

Abstract
Current high-throughput screening methods for drug discovery rely on the existence of targets. Moreover, most of the hits generated during screenings turn out to be invalid after further testing in animal models. To by-pass these limitations, efforts are now being made to screen chemical libraries on whole animals. One of the most commonly used animal model in biology is the murine model Mus musculus. However, its cost limit its use in large-scale therapeutic screening. In contrast, the nematode Caenorhabditis elegans, the fruit fly Drosophila melanogaster, and the fish Danio rerio are gaining momentum as screening tools. These organisms combine genetic amenability, low cost and culture conditions that are compatible with large-scale screens. Their main advantage is to allow high-throughput screening in a whole-animal context. Moreover, their use is not dependent on the prior identification of a target and permits the selection of compounds with an improved safety profile. This review surveys the versatility of these animal models for drug discovery and discuss the options available at this day.
Keywords: Caenorhabditis elegans, Drosophila melanogaster, Danio rerio, high throughput screening, drug discovery, chemical genetics, disease, drug target
Introduction
During the last two decades, drug research has been subject to major mutations. Entire genome sequencing, DNA microarrays, miniaturization, informatics and robotics have drastically changed the approach of this art. Pharmaceutical companies were the first to fully combine these emerging technologies and knowledge to establish robust methods for drug discovery (Figure 1.1). One is called High Throughput Screening (HTS) and consists in random screens of compounds to find hits showing an activity or an affinity on a selected target and/or in a model considered representative of a disease (Spring, 2005). This approach has contributed to identify therapeutic compounds, pathway, cell functions, chemical probes, with the ultimate goal of comprehensively delineating relationships between chemical structures and biological activities. Another one is called virtual screening and consists in in silico intelligent drug design against characterized targets (Bajorath, 2002).
Figure 1.
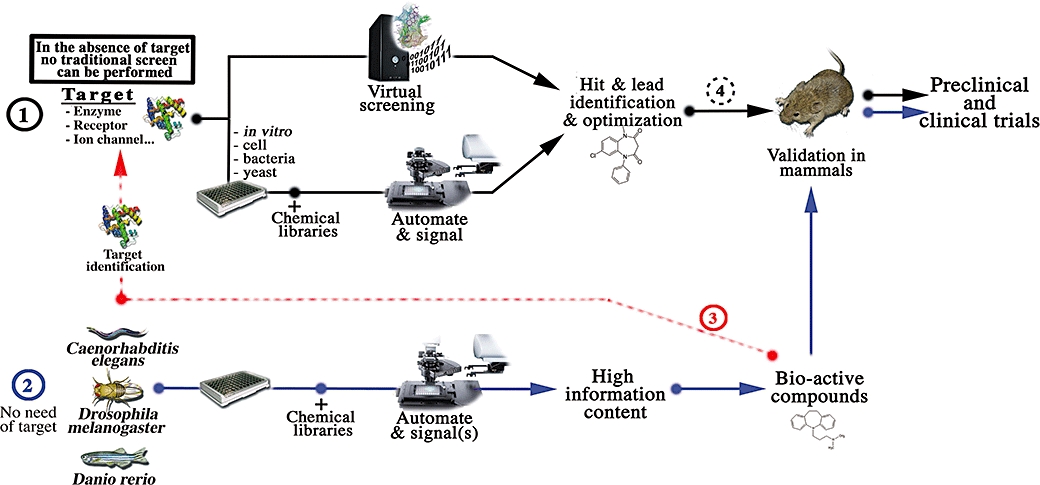
Whole-animal screening and drug discovery process. (1, black lines) show a schematic view of the different stages which came upon a drug discovery process based on traditional HTS. In the absence of target or in complex mechanism, screen can hardly set up. (2, blue line) An alternative may come from phenotypic chemical screens with small animal models like C. elegans, D. melanogaster and D. rerio. (3, red line) Identification of hits in these models may reveals new molecular mechanisms and targets. The target could be further used in traditional HTS. (4) C. elegans, D. melanogaster and D. rerio may also bridge the gap between traditional high-throughput screening and validation in mammalian models. HTS, High Throughput Screening.
Despite numerous successes, the pharmaceutical industry is experiencing a slow-down in the development of new and innovative medical products, due to three major reasons. First, these methods are roughly reliant on the existence of identifiable and screenable targets (Lindsay, 2003). Despite a wealth of information about normal physiology and disease pathology, it is still difficult to predict which targets will effectively reverse a disease phenotype; this fact is particularly true for loss-of-function disease where the identification of pharmaceutically relevant targets is often difficult (Segalat, 2007b). Second, mechanisms involved in some diseases cannot be reproduced in vitro. Cells and tissues are physiologically connected and this interplay may be crucial in the evolution of some disorders. Third, most of the hits generated by traditional screening turn out to be invalid once tested in mouse, resulting in a waste of funds and efforts. Absorption, solubility, distribution, metabolic stability, toxicological problems in later animal studies present numerous difficulties which turn to a dead-end for most hits (Bleicher et al., 2003).
To by pass these limitations, efforts are now being made to screen chemical libraries on whole-animals (Figure 1.2). In medical sciences, the mouse Mus musculus is one of the most commonly used animal model due to its genetic, physiological and anatomical similarities to the human system (West et al., 2000). However, its cost limit its use in large-scale therapeutic screening. On the contrary, with the development of robotic and automated imaging, several tiny animal models, like C. elegans, D. melanogaster and D. rerio, are gaining momentum as screening tools for drug discovery (Lieschke and Currie, 2007; Segalat, 2007a). These organisms combine genetic amenability, low cost and culture conditions compatible with large-scale screening. Their main advantage is to allow high-throughput screening in a whole animal context. Moreover, their use is not dependent on the prior identification of target.
Finally, these new pharmaceutical screening tools will allow (i) the identification of new active compounds that may eventually be validated on mammals (Figure 1.2), (ii) the identification of unsuspected targets and/or molecular mechanisms which could be used in traditional HTS based on target binding or function (Figure 1.3), (iii) the generation of supplementary information which can turn useful to establish a database linking phenotypic activity to chemical structure.
This review discusses the versatility of these models for drug discovery, and try to make an overview of the options currently available at each step of the screening process.
Model attributes overview
C. elegans was introduced by Sydney Brenner in the 1960s as a model organism to study animal development and the nervous system. Since, this little worm has led to landmark discoveries on many molecular mechanisms such as cell death, ageing, development and neuronal function (Lendahl and Orrenius, 2002; Putcha and Johnson, 2004; Kenyon, 2005). It has also been used as a model for host-pathogen interactions and for neurotoxicological research (Moy et al., 2006; Leung et al., 2008; Peterson et al., 2008). Its genome is fully sequenced and more than 50% of human genes have a C. elegans counterpart (Harris et al., 2004). Several human diseases can be recapitulated in this model by knocking down a selected gene or by expressing a deleterious version of it. Unfortunately, the absence of tools to perform targeted mutations complexify this approach. However, many strains have already been generated by large-scale mutagenesis and are available at the Caenorhabditis Genetics Center (Table 1). Indeed, the conservation of such strains is easy and systematic as C. elegans can be stored indefinitely by cryo-conservation (Stiernagle, 2006). In parallel, the NemaGENETAG project has generated a large collection of 14 000 transposon-tagged mutants strains which can be obtained upon request (Bazopoulou and Tavernarakis, 2009). Additionally, double-stranded RNA-mediated interference (RNAi) is an alternative and useful method for gene disruption in C. elegans (Fire et al., 1998). Moreover, several genetic tools have been developed to generate genetically engineered strains, like transgenesis or the MosTIC technology (Rieckher et al., 2009; Robert et al., 2009). Finally, the Wormbase website allows access to an exhaustive database containing comprehensive data on gene structures, mutants and RNAi phenotypes, microarray data, protein-protein interactions and more (Chen et al., 2005).
Table 1.
Overview of animal models characteristics for drug discovery
| C. elegans | D. melanogaster | D. rerio | M. musculus | |
|---|---|---|---|---|
| Practical issues | ||||
| Generation time | 3 to 5 days | 10 to 14 days | 3 to 4 months | 3 to 4 weeks |
| Embryos size | 50 µm | 100 µm | 1 mm | N/A |
| Adult size | 1 mm | 3 mm | 6 cm | 10 cm |
| Growth conditions | Solid or liquid medium | Solid medium | Liquid medium | Cage |
| Easiness to obtain individuals for testing | ++++ | ++++ | +++ | + |
| Ressource | ||||
| Databases | Wormbase (Harris et al., 2004; Chen et al., 2005; Kaletta and Hengartner, 2006) | Flybase (Matthews et al., 2005; Drysdale, 2008) | ZFIN (Sprague et al., 2003; Henken et al., 2004; Meli et al., 2008) | N/A |
| Stock centre | CGC (Stiernagle, 2006) | BDSC (Matthews et al., 2005) | ZIRC (Henken et al., 2004) | N/A |
| Storage | ++++ (frozen) | ++ | +++ (sperm frozen) | + |
| Strains available | ++++ | +++ | + | ++ |
| Characterized inbred strains | +++ | ++ | + | +++ |
| Annual cost | ++++ | ++++ | +++ | + |
| Similarity to human | ||||
| Number of gene (approximately) | 19 000 | 13 000 | 25 000 | 25 000 |
| Homology to human (genome) | >50% | >60% | >70% | >90% |
| Anatomical similarity | + | ++ | +++ | ++++ |
| Pathological similarity | + | + | ++ | ++++ |
| Molecular and genetic similarity | ++ | ++ | +++ | ++++ |
| Molecular tools | ||||
| Targeted gene KO/time | No | No | No | Yes months |
| Forward genetic tools | Yes (Jorgensen and Mango, 2002) | Yes (Venken and Bellen, 2005) | Yes (Solnica-Krezel et al., 1994; Amsterdam, 2003) | No |
| Reverse genetic tools | Yes (Fire et al., 1998; Bazopoulou and Tavernarakis, 2009) | Yes (Venken and Bellen, 2005; Matsushima et al., 2007) | Yes (Nasevicius and Ekker, 2000; Wienholds et al., 2002) | No (excepted targeted KO) |
| Generation of transgenic organism | Weeks (Rieckher et al., 2009) | Weeks (Venken and Bellen, 2005) | Months (Davidson et al., 2003; Esengil and Chen, 2008; Ogura et al., 2009) | Months |
| Tissue-specific promoters available | ++++ | ++++ | ++ | + |
| Antibody reagents | + | + | + | +++ |
| Issues for drug discovery | ||||
| Current use in drug discovery | + | + | ++ | ++++ |
| Available sorting equipment | Eggs to adults (Pulak, 2006; Rohde et al., 2007) | Eggs to larvae (Pulak, 2006) | Eggs to embryos (Pulak, 2006) | N/A |
| Culture in microtiter plate | Eggs to adults | Eggs to larvae | Eggs to larvae | N/A |
| Equipment cost | ++++ | ++++ | +++ | + |
| Screenning throughput | Medium to high | Low to medium | Low to medium | Very low |
| Examples of chemical screen | Gill et al., 2003; Burns et al., 2006; Kwok et al., 2006; Breger et al., 2007; Boyd et al., 2009) | Stilwell et al., 2006 | Burns et al., 2005; Murphey and Zon, 2006; Tran et al., 2007; Hong, 2009 | N/A |
| Review on drug discovery | Artal-Sanz et al., 2006; Kaletta and Hengartner, 2006; Segalat, 2007a | Nichols, 2006; Whitworth et al., 2006 | Parng et al., 2002; Lieschke and Currie, 2007; Rocke et al., 2009 | N/A |
+,++,+++,++++, relative strength of the model in each category; CGC, Caenorhabditis Genetics Center; BDSC, Bloomington Drosophila Stock Center; N/A, not available; ZFIN, Zebrafish Information Network; ZIRC, Zebrafish International Resource Center.
Moreover, due to its little size, its short life cycle, its simple growth conditions and its low-cost, this animal has extensively been used in large-scale genetic screens (Sugimoto, 2004; Bazopoulou and Tavernarakis, 2009). The adult C. elegans reach ∼1 mm in long and ∼80 µm in diameter, whereas embryos are about ∼50 µm long and ∼30 µm in height. This little size allow the use of a specific automated pippeting system at all developmental stages (see below). Animals can easily be cultivated in multi-well plates in agar or liquid medium with a diet of Escherichia coli. Its rapid life cycle is temperature-dependent (3.5 days at 20°C) which is convenient for experiment planification. Moreover, its optically transparent body allow to detect functional and morphological changes without having to kill or dissect the organism.
Despite these advantages for modelling molecular disorders and for large-scale screening, C. elegans has several drawbacks. The major one is that some diseases cannot be reproduced because the animal does not have the corresponding genes or organs. In this case, an alternative could be the fly Drosophila melanogaster or the fish Danio rerio, closer to mammals in the evolutionary tree.
The fruit fly D. melanogaster was introduced as an animal model in the beginning of the 20th century. D. melanogaster has typically been used over the years for genetics, development, signal transduction and cell biology studies, but has also recently been employed for pharmacological research purposes (Manev and Dimitrijevic, 2004; Arias, 2008). The entire Drosophila genome has been sequenced and annoted, and more than 60% of human genes have functional orthologs in D. melanogaster (Table 1) (Bernards and Hariharan, 2001; Celniker and Rubin, 2003; Bier, 2005). Over 75% of the human disease genes in the Online Mendelian Inheritance in Man database present strong protein sequence conservation with D. melanogaster genes (Reiter et al., 2001). Then, this animal model has become a popular organism for studying human diseases (Botas, 2007; Doronkin and Reiter, 2008). D. melanogaster also benefit from a comprehensive range of methods for carrying out molecular genetic research such as mutagenesis, RNAi and transgenesis (Table 1) (Venken and Bellen, 2005; Matsushima et al., 2007). Moreover, there are several efforts under way to mutate every predicted gene. Although D. melanogaster cannot be conserved frozen, it is easy to maintain, and thousands of strains are available from the Bloomington Stock Center (Matthews et al., 2005). Additionally, an exhaustive database information relative to its genetics and its molecular biology is freely available (Drysdale, 2008). Indeed, the FlyBase database contains detailed phenotypic reports for over 2500 mutants and more are characterized every year.
Its size, although larger than C. elegans, allow manipulation of its eggs and its embryos (∼100 µm) with a specific pipetting automate (see below). Although D. melanogaster has already proven its potential for large-scale genetic screening, this animal cannot be grown in liquid medium which limit its use in HTS (Nusslein-Volhard and Wieschaus, 1980; Nagy et al., 2003). However, more and more chemical screens take advantage of it (Nichols, 2006; Whitworth et al., 2006; Segalat, 2007a).
The high degree of functional conservation in cell-biological processes between mammals and invertebrates suggests that diseases resulting from disruption of conserved cellular processes can be recapitulated at a genetic and molecular level in flies and worms. However, one has to keep in mind that the anatomy and physiology of invertebrates are significantly different from those of humans and that, as a consequence, these animals can produce only a partial picture of the human symptoms. On the contrary, the zebrafish (Danio rerio) is a vertebrate model which is more similar to humans. During the last 20 years, this tropical aquarium fish was almost exclusively used to study organ development (Streisinger et al., 1989; Grunwald and Eisen, 2002). Indeed, it was established as a model to bridge the gap between worm/fly and mouse/human for understanding embryonic development. Its interest for research increased substantially approximately 10 years ago, following the demonstration that it was amenable to large-scale forward genetic screens (Eisen, 1996). This resulted in the characterization of an exceptionally large number of genes involved in vertebrate pathways, and contributed to the establishment of the zebrafish as a relevant model for human diseases and pharmaceutical research (Driever et al., 1996; Alestrom et al., 2006). Moreover, the zebrafish genome has now been sequenced, and is part of a large database freely available at the Zebrafish Information Network (ZFIN) and at the FishMap website (Sprague et al., 2003; Meli et al., 2008). Additionally, ZFIN provides exhaustive information to serve the needs of the research community, such as methods, anatomical descriptions, developmental processes, mutants phenotype and more. Finally, working closely with ZFIN, the Zebrafish International Resource Center maintain numerous zebrafish strains and frozen sperm which can be obtained upon request (Henken et al., 2004). Unfortunately, the number of available mutant strains with respect to the number of genes is still limited. The genetic resources for zebrafish continue to expand steadily with the existence of hundreds of genetic mutants from large-scale mutagenesis experiments, the availability of transgenic morpholino knock-down techniques and of the Targeting Induced Local Lesion In Genome and other genetic manipulations such as the sleeping beauty transposase system which allow gene insertion under the control of tissue- and/or time-specific promoters (Table 1) (Nasevicius and Ekker, 2000; Wienholds et al., 2002; Davidson et al., 2003; Esengil and Chen, 2008; Ogura et al., 2009).
An attractive feature of zebrafish assays for pharmacology investigations is the optical transparency of its embryos and its larvae which allow in vivo observation of morphological defects. Moreover, in contrast to rodents, the zebrafish embryos develop externally from the mothers and most of the internal organs, including the cardiovascular system, gut, liver, kidney and muscle develop rapidly in the first 24–48 h (Westerfield, 1995). Then, a wide range of biological and disease process may be studied at an early developmental stages (Chico et al., 2008). In addition, the zebrafish embryo is small (5 mm at 7 days post fertilization), inexpensive, hardy and easy to produce in large numbers. Then, screens may be conducted in microtiter plates (Barros et al., 2008; Hong, 2009).
Screen for what?
HTS with whole animals can be subdivided in two distinct approaches (Figure 2). The first approach is comparable to the classical forward genetic screens which consist in random genetic modifications of wild-type animals by mutagenesis. Similarly, chemical libraries may be tested on wild-type animals for their ability to produce a particular phenotype such as slow growth, lethality, uncoordinated movement, morphological defects and more (Figure 2A). In contrast to traditional biochemical assays that focus on specific molecular targets, a screen based on a phenotypic observation has the advantage of being independent of the specific molecular target involved. Then, depending on the end-point measure (readout), a large variety of bio-active molecules may be detected in the same screen. Additionally, experiments could further lead to the identification of unsuspected targets (see below) (Artal-Sanz et al., 2006; Perrimon et al., 2007). An example of this approach is well illustrated by the screen made by Kwok et al. which screened 14 100 small molecules for bioactivity in wild-type C. elegans and identified 308 compounds that induce a variety of phenotypes, including slow growth, uncoordinated movements and morphology defects (Kwok et al., 2006). One of these compounds, named nemadipine-A, induces morphology and egg-laying defects. Through a genetic suppressor screen, Kwok et al. further identified egl-19, a calcium channel, as the sole candidate target of this compound in C. elegans (Burns et al., 2006; all drug/molecular targets nomenclature follows Alexander et al., 2008). Moreover, by showing that nemadipine-A can also antagonize vertebrate L-type calcium channels, they demonstrated the relevance of this approach for drug discovery.
Figure 2.

High Throughput Screening with whole animals can be separated in two distinct approaches. (A) Hits are selected for their ability to induce a phenotype in a wild-type strain (growth, behaviour, morphology defects or other detectable trait). (B) Hits are selected for their ability to reverse an abnormal phenotype to the wild-type phenotype. Once a bioactive compound is identified, the flexibility and the versatility of these small animals models may rapidly conduct to understand the mechanism of action of each lead at a low cost.
The second approach consists in testing chemical libraries for their ability to reverse an abnormal phenotype to the wild-type phenotype (Figure 2B). HTS is performed on animals that reproduce a disorder or at least, some features of it. For this purpose, mutants strains which mimic human disease are already available or may be generated by mutation, transgenesis or in certain case by chemical treatments (Table 2). As an example, C. elegans and D. melanogaster models of Alzheimer's, Parkinson's and Huntington's disease are available and are being exploited (Link, 2006; Nichols, 2006; Kuwahara et al., 2008). In the case of Alzheimer, loss-of-function mutations in C. elegans homologs of presenilin cause a defect in egg-laying (Smialowska and Baumeister, 2006). Then, the amount of eggs layed may be quantitated to monitor the therapeutical potential of chemical compounds. Comparably, amyloid deposits may be observed in transgenic C. elegans expressing the human β-amyloid peptide (Link, 2006). As this accumulation induces a paralysis phenotype, a screen based on a locomotion readout could be designed (see below). Similarly, because C. elegans is killed by many pathogens such as Pseudomonas aeruginosa, Serretia marcescens, Salmonella enterica, Staphylococus aureus and Streptococcus pneumoniae, a basic screen based on survival may lead to the identification of new anti-microbials agents (Tenor et al., 2004; Sifri et al., 2005; Moy et al., 2006). Finally, in a more subtle approach, the help of fluorescent markers is an additional way of measuring a disorder parameter. Like C. elegans, many mutants and transgenic strains of D. melanogaster present relevant defects that may be used to design a screen (Table 2). Nevertheless, culture conditions of D. melanogaster limit its use to embryos in an HTS process.
Table 2.
Example of disease models showing quantifiable defects
| Disease or disorder | Example of model | Quantifiable phenotype/defect | Reference |
|---|---|---|---|
| Alzheimer's disease | |||
| C. elegans | Transgenics: human beta-amyloid expression | Paralysis due to amyloid deposit | Link, 2006 |
| Mutants: sel-12, homologue of Human presenilin | Egg-laying defect | Smialowska and Baumeister, 2006 | |
| D. melanogaster | Transgenics: human beta-amyloid expression | Progressive locomotor defects, premature death | Nichols, 2006 |
| D. rerio | Knock-down: Zebrafish psen1 or psen2 morpholinos injection | Developmental and morphological defects | Nornes et al., 2009 |
| Knock-down: zebrafish pen-2 morpholinos injection | Neuronal cells loss | Campbell et al., 2006 | |
| Parkinson's disease | |||
| C. elegans | Transgenics: human alpha-synuclein expression | Locomotor defects, dopaminergic cell loss | Kuwahara et al., 2008 |
| D. melanogaster | Transgenics: human alpha-synuclein expression | Locomotor defects | Nichols, 2006 |
| D. rerio | Knock-down: zebrafish parkin morpholinos injection | Dopaminergic cell loss | Flinn et al., 2009 |
| Chemical treatment: PD-inducing neurotoxins | Behavioural and locomotor defects | Bretaud et al., 2004 | |
| Duchenne muscular dystrophy | |||
| C. elegans | Mutants: dys-1, homologue of human dystrophin | Progressive muscle degeneration and paralysis | Gieseler et al., 2000 |
| D. melanogaster | RNAi: reduction of all dystrophin isoform expression | Progressive muscle degeneration and increased pupae lethality | van der Plas et al., 2007 |
| D. rerio | Knock-down: zebrafish dystrophin morpholinos injection | Locomotor and morphological defects | Guyon et al., 2003 |
| Spinal muscular atrophy | |||
| C. elegans | Mutants: SMN-1, homologue of human SMN | Locomotor defects, pharyngeal pumping defects | Briese et al., 2009 |
| RNAi: reduction of SMN-1 expression | Egg-laying defect | Briese et al., 2009 | |
| D. melanogaster | Mutants: SMN, homologue of human SMN | Larval lethality and developmental defects | Chang et al., 2008 |
| D. rerio | Knock-down: zebrafish SMN morpholinos injection | Embryonic lethality and developmental defects | Schmid and DiDonato, 2007 |
| Other muscular disorders | |||
| C. elegans | Mutants: unc-52, homologue of human perlecan | Progressive paralysis | Rogalski et al., 2001 |
| D. melanogaster | Mutants: drosophila delta-sarcoglycan | Reduced life span and locomotor defects | Allikian et al., 2007 |
| D. rerio | Mutants: zebrafish laminin a2 (sapje or candy½oss) | Progressive detachment of muscle ¼bres | Ingham, 2009 |
| Mutants: zebrafish ryr1b, homologue of human RYR1 | Locomotor defects (slow swimming behaviour) | Ingham, 2009 | |
| Cancer | |||
| D. rerio | Transgenics: mouse c-myc expression | Lethal acute lymphoblastic leukemia | Langenau et al., 2003; Langenau et al., 2005a |
| Transgenics: human BRAF melanocyte-specific expression | Malignant melanoma | Patton et al., 2005 | |
| Chemical carcinogen exposure | Assorted tumours (sarcoma, seminoma …) | Beckwith et al., 2000; Spitsbergen et al., 2000 | |
| Fat metabolism | |||
| C. elegans | Mutant: daf-2 | Fat accumulation | Ashrafi et al., 2003 |
| D. melanogaster | Mutants: Adp (Adipose) | Fat accumulation and sterility | Schlegel and Stainier, 2007 |
| D. rerio | Transgenics: AgRP overexpression (agouti-related Protein) | Fat accumulation | Song and Cone, 2007 |
| Pathogen/immunity | |||
| C. elegans | A variety of human pathogens | Premature death | Sifri et al., 2005 |
| D. melanogaster | Tuberculosis-like disease (mycobacterium marinum) | Premature death | Dionne et al., 2003 |
| D. rerio | Streptococcal infections (streptococcus iniae) | Premature death | Neely et al., 2002 |
Although invertebrate models present a high degree of functional conservation with human (Table 1), they often show a partial picture of human processes and, in certain cases, cannot reproduce the disorder as they do not have the corresponding gene and organ. To by-pass this drawback, the vertebrate fish D. rerio could be an alternative, in particular in immunity and cardiovascular disorders, but also in inflammation and in cancer (Table 2). For example, zebrafish develops malignant tumours in response to mutagens, carcinogens or by gene mis-expression (Beckwith et al., 2000; Spitsbergen et al., 2000; Mizgireuv and Revskoy, 2006). Inducing malignancy in transgenic fish carrying oncogenes with fluorescently tags, or fish with appropriate fluorescently marked cell types, results in fluorescent tumours, allowing recognition of tumour onset, location and the estimation of tumour bulk (Langenau et al., 2003; 2005a,b;). This feature, coupled with the optical transparency of its larvae, may be used in large-scale screens to identify enhancers or suppressors of tumours (Stern and Zon, 2003). Similarly, even if zebrafish do not spontaneously develop cardiovascular diseases analogous to those seen in humans, some related mechanisms may be modelled in zebrafish embryos such as cardiomyopathy, thrombosis, arteriogenesis and cardiac regeneration (Chico et al., 2008). Unfortunately, at this day, the lack of appropriate tools for automated manipulation and analysis limit the throughput of screens based on the zebrafish model.
Finally, in comparison with D. melanogaster and D. rerio, an advantage of the C. elegans model for modelling molecular disorders is the possibility to interfere with almost any gene function at any time in its life cycle by delivering RNAi by feeding (Timmons and Fire, 1998). Therefore, chemical screens with essential genes are also possible with this model. For this purpose, a C. elegans RNAi feeding library may be obtained from the Geneservice company (Cambridge UK). This library consists of 16 757 bacterial strains dispensed in 384-well plates, which cover 87% of C. elegans genes. As RNAi is delivered by feeding, it is easy to incorporate it an HTS process in combination with chemical compounds. One has to keep in mind that RNAi is poorly active on the nervous system of C. elegans, whereas a lot of molecule may act in this way. However, a recent study showed that this limitation could be overturned by using a specific strain (Kennedy et al., 2004). Indeed, Seiburth et al. used an eri-1; lin-15B strain to perform a large-scale RNAi screen that identified more than 100 novel genes involved in synaptic transmission (Sieburth et al., 2005).
Options in process design
Defining a relevant model is a crucial step for the success of a screening campaign. In addition, to identify hits among the multitude of tested compounds, a screen must generate comparable, homogeneous and reliable data. Moreover, it needs to be performed in an automatic manner to allow a high-throughput (Zhang et al., 1999). Even if small animal models have already proved their versatility for large-scale studies, scientists usually manipulate these organisms by manually collecting, sorting and transferring individual animals. As a result, large-scale screens often take months or even years, increasing the risk of generating heterogeneous and non-quantitative data. Indeed, whole-animal screens are much more difficult to automate than in vitro screens. Animals size is not suitable with traditional HTS equipment, and culture volumes are often incompatible with large-scale chemical screens. However, emerging methods where culture is performed in minute volumes in 96- or 384-well plates, coupled with the development of robotics, open the way to new possibilities (Table 1) (Lehner et al., 2006; Murphey and Zon, 2006; Pulak, 2006; Vogt et al., 2009). For example, the development of the Complex Object Parametric Analyzer and Sorter (COPAS) Biosort (Union Biometrica, MA, USA), which allows the automatic dispensing of precise numbers of animals into multiwell plates, minimize the initial manipulation of individual animals and therefore dramatically increases the quality and the throughput of the screen. Indeed, the COPAS, like a flow-through sorter, can rapidly sorts objects based on several criteria including length, optical density and several channels of fluorescence (Pulak, 2006). Although all C. elegans stages can be manipulated with this machine, only eggs and embryos of D. melanogaster and D. rerio can.
Another important point of the assay design is the compounds concentration. This question is well debated for in vitro- or cell-based assays. On the contrary, in the case of whole animals, as the target is not always known and the treatment usually delivered through the media, it is difficult to accurately predict the range of doses which have to be tested. Compounds penetrate by both ingestion and diffusion through the animal epiderm (Kaletta and Hengartner, 2006). Indeed, the concentration of a given compound within the animals cannot be predicted and it varies depending on the chemical properties of the compound. As a consequence, negative results cannot be interpreted because it is hardly possible to determine if a negative result is due to poor penetration, docking problems or a true absence of biological activity in the model. It is particularly true for the C. elegans model, which is surrounded by a thick cuticle which provides protection from environmental chemicals (Page and Johnstone, 2007). A recent study evaluated that the absorbed concentration of the 5-hydroxy tryptophane is approximately 100–1000× lower than in the medium (Carre-Pierrat et al., 2006). To avoid missing hits, a conceivable approach may be to test compounds at several concentrations, defined by preexisting data and by the molecular properties of the chemical library. If the number of assays are limited, the compounds may be tested at a high concentration in the medium. With this approach, a lot of of compounds will display a toxic activity, but they could be further re-tested at a lower concentration.
Options for automatic and quantitative read-out?
Another crucial step which has a profound effect upon the quality of the information produced, and upon the throughput, is the output measure (the readout). As a machine is not appropriate to detect the unsuspected, whole animal screening have often relied on laborious observation and manual scoring by small teams of highly trained students and postdocs (Evanko, 2006). Moreover, data generated were often non quantitative and ambiguous. For example, a recent study described a valuable procedure to screen molecules for their ability to induce a phenotype, nevertheless, their throughput was hampered due to the manual interpretation of the data (Burns et al., 2006). In this study, C. elegans larvae were deposited using the COPAS biosort into 24-well plates on top of the agar-chemical mixture. Three to 5 days later, images of the wells were acquired and archived using a HiDI2100 automated imaging system. With this automated method, up to 2400 compounds per week could be screened. Unfortunately, in absence of robust statistical methods to extract data, phenotypes had to be characterized from the archived images which was not better or more rapid than manually scoring phenotype at the dissection microscope. Moreover, the manual interpretation of the data was ambiguous and non quantitative. There is thus a need for more rapid, and more consistent, methods for scoring phenotypes. It is obvious that the ultimate goal of the automatization will be to attempt a comprehensive description of all ‘observable perturbation’ using a large list of numerical parameters. This approach is very complex, it will be highly demanding of programming skills and need considerable progress of the current automated imaging systems. However, several methods already exist at this day to measure the switch between abnormal phenotype to wild-type phenotype, or reciproquely (Table 3).
Table 3.
Example of automated phenotypic observation
| Example of observation | Example of automatic readout | References |
|---|---|---|
| C. elegans | ||
| Global picture of well | Automated imaging of 24-well plate (solid media) | Burns et al., 2006 |
| Life span/death | Measurement of fluorescent dye SYTOX (marker of nematode death) | Gill et al., 2003 |
| Egg-laying | Quantification of the chitinase released into the culture medium | Ellerbrock et al., 2004 |
| Locomotion | Measurement of average speed and paralysis into 35 mm petri plates (solid media) | Tsibidis and Tavernarakis, 2007; Ramot et al., 2008 |
| Measurement of swimming into multiwell plate (liquid media) | Restif and Metaxas, 2008; Tsechpenakis et al., 2008 | |
| Fluorescent cells or tissues | Automated profiling of individual animals into multiwell plate (liquid media) | Pulak, 2006; Rohde et al., 2007 |
| D. melanogaster | ||
| Behaviour and locomotion | Quantification of locomotor activity rhythms | Branson et al., 2009 |
| Measurement of jump reflex (habituation/learning) | Sharma et al., 2009 | |
| Measurement of aggression and courtship | Dankert et al., 2009 | |
| Measurement of locomotion and social behaviours | Reiser, 2009 | |
| Fluorescent cells or tissues | Automated imaging and analysis of Drosophila embryos | Pulak, 2006; Peng et al., 2007 |
| D. rerio | ||
| Histology overview | Automated process which generate digital larval slides for review and annotation | Sabaliauskas et al., 2006 |
| Locomotion/behaviour | Analysis of location and orientation of zebrafish larvae into multiwell plate | Winter et al., 2008; Creton, 2009 |
| Fluorescent cells or tissues | Automated imaging and analysis of zebrafish embryos into multiwell plate | Burns et al., 2005; Tran et al., 2007; Vogt et al., 2009 |
As an example, Gill et al. have developed a method to study the life span of worms. This ‘survival assay’ combine automated worm-handling technology with automated real-time fluorescence detection (Gill et al., 2003). To measure nematode death, they added to the culture the fluorescent dye SYTOX, a nucleic acid stain that binds to DNA in damaged cells. Then, fluorescence was measured using a traditional fluorescence plate reader, allowing the quantification of the amount of dead worms per well. This approach may, in principle, be applied to the identification of pharmacological agents that extend life span of worms or kill them, as well. Additionally, with addition of pathogens in the culture, this method may be used to screen for anti-microbial or anti-fungal compounds. The same approach could be developed for D. melanogaster et D. rerio (Kang et al., 2002; Gerhard, 2007).
With traditional equipment, several other events may be measured in an automatic manner. For example, to measure egg-laying behaviour of C. elegans, a pharmaceutical company has developed an indirect method, called the ‘chitinase assay’ (Ellerbrock et al., 2004). It consists of measuring the chitinase activity that is released from the eggs by hatching larvae. Therefore, this activity reflects the amount of hatching larvae per well. As many disorders models, like neurodegenerative disorders, affect egg-laying behaviour of C. elegans, this rapid measure could be used to monitor the efficiency of the compounds (Smialowska and Baumeister, 2006; Liau et al., 2007; Briese et al., 2009). Nevertheless, one has to keep in mind that the more indirect the observation is, the more false negative are expected.
Additionally, neuro-degenerative, neuro-muscular and several other disorders often result in locomotion or behavioural defects in these models (Table 2). As this type of injury is particularly relevant to human disease, different teams have developed systems to monitor locomotion and behaviour in an automatic and quantitative manner (Tsibidis and Tavernarakis, 2007; Buckingham and Sattelle, 2008; Ramot et al., 2008; Restif and Metaxas, 2008; Tsechpenakis et al., 2008). As an example, a worm-tracker system called the ‘thrashing assay’ is used to evaluate the effect of drugs and mutations on locomotion (Tsechpenakis et al., 2008). Nevertheless, these worm-trackers avoid excessive computational demand by extracting a limited set of features and are time-consuming at this day. In parallel, similar efforts are being made by Drosophila and Zebrafish laboratories (Hicks et al., 2006; Rosato and Kyriacou, 2006; Flinn et al., 2008; Winter et al., 2008; Branson et al., 2009; Creton, 2009; Dankert et al., 2009; Reiser, 2009; Sharma et al., 2009).
Additionally, the optical transparency of the C. elegans body, the D. melanogaster embryos and the D. rerio larvae allows in vivo vizualization and tracking of their cells, tissues and fluorescent markers. Furthermore, the known and invariant cell lineage of C. elegans may be used to establish robust and accurate measurements. Indeed, C. elegans is composed of 959 somatic cells including muscles cells (111), neurons (302), intestine (34 cells), epidermis (213 cells), which are not able to regenerate (Harris et al., 2004). Although this last feature is advantageous for quantitative measurements, it may also be seen as a drawback because many diseases may be treated through the regeneration process.
As example of this approach, mutations in the C. elegans homologue of the human dystrophin, a protein involved in the Duchenne muscular dystrophy, result in a progressive paralysis phenotype (Gieseler et al., 2000). As this paralysis is due to a muscle degenerescence, the use of fluorescent markers under the control of a muscle-specific promoter allow to monitor, in vivo, the degree of injury without having to estimate the locomotion or to perform a muscle-specific staining (Figure 3). Then, with appropriate equipment, the fluorescent pattern of each animals may be collected to establish a quantitative measurement of the muscle degeneresence (Figure 3). Two systems are available at this day to monitor the fluorescent profile of C. elegans. The first one is the multi-well plate reader of the COPAS biosort which has been previously described (Pulak, 2006). The second one consists in a high-speed microfluidic analyser that can isolate and immobilize C. elegans animals in a well defined geometry. Then, fluorescent profiles may be recorded at a subcellular resolution in physiologically active animals (Rohde et al., 2007). Although this recent technology has been designed for sorting, it is attractive for phenotypic analysis at a cellular and subcellular level.
Figure 3.
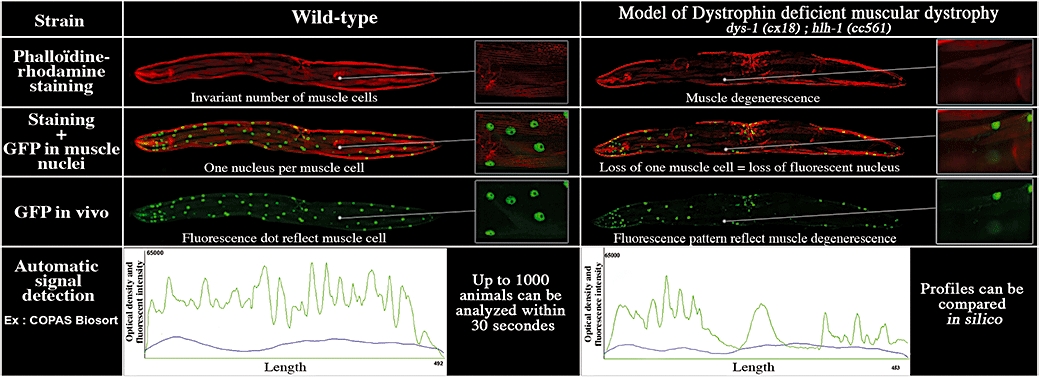
Example of disease model and engineering with C. elegans. Mutations in the C. elegans homologue of the human dystrophin result in a progressive paralysis phenotype due to a muscle degenerescence. Traditionally, this phenotype is quantitated by manually scoring the number of absent muscle cells after labelling of actin fibres. This approach is laborious and time-consuming. However, the use of a fluorescent marker under the control of tissue-specific promoter allows the in vivo quantification of the disorder. Phenotype comparison is possible with automated imaging system like the COPAS Biosort which records the fluorescent profile of individual animals (bottom panels).
The same strategy may be applied to D. melanogaster and D. rerio in their early stages (Table 3). For example, several teams use fluorescent marker coupled to automated imaging systems and artificial intelligence-based image analysis to track the angiogenesis process and find compounds which modulate it (Tran et al., 2007; Vogt et al., 2009). Furthermore, their results demonstrated that it is feasible to adapt image-based high-content screening methodology to measure complex whole organism phenotypes in an automatic manner. Another example is described by Burns et al. who developed a rapid assay to quantify the effect of novel drugs on heart rate by generating transgenic zebrafish with fluorescent cardiomyocytes. Embryos were distributed in 96-well plates and the heart rate of individual animals was measured by an automated imaging system (Burns et al., 2005).
After the screen?
One of the concerns of using small animal models for drug discovery is the determination of the effective dose for a lead compound. Dose-response experiments are easily feasible but one has to keep in mind that the compounds are provided to the animals through the media (Kaletta and Hengartner, 2006). To meet this challenge, the results obtained with these models will have to be linked to data obtained on laboratory rodents and, when possible, in humans. It is too early to establish if the range of doses to be tested in mammals can be extrapolated from data obtained on C. elegans, D. melanogaster and D. rerio. Furthermore, it may be different from drug to drug. At this day, the confirmation in mammals remains essential. Indeed, mammalian models will remain necessary in drug development to answer fundamental questions of drug pharmacology and toxicity.
For the future of drug discovery, the identification of novel targets seems critical. Then, the flexibility of small animal models is a powerful tool to rapidly understand the mechanism of action of each lead at a low cost (Artal-Sanz et al., 2006; Kaletta and Hengartner, 2006). Several approaches may be used to determine the targets and modes of action of active compounds, such as candidate-based approaches, affinity chromatography coupled to mass spectrometry, micro-array technologies and more global approaches like genome-wide RNAi screen, random mutagenesis or metaboprofiling studies (Kaletta and Hengartner, 2006; Lehner et al., 2006; Blaise et al., 2007; Sleno and Emili, 2008; Blaise et al., 2009).
For example, a screen of a collection of 1000 already approved compounds conducted on the C. elegans model of dystrophin-deficient muscular dystrophy led to the identification of several new active compounds (L. Segalat, unpublished results). Two of the most active hits obtained in this screen were methazolamide and dichlorphenamide, which are sulfonamides. As these chemicals are known to be strong inhibitors of human carbonic anhydrase enzymes, implication of these enzymes was investigated in the worm (Giacomotto et al., 2009). For this purpose, RNAi experiments against all putative carbonic anhydrase of the worm were performed, and conducted to identify CAH-4 as the sole target among the six carbonic anhydrases of the worm.
Alternatively, in the absence of candidate genes, the use of a global approach such as large-scale mutagenesis or genome-wide RNAi screens is an option which may rapidly conduct to the identification of potential targets. For example, as RNAi can be delivered by feeding in C. elegans, a secondary RNAi screen – based on the process used for hit identification – may be run in which C. elegans genes are tested one by one for their ability to modify the response to the drug. Then, candidate genes are identified by their ability to induce a resistance or a hypersensivity to the hit when inactivated (Burns et al., 2006). The major drawback of this method is that the process needs considerable amount of chemicals.
Translation to human
How predictable are the findings made on model organisms such as C. elegans, D. melanogaster or Danio rerio, when it comes to treating humans?
One has to keep in mind that the anatomical and molecular differences of small model organisms with humans may cause the elimination of a significant fraction of the hits generated. At this date, the question of hit predictability in this strategy cannot be answered because there is not enough feedback. One may imagine that the number of compounds which can be translated to human may not be much higher than the attrition rate seen with cellular screens. Along the same lines, the targets identified in these models might help to understand normal and disease biology of these tiny animals but not necessarily be relevant for human.
The lead-to-drug bottleneck, which remains a critical problem of today's drug discovery, will likely not be alleviated by screening on small model organisms. These models should be viewed as complementary alternatives to cellular or in-vitro screening devices, rather than as universal shortcuts to human treatments. Their limitations are numerous. However, the glass should be seen as half-full rather than half-empty. As it was already demonstrated in a few cases, the real added value of these models is more in their ability to reveal targets and pathways that would be missed by more conventional devices. Thus, they feed the downstream pipeline of drug discovery.
Conclusion
All in all, whole-animal screening based on C. elegans, D. melanogaster and D. rerio appears as a new tool in the drug discovery process. Their added value for drug discovery varies from disease to disease, and mainly depends on what alternative options are. Indeed, they are complementary to in vitro and cellular systems because their small size and their culture conditions fulfil the requirements for large-scale screens. Moreover, such approaches may permit the selection of potential therapeutic molecules with an improved safety profile earlier in the drug discovery phase, saving both time and funds. Furthermore, the versatility of these small animals for genetic studies potentially allows the rapid target identification of each lead at a low cost. Such new target may eventually feed the traditional HTS strategy based on target binding or function.
Acknowledgments
This work was supported by the Association Française contre les Myopathies (AFM) and by the European Muscle Development Network (MYORES).
Glossary
Abbreviations:
- BDSC
Bloomington Drosophila Stock Center
- CGC
Caenorhabditis Genetics Center
- COPAS
Complex Object Parametric Analyzer and Sorter
- HTS
High Throughput Screening
- RNAi
RNA-mediated interference
- TILLING
Targeting Induced Local Lesion In Genome
- ZFIN
Zebrafish Information Network
- ZIRC
Zebrafish International Resource Center
Conflicts of interest
The authors state that they have no conflict of interest.
References
- Alestrom P, Holter JL, Nourizadeh-Lillabadi R. Zebrafish in functional genomics and aquatic biomedicine. Trends Biotechnol. 2006;24:15–21. doi: 10.1016/j.tibtech.2005.11.004. [DOI] [PubMed] [Google Scholar]
- Alexander SPH, Mathie A, Peters JA. Guide to receptors and Channels (GRAC), 3rd edition (2008 revision) Br J Pharmacol. 2008;153:S1–S209. doi: 10.1038/sj.bjp.0707746. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Allikian MJ, Bhabha G, Dospoy P, Heydemann A, Ryder P, Earley JU, et al. Reduced life span with heart and muscle dysfunction in Drosophila sarcoglycan mutants. Hum Mol Genet. 2007;16:2933–2943. doi: 10.1093/hmg/ddm254. [DOI] [PubMed] [Google Scholar]
- Amsterdam A. Insertional mutagenesis in zebrafish. Dev Dyn. 2003;228:523–534. doi: 10.1002/dvdy.10381. [DOI] [PubMed] [Google Scholar]
- Arias AM. Drosophila melanogaster and the development of biology in the 20th century. Methods Mol Biol. 2008;420:1–25. doi: 10.1007/978-1-59745-583-1_1. [DOI] [PubMed] [Google Scholar]
- Artal-Sanz M, de Jong L, Tavernarakis N. Caenorhabditis elegans: a versatile platform for drug discovery. Biotechnol J. 2006;1:1405–1418. doi: 10.1002/biot.200600176. [DOI] [PubMed] [Google Scholar]
- Ashrafi K, Chang FY, Watts JL, Fraser AG, Kamath RS, Ahringer J, et al. Genome-wide RNAi analysis of Caenorhabditis elegans fat regulatory genes. Nature. 2003;421:268–272. doi: 10.1038/nature01279. [DOI] [PubMed] [Google Scholar]
- Bajorath J. Integration of virtual and high-throughput screening. Nat Rev Drug Discov. 2002;1:882–894. doi: 10.1038/nrd941. [DOI] [PubMed] [Google Scholar]
- Barros TP, Alderton WK, Reynolds HM, Roach AG, Berghmans S. Zebrafish: an emerging technology for in vivo pharmacological assessment to identify potential safety liabilities in early drug discovery. Br J Pharmacol. 2008;154:1400–1413. doi: 10.1038/bjp.2008.249. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bazopoulou D, Tavernarakis N. The NemaGENETAG initiative: large scale transposon insertion gene-tagging in Caenorhabditis elegans. Genetica. 2009;137:39–46. doi: 10.1007/s10709-009-9361-3. [DOI] [PubMed] [Google Scholar]
- Beckwith LG, Moore JL, Tsao-Wu GS, Harshbarger JC, Cheng KC. Ethylnitrosourea induces neoplasia in zebrafish (Danio rerio) Lab Invest. 2000;80:379–385. doi: 10.1038/labinvest.3780042. [DOI] [PubMed] [Google Scholar]
- Bernards A, Hariharan IK. Of flies and men – studying human disease in Drosophila. Curr Opin Genet Dev. 2001;11:274–278. doi: 10.1016/s0959-437x(00)00190-8. [DOI] [PubMed] [Google Scholar]
- Bier E. Drosophila, the golden bug, emerges as a tool for human genetics. Nat Rev Genet. 2005;6:9–23. doi: 10.1038/nrg1503. [DOI] [PubMed] [Google Scholar]
- Blaise BJ, Giacomotto J, Elena B, Dumas ME, Toulhoat P, Segalat L, et al. Metabotyping of Caenorhabditis elegans reveals latent phenotypes. Proc Natl Acad Sci U S A. 2007;104:19808–19812. doi: 10.1073/pnas.0707393104. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Blaise BJ, Giacomotto J, Triba MN, Toulhoat P, Piotto M, Emsley L, et al. Metabolic profiling strategy of Caenorhabditis elegans by whole-organism nuclear magnetic resonance. J Proteome Res. 2009;8:2542–2550. doi: 10.1021/pr900012d. [DOI] [PubMed] [Google Scholar]
- Bleicher KH, Bohm HJ, Muller K, Alanine AI. Hit and lead generation: beyond high-throughput screening. Nat Rev Drug Discov. 2003;2:369–378. doi: 10.1038/nrd1086. [DOI] [PubMed] [Google Scholar]
- Botas J. Drosophila researchers focus on human disease. Nat Genet. 2007;39:589–591. doi: 10.1038/ng0507-589. [DOI] [PubMed] [Google Scholar]
- Boyd WA, Smith MV, Kissling GE, Freedman JH. Medium- and high-throughput screening of neurotoxicants using C. elegans. Neurotoxicol Teratol. 2009 doi: 10.1016/j.ntt.2008.12.004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Branson K, Robie AA, Bender J, Perona P, Dickinson MH. High-throughput ethomics in large groups of Drosophila. Nat Methods. 2009;6:451–457. doi: 10.1038/nmeth.1328. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Breger J, Fuchs BB, Aperis G, Moy TI, Ausubel FM, Mylonakis E. Antifungal chemical compounds identified using a C. elegans pathogenicity assay. Plos Pathog. 2007;3:e18. doi: 10.1371/journal.ppat.0030018. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bretaud S, Lee S, Guo S. Sensitivity of zebrafish to environmental toxins implicated in Parkinson's disease. Neurotoxicol Teratol. 2004;26:857–864. doi: 10.1016/j.ntt.2004.06.014. [DOI] [PubMed] [Google Scholar]
- Briese M, Esmaeili B, Fraboulet S, Burt EC, Christodoulou S, Towers PR, et al. Deletion of smn-1, the Caenorhabditis elegans ortholog of the spinal muscular atrophy gene, results in locomotor dysfunction and reduced lifespan. Hum Mol Genet. 2009;18:97–104. doi: 10.1093/hmg/ddn320. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Buckingham SD, Sattelle DB. Strategies for automated analysis of C. elegans locomotion. Invert Neurosci. 2008;8:121–131. doi: 10.1007/s10158-008-0077-3. [DOI] [PubMed] [Google Scholar]
- Burns AR, Kwok TC, Howard A, Houston E, Johanson K, Chan A, et al. High-throughput screening of small molecules for bioactivity and target identification in Caenorhabditis elegans. Nat Protoc. 2006;1:1906–1914. doi: 10.1038/nprot.2006.283. [DOI] [PubMed] [Google Scholar]
- Burns CG, Milan DJ, Grande EJ, Rottbauer W, MacRae CA, Fishman MC. High-throughput assay for small molecules that modulate zebrafish embryonic heart rate. Nat Chem Biol. 2005;1:263–264. doi: 10.1038/nchembio732. [DOI] [PubMed] [Google Scholar]
- Campbell WA, Yang H, Zetterberg H, Baulac S, Sears JA, Liu T, et al. Zebrafish lacking Alzheimer presenilin enhancer 2 (Pen-2) demonstrate excessive p53-dependent apoptosis and neuronal loss. J Neurochem. 2006;96:1423–1440. doi: 10.1111/j.1471-4159.2006.03648.x. [DOI] [PubMed] [Google Scholar]
- Carre-Pierrat M, Mariol MC, Chambonnier L, Laugraud A, Heskia F, Giacomotto J, et al. Blocking of striated muscle degeneration by serotonin in C. elegans. J Muscle Res Cell Motil. 2006;27:253–258. doi: 10.1007/s10974-006-9070-9. [DOI] [PubMed] [Google Scholar]
- Celniker SE, Rubin GM. The Drosophila melanogaster genome. Annu Rev Genomics Hum Genet. 2003;4:89–117. doi: 10.1146/annurev.genom.4.070802.110323. [DOI] [PubMed] [Google Scholar]
- Chang HC, Dimlich DN, Yokokura T, Mukherjee A, Kankel MW, Sen A, et al. Modeling spinal muscular atrophy in Drosophila. Plos One. 2008;3 doi: 10.1371/journal.pone.0003209. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chen N, Harris TW, Antoshechkin I, Bastiani C, Bieri T, Blasiar D, et al. WormBase: a comprehensive data resource for Caenorhabditis biology and genomics. Nucleic Acids Res. 2005;33:D383–D389. doi: 10.1093/nar/gki066. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chico TJ, Ingham PW, Crossman DC. Modeling cardiovascular disease in the zebrafish. Trends Cardiovasc Med. 2008;18:150–155. doi: 10.1016/j.tcm.2008.04.002. [DOI] [PubMed] [Google Scholar]
- Creton R. Automated analysis of behavior in zebrafish larvae. Behav Brain Res. 2009;203:127–136. doi: 10.1016/j.bbr.2009.04.030. [DOI] [PubMed] [Google Scholar]
- Dankert H, Wang L, Hoopfer ED, Anderson DJ, Perona P. Automated monitoring and analysis of social behavior in Drosophila. Nat Methods. 2009;6:297–303. doi: 10.1038/nmeth.1310. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Davidson AE, Balciunas D, Mohn D, Shaffer J, Hermanson S, Sivasubbu S, et al. Efficient gene delivery and gene expression in zebrafish using the Sleeping Beauty transposon. Dev Biol. 2003;263:191–202. doi: 10.1016/j.ydbio.2003.07.013. [DOI] [PubMed] [Google Scholar]
- Dionne MS, Ghori N, Schneider DS. Drosophila melanogaster is a genetically tractable model host for Mycobacterium marinum. Infect Immun. 2003;71:3540–3550. doi: 10.1128/IAI.71.6.3540-3550.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Doronkin S, Reiter LT. Drosophila orthologues to human disease genes: an update on progress. Prog Nucleic Acid Res Mol Biol. 2008;82:1–32. doi: 10.1016/S0079-6603(08)00001-9. [DOI] [PubMed] [Google Scholar]
- Driever W, Solnica-Krezel L, Schier AF, Neuhauss SC, Malicki J, Stemple DL, et al. A genetic screen for mutations affecting embryogenesis in zebrafish. Development. 1996;123:37–46. doi: 10.1242/dev.123.1.37. [DOI] [PubMed] [Google Scholar]
- Drysdale R. FlyBase: a database for the Drosophila research community. Methods Mol Biol. 2008;420:45–59. doi: 10.1007/978-1-59745-583-1_3. [DOI] [PubMed] [Google Scholar]
- Eisen JS. Zebrafish make a big splash. Cell. 1996;87:969–977. doi: 10.1016/s0092-8674(00)81792-4. [DOI] [PubMed] [Google Scholar]
- Ellerbrock BR, Coscarelli EM, Gurney ME, Geary TG. Screening for presenilin inhibitors using the free-living nematode, Caenorhabditis elegans. J Biomol Screen. 2004;9:147–152. doi: 10.1177/1087057103261038. [DOI] [PubMed] [Google Scholar]
- Esengil H, Chen JK. Gene regulation technologies in zebrafish. Mol Biosyst. 2008;4:300–308. doi: 10.1039/b718447f. [DOI] [PubMed] [Google Scholar]
- Evanko D. In praise of manual high-throughput screening. Nat Methods. 2006;3:662–663. doi: 10.1038/nmeth0906-662b. [DOI] [PubMed] [Google Scholar]
- Fire A, Xu S, Montgomery MK, Kostas SA, Driver SE, Mello CC. Potent and specific genetic interference by double-stranded RNA in Caenorhabditis elegans. Nature. 1998;391:806–811. doi: 10.1038/35888. [DOI] [PubMed] [Google Scholar]
- Flinn L, Bretaud S, Lo C, Ingham PW, Bandmann O. Zebrafish as a new animal model for movement disorders. J Neurochem. 2008;106:1991–1997. doi: 10.1111/j.1471-4159.2008.05463.x. [DOI] [PubMed] [Google Scholar]
- Flinn L, Mortiboys H, Volkmann K, Koster RW, Ingham PW, Bandmann O. Complex I deficiency and dopaminergic neuronal cell loss in parkin-deficient zebrafish (Danio rerio) Brain. 2009;132:1613–1623. doi: 10.1093/brain/awp108. [DOI] [PubMed] [Google Scholar]
- Gerhard GS. Small laboratory fish as models for aging research. Ageing Res Rev. 2007;6:64–72. doi: 10.1016/j.arr.2007.02.007. [DOI] [PubMed] [Google Scholar]
- Giacomotto J, Pertl C, Borrel C, Walter MC, Bulst S, Johnsen B, et al. Evaluation of the therapeutic potential of carbonic anhydrase inhibitors in two animal models of dystrophin deficient muscular dystrophy. Hum Mol Genet. 2009;18:4089–4101. doi: 10.1093/hmg/ddp358. [DOI] [PubMed] [Google Scholar]
- Gieseler K, Grisoni K, Segalat L. Genetic suppression of phenotypes arising from mutations in dystrophin-related genes in Caenorhabditis elegans. Curr Biol. 2000;10:1092–1097. doi: 10.1016/s0960-9822(00)00691-6. [DOI] [PubMed] [Google Scholar]
- Gill MS, Olsen A, Sampayo JN, Lithgow GJ. An automated high-throughput assay for survival of the nematode Caenorhabditis elegans. Free Radic Biol Med. 2003;35:558–565. doi: 10.1016/s0891-5849(03)00328-9. [DOI] [PubMed] [Google Scholar]
- Grunwald DJ, Eisen JS. Headwaters of the zebrafish – emergence of a new model vertebrate. Nat Rev Genet. 2002;3:717–724. doi: 10.1038/nrg892. [DOI] [PubMed] [Google Scholar]
- Guyon JR, Mosley AN, Zhou Y, O'Brien KF, Sheng X, Chiang K, et al. The dystrophin associated protein complex in zebrafish. Hum Mol Genet. 2003;12:601–615. [PubMed] [Google Scholar]
- Harris TW, Chen N, Cunningham F, Tello-Ruiz M, Antoshechkin I, Bastiani C, et al. WormBase: a multi-species resource for nematode biology and genomics. Nucleic Acids Res. 2004;32:D411–D417. doi: 10.1093/nar/gkh066. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Henken DB, Rasooly RS, Javois L, Hewitt AT. The National Institutes of Health and the growth of the zebrafish as an experimental model organism. Zebrafish. 2004;1:105–110. doi: 10.1089/zeb.2004.1.105. [DOI] [PubMed] [Google Scholar]
- Hicks C, Sorocco D, Levin M. Automated analysis of behavior: a computer-controlled system for drug screening and the investigation of learning. J Neurobiol. 2006;66:977–990. doi: 10.1002/neu.20290. [DOI] [PubMed] [Google Scholar]
- Hong CC. Large-scale small-molecule screen using zebrafish embryos. Methods Mol Biol. 2009;486:43–55. doi: 10.1007/978-1-60327-545-3_4. [DOI] [PubMed] [Google Scholar]
- Ingham PW. The power of the zebrafish for disease analysis. Hum Mol Genet. 2009;18:R107–R112. doi: 10.1093/hmg/ddp091. [DOI] [PubMed] [Google Scholar]
- Jorgensen EM, Mango SE. The art and design of genetic screens: caenorhabditis elegans. Nat Rev Genet. 2002;3:356–369. doi: 10.1038/nrg794. [DOI] [PubMed] [Google Scholar]
- Kaletta T, Hengartner MO. Finding function in novel targets: C. elegans as a model organism. Nat Rev Drug Discov. 2006;5:387–398. doi: 10.1038/nrd2031. [DOI] [PubMed] [Google Scholar]
- Kang HL, Benzer S, Min KT. Life extension in Drosophila by feeding a drug. Proc Natl Acad Sci U S A. 2002;99:838–843. doi: 10.1073/pnas.022631999. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kennedy S, Wang D, Ruvkun G. A conserved siRNA-degrading RNase negatively regulates RNA interference in C. elegans. Nature. 2004;427:645–649. doi: 10.1038/nature02302. [DOI] [PubMed] [Google Scholar]
- Kenyon C. The plasticity of aging: insights from long-lived mutants. Cell. 2005;120:449–460. doi: 10.1016/j.cell.2005.02.002. [DOI] [PubMed] [Google Scholar]
- Kuwahara T, Koyama A, Koyama S, Yoshina S, Ren CH, Kato T, et al. A systematic RNAi screen reveals involvement of endocytic pathway in neuronal dysfunction in alpha-synuclein transgenic C. elegans. Hum Mol Genet. 2008;17:2997–3009. doi: 10.1093/hmg/ddn198. [DOI] [PubMed] [Google Scholar]
- Kwok TC, Ricker N, Fraser R, Chan AW, Burns A, Stanley EF, et al. A small-molecule screen in C. elegans yields a new calcium channel antagonist. Nature. 2006;441:91–95. doi: 10.1038/nature04657. [DOI] [PubMed] [Google Scholar]
- Langenau DM, Feng H, Berghmans S, Kanki JP, Kutok JL, Look AT. Cre/lox-regulated transgenic zebrafish model with conditional myc-induced T cell acute lymphoblastic leukemia. Proc Natl Acad Sci U S A. 2005a;102:6068–6073. doi: 10.1073/pnas.0408708102. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Langenau DM, Jette C, Berghmans S, Palomero T, Kanki JP, Kutok JL, et al. Suppression of apoptosis by bcl-2 overexpression in lymphoid cells of transgenic zebrafish. Blood. 2005b;105:3278–3285. doi: 10.1182/blood-2004-08-3073. [DOI] [PubMed] [Google Scholar]
- Langenau DM, Traver D, Ferrando AA, Kutok JL, Aster JC, Kanki JP, et al. Myc-induced T cell leukemia in transgenic zebrafish. Science. 2003;299:887–890. doi: 10.1126/science.1080280. [DOI] [PubMed] [Google Scholar]
- Lehner B, Tischler J, Fraser AG. RNAi screens in Caenorhabditis elegans in a 96-well liquid format and their application to the systematic identification of genetic interactions. Nat Protoc. 2006;1:1617–1620. doi: 10.1038/nprot.2006.245. [DOI] [PubMed] [Google Scholar]
- Lendahl U, Orrenius S. [Sydney Brenner, Robert Horvitz and John Sulston. Winners of the 2002 Nobel Prize in medicine or physiology. Genetic regulation of organ development and programmed cell death] Lakartidningen. 2002;99:4026–4032. [PubMed] [Google Scholar]
- Leung MC, Williams PL, Benedetto A, Au C, Helmcke KJ, Aschner M, et al. Caenorhabditis elegans: an emerging model in biomedical and environmental toxicology. Toxicol Sci. 2008;106:5–28. doi: 10.1093/toxsci/kfn121. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Liau WS, Gonzalez-Serricchio AS, Deshommes C, Chin K, LaMunyon CW. A persistent mitochondrial deletion reduces fitness and sperm performance in heteroplasmic populations of C. elegans. BMC Genet. 2007;8:8. doi: 10.1186/1471-2156-8-8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lieschke GJ, Currie PD. Animal models of human disease: zebrafish swim into view. Nat Rev Genet. 2007;8:353–367. doi: 10.1038/nrg2091. [DOI] [PubMed] [Google Scholar]
- Lindsay MA. Target discovery. Nat Rev Drug Discov. 2003;2:831–838. doi: 10.1038/nrd1202. [DOI] [PubMed] [Google Scholar]
- Link CD. C. elegans models of age-associated neurodegenerative diseases: lessons from transgenic worm models of Alzheimer's disease. Exp Gerontol. 2006;41:1007–1013. doi: 10.1016/j.exger.2006.06.059. [DOI] [PubMed] [Google Scholar]
- Manev H, Dimitrijevic N. Drosophila model for in vivo pharmacological analgesia research. Eur J Pharmacol. 2004;491:207–208. doi: 10.1016/j.ejphar.2004.03.030. [DOI] [PubMed] [Google Scholar]
- Matsushima Y, Adan C, Garesse R, Kaguni LS. Functional analysis by inducible RNA interference in Drosophila melanogaster. Methods Mol Biol. 2007;372:207–217. doi: 10.1007/978-1-59745-365-3_15. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Matthews KA, Kaufman TC, Gelbart WM. Research resources for Drosophila: the expanding universe. Nat Rev Genet. 2005;6:179–193. doi: 10.1038/nrg1554. [DOI] [PubMed] [Google Scholar]
- Meli R, Prasad A, Patowary A, Lalwani MK, Maini J, Sharma M, et al. FishMap: a community resource for zebrafish genomics. Zebrafish. 2008;5:125–130. doi: 10.1089/zeb.2008.0531. [DOI] [PubMed] [Google Scholar]
- Mizgireuv IV, Revskoy SY. Transplantable tumor lines generated in clonal zebrafish. Cancer Res. 2006;66:3120–3125. doi: 10.1158/0008-5472.CAN-05-3800. [DOI] [PubMed] [Google Scholar]
- Moy TI, Ball AR, Anklesaria Z, Casadei G, Lewis K, Ausubel FM. Identification of novel antimicrobials using a live-animal infection model. Proc Natl Acad Sci U S A. 2006;103:10414–10419. doi: 10.1073/pnas.0604055103. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Murphey RD, Zon LI. Small molecule screening in the zebrafish. Methods. 2006;39:255–261. doi: 10.1016/j.ymeth.2005.09.019. [DOI] [PubMed] [Google Scholar]
- Nagy A, Perrimon N, Sandmeyer S, Plasterk R. Tailoring the genome: the power of genetic approaches. Nat Genet. 2003;33:276–284. doi: 10.1038/ng1115. [DOI] [PubMed] [Google Scholar]
- Nasevicius A, Ekker SC. Effective targeted gene ‘knockdown’ in zebrafish. Nat Genet. 2000;26:216–220. doi: 10.1038/79951. [DOI] [PubMed] [Google Scholar]
- Neely MN, Pfeifer JD, Caparon M. Streptococcus-zebrafish model of bacterial pathogenesis. Infect Immun. 2002;70:3904–3914. doi: 10.1128/IAI.70.7.3904-3914.2002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nichols CD. Drosophila melanogaster neurobiology, neuropharmacology, and how the fly can inform central nervous system drug discovery. Pharmacol Ther. 2006;112:677–700. doi: 10.1016/j.pharmthera.2006.05.012. [DOI] [PubMed] [Google Scholar]
- Nornes S, Newman M, Wells S, Verdile G, Martins RN, Lardelli M. Independent and cooperative action of Psen2 with Psen1 in zebrafish embryos. Exp Cell Res. 2009;315:2791–2801. doi: 10.1016/j.yexcr.2009.06.023. [DOI] [PubMed] [Google Scholar]
- Nusslein-Volhard C, Wieschaus E. Mutations affecting segment number and polarity in Drosophila. Nature. 1980;287:795–801. doi: 10.1038/287795a0. [DOI] [PubMed] [Google Scholar]
- Ogura E, Okuda Y, Kondoh H, Kamachi Y. Adaptation of GAL4 activators for GAL4 enhancer trapping in zebrafish. Dev Dyn. 2009;238:641–655. doi: 10.1002/dvdy.21863. [DOI] [PubMed] [Google Scholar]
- Page AP, Johnstone IL. The cuticle. WormBook. 2007:1–15. doi: 10.1895/wormbook.1.138.1. 19 March. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Parng C, Seng WL, Semino C, McGrath P. Zebrafish: a preclinical model for drug screening. Assay Drug Dev Technol. 2002;1:41–48. doi: 10.1089/154065802761001293. [DOI] [PubMed] [Google Scholar]
- Patton EE, Widlund HR, Kutok JL, Kopani KR, Amatruda JF, Murphey RD, et al. BRAF mutations are sufficient to promote nevi formation and cooperate with p53 in the genesis of melanoma. Curr Biol. 2005;15:249–254. doi: 10.1016/j.cub.2005.01.031. [DOI] [PubMed] [Google Scholar]
- Peng H, Long F, Zhou J, Leung G, Eisen MB, Myers EW. Automatic image analysis for gene expression patterns of fly embryos. BMC Cell Biol. 2007;8:S7. doi: 10.1186/1471-2121-8-S1-S7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Perrimon N, Friedman A, Mathey-Prevot B, Eggert US. Drug-target identification in Drosophila cells: combining high-throughout RNAi and small-molecule screens. Drug Discov Today. 2007;12:28–33. doi: 10.1016/j.drudis.2006.10.006. [DOI] [PubMed] [Google Scholar]
- Peterson RT, Nass R, Boyd WA, Freedman JH, Dong K, Narahashi T. Use of non-mammalian alternative models for neurotoxicological study. Neurotoxicology. 2008;29:546–555. doi: 10.1016/j.neuro.2008.04.006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pulak R. Techniques for analysis, sorting, and dispensing of C. elegans on the COPAS flow-sorting system. Methods Mol Biol. 2006;351:275–286. doi: 10.1385/1-59745-151-7:275. [DOI] [PubMed] [Google Scholar]
- Putcha GV, Johnson EM., Jr Men are but worms: neuronal cell death in C elegans and vertebrates. Cell Death Differ. 2004;11:38–48. doi: 10.1038/sj.cdd.4401352. [DOI] [PubMed] [Google Scholar]
- Ramot D, Johnson BE, Berry TL, Jr, Carnell L, Goodman MB. The Parallel Worm Tracker: a platform for measuring average speed and drug-induced paralysis in nematodes. Plos One. 2008;3:e2208. doi: 10.1371/journal.pone.0002208. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Reiser M. The ethomics era? Nat Methods. 2009;6:413–414. doi: 10.1038/nmeth0609-413. [DOI] [PubMed] [Google Scholar]
- Reiter LT, Potocki L, Chien S, Gribskov M, Bier E. A systematic analysis of human disease-associated gene sequences in Drosophila melanogaster. Genome Res. 2001;11:1114–1125. doi: 10.1101/gr.169101. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Restif C, Metaxas D. Tracking the swimming motions of C. elegans worms with applications in aging studies. Med Image Comput Comput Assist Interv Int Conf Med Image Comput Comput Assist Interv. 2008;11:35–42. doi: 10.1007/978-3-540-85988-8_5. [DOI] [PubMed] [Google Scholar]
- Rieckher M, Kourtis N, Pasparaki A, Tavernarakis N. Transgenesis in Caenorhabditis elegans. Methods Mol Biol. 2009;561:21–39. doi: 10.1007/978-1-60327-019-9_2. [DOI] [PubMed] [Google Scholar]
- Robert VJ, Katic I, Bessereau JL. Mos1 transposition as a tool to engineer the Caenorhabditis elegans genome by homologous recombination. Methods. 2009;49:263–269. doi: 10.1016/j.ymeth.2009.02.013. [DOI] [PubMed] [Google Scholar]
- Rocke J, Lees J, Packham I, Chico T. The zebrafish as a novel tool for cardiovascular drug discovery. Recent Pat Cardiovasc Drug Discov. 2009;4:1–5. doi: 10.2174/157489009787260043. [DOI] [PubMed] [Google Scholar]
- Rogalski TM, Mullen GP, Bush JA, Gilchrist EJ, Moerman DG. UNC-52/perlecan isoform diversity and function in Caenorhabditis elegans. Biochem Soc Trans. 2001;29:171–176. doi: 10.1042/0300-5127:0290171. [DOI] [PubMed] [Google Scholar]
- Rohde CB, Zeng F, Gonzalez-Rubio R, Angel M, Yanik MF. Microfluidic system for on-chip high-throughput whole-animal sorting and screening at subcellular resolution. Proc Natl Acad Sci U S A. 2007;104:13891–13895. doi: 10.1073/pnas.0706513104. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rosato E, Kyriacou CP. Analysis of locomotor activity rhythms in Drosophila. Nat Protoc. 2006;1:559–568. doi: 10.1038/nprot.2006.79. [DOI] [PubMed] [Google Scholar]
- Sabaliauskas NA, Foutz CA, Mest JR, Budgeon LR, Sidor AT, Gershenson JA, et al. High-throughput zebrafish histology. Methods. 2006;39:246–254. doi: 10.1016/j.ymeth.2006.03.001. [DOI] [PubMed] [Google Scholar]
- Schlegel A, Stainier DY. Lessons from ‘lower’ organisms: what worms, flies, and zebrafish can teach us about human energy metabolism. Plos Genet. 2007;3:e199. doi: 10.1371/journal.pgen.0030199. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Schmid A, DiDonato CJ. Animal models of spinal muscular atrophy. J Child Neurol. 2007;22:1004–1012. doi: 10.1177/0883073807305667. [DOI] [PubMed] [Google Scholar]
- Segalat L. Invertebrate animal models of diseases as screening tools in drug discovery. ACS Chem Biol. 2007a;2:231–236. doi: 10.1021/cb700009m. [DOI] [PubMed] [Google Scholar]
- Segalat L. Loss-of-function genetic diseases and the concept of pharmaceutical targets. Orphanet J Rare Dis. 2007b;2:30. doi: 10.1186/1750-1172-2-30. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sharma P, Keane J, O'Kane CJ, Asztalos Z. Automated measurement of Drosophila jump reflex habituation and its use for mutant screening. J Neurosci Methods. 2009;182:43–48. doi: 10.1016/j.jneumeth.2009.05.024. [DOI] [PubMed] [Google Scholar]
- Sieburth D, Ch'ng Q, Dybbs M, Tavazoie M, Kennedy S, Wang D, et al. Systematic analysis of genes required for synapse structure and function. Nature. 2005;436:510–517. doi: 10.1038/nature03809. [DOI] [PubMed] [Google Scholar]
- Sifri CD, Begun J, Ausubel FM. The worm has turned – microbial virulence modeled in Caenorhabditis elegans. Trends Microbiol. 2005;13:119–127. doi: 10.1016/j.tim.2005.01.003. [DOI] [PubMed] [Google Scholar]
- Sleno L, Emili A. Proteomic methods for drug target discovery. Curr Opin Chem Biol. 2008;12:46–54. doi: 10.1016/j.cbpa.2008.01.022. [DOI] [PubMed] [Google Scholar]
- Smialowska A, Baumeister R. Presenilin function in Caenorhabditis elegans. Neurodegener Dis. 2006;3:227–232. doi: 10.1159/000095260. [DOI] [PubMed] [Google Scholar]
- Solnica-Krezel L, Schier AF, Driever W. Efficient recovery of ENU-induced mutations from the zebrafish germline. Genet. 1994;136:1401–1420. doi: 10.1093/genetics/136.4.1401. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Song Y, Cone RD. Creation of a genetic model of obesity in a teleost. FASEB J. 2007;21:2042–2049. doi: 10.1096/fj.06-7503com. [DOI] [PubMed] [Google Scholar]
- Spitsbergen JM, Tsai HW, Reddy A, Miller T, Arbogast D, Hendricks JD, et al. Neoplasia in zebrafish (Danio rerio) treated with N-methyl-N'-nitro-N-nitrosoguanidine by three exposure routes at different developmental stages. Toxicol Pathol. 2000;28:716–725. doi: 10.1177/019262330002800512. [DOI] [PubMed] [Google Scholar]
- Sprague J, Clements D, Conlin T, Edwards P, Frazer K, Schaper K, et al. The Zebrafish Information Network (ZFIN): the zebrafish model organism database. Nucleic Acids Res. 2003;31:241–243. doi: 10.1093/nar/gkg027. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Spring DR. Chemical genetics to chemical genomics: small molecules offer big insights. Chem Soc Rev. 2005;34:472–482. doi: 10.1039/b312875j. [DOI] [PubMed] [Google Scholar]
- Stern HM, Zon LI. Cancer genetics and drug discovery in the zebrafish. Nat Rev Cancer. 2003;3:533–539. doi: 10.1038/nrc1126. [DOI] [PubMed] [Google Scholar]
- Stiernagle T. Maintenance of C. elegans. WormBook. 2006:1–11. doi: 10.1895/wormbook.1.101.1. 11 February. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Stilwell GE, Saraswati S, Littleton JT, Chouinard SW. Development of a Drosophila seizure model for in vivo high-throughput drug screening. Eur J Neurosci. 2006;24:2211–2222. doi: 10.1111/j.1460-9568.2006.05075.x. [DOI] [PubMed] [Google Scholar]
- Streisinger G, Coale F, Taggart C, Walker C, Grunwald DJ. Clonal origins of cells in the pigmented retina of the zebrafish eye. Dev Biol. 1989;131:60–69. doi: 10.1016/s0012-1606(89)80038-7. [DOI] [PubMed] [Google Scholar]
- Sugimoto A. High-throughput RNAi in Caenorhabditis elegans: genome-wide screens and functional genomics. Differentiation. 2004;72:81–91. doi: 10.1111/j.1432-0436.2004.07202004.x. [DOI] [PubMed] [Google Scholar]
- Tenor JL, McCormick BA, Ausubel FM, Aballay A. Caenorhabditis elegans-based screen identifies Salmonella virulence factors required for conserved host-pathogen interactions. Curr Biol. 2004;14:1018–1024. doi: 10.1016/j.cub.2004.05.050. [DOI] [PubMed] [Google Scholar]
- Timmons L, Fire A. Specific interference by ingested dsRNA. Nature. 1998;395:854. doi: 10.1038/27579. [DOI] [PubMed] [Google Scholar]
- Tran TC, Sneed B, Haider J, Blavo D, White A, Aiyejorun T, et al. Automated, quantitative screening assay for antiangiogenic compounds using transgenic zebrafish. Cancer Res. 2007;67:11386–11392. doi: 10.1158/0008-5472.CAN-07-3126. [DOI] [PubMed] [Google Scholar]
- Tsechpenakis G, Bianchi L, Metaxas D, Driscoll M. A novel computational approach for simultaneous tracking and feature extraction of C. elegans populations in fluid environments. IEEE Trans Biomed Eng. 2008;55:1539–1549. doi: 10.1109/TBME.2008.918582. [DOI] [PubMed] [Google Scholar]
- Tsibidis GD, Tavernarakis N. Nemo: a computational tool for analyzing nematode locomotion. BMC Neurosci. 2007;8:86. doi: 10.1186/1471-2202-8-86. [DOI] [PMC free article] [PubMed] [Google Scholar]
- van der Plas MC, Pilgram GS, de Jong AW, Bansraj MR, Fradkin LG, Noordermeer JN. Drosophila Dystrophin is required for integrity of the musculature. Mech Dev. 2007;124:617–630. doi: 10.1016/j.mod.2007.04.003. [DOI] [PubMed] [Google Scholar]
- Venken KJ, Bellen HJ. Emerging technologies for gene manipulation in Drosophila melanogaster. Nat Rev Genet. 2005;6:167–178. doi: 10.1038/nrg1553. [DOI] [PubMed] [Google Scholar]
- Vogt A, Cholewinski A, Shen X, Nelson SG, Lazo JS, Tsang M, et al. Automated image-based phenotypic analysis in zebrafish embryos. Dev Dyn. 2009;238:656–663. doi: 10.1002/dvdy.21892. [DOI] [PMC free article] [PubMed] [Google Scholar]
- West DB, Iakougova O, Olsson C, Ross D, Ohmen J, Chatterjee A. Mouse genetics/genomics: an effective approach for drug target discovery and validation. Med Res Rev. 2000;20:216–230. doi: 10.1002/(sici)1098-1128(200005)20:3<216::aid-med6>3.0.co;2-0. [DOI] [PubMed] [Google Scholar]
- Westerfield M. The Zebrafish Book. Eugene, OR: University of Oregon Press; 1995. [Google Scholar]
- Whitworth AJ, Wes PD, Pallanck LJ. Drosophila models pioneer a new approach to drug discovery for Parkinson's disease. Drug Discov Today. 2006;11:119–126. doi: 10.1016/S1359-6446(05)03693-7. [DOI] [PubMed] [Google Scholar]
- Wienholds E, Schulte-Merker S, Walderich B, Plasterk RH. Target-selected inactivation of the zebrafish rag1 gene. Science. 2002;297:99–102. doi: 10.1126/science.1071762. [DOI] [PubMed] [Google Scholar]
- Winter MJ, Redfern WS, Hayfield AJ, Owen SF, Valentin JP, Hutchinson TH. Validation of a larval zebrafish locomotor assay for assessing the seizure liability of early-stage development drugs. J Pharmacol Toxicol Methods. 2008;57:176–187. doi: 10.1016/j.vascn.2008.01.004. [DOI] [PubMed] [Google Scholar]
- Zhang JH, Chung TD, Oldenburg KR. A simple statistical parameter for use in evaluation and validation of high throughput screening assays. J Biomol Screen. 1999;4:67–73. doi: 10.1177/108705719900400206. [DOI] [PubMed] [Google Scholar]