infected with either RAd-scrambled shRNA or RAd-DR3 shRNA at an MOI of 100 for 72 h. DR3 RT-PCR was performed and 28S rRNA was used as a housekeeping gene. RT-PCR was used instead of Western blotting because of detection problems with anti-DR3 Abs. −RT indicates that reverse transcriptase was omitted in the cDNA synthesis step using RNA from RAd-scrambled shRNA infection. M indicates m.w. markers. Data are indicative of two separate experiments. RAd-scrambled shRNA or RAd-DR3 shRNA-infected THP-1 macrophages or 7 d differentiated HMDMs were incubated with 10 μg/ml DiI-AcLDL for 24 h (C) or with 50 μg/ml AcLDL and 0.5 μCi/ml [4-14C]cholesterol for 24 h, followed by treatment with 10 μg/ml apoA-I (D) in the presence or absence of 100 ng/ml TL1A. DiI-AcLDL uptake and cholesterol efflux was then assayed. Filled histogram indicates data for THP-1 macrophages, and the striped histogram indicates data for HMDMs. Data are derived from three separate experiments (n = 3) and represent mean ± SD. One-way ANOVA with Dunnett post hoc test was used to ascertain the statistical significance of the results. *p < 0.05; ***p < 0.001.
We next analyzed the uptake of DiI-AcLDL and efflux of [14C]-labeled cholesterol by THP-1 and HMDMs infected with either a scrambled shRNA adenovirus or a DR3 shRNA adenovirus that were then stimulated in the presence or absence of TL1A (Fig. 5C, 5D). In cells infected with a scrambled shRNA adenovirus, TL1A produced a statistically significant increase in DiI-AcLDL uptake and reduction in [14C]cholesterol efflux by both THP-1 macrophages and HMDMs respectively mirroring that seen in uninfected cells (Figs. 1, 2). However, in THP-1 macrophages and HMDMs infected with a DR3 shRNA adenovirus, TL1A did not produce a statistically significant increase in DiI-AcLDL uptake or reduction in [14C]cholesterol efflux by THP-1 macrophages and HMDMs. In addition, a statistically significant reduction in DiI-AcLDL uptake and increase in [14C]cholesterol efflux was seen in unstimulated cells. These data suggest that the uptake of AcLDL and efflux of cholesterol by non-cytokine stimulated macrophages is DR3-dependent and that attenuation of DR3 expression in these cells has prevented TL1A from regulating AcLDL uptake and cholesterol efflux. Collectively, these data indicate that TL1A promotes macrophage foam cell formation in a potentially DR3-dependent fashion and that DR3 can regulate macrophage foam cell formation by itself.
DR3 regulates basal cholesterol uptake and efflux in mouse BMMs
To confirm the role of DR3 in noncytokine stimulated macrophage foam cell formation, we compared DiI-AcLDL uptake and [14C] cholesterol efflux in BMMs isolated from DR3WT and DR3KO mice on the same genetic background. Fig. 6A demonstrates that DiI-AcLDL uptake by DR3KO BMMs was significantly lower than that seen by DR3WT BMMs. Fig. 6B shows that basal cholesterol efflux from DR3KO BMMs was significantly higher than that seen from DR3WT BMMs. These results strongly suggest that DR3 promotes macrophage cholesterol uptake, limits cholesterol efflux, and supports the data shown in human cells (Fig. 5C, 5D). Real-time quantitative PCR assays demonstrated that the relative mRNA expression of mouse SR-A, CD36, and SR-BI was significantly lower in DR3KO BMMs compared with that seen in DR3WT BMMs; conversely, the relative expression of mouse apoE and ABCG-1 was significantly higher in DR3KO BMMs compared with that seen in DR3WT BMMs. Mouse ABCA-1 mRNA expression was higher in DR3KO BMMs, although it was not statistically significant (Fig. 6C, 6D). These data indicate that the reduced level of AcLDL uptake and increased level of cholesterol efflux in DR3KO BMMs is elicited by changes in the expression of these key scavenger receptor and cholesterol efflux genes. Collectively, these data strongly suggest that DR3 is able to promote macrophage foam cell formation by regulating the expression of key genes implicated in the uptake and efflux of cholesterol.
FIGURE 6.

DR3 regulates foam cell formation in mouse BMMs. DR3WT and DR3KO BMMs were incubated with 10 μg/ml DiI-AcLDL for 24 h (A) or with 50 μg/ml AcLDL and 0.5 μCi/ml [4-14C]cholesterol for 24 h, followed by treatment with 10 μg/ml apoA-I for 24 h (B). DiI-AcLDL uptake and cholesterol efflux was then assayed. Data are derived from three to four separate experiments (A, n =3; B, n =4) and represents mean ± SD. The value from DR3WT cells in A and B has been arbitrarily assigned as 100% and 1, respectively. Real-time quantitative PCR for mouse CD36, SR-A, SR-BI (C) and mouse apoE, ABCA-1, and ABCG-1 (D) was performed on cDNA generated from DR3WT and DR3KO BMMs. Relative expression levels were calculated using comparative Ct method and normalized to mouse β-actin, with DR3WT BMMs given an arbitrary value of 1. Data are derived from four separate experiments (n =4) and represent mean ± SD. Student t test was used ascertain the statistical significance of all the results. *p < 0.05; **p < 0.01; ***p < 0.001.
Discussion
This study is the first to report a role for TL1A and DR3 in promoting macrophage foam cell formation and contributes further to published evidence implicating both as proatherogenic mediators. This study also reveals a mechanism for such actions, which is mediated through corresponding changes in the expression of key genes implicated in the uptake and efflux of cholesterol by macrophages. For example, the TL1A-mediated stimulation of modified LDL uptake (Fig. 1) is associated with induced expression of three key proteins, SR-A, CD36, and LPL (Figs. 3, 4), that have been demonstrated to promote such cellular LDL uptake in vitro and in vivo (26-28). In further support, our recent studies have shown that TL1A induces the mRNA expression of the oxLDL-binding, scavenger receptor-PSOX (SR-PSOX/CXCL16; Supplemental Fig. 5), another key gene implicated in cellular lipoprotein uptake (29). Alternatively, TL1A inhibits cholesterol efflux (Fig. 2) and suppresses the expression of three proteins, apoE, ABCA1, and ABCG1 (Figs. 3, 4), which have been shown similarly by numerous studies to promote this process in vitro and in vivo (1, 30). TL1A also induces the expression of SR-BI (Figs. 3, 4), although this is likely to contribute partly to its actions, because it has been found to be involved in both the uptake and efflux of cholesterol (26, 28).
The findings presented in this study are consistent with a number of studies showing that TL1A can steer the progression of inflammatory-driven disease (5, 7, 9, 10, 12-14). The effect of TL1A in promoting macrophage foam cell formation reflects the effect observed for other TNFSF members (31, 32). TNF-α has been shown to enhance macrophage foam cell formation by increasing total cholesterol content through ABCA-1 inhibition (31) and by ablating the catabolism of intracellular lipids (32). Given that TL1A is found localized to atherosclerotic plaques (6), as is TNF-α (33), it is clear that members of this cytokine superfamily play a role in driving atherosclerotic progression in part by promoting macrophage foam cell formation. In addition, it is possible that TL1A contributes to the formation of foam cells derived from vascular smooth muscle cells, because TNF-α promotes vascular smooth muscle cell foam cell formation by increasing SR expression and AcLDL uptake (34).
The identification of a role for DR3 in basal macrophage foam cell formation in this study provides evidence that this receptor is able to regulate this atherosclerotic mechanism alone. This observation is not unique to TNF superfamily receptors, because TNFR1 can regulate macrophage foam cell formation (35). This evidence for TNFR1 and now DR3 indicates that TNF superfamily ligands and their receptors are both able to modulate this atherosclerotic process. The role of DR3 in basal macrophage foam cell formation can be observed owing to a direct regulatory effect of DR3 on the expression of genes implicated in modified LDL uptake and cholesterol efflux, because the expression of genes such as SR-A and ABCA-1 are altered in mice lacking DR3 (Fig. 6). This observation again mirrors TNFR1, because macrophages from irradiated LDL receptorKO mice reconstituted with TNFR1KO hematopoietic stem cells displayed reduced SR-A expression (35). It is also possible that DR3 can trigger multiple signaling cascades, such as the NF-kB and JNK 2 pathways that mediate DR3-specific regulation of IL-8 expression (36), to induce its regulatory response on downstream gene expression. Collectively, this indicates that TNF superfamily ligands and also their receptors, such as TL1A and DR3, are both able to drive macrophage foam cell formation.
We showed that the ability for TL1A to induce macrophage foam cell formation was potentially dependent on the presence of DR3. However, TL1A is also a target for a soluble TNF receptor, decoy receptor 3 (DcR3), which functions as a TL1A antagonist (8). Although DcR3 can bind TL1A, our study shows that TL1A is unlikely to promote macrophage foam cell formation through DcR3. This is because the ability of TL1A to both increase the uptake of AcLDL into human macrophages and reduce the efflux of cholesterol was attenuated by soluble DR3-Fc and by adenovirally delivered DR3 shRNA (Figs. 1, 2, 5). These data also strongly suggest that the actions of TL1A are direct and not a secondary response to the production of other proatherogenic factors. Indeed, in a previous study by Kang et al. (6), TL1A or activation of DR3 by cross-linking with immobilized Abs only produced a marked induction in macrophages of the proinflammatory cytokines TNF-α and IL-8 with matrix metalloproteinase-9 following coculture with IFN-γ. The alteration in macrophage foam cell formation seen in DR3KO mice does not involve DcR3, because mice lack the DcR3 gene in humans (37). These findings strongly suggest that TL1A is DR3-dependent in its proatherogenic ability to promote macrophage foam cell formation. However, it is possible for DcR3 to be involved in macrophage foam cell formation and to have an inhibitory function in this process.
The data presented in this paper emphasize a proatherogenic role for both TL1A and DR3 in driving macrophage foam cell formation. We show that TL1A produces an imbalance in cholesterol homeostasis by orchestrating changes in the expression of key genes implicated in the uptake and efflux of cholesterol, such as SR-A and ABCA-1, and that the ability of TL1A to promote foam cell formation is potentially dependent on DR3. Our data suggest that the TL1A–DR3 pathway is an important contributing factor to atherogenesis and thus represents itself as a novel therapeutic target for the intervention of atherosclerosis.
Supplementary Material
Acknowledgments
This work was supported by grants from the British Heart Foundation (PG/07/031/22716toD.P.R), the Biotechnology and Biological Sciences Research Council (BBF0098361 to G.W.G.W), the Wellcome Trust (GR07075 to G.W.G.W), and the Medical Research Council (G0700142 to G.W.G.W and G0300180 to E.C.Y.W).
Abbreviations used in this paper
- AcLDL
acetylated low-density lipoprotein
- Ad5
adenovirus type 5
- apoA-I
apolipoprotein A-I
- apoE
apolipoprotein E
- BMM
bone-marrow macrophage
- DcR3
decoy receptor 3
- DiI
1,1′-dioctadecyl-3,3,3′,3′-tetra-methyllindocarbocyane
- DR3
death receptor 3
- HMDM
human monocyte-derived macrophage
- KO
knockout
- LDL
low-density lipoprotein
- LPL
lipoprotein lipase
- MOI
multiplicity of infection
- SR
scavenger receptor
- TL1A
TNF-like protein 1A
- TNFRSF
TNFR superfamily
- WT
wild type
Footnotes
Disclosures The authors have no financial conflicts of interest.
The online version of this article contains supplemental material.
References
- 1.Lusis AJ. Atherosclerosis. Nature. 2000;407:233–241. doi: 10.1038/35025203. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Lusis AJ, Mar R, Pajukanta P. Genetics of atherosclerosis. Annu. Rev. Genomics Hum. Genet. 2004;5:189–218. doi: 10.1146/annurev.genom.5.061903.175930. [DOI] [PubMed] [Google Scholar]
- 3.McLaren JE, Ramji DP. Interferon gamma: a master regulator of atherosclerosis. Cytokine Growth Factor Rev. 2009;20:125–135. doi: 10.1016/j.cytogfr.2008.11.003. [DOI] [PubMed] [Google Scholar]
- 4.Kavurma MM, Tan NY, Bennett MR. Death receptors and their ligands in atherosclerosis. Arterioscler. Thromb. Vasc. Biol. 2008;28:1694–1702. doi: 10.1161/ATVBAHA.107.155143. [DOI] [PubMed] [Google Scholar]
- 5.Croft M. The role of TNF superfamily members in T-cell function and diseases. Nat. Rev. Immunol. 2009;9:271–285. doi: 10.1038/nri2526. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Kang YJ, Kim WJ, Bae HU, Kim DI, Park YB, Park JE, Kwon BS, Lee WH. Involvement of TL1A and DR3 in induction of proinflammatory cytokines and matrix metalloproteinase-9 in atherogenesis. Cytokine. 2005;29:229–235. doi: 10.1016/j.cyto.2004.12.001. [DOI] [PubMed] [Google Scholar]
- 7.Fang L, Adkins B, Deyev V, Podack ER. Essential role of TNF receptor superfamily 25 (TNFRSF25) in the development of allergic lung inflammation. J. Exp. Med. 2008;205:1037–1048. doi: 10.1084/jem.20072528. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Migone TS, Zhang J, Luo X, Zhuang L, Chen C, Hu B, Hong JS, Perry JW, Chen SF, Zhou JX, et al. TL1A is a TNF-like ligand for DR3 and TR6/DcR3 and functions as a T cell costimulator. Immunity. 2002;16:479–492. doi: 10.1016/s1074-7613(02)00283-2. [DOI] [PubMed] [Google Scholar]
- 9.Pappu BP, Borodovsky A, Zheng TS, Yang X, Wu P, Dong X, Weng S, Browning B, Scott ML, Ma L, et al. TL1A-DR3 interaction regulates Th17 cell function and Th17-mediated autoimmune disease. J. Exp. Med. 2008;205:1049–1062. doi: 10.1084/jem.20071364. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Meylan F, Davidson TS, Kahle E, Kinder M, Acharya K, Jankovic D, Bundoc V, Hodges M, Shevach EM, Keane-Myers A, et al. The TNF-family receptor DR3 is essential for diverse T cell-mediated inflammatory diseases. Immunity. 2008;29:79–89. doi: 10.1016/j.immuni.2008.04.021. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Bamias G, Martin C, III, Marini M, Hoang S, Mishina M, Ross WG, Sachedina MA, Friel CM, Mize J, Bickston SJ, et al. Expression, localization, and functional activity of TL1A, a novel Th1-polarizing cytokine in inflammatory bowel disease. J. Immunol. 2003;171:4868–4874. doi: 10.4049/jimmunol.171.9.4868. [DOI] [PubMed] [Google Scholar]
- 12.Takedatsu H, Michelsen KS, Wei B, Landers CJ, Thomas LS, Dhall D, Braun J, Targan SR. TL1A (TNFSF15) regulates the development of chronic colitis by modulating both T-helper 1 and T-helper 17 activation. Gastroenterology. 2008;135:552–567. doi: 10.1053/j.gastro.2008.04.037. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Bull MJ, Williams AS, Mecklenburgh Z, Calder CJ, Twohig JP, Elford C, Evans BA, Rowley TF, Slebioda TJ, Taraban VY, et al. The Death Receptor 3-TNF-like protein 1A pathway drives adverse bone pathology in inflammatory arthritis. J. Exp. Med. 2008;205:2457–2464. doi: 10.1084/jem.20072378. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Al-Lamki RS, Wang J, Tolkovsky AM, Bradley JA, Griffin JL, Thiru S, Wang EC, Bolton E, Min W, Moore P, et al. TL1A both promotes and protects from renal inflammation and injury. J. Am. Soc. Nephrol. 2008;19:953–960. doi: 10.1681/ASN.2007060706. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Wang EC, Thern A, Denzel A, Kitson J, Farrow SN, Owen MJ. DR3 regulates negative selection during thymocyte development. Mol. Cell. Biol. 2001;21:3451–3461. doi: 10.1128/MCB.21.10.3451-3461.2001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Calder CJ, Nicholson LB, Dick AD. A selective role for the TNF p55 receptor in autocrine signaling following IFN-gamma stimulation in experimental autoimmune uveoretinitis. J. Immunol. 2005;175:6286–6293. doi: 10.4049/jimmunol.175.10.6286. [DOI] [PubMed] [Google Scholar]
- 17.McLaren J, Rowe M, Brennan P. Epstein-Barr virus induces a distinct form of DNA-bound STAT1 compared with that found in interferon-stimulated B lymphocytes. J. Gen. Virol. 2007;88:1876–1886. doi: 10.1099/vir.0.82741-0. [DOI] [PubMed] [Google Scholar]
- 18.Chung KH, Hart CC, Al-Bassam S, Avery A, Taylor J, Patel PD, Vojtek AB, Turner DL. Polycistronic RNA polymerase II expression vectors for RNA interference based on BIC/miR-155. Nucleic Acids Res. 2006;34:e53. doi: 10.1093/nar/gkl143. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Stanton RJ, McSharry BP, Armstrong M, Tomasec P, Wilkinson GWG. Re-engineering adenovirus vector systems to enable high-throughput analyses of gene function. Biotechniques. 2008;45:659–662. 664–668. doi: 10.2144/000112993. [DOI] [PubMed] [Google Scholar]
- 20.Geng YJ, Hansson GK. Interferon-gamma inhibits scavenger receptor expression and foam cell formation in human monocyte-derived macrophages. J. Clin. Invest. 1992;89:1322–1330. doi: 10.1172/JCI115718. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Goldstein JL, Ho YK, Basu SK, Brown MS. Binding site on macrophages that mediates uptake and degradation of acetylated low density lipoprotein, producing massive cholesterol deposition. Proc. Natl. Acad. Sci. USA. 1979;76:333–337. doi: 10.1073/pnas.76.1.333. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Grewal T, Priceputu E, Davignon J, Bernier L. Identification of a gamma-interferon-responsive element in the promoter of the human macrophage scavenger receptor A gene. Arterioscler. Thromb. Vasc. Biol. 2001;21:825–831. doi: 10.1161/01.atv.21.5.825. [DOI] [PubMed] [Google Scholar]
- 23.Devaraj S, Hugou I, Jialal I. Alpha-tocopherol decreases CD36 expression in human monocyte-derived macrophages. J. Lipid Res. 2001;42:521–527. [PubMed] [Google Scholar]
- 24.Reiss AB, Patel CA, Rahman MM, Chan ES, Hasneen K, Montesinos MC, Trachman JD, Cronstein BN. Interferon-gamma impedes reverse cholesterol transport and promotes foam cell transformation in THP-1 human monocytes/macrophages. Med. Sci. Monit. 2004;10:BR420–BR425. [PubMed] [Google Scholar]
- 25.Panousis CG, Zuckerman SH. Interferon-gamma induces down-regulation of Tangier disease gene (ATP-binding-cassette transporter 1) in macrophage-derived foam cells. Arterioscler. Thromb. Vasc. Biol. 2000;20:1565–1571. doi: 10.1161/01.atv.20.6.1565. [DOI] [PubMed] [Google Scholar]
- 26.Plüddemann A, Neyen C, Gordon S. Macrophage scavenger receptors and host-derived ligands. Methods. 2007;43:207–217. doi: 10.1016/j.ymeth.2007.06.004. [DOI] [PubMed] [Google Scholar]
- 27.Irvine SA, Foka P, Rogers SA, Mead JR, Ramji DP. A critical role for the Sp1-binding sites in the transforming growth factor-beta-mediated inhibition of lipoprotein lipase gene expression in macrophages. Nucleic Acids Res. 2005;33:1423–1434. doi: 10.1093/nar/gki280. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Moore KJ, Freeman MW. Scavenger receptors in atherosclerosis: beyond lipid uptake. Arterioscler. Thromb. Vasc. Biol. 2006;26:1702–1711. doi: 10.1161/01.ATV.0000229218.97976.43. [DOI] [PubMed] [Google Scholar]
- 29.Shimaoka T, Kume N, Minami M, Hayashida K, Kataoka H, Kita T, Yonehara S. Molecular cloning of a novel scavenger receptor for oxidized low density lipoprotein, SR-PSOX, on macrophages. J. Biol. Chem. 2000;275:40663–40666. doi: 10.1074/jbc.C000761200. [DOI] [PubMed] [Google Scholar]
- 30.Tall AR, Yvan-Charvet L, Terasaka N, Pagler T, Wang N. HDL, ABC transporters, and cholesterol efflux: implications for the treatment of atherosclerosis. Cell Metab. 2008;7:365–375. doi: 10.1016/j.cmet.2008.03.001. [DOI] [PubMed] [Google Scholar]
- 31.Mei CL, Chen ZJ, Liao YH, Wang YF, Peng HY, Chen Y. Interleukin-10 inhibits the down-regulation of ATP binding cassette transporter A1 by tumour necrosis factor-alpha in THP-1 macrophage-derived foam cells. Cell Biol. Int. 2007;31:1456–1461. doi: 10.1016/j.cellbi.2007.06.009. [DOI] [PubMed] [Google Scholar]
- 32.Persson J, Nilsson J, Lindholm MW. Interleukin-1beta and tumour necrosis factor-alpha impede neutral lipid turnover in macrophage-derived foam cells. BMC Immunol. 2008;9:70. doi: 10.1186/1471-2172-9-70. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Rus HG, Niculescu F, Vlaicu R. Tumor necrosis factor-alpha in human arterial wall with atherosclerosis. Atherosclerosis. 1991;89:247–254. doi: 10.1016/0021-9150(91)90066-c. [DOI] [PubMed] [Google Scholar]
- 34.Li H, Freeman MW, Libby P. Regulation of smooth muscle cell scavenger receptor expression in vivo by atherogenic diets and in vitro by cytokines. J. Clin. Invest. 1995;95:122–133. doi: 10.1172/JCI117628. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Xanthoulea S, Gijbels MJ, van der Made I, Mujcic H, Thelen M, Vergouwe MN, Ambagts MH, Hofker MH, de Winther MP. P55 tumour necrosis factor receptor in bone marrow-derived cells promotes atherosclerosis development in low-density lipoprotein receptor knock-out mice. Cardiovasc. Res. 2008;80:309–318. doi: 10.1093/cvr/cvn193. [DOI] [PubMed] [Google Scholar]
- 36.Su WB, Chang YH, Lin WW, Hsieh SL. Differential regulation of interleukin-8 gene transcription by death receptor 3 (DR3) and type I TNF receptor (TNFRI) Exp. Cell Res. 2006;312:266–277. doi: 10.1016/j.yexcr.2005.10.015. [DOI] [PubMed] [Google Scholar]
- 37.Han B, Moore PA, Wu J, Luo H. Overexpression of human decoy receptor 3 in mice results in a systemic lupus erythematosus-like syndrome. Arthritis Rheum. 2007;56:3748–3758. doi: 10.1002/art.22978. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.