

Abstract
Figure-ground organization involves assignment of edges to a figural shape on one or the other side of each dividing edge. Established visual cues for edge-assignment primarily concern relatively local rather than contextual factors. Here we show that assignment for a locally-unbiased edge can be affected by assignment of a remote contextual edge that has its own locally-biased assignment. We find that such propagation of edge-assignment from the biased remote context occurs only when the biased and unbiased edges are grouped. This new principle, whereby grouping constrains propagation of figural edge-assignment, emerges from both subjective reports and from an objective short-term edge-matching task. It generalizes from moving displays involving grouping by common fate and collinearity, to static displays with grouping by similarity of edge-contrast polarity, or apparent occlusion. Our results identify a new contextual influence upon edge-assignment. They also identify a new mechanistic relation between grouping and figure-ground processes, whereby grouping between remote elements can constrain propagation of edge-assignment between those elements.
Edge-assignment is a key aspect of figure-ground organization. When a dividing edge is assigned to one of its adjacent regions, that region becomes ‘figural’ and is perceived as shaped along the edge and in front, while the other adjacent ‘ground’ region is not shaped and appears behind the figural region (Baylis & Cale, 2001; Rubin, 1921; Driver & Baylis, 1996). This is illustrated by the change in perceived shape associated with reversals of figural edge-assignment, as for the well-known faces-vase example. Furthermore, figural edge-assignment affects the way that visual input is processed by brain regions involved in shape representation and object recognition (e.g., Appelbaum et al., 2006; Baylis & Driver, 2001; Kourtzi & Kanwisher, 2001).
Established image cues for determining figural edge-assignment typically involve relatively local edge or region properties, such as convexity (Kanizsa & Gerbino, 1976; Metzger, 1953; Stevens & Brookes, 1988), symmetry (Kanizsa & Gerbino, 1976; Rubin, 1921), relative size and contrast (Palmer, 1999; Rubin, 1921), relative position in the scene (Hulleman & Humphreys, 2004; Vecera et al., 2002), or local edge-region grouping relationships (Palmer & Brooks, 2008), among other image-based factors. Higher-level factors such as attention (Baylis & Driver, 1995; Driver & Baylis, 1996; Vecera et al., 2004; although see Huang & Pashler, 2009) and previous experience (Peterson & Gibson, 1994b; Peterson & Gibson, 1994a; Peterson, 1994) have also been suggested to play some role in determining figural edge-assignment.
However, the possible influences of distant, contextual parts of an image upon figural edge-assignment elsewhere in that image remain less explored. Recently, Peterson and Salvagio (2008) demonstrated that the effectiveness of convexity for local figural edge-assignment can be affected by more global context. For a bipartite display (i.e., comprising two adjacent regions) with a single critical dividing edge, the more convex region was only weakly biased toward figural status. However, when additional surrounding edges and regions were added together, to form a chain of convex/concave/convex regions, the convex regions became significantly more likely to be chosen as figural. Evidently the presence of the convexity cue at multiple edges affected the impact of that cue. Interestingly, this effect of context only occurred when the concave regions that intervened between the successive convex regions were similarly colored, thereby providing a homogeneous background made up of all the commonly colored concave regions. These results indicate that, under certain conditions, the strength of existing figural biases within a scene may be enhanced contextually by other edges/regions in the scene. However, we know of no examples to date in which figural edge-assignment for a locally unbiased edge (i.e., where no local figural cues are present) has been shown unequivocally to be determined by figural assignment of remote contextual edges elsewhere in the image (whose figural assignment is determined by their own local biasing factors, that do not apply for the unbiased edge elsewhere). Thus, although Peterson & Salvagio have provided an initial contextual demonstration that a weak local cue can have more impact when combined with matching context, it still remains unknown whether figural assignment for a locally unbiased edge can be determined by propagation of figural assignment from a remote but locally-biased contextual edge.
Here we demonstrate that figural edge-assignment of a locally ambiguous edge (i.e., with no local cues to figural assignment along that edge, nor in its adjoining regions) can in fact be determined by assignment of remote contextual edges. We also investigate the boundary conditions for contextual edges to propagate their influence to other edges in this way. We propose that perceptual grouping can constrain whether or not a given contextual edge will influence figural edge-assignment of the locally ambiguous edge. Specifically, when an unbiased-edge is grouped with a separate locally-biased-edge, we predict that the figural assignment of the latter will propagate to the former. But we expect such propagation to be curtailed when biased and unbiased edges are not grouped together. This prediction arises intuitively from an ecological rationale. When two edges are grouped together, this indicates that those two edges are likely to belong to the same external source, and hence may also be likely to have congruent figure/ground (depth) relations in the physical world, along the grouped edges. By contrast, for edges that are not grouped together by the visual system, corresponding figure-ground assignment may be less likely.
From a more formal perspective, the new prediction that we test for the first time here also appears to be in line with some recent computational models of edge-assignment, which in practice rely on “grouping” operations in order to propagate depth information from local t-junction cues to other locations along a complete or partially complete contour (Craft et al., 2007; Heitger et al., 1998; Peterhans & Heitger, 2001; Thielscher & Neumann, 2008). Here we test for human vision whether grouping operations do indeed serve as a mechanism for constraining propagation of figure-ground assignment from one contour to another in separate parts of the image.
Our proposal that grouping should constrain propagation of edge-assignment in this specific way appears to contrast with some other influential proposals about the relationship between figural edge-assignment and grouping. For instance, Palmer and Rock (1994b) initially proposed that figure-ground assignment invariably arises before perceptual grouping, so that the latter should presumably not influence the former (although they later finessed this to acknowledge a possible role for interactive activation between successive stages of visual processing; see Palmer & Rock (1994b; 1994a). Their initial claim was largely based on an argument from first principles, which had assumed that figure-ground assignment must always operate first, to provide candidate items to undergo grouping. Our experiments directly test whether instead grouping can in fact constrain figure-ground assignment (indicating the reverse influence), to determine whether or not figure-ground assignment will be propagated from one contour to another. Although ‘figure-ground organization’ (i.e. edge-assignment) and ‘grouping’ (i.e. perceptual linking of separate elements in an image) are often jointly discussed in many textbook chapters on Gestalt psychology, typically little or no explicit mechanistic relation is specified between the two types of process in such chapters, unlike the specific proposal we test here.
To assess our proposal that figure-ground organization should propagate from contextual contours to unbiased contours, specifically only when the latter are grouped with the former, we created novel displays comprising two main ‘sections’ plus an occluder separating them (Figure 1A). The top and bottom bipartite sections each comprised two differently-colored regions (shown as black and white in Figure 1A), separated by a vertical contrast edge. The vertical edge for the locally biased-section (bottom section in Figure 1A) was assigned by local figure-ground cues towards one or the other side (see below for a description of the determining local cues, which involved moving dots in our initial experiments; but see the later experiments for different cues in static displays). The other unbiased-section (top section in Figure 1A) had no such local edge-assignment cues. A rectangular occluder (shown as gray in Figure 1A, actually red in our initial experiments) clearly separated the biased and unbiased-sections in the displayed image. The original rationale for this occluder was to demarcate a clear separation in the image between biased- and unbiased-sections, but we return to its possible perceptual role later.
Figure 1.

(A) Example static frame from the dynamic displays in Experiment 1A. The top (above rectangular occluder) black-and-white section is the unbiased-section (no dots) in this example, whereas the bottom section (below rectangular occluder) is the biased-section with texture dots on both sides of the edge. Edge-assignment for the edge within the biased-section was determined by local edge-assignment cues, namely the relationship between the motion of the edge and the dots in the two regions (see main text). The color of the central ‘occluding’ horizontal bar was red in the actual displays. Animated color versions of all displays are available in the Supplemental Materials. (B) Annotated cartoon of the motion for different sections of the display in the edges-grouped with similar-region-color condition. Arrows within a region indicate relative direction of motion for dotted region texture. Arrows above or below a section indicate the relative motion direction of the vertical dividing edge for that section. The frequencies listed indicate the rate of oscillation associated with that display element (e.g. 1.0 Hz below a region arrow indicates a 1.0 Hz oscillation for texture dots in that region). “F” and “G” indicate anticipated figural or ground status (respectively) within the biased-section due to dot motion in relation to edge motion, as was confirmed by phenomenal judgments for the biased section. (C) Edge-only grouped condition, region-color-dissimilar. (D) Edges-ungrouped with region-color-similar condition. (E) Edges-ungrouped with region-color-dissimilar condition. Blue and green region colors are replaced by dark gray and light gray here. In the actual displays the blue and green were equiluminant (by flicker photometry for each participant) with the “black” (dark gray) and “white” (light gray) of the unbiased-section, respectively.
We measured whether edge-assignment for the central vertical edge in the biased-section of the display would affect edge-assignment for the central vertical edge in the unbiased-section, to produce a remote contextual effect. To test whether grouping would constrain any such propagation of edge-assignment from biased to unbiased-sections, we manipulated perceptual grouping between the central dividing edges (henceforth termed edge-grouping) of the biased and unbiased-sections. In Experiment 1, the central vertical edges within biased and-unbiased sections moved from left to right in an oscillatory manner (see animations in Supplementary Materials). These edges clearly grouped when they moved collinearly together with common fate (Figs 1B-C). In this case, we expected edge-grouping to cause propagation of local figural edge-assignment from the biased-section's dividing edge to the unbiased-section's dividing edge. On the other hand, these edges were clearly ungrouped when they moved at different rates and thus no longer shared common fate (Figures 1D-E), nor collinearity (except at the instant when they ‘crossed’). In this edge-ungrouped case, we expected that propagation of figural edge-assignment from the biased to the unbiased section would become minimal or absent. In subsequent experiments (see below), we used displays with static figure-ground cues instead, imposing other grouping manipulations, to test the generality of our findings and the new principle that they suggest.
Orthogonally to the edge-grouping manipulation described above, for completeness we also manipulated region-grouping, by color similarity (henceforth termed region-color-similarity) in Experiment 1. Analogously to edge-grouping, when the regional colors of the biased and unbiased-sections were matched (i.e. region-grouped by color similarity) we anticipated that figural edge-assignment for the central dividing edge might be more likely to propagate between the two sections than when they had differently colored regions. Regions on either side of the critical vertical edges in both the biased and unbiased-sections either shared region-color-similarity (i.e. both sections were black-at-left and white-at-right, or vice-versa, in Experiment 1; Figures 1B/D), or else they did not (unbiased-section regions were black and white, whereas the biased-section regions were blue and green, or vice versa in Experiment 1; Figures 1C/E). Manipulating edge-grouping and region-color-similarity orthogonally allowed us to assess whether only edge-grouping (initially manipulated by common fate and collinearity, but see our later experiments also) affects figural edge-assignment propagation; or whether region-color-similarity also matters, plus if these factors interact.
Experiment 1: Dynamic edge-grouping cues with subjective reports of figural assignment
In Experiment 1, we used a subjective-report procedure to measure participants' phenomenal experience of figural edge-assignment for the biased or unbiased sections of dynamic displays. On some trials, we measured the participants' perception of the biased section, in order to determine whether local cues were effective in determining figural edge-assignment as expected along the dividing edge in that section. In other trials, we measured figural edge-assignment for the unbiased-section's dividing edge instead. This allowed us to assess whether edge-assignment propagated from the biased-section's edge to the unbiased-section's edge. We manipulated the edge-grouping and region-color-similarity factors as described above in order to determine whether perceptual grouping affected propagation of subjective figural edge-assignment from the biased to the unbiased dividing edge.
Method
Participants
12 right-handed people from the local community participated (6 male and 6 female, mean age 21.4 years). All reported normal vision and gave informed consent to participate in the study.
Displays and Design
Each display comprised three sections (see five examples of alternative three-section displays in Figure 1); the top bipartite section, the bottom bipartite section, and the rectangular occluder between them. The top and bottom bipartite sections were each 5.3° square and were separated by a red rectangle (16.8° by 2.0°) centered at fixation. The vertical dividing edges within each bipartite section oscillated horizontally (the oscillation distance was 0.85°) at either 1.0 Hz or 1.5 Hz.
Edge-assignment for the biased-section's dividing edge was determined by powerful local cues to figural edge-assignment (Yonas et al., 1987; Palmer & Brooks, 2008). Biased-section regions on both sides of its dividing edge were filled with a sparse dot texture (Figure 1A, bottom). The dot texture on one side of the edge moved with the dividing edge, which should assign the edge to that region figurally (see Palmer & Brooks, 2008). The dot texture on the other side moved in counter-phase to the edge, so that region should become ground.
When the biased and unbiased-sections were related in terms of region-color-similarity, the regions in both sections were black (average 9.0 cd/m2, herein ‘black’) and white (average 53.5 cd/m2, herein ‘white’), with corresponding left-right assignments for colors within the two sections. The exact luminance for each color was determined for each participant individually by flicker photometry (Wagner & Boynton, 1972). White was made equiluminant to green and black was made equiluminant to blue. This was done to ensure that the region colors were similar in luminance to the green and blue colors used in the region-color-dissimilar conditions. In the region-color-dissimilar conditions, biased-section regions were green (53 cd/m2) and blue (8 cd/m2).
The design was a 2 × 2 within-subjects factorial design with edge-grouping and region-color-similarity as the factors. There were 32 repetitions of each of the 4 conditions within each block. Several other aspects of the displays were counterbalanced. In the biased-section, the dot-motion-determined figure was either on the left or right side of the vertical edge equiprobably. Likewise the initial direction of motion for the biased-section's dividing edge was either leftward or rightward equally often. This counterbalanced for the effects of the recently described figure-ground cue of advancing versus receding motion (Barenholtz & Tarr, 2009), which might arise in the form of initial expansion versus contraction here for either side of the moving dividing edge, from display onset. The biased-section was equally often above or below the occluder. In edge-grouped conditions, the initial direction of motion for the biased-section's edge was the same for the unbiased-section also, whereas it began with opposite motion in edge-ungrouped conditions. Edge oscillation frequency in the biased-section was either 1.0 Hz or 1.5 Hz, equiprobably. In edge-grouped conditions, the frequency was the same for the unbiased-section, whereas for edge-ungrouped conditions it was different. The color of the two regions within the unbiased-section was counterbalanced (white on left or right), and the color in the biased-section was equiprobably either the same (in region-color-similar conditions), or differed in being a green/blue combination (region-color-dissimilar conditions). In the latter case, contrast polarity across the edge (i.e. dark/light or light/dark) was counterbalanced by using either blue/green (on the left/right respectively) or green/blue instead.
Participants reported phenomenal figure-ground assignment for only one section of the display (upper or lower) in each block. Because the judged section was equiprobably biased or unbiased, there were two separate sets of results. Judgments of the biased-section allowed us to confirm whether the local figure-ground bias induced by the dots was indeed effective. Judgments of the unbiased-section provided the critical new test for any propagation of figural edge-assignment from the biased-section to the unbiased-section.
Procedure
Displays were presented on a 21-inch CRT computer monitor (60 Hz, 1280 × 1024 pixel resolution) using Presentation software (Neurobehavioral Systems, Inc., http://www.neurobs.com). Each trial began with a central fixation cross for 1000 ms followed by the 2800 ms display. Participants reported, for just one section of the display (top or bottom, blocked) whether the left or right region within that section appeared to be “in front”, using corresponding buttons. Because the judged-section was equally often biased or unbiased, there were two sets of results; biased-section judgments (assessing whether the local dot-motion biased that section's figural assignment as expected) and unbiased-section judgments (assessing any propagation from the biased-section). Eye position was monitored throughout displays using an ASL-5000 Tracker (Applied Science Laboratories, Inc.; Bedford, MA, USA; http://www.a-s-l.com). Trials with >1° deviations from fixation or with pursuit eye movements (>0.5° regular oscillatory structure) during the displays were excluded (0.9%). The procedure was approved by the UCL Psychology Department Ethics Committee.
Results and Discussion
As expected for the biased-section, participants primarily reported as figural (“in front”) the side on which the dot-texture moved together with the dividing edge. They selected this for 94.7% of judgments on the biased-section; significantly greater than 50%, t = 23.31, p < 0.001. This is as expected, in accord with the local figural edge-assignment cues favoring that region (Palmer & Brooks, 2008; Yonas et al., 1987). There were no significant effects of any of the counterbalanced factors on biased-section figure-ground judgments. At the request of a reviewer, we assessed any impact from the starting direction of motion for the dividing edge, which made one side of the section expand and the other contract (Barenholtz & Tarr, 2009) before the motion reversed. Participants chose the initially expanding region 50.3% of the time, no greater than chance, t(11) = 1.86, p < 0.09. The lack of a significant impact from this initial expansion/contraction here might be due to the much stronger impact of our intended figural edge-assignment cue for the biased-section (due to how the dots moved in relation to the dividing edge, cf. Palmer & Brooks, 2008). That dot-motion factor determined figure-ground assignment 94.7% of the time for the biased section.
For our critical new test of whether figural assignment propagated from the biased-section to the unbiased-section, the unbiased-section judgments were re-coded as either context-consistent (e.g. when the biased-section's edge should be assigned leftward via the dots, participants correspondingly reported leftward assignment for the unbiased-section), or context-inconsistent. Figure 2 plots the percentage of context-consistent responses in the four main conditions. These context-consistent responses were significantly higher when the edges were grouped, F(1,11) = 63.36, p < 0.0001 and when region-color-similarity was present, F(1,11) = 8.27, p < 0.015. These results indicate that figural edge-assignment propagated from the biased- to the unbiased-section, but only when grouping between these two sections was present. In addition to the main effects, the two factors also interacted, F(1,11) = 4.96, p < 0.04. This interaction reflects an impact of region-color-similarity only when edge-grouping was present, F(1,11) = 16.04, p < 0.002, not when the two critical vertical edges were ungrouped, p = 0.46, n.s.
Figure 2.

Experiment 1 results. The average percentage of trials on which the context-consistent side within the unbiased-section of the display was chosen as figural (i.e., in accord with the side biased as figural in the separate biased-section) is plotted as a function of edge-grouping and region-color-similarity factors. Error bars represent 95% within-subjects confidence intervals. Note the higher context-consistent responses in the edge-grouped condition, with this slightly increased when regional color is also similar.
The percentage of context-consistent responses for the unbiased-section was significantly greater than 50% only in edge-grouped conditions (for edge-and-region-grouped, t = 11.22, p < 0.001; edge-only-grouped, t = 8.11, p < 0.001), but not in edge-ungrouped conditions (region-only-grouped, p = 0.64, n.s.; neither-grouped, p = 0.87, n.s.). For completeness we report also that the counterbalanced factor of edge-contrast-similarity (i.e. same or different contrast polarity across edges in biased and unbiased-sections, which was varied intrinsically but only for region-color-dissimilar conditions) showed a very small yet reliable influence (same 61% vs. different 59%; F(1,11) = 5.21, p < 0.043).
Finally a reviewer asked us to examine any impact of the expansion/contraction on one or other side of the dividing edge, to its initial direction of motion. The initially expanding side within the unbiased-section was chosen as figural 59.5% of the time (in accord with Barenholtz & Tarr), slightly but significantly above chance, t(11) = 8.48, p < 0.001. This effect for the unbiased-section judgments (that was not significant for the biased-section) might reflect different strengths of figure-ground assignment for the two sections. Within the unbiased-section there were no local cues to edge-assignment and the propagated contextual effect that we uncover here for the first time (see Figure 2) did not reach ceiling levels. This might have left some room for modulation by other figure-ground cues, such as expansion/contraction.
The critical new aspect of our results lies in showing show that subjective figural edge-assignment of a locally-unbiased edge can be affected by the figural edge-assignment of a remote contextual biased edge. Moreover, this assignment propagates from biased to unbiased-edges only when they are grouped together (here by common fate and collinearity, but see our later experiments for different grouping cues). Region-color-similarity had some influence but only when it was not vetoed by edge-ungrouping. Same or different edge-contrast-polarity also had a minor influence, but was only varied in a minor way in Experiment 1, i.e. only for the region-color-dissimilar conditions, since contrast polarity had not been of a priori interest to us.
Experiment 2: Control for salient dots potentially attracting attention when figural assignment is measured by subjective report
Experiment 1 demonstrated that figural edge-assignment along an edge with no local cues to edge-assignment (e.g. our unbiased-section edge) can be determined by edge-assignment along an edge elsewhere in the image (i.e., our biased-section edge), provided that this contextual edge is grouped with the unbiased edge. Our interpretation of this finding is that perceptual grouping can gate propagation of figural edge-assignment between remote parts of an image (here the biased and unbiased sections). In Experiment 1, the local edge-assignment cues within the biased-section involved moving dots. These dots were presented on both sides of the dividing edge in the biased-section (see Figure 1A), but the dots on one side moved together with the central edge (see Supplementary Materials animations) causing that side to become figural, in line with previous results (Palmer & Brooks, 2008; Yonas et al., 1987). Dots on the other side of the biased-section moved in counter-phase to the dividing edge and thus caused that side to be perceived as ground. Thus, in our initial conditions, regions on both sides of the biased-edge had moving dots. Nevertheless, one might still potentially be concerned that attention could be drawn to salient moving dots (Franconeri & Simons, 2005; Hillstrom & Yantis, 1994), perhaps more so on the figural side. Furthermore, there have been some suggestions that familiar figural regions may attract attention (Nelson & Palmer, 2007) although it is unclear if non-familiar regions (like those used in the present study) can attract attention by their figural status alone, a point we return to in the General Discussion. Returning to interpretation of the current Experiment 1, if the dots on one side indeed became more salient and thereby attracted attention, one might then in principle argue that it could be attention towards the side of more salient dots in biased-section, rather than figural assignment per se, that ultimately leads to an influence on the unbiased section. This might conceivably occur, for instance, via object-based or grouping-related attention mechanisms (e.g., Egly et al., 1994) that might conceivably allow attention to spread to the unbiased-section.
To address this concern for the specific issue of salient dots potentially attracting attention towards one side, in Experiment 2 we now used the dots within the biased-section's regions to manipulate which side of the dividing edge should be most ‘salient’ (in terms of containing moving dots) within that biased-section, separately from which region should be figural. In some displays we now had moving dots present on only one side of the biased-section, rather than on both. This is particularly telling when the dots appeared only on the ground side in the biased-section (henceforth termed the ground-salient condition). In this case, the dots move in counter-phase to the dividing edge's motion and thus should still make that region appear as the ground , leading to figural assignment of the dividing edge to the other region (with no dots). But any account in terms of attention- capture by salient moving dots should presumably then clearly favor the biased-section's ground side (now containing the only moving dots) rather than the anticipated figural side (now with none). For an account solely in terms of attention being attracted to salient moving dots (see also our General Discussion for consideration of further accounts), this should lead to the opposite prediction to that expected from our own perspective in terms of grouping-mediated propagation of edge-assignment.
We compared the ground-salient condition to a condition with dots on both sides (salience-balanced, the same as the stimuli in Experiment 1), and also for completeness to a condition with the moving dot texture appearing on only the biased-section's figural side (figure-salient). If our grouping-mediating propagation of figural edge-assignment account is correct, then the pattern of results should be similar for all three conditions. In particular, having dots present only on the ground side of the biased section should not reverse the pattern of results for the unbiased section, provided that the expected figural assignment still applies for the biased section. By contrast, if attention capture by salient moving dots is critical, then the ground-salient condition in particular should lead to a reversed outcome for the unbiased section.
Method
Participants
16 people from the local community participated (7 male, 9 female, average age 26.1 years). All reported normal vision and gave informed consent. None of these participants had taken part in Experiment 1.
Displays and Design
In the salience-balanced conditions, the displays were exactly as in Experiment 1. In the ground-salient condition, the displays were as for those in Experiment 1, except that the moving dots on the figural side of the biased section were now removed so that only those moving in counter-phase to the edge (in the ground side) remained. In the figure-salient condition, the displays were as for Experiment 1 except that the moving dots on the ground side of the biased section were removed, so that only those moving in common fate with the edge (on the figural side) remained. We note in passing that having moving dots only on the figural side might slightly reduce the strength of figural assignment by the dots (since this condition removes the dots which move in counterphase to the dividing edge). By contrast, in terms of possible attention-capture the figure-salient condition should presumably instead only act to increase any effects. All other aspects of the displays were identical to those in Experiment 1.
The design was a 2 × 2 × 3 within-subjects factorial design. The first two factors were the same edge-grouping and region-color-similarity factors as in Experiment 1. The third factor specified the salient side due to the dots: figure-salient, ground-salient, or salience-balanced (only the latter having dots on both sides of the dividing edge in the biased section, equivalent to Experiment 1). The counterbalancing of other display factors was the same as in Experiment 1. There were 32 repeated measures per condition within each block.
Procedure
The procedure was the same as in Experiment 1. In one block the participant judged the top section and in the other block they judged the lower section. There were 4 breaks in each block. Trials with >1° deviations from fixation or with pursuit eye movements (>0.5° regular oscillatory structure) during the displays were excluded (intersubject mean of 1.8% trials excluded).
Results and Discussion
The local edge-assignment cues due to the dot motion within the biased-section were effective (significantly greater than 50%) at determining edge-assignment within that section in all three dot-salience conditions; figure-salient, 87.2%, t(15) = 17.11, p < 0.0001; ground-salient, 97.9%, t(15) = 96.23, p < 0.0001; salience-balanced, 99.2%, t(15) = 181.64, p < 0.0001. Notably the local edge-assignment cue from dot motion remained equally effective in the balanced-salience and ground-salient conditions. Its influence was somewhat reduced in the figure-salient condition (significantly so in comparison with either of the other two dot condition, both, t(15) > 4.9, p < 0.001) but still remained significant. As briefly anticipated in the introduction to this experiment, this reduction may have occurred because (contrary to an account solely in terms of attention capture by salient dots) the counter-phase motion of dots on the ground side may provide a particularly strong figural-assignment depth cue (Palmer & Brooks, 2008), when present.
As for Experiment 1, at the request of a reviewer we also analyzed any impact from the counterbalanced cue of initial expansion versus contraction on one of other side of the dividing edge in the biased section, due to its initial direction of motion before the oscillatory reversal. We did this separately for each of the dot salience conditions. In the salience-balanced condition, the initially expanding region was chosen 50.1% of the time, no greater than chance, t(15) = 0.56, p < 0.58. In the ground-salient condition, the initially expanding region was chosen 50.6% of the time, again no greater than chance, t(15) = 1.21, p < 0.24. The results in these two conditions are consistent with Experiment 1's biased-section results. In contrast, there was a small but significant effect of initial expansion versus contraction for biased-section judgments in the figure-salient condition, t(15) = 2.23, p < 0.04, with the initially expanding side selected on 52.6% of trials. We note that this same figure-salient condition showed slightly less impact of the local dot-motion figure-ground cue (presumably due to the removal of dots moving in counter-phase to the dividing edge), and so was not as close to ceiling as the other two conditions. This might have led to more sensitivity to any impact from expansion/contraction at onset (see also our Discussion of Experiment 1), although we note that expansion/contraction was fully counterbalanced with respect to the main factors of interest here, so cannot explain any of our critical results.
We turn next to the critical judgments for the unbiased section (Figure 3) that allow us to measure any propagation of figural assignment from the biased section. In all salience conditions there was always an impact of edge-grouping: for salience-balanced, F(1,15) = 201.05, p < 0.0001; ground-salient, F(1,15) = 285.53, p < 0.0001; figure-salient, F(1,15) = 102.20, p < 0.0001. There were also main effects of region-color-similarity (for salience-balanced, F(1,15) = 4.7, p < 0.047; ground-salient, F(1,15) = 5.43, p < 0.034; figure-salient, F(1,15) = 5.94, p < 0.028); plus the usual interaction between edge-grouping and region-color-similarity (for salience-balanced, F(1,15) = 9.56, p < 0.007; ground-salient, F(1,15) = 10.69, p < 0.005; figure-salient, F(1,15) = 41.03, p < 0.0001). These interactions all arose because (as in Experiment 1 also, compare Figs 2 and 3), region-color-similarity only had an impact when the edges were grouped. The overall 2×2×3 within-subjects ANOVA showed main effects of edge-grouping, F(1,15) = 441.96, p < 0.0001; region-color-similarity, F(1,15) = 11.56, p < 0.004; and the interaction of these two factors, F(1,15) = 36.21, p < 0.0001. For all three dot-salience conditions, the above statistics confirm that the basic pattern of results we saw in Experiment 1 (i.e. that edge-assignment propagates from the biased to the unbiased edge when edge-grouping is present and is further facilitated by additional region-grouping) is still found even with the moving dots on just one or other side of the dividing edge in the biased section, rather than on both sides. Most notably, the pattern of results for the unbiased section was unchanged when removing the dots on the figural side of the biased section, even though in terms of putative attention-capture by salient moving dots, that situation might have been expected to reverse the outcome for the unbiased section.
Figure 3.

Experiment 2 results. (A) Results for the salience-balanced condition (i.e. moving dots on both sides in biased-section). (B) Results for the ground-salient condition (i.e. moving dots now only on ground side in biased-section). (C) Results for the figure-salient condition (moving dots now only on figural side in biased-section). Error bars represent 95% within-subjects confidence intervals. Note the similar outcome in all three salience conditions, namely higher context-consistent responses in the edge-grouped condition, with this slightly increased when regional color is also similar.
As noted above, there was a small effect of dot-salience on the strength of the local cue within the biased-section (i.e. this local cue was somewhat weaker in the figure-salient condition, presumably due to the lack of any counter-phase dot motion). Given that figural edge-assignment in the biased section must be present in order to propagate to the unbiased section, it follows that any changes in the strength of the local cue for the biased section may also propagate through to the unbiased section (provided, of course, that edge-grouping and region-grouping are present to allow such propagation). Consistent with this, there was a main effect of dot-salience for the biased section results, F(2,30) = 6.17, p < 0.006, reflecting somewhat lower context-consistent responses in the figure-salient condition. Moreover, consistent with our account, this reduction was most apparent in the edge-grouped conditions, leading to an interaction with edge-grouping, F(2,15) = 11.29, p < 0.0001. There was no higher-level three-way interaction (p > 0.75, n.s.)
As in the unbiased-section judgments of Experiment 1, we also found a small but reliable impact effect of the initial expansion/contraction cue. Participants chose the initially expanding region as figure significantly more than 50% in all three dot-salience conditions; for salience-balanced, 52.9%, t(15) = 2.47, p < 0.02; figure-salient, 54.3%, t(15) = 2.36, p < 0.03; ground-salient, 55.2%, t(15) = 3.45, p < 0.004. We note once again that this initial expansion/contraction factor (determined by the initial direction of motion of the dividing edge, prior to the oscillatory change in direction) was fully counterbalanced with respect to our main factors of interest, and therefore cannot explain the effects of the latter factors.
Figure 3 displays a total of three replications of our key findings from Experiment 1 (cf. Fig 2). Once again, figural edge-assignment of the biased-edge propagated to the unbiased-edge only when these edges were grouped (here by common fate and collinearity, but see our later experiments for other grouping cues); while region-color-similarity exerted a small but reliable influence on such propagation only when the edges were grouped. Most importantly, all of this still occurred in the ground-salient condition, in which the salient dots should if anything presumably attract attention towards the ground rather than figural region of the biased section. If such attention-capture by moving dots in itself mediated the spread of figural edge-assignment, then one should expect the effects to reverse in the ground-salient condition because the presence of moving dots should now favor the side opposite to the figural side. But in fact, the effects were just as strong in the ground-salient condition as they were in the balanced-salience condition (which further replicated Experiment 1).
In the figure-salient condition, context-consistent responses were slightly lower, but this was in accord with the slightly weakened local impact of the dot-motion cue within the biased-section of those particular displays (which now had no dots in counter-phase to the dividing edge of the biased section). Nevertheless, even the (correspondingly) slightly weakened impact on the unbiased section in this situation still showed a similar overall pattern to the other salience conditions (see Figure 3). Once again, figural edge-assignment propagated to the unbiased-section's edge when edge-grouping was present between the dividing edges in the two sections, with this effect being modulated by region-color-similarity but only in the presence of edge-grouping.
Experiment 3: Objective short term edge-matching as an index for the consequences of figural assignment
In Experiment 3, we sought to generalize our key findings to an objective consequence of edge-assignment, rather than for phenomenal judgments as studied in Experiments 1 and 2. Driver & Baylis (1996) introduced a short-term edge-matching (STEM) procedure that reflects figural edge-assignment. Their participants judged whether a curvy edge in a ‘probe’ display matched the curvy dividing edge in a preceding ‘prime’ display. Performance was better if the probe display's edge had the same figural edge-assignment as in the preceding prime display, than when a ‘ground’ probe was used instead (though in either case, equivalent curvy edges were presented).
We adapted this STEM procedure here so that participants now saw a prime like the displays in our Experiment 1, but the unbiased-section's dividing edge was now ‘curvy’ rather than straight (Figure 4A). Each prime was followed by either of two types of probe-pair displays (see Figure 4B or 4C), each containing two shapes with curvy edges. For both types, one of the curvy probe edges matched the curvy edge in the unbiased-section of the preceding prime display. The other was a previously unseen foil. The objective task was to select the matching probe. Each probe-pair either comprised ‘figure’ probes (i.e. with their curvy edges assigned in the same direction as the edge in the preceding prime biased-section); or ‘ground’ probes, both with edges assigned to the other side. The task was to pick whether the upper or lower edge in a given probe display had the same curvy edge as the preceding unbiased prime. This now allowed us to test for objective consequences (rather than subjective reports, cf. Experiments 1 and 2) of our grouping manipulations, upon propagation of figural edge-assignment from the biased-section to the locally-unbiased-section of the prime display. Under the appropriate grouping conditions for the prime displays, ‘figure’ probes should lead to more efficient matching performance than ‘ground’ probes (analogously to Driver & Baylis, 1996), if figural edge-assignment does indeed propagate from biased to unbiased-sections when grouped, and if this has objective consequences for edge-assignment. If figural edge-assignment does not propagate to the unbiased-section then we should see no difference between ‘figure’ and ‘ground’ probe performance in any of the conditions.
Figure 4.

Example displays from Experiment 3 for the STEM task, see main text for description of that task. (A) Prime display shown with curved dividing edge in upper unbiased-section. (B) ‘Figural’ probes, assuming that left side was defined (by the biased section moving dots) as figural in preceding prime. The top probe shape in B matches the prime curvy edge in A. (C) ‘Ground’ probes assuming that left side was defined as figural in preceding prime. The top probe shape in C matches the prime curvy edge in A.
Method
Participants
16 people from the local community participated (6 male, 10 female, average age 28.9 years). All reported normal vision and gave informed consent. None of these participants had taken part in Experiments 1 or 2.
Displays and Design
The STEM task prime displays were similar to Experiment 1 displays although instead of a straight dividing edge the unbiased-section now had a unique curvy edge on each trial, with no local edge-assignment cues. The curvy edges were generated by a pseudo-random algorithm that divided each edge into a set of vertical segments of 10 – 30 pixels in length. Each segment was then randomly assigned a horizontal deviation from the vertical midline of the box that could range from 0 to 45 pixels in either direction from the horizontal center of the box. The top and bottom segments were constrained to have a horizontal deviation of 0. The total area of the regions to the left and the right side of the edge was then approximately equated by displacing all of the edge segments (except the top and bottom) an equal amount in the required direction. The resulting edge was then smoothed with a 5-pixel-wide averaging kernel. Overall convexity of the regions was approximately equated by ensuring that the horizontal deviations occurred equally often to the left and right of the horizontal center. The same set of edges was used for each participant. However, these edges were randomly assigned to conditions separately for each participant. The luminance values of the ‘black’ and ‘white’ regions were 8.8 cd/m2 and 53.3 cd/m2.
Probe displays always comprised two shapes (each 3.97° horizontally by 5.3°). The curvy edge of one shape exactly matched the preceding prime display's curvy edge, while the other (foil) was a new unique curvy edge. Matching and foil edges in ‘figure’ probes were each assigned (by closure/surroundedness, see Figure 4B and 4C) to the same side as the biased-section's straight edge in the preceding prime display. ‘Ground’ probe curvy edges were each assigned oppositely. Contrast polarity across the probes' curvy edge was always the same as in the prime display (see Figure 4). Curvy edges were aligned with the screen's horizontal meridian and centered 5.3° vertically above and below fixation. The matching probe within each pair was equiprobably above or below fixation. Top or bottom location for the unbiased-section in the prime display was now counterbalanced between subjects, rather than within as had been done in Experiments 1 and 2. Type-of-probe (figure or ground, with respect to the preceding biased section) was an additional within-subjects factor in this design. All other display counterbalancing was as for Experiment 1. There were 32 repeated measures per condition within each block. Two blocks were presented.
Procedure
Trials began with a 1000 ms fixation cross followed by the 2800 ms prime display, a 500 ms prime-probe ISI, and finally the probe display until response. Participants judged which curvy probe edge (upper or lower) matched the prime display's curvy edge, using a mouse button. Participants were instructed to respond as quickly as possible without sacrificing accuracy. They were given 40 practice trials before the experimental trials began. Trials with >1° deviations from fixation or with pursuit eye movements (>0.5° regular oscillatory structure) during the prime displays were excluded (intersubject mean was 0.85% of trials excluded).
Results and Discussion
Because the pattern of results was the same for STEM error and reaction time measures, the two measures were combined into an inverse efficiency (IE) score (mean-correct RT divided by accuracy-rate) to simplify presentation and analysis (Townsend & Ashby, 1978). See Supplemental Materials for error and RT separately. IE scores (Figure 5) showed main effects of probe-type, F(1,15) = 23.08, p < 0.0001; and edge-grouping, F(1,15) = 11.49, p < 0.004. There were interactions of probe-type and edge-grouping, F(1,15) = 26.37, p < 0.0001; probe-type and region-color-similarity, F(1,15) = 12.74, p < 0.003; and probe x edge-grouping x region-color-similarity, F(1,15) = 4.81, p < 0.04. These interactions show that performance was better for figure than ground probes when edge-grouping and region-color-similarity were present, t(15) = 7.52, p < 0.0001; or in the presence of edge-grouping-only, t(15) = 4.91, p < 0.0001; but that region-color-similarity had no effect when edges were ungrouped in the prime display (no effect for the region-color-similarity only conditions, t(15)= 0.23, p < 0.82, n.s.). There was no difference between figural and ground probes when neither edge grouping nor regional-color-similarity was applied during the prime (t(15) = 0.93, p < 0.36, n.s.). This lack of an effect when edge and region grouping were not present demonstrates once again that simply having a biased-section elsewhere in the image in not sufficient to cause propagation of edge-assignment. Rather, the biased and unbiased section edges must be grouped in order for edge-assignment to propagate from the biased to the unbiased section.
Figure 5.
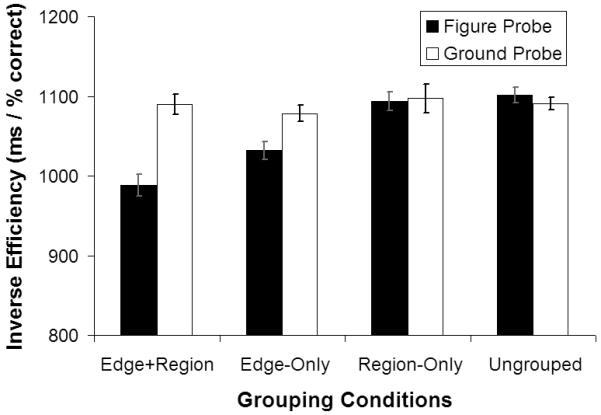
Experiment 3 Results. Average Inverse Efficiency (IE, corresponding to mean correct reaction time divided by proportion correct responses) is plotted as a function of edge-grouping, region-color-similarity, and probe type. Error bars represent 95% within-subjects confidence intervals. Note the enhanced efficiency of performance for figure compared to group probes, when edges were grouped (second pair of bars from left), and even more so when edges were grouped and regional colors were matched (leftmost pair of bars).
Although providing a very different measure, the STEM results of Experiment 3 are perfectly analogous to those for the subjective reports in Experiment 1 (and Experiment 2), in terms of the impact of the edge-grouping and region-color-similarity factors. These factors now affected performance in recognizing different types of probe shapes, rather than merely affecting subjective report as before. As in Experiments 1 and 2, figural edge-assignment propagated from the biased-section's edge to the (now curvy) edge within the unbiased-section, but only when those two sections were grouped by edge-grouping. Because of this, the curvy edge in the unbiased-section was then more likely to be seen as assigned in the corresponding direction. Hence participants were more efficient (faster and more accurate, see Supplementary Materials) at recognizing that edge in the probe phase when it was figurally assigned in the same direction for the probe as for the unbiased section in the prime. Also as in Experiments 1 and 2, region-color-similarity had a small effect but only when the edges were also grouped. These results provide a further replication of our effects that now demonstrate the consequences for objective performance.
For completeness, at the request of a reviewer we again tested further for any impact from the fully counterbalanced factor of which side of the unbiased section initially expanded (versus contracted) due to the initial direction of motion of the dividing wavy edge, prior to the oscillatory reversal in direction. To assess this we had to re-code the data such that if, for instance, the left region of the prime display's unbiased-section initially expanded (i.e. the wavy dividing edge moved rightward first), then a matching subsequent static probe edge that was assigned leftward would be re-coded as congruent probe. For this example, a matching edge that was assigned rightward instead in the static probe would be re-coded as a potential incongruent probe. The opposite would apply if the right region of the unbiased-section had expanded first during initial moving exposure. We found no impact of the initial expansion/contraction factor on STEM performance (mean RT for congruent probes (993.6 ms) and incongruent probes (990.6 ms) did not differ, t(15) = 0.404, p = 0.602, when the data were coded this way. Error rates also showed no impact (8.0% versus 7.2%, t(15) = −1.16, p = 0.264. These results indicate that, unlike our more critical factors of interest, the counterbalanced initial expansion/contraction cue does not affect performance in the STEM task for the current context. This differs from some of the small but reliable impacts of this counterbalanced factor on subjective judgments in Experiments 1 and 2. This might reflect some potential differences between phenomenal versus objective measures; and/or perhaps greater sensitivity of the phenomenal measure to a subtle influence originating at initial display onset. The probes for the STEM task appeared 3300 ms after initial onset of the prime displays, and subsequent to oscillatory motion in the primes which meant that either side of the dividing edges had both undergone expansion and contraction by then.
Fortunately, these consideration of the fully counterbalanced expansion/contraction cue are all orthogonal to our main factors of interest, which showed in Experiment 3 that the key results of Experiments 1, in terms of grouping-constrained propagation of figural assignment, do extend also to an objective consequence of figural assignment, namely for the STEM task.
Experiment 4: Control for salient dots potentially attracting attention when using the STEM task as the measure for the consequences of figural assignment
Analogously to Experiment 2, we next manipulated the location of the moving dots (appearing on one or other or both sides of the dividing edge in the biased section of the prime). This was in order to address the potential concern of moving dots potentially attracting attention to one side, as described above in the introduction for Experiment 2. We now verified the results using the objective STEM task, as per Experiment 3, rather than the subjective measure as used in Experiment 2.
Method
Participants
16 people from the local community (8 male, 8 female, average age 23.4 years) participated. All reported normal vision and gave informed consent. None had taken part in Experiments 1, 2 or 3.
Displays and Design
The prime displays were the same as in Experiment 2. The parameters of the probe displays were the same as in Experiment 3. Counterbalancing of display parameters was as for Experiment 2. Within each block there were 32 repeated measures per condition in the 2 × 2 × 2 × 3 within-subjects design. The factors were edge-grouping (2), region-color-similarity (2), probe-type (2), and salient-dots-location (3). Each participant completed 2 blocks. There were 4 breaks within each block.
Procedure
The procedure was the same as in Experiment 3, except for the further manipulation of salient-dots-location. Trials with >1° deviations from fixation or with pursuit eye movements (>0.5° regular oscillatory structure) during the prime displays were excluded (intersubject mean of 1.3% trials excluded).
Results and Discussion
In the overall 2 × 2 × 2 × 3 ANOVA, there was a significant main effect of probe-type, F(1,15) = 31.67, p < 0.0001, indicating that some propagation of figural assignment did occur (because the difference between probe types reflects propagation of figural assignment from the biased-section to the unbiased-section); see Figure 6. There were also significant effects of edge-grouping, F(1,15) = 24.86, p < 0.0001; region-color-similarity, F(1,15) = 4.52, p < 0.049; and their interaction, F(1,15) = 7.72, p < 0.014, indicating that these factors had a combined impact on performance. The probe effect interacted with both edge-grouping, F(1,15) = 133.46, p < 0.0001, and region-color-similarity, F(1,15) = 10.35, p < 0.006, confirming that the difference between figure and ground probes (the critical measure of edge-assignment propagation here) depended on edge-grouping and was also affected by the presence of region-color-similarity. As in all of our previous experiments, region-color-similarity only had an effect when edge-grouping was also present, as reflected in the 3-way interaction of probe-type, edge-grouping, and region-color-similarity, F(1,15) = 12.15, p < 0.003. None of the factors interacted with salience (all p > 0.189) demonstrating that the results were equivalent for the three dot-salience conditions. There was no main effect of salience either, F(2,30) = 0.37, p < 0.692.
Figure 6.

Experiment 4 Results. (A) Results for the salience-balanced condition (i.e. moving dots on both sides of biased-section in prime). (B) Results for the ground-salient condition (i.e. dots only on ground side for biased-section in prime). (C) Results for the figure-salient condition (i.e. dots only on figural side for biased-section in prime). Error bars represent 95% within-subjects confidence intervals. Note the similar outcome for all three graphs, all of which replicate the pattern from Experiment 3 shown in Figure 5.
Within each salience condition (assessed by separate 2 × 2 × 2 ANOVAs on the three distinct dot arrangements), there was always an impact of probe-type (for salience-balanced, F(1,15) = 26.85, p < 0.0001; ground-salient, F(1,15) = 18.39, p < 0.001; figure-salient, F(1,15) = 23.18, p < 0.0001) and of edge-grouping (salience-balanced, F(1,15) = 17.77, p < 0.001; ground-salient, F(1,15) = 22.37, p < 0.0001; figure-salient, F(1,15) = 10.75, p < 0.005). There was also a 3-way interaction for all dot conditions (i.e. between probe-type, edge-grouping and region-color-similarity: for salience-balanced, F(1,15) = 7.03, p < 0.018; ground-salient, F(1,15) = 5.76, p < 0.03; figure-salient, F(1,15) = 8.16, p < 0.01). In all cases, this interaction arose because region-color-similarity mattered for the impact of probe-type only when edges were grouped (as in all of our previous experiments also). As in Experiment 3, at the request of a reviewer we tested for effects of the counterbalanced expansion/contraction cue. As in Experiment 3, we found no significant impact on the STEM task: mean RTs of 1070.6 ms versus 1067.2 ms for congruent and incongruent probes respectively, t(15) = 0.148, p = 0.881, n.s.; 7.5% and 7.3% error-rates respectively, t(15) = 0.718, p = 0.483, n.s. These results are consistent with those found in Experiment 3.
The three replications (see Figure 6) of the STEM task here all converge perfectly with the previous STEM task results from Experiment 3 (compare Figures 5 and 6), as well as with the subjective report results of Experiments 1 and 2., with respect to our factors of main interest. Experiment 4 also provides further evidence (in addition to Experiment 2) that none of our critical effects depend on dot salience favoring one or other side of the biased section.
Experiment 5: Extension to static displays and the role of the occluder
We next tested whether our new principle, that grouping constrains propagation of figural edge-assignment between edges, would generalize to different displays and grouping factors. We now manipulated edge-grouping in a completely different way than via common fate, and also used a very different local edge-assignment cue for the biased-section, now within static displays. Experiment 5 returned to phenomenal measures (as in Experiments 1 and 2). We manipulated T-junctions as the local cue for edge-assignment within the biased-section (see Figure 7A-D, noting the different vertical extents of the black and white regions at the bottom of each display, and the resulting T-junction, as arrowed in Figure 7A). T-junctions have previously been recognized as an important cue to relative depth at edges (Clowes, 1971; Guzman, 1969; Huffman, 1971) and have been used to manipulate edge-assignment in some neurophysiological work (e.g. Zhou et al., 2000). Furthermore, several computational models of edge-assignment (as mentioned in the introduction) that consider propagation of such assignment along complete or partially complete contours actually rely on T-junction depth cues (e.g. Craft et al., 2007; Heitger et al., 1998; Peterhans & Heitger, 2001; Thielscher & Neumann, 2008).
Figure 7.

Experiment 5, example displays and results. (A) Horizontal segment serving as an occluder that appears pictorially ‘in-front’, with edge-grouping by edge-contrast similarity between biased (bottom) and unbiased (top) sections. Local t-junction cues (see arrow, not present in actual display) assign the dividing edge to the right within the biased-section at bottom. (B) Horizontal segment now appears pictorially ‘behind’, so cannot serve as an occluder, despite edge-contrast similarity between biased (bottom) and unbiased (top) sections. Note that the new pictorial cues to the horizontal segment being ‘behind’ actually bring the unbiased and biased-sections slightly closer together, but nevertheless less propagation of edge-assignment is found between them. (C) Horizontal segment pictorially ‘in front’ (as for A), but now with edges in biased and unbiased-sections having opposite contrast polarity. (D) Horizontal segment pictorially ‘behind’ (see also B), now with edges in biased and unbiased-sections having opposite contrast polarity. (E) Results of Experiment 5 for the conditions with the horizontal segment in front. (F) Results of Experiment 5 for the conditions with the horizontal segment behind, so that it unlikely to look like an occluder, which eliminates the impact of edge-grouping and its interaction with region-color-similarity.
In Experiment 5, we also took a closer look at any role for contrast-polarity across the critical dividing edges as a grouping factor (previously we had varied contrast-polarity only when region-color was dissimilar rather than varying this in a fully orthogonal manner, see Experiment 1). Contrast polarity now served as a strictly orthogonal edge-grouping cue (see below). Manipulating just contrast polarity allowed us to hold collinearity constant now between the biased and unbiased dividing edges, while also eliminating common fate, unlike our four preceding experiments. We now used static rather than moving displays, seeking to generalize our findings.
Finally, in order to understand the role of the ‘occluder’ that had previously intervened between biased and unbiased dividing edges, and to study how this might affect propagation of edge-assignment, we now varied the perceived depth of the potentially ‘occluding’ horizontal segment that had separated biased and unbiased-sections hitherto (see Figure 1A). When this segment appeared to be an occluder, apparently in front of the biased and unbiased-sections (see Figures 7A & 7C), the dividing edges in those two sections could actually be continuous in the external world, being interrupted in the image only by partial occlusion. But when the same horizontal segment clearly appeared pictorially as ‘behind’ (Figures 7B & 7D) the biased and unbiased sections, as indicated by overlaps that break the T-junctions where the horizontal section meets those sections, this interpretation becomes invalid, with the upper and lower sections now less ‘grouped’ in that sense. Hence figural edge-assignment may then no longer propagate between the biased and unbiased sections as effectively in cases like Figures 7B and 7D, as for apparent-occlusion cases such as Figures 7A and 7C.
Methods
Participants
12 people from the local community participated (6 male, 6 female, average age 25.3 years). All reported normal vision and gave informed consent. None had taken part in the previous experiments.
Displays and Design
The displays were conceptually similar to Experiment 1, but now without any motion or dots. Instead, assignment of the biased-section's dividing edge was determined locally by T-junctions (see Figure 7A-D) at the far end of the biased section that was furthest away from the unbiased section (and the horizontal potentially occluding segment; see bottom of each panel in Fig 7A-D). Region-color-similarity was analogous to Experiment 1, in the sense that either the biased- and unbiased sections were both composed of black and white, or else one section was colored while the other was black and white, see below. Similarity of edge-contrast-polarity was now for the first time a fully orthogonal factor, regardless of whether region-color was similar or dissimilar between biased and unbiased-sections. A final factor was whether the segment intervening between these sections appeared to be in front (e.g., Figures 7A & 7C) or behind both the biased and unbiased-sections (Figures 7B & 7D) as manipulated via pictorial overlap cues.
In the condition with the intervening segment now pictorially “behind”, we extended the ends of both the biased and unbiased sections into the horizontal intervening segment. This created t-junctions along the outer vertical edges of the black and white regions (i.e. the biased and unbiased section) which caused the horizontal segment to appear behind them (see Figures 7B and 7D). Within the unbiased-section both the regions protruded 0.5° into the horizontal segment (upper sections in Figures 7B and 7D). However, in the biased-section, the ground region extended 0.5° into the horizontal segment, whereas the figural region extended 0.8° into the horizontal segment (lower section in Figures 7B and 7D). This was done for two reasons. First, it maintained the local edge-assignment cue within the biased-section that was also present at the other end of the biased-section. Second, the extended figural region was designed to eliminate the otherwise possible perception of the biased and unbiased section regions as connecting via an apparent ‘slit’ in the grey section. Only a slit that was cut just so and then perfectly aligned with the edge between the black and white regions would accord with this perception. This perceptual interpretation should thus be highly unlikely, given the visual system's known bias against highly “accidental” interpretations of a scene (Rock, 1983; Lowe, 1985; Albert & Hoffman, 1995). Accordingly, we expected that the new stimulus configuration would lead to the horizontal segment being perceived as behind the abutting biased and unbiased sections, hence our description of this as the ‘behind’ condition, even though only pictorial depth cues were utilized.
Procedure
This was as for Experiment 1 except displays were shown for only 500 ms (intended to minimize any perceptual reversals), unlike the moving displays in Experiment 1 which required longer display times. Trials with >1° deviations from fixation or with pursuit eye movements (>0.5° regular oscillatory structure) during the displays were excluded (intersubject mean of 0.5% trials discarded)
Results and Discussion
For the biased-section, t-junction cues determined phenomenal figural edge-assignment as expected in 90.9% of trials (significantly more than 50% chance, t(11) = 28.04, p < 0.001). This did not differ between the conditions when the horizontal segment was in front (90.0%) versus when it was behind (91.8%), t(11) = 1.63, p < 0.134, confirming that the local bias we had applied to the biased section via pictorial cues was equally effective in either case. For the unbiased-section, the percentage of context-consistent judgments (see Figures 7E and 7F) depended on our new grouping cue of whether the horizontal segment appeared as an occluder or not, F(1,11) = 14.05, p < 0.003; plus also depended upon on region-color-similarity, F(1,11) = 6.25, p < 0.029, and on edge-contrast-polarity matching or otherwise, F(1,11) = 19.33, p < 0.001. Occlusion interacted with the region-color-similarity factor, F(1,11) = 5.71, p < 0.036, and the edge-contrast-polarity factor, F(1,11) = 19.61, p < 0.001, because in the absence of apparent occlusion the other factors then had no impact (i.e. all segment-behind conditions had chance-levels of context-consistent responses when reporting the unbiased-section's figural edge-assignment; see Figure 7F). By contrast, when the horizontal intervening segment appeared to be an occluder, figural assignment of the dividing edge propagated from the biased to the unbiased-section, and did so most when both region-color was similar and contrast polarity of the edges matched; see Figure 7E. These results are logically analogous to those in Experiment 1 (and all of our other experiments). But they now show that our new principle, whereby grouping constrains propagation of figural edge-assignment, extends to very different displays and to different grouping cues.
In particular, Experiment 5 shows that our findings on propagation of figural edge-assignment from the biased to the unbiased-section can generalize to static displays, in which T-junctions serve as the local figural cues (at the extremity of the biased section). The results further show that this propagation is constrained by occlusion relations; and by region-color-similarity and edge-contrast-polarity, provided that the intervening segment appears to be an occluder, consistent with biased and unbiased-edges potentially reflecting a common edge that is partially occluded.
General Discussion
Our results show that figural assignment of a dividing edge can be affected by contextual information well beyond the two local regions immediately on either side of that edge, providing the basis for a new class of contextual figure-ground cues. But importantly, not all contextual edges have equal influence. Edge-assignment along one dividing edge (here in the locally-biased-section) only propagated to the other dividing edge (in the unbiased-section) when the two edges grouped with one another. This gating of edge-assignment propagation by grouping was shown for common fate motion and collinearity (Experiments 1-4), and separately for grouping by edge-contrast-polarity and occlusion (Experiment 5). When the occlusion relation was reversed pictorially, rendering the separate edges perceptually discontinuous (Figs 7B and 7D), propagation of edge-assignment from the biased- to the unbiased-section was eliminated. Region-color-similarity also had some facilitatory influence on propagation of edge-assignment, but could be vetoed by edge-ungrouping. In other words, region-color-similarity only mattered when the dividing edges were grouped, as we confirmed in all five experiments.
It may be worth noting that here we always manipulated region-color-similarity in the presence of relatively powerful edge-grouping factors (such as contrast polarity and common fate). In the present studies we did not explore the influence of region-color-similarity on its own. Moreover, in all the cases where edge-grouping was absent in our study, one could argue that in fact there was a strong edge-ungrouping cue (e.g. different speeds and out-of-phase motion for the common fate manipulations; or opposite contrast polarity in our static experiment). In documenting for the first time the contextual propagation of figure-ground assignment between disconnected elements of a display under the constraint of grouping, our study raises many new questions for future research. These include whether region-color-similarity can be effective on its own, when edge-grouping is kept neutral; what happens if color-similarity for potential grounds is manipulated orthogonally to that for potential figures; whether absolute or relative color or contrast values are more critical; and how figural assignment of the unbiased section arises in time, relative to that for the biased section. Future variations on the approach introduced here could address these issues, together with many further interesting questions that arise from our initial demonstrations.
We propose that our existing demonstrations reflect a mechanistic link between perceptual grouping mechanisms and figural edge-assignment mechanisms. From this perspective, propagation of figural edge-assignment proceeds when the visual system judges that the edges in question are likely to be related and probably reflect a common external source, as when the edges are grouped together. However, as we briefly discussed when introducing Experiments 2 and 4, it might be possible to suggest that attention could play some mediating role in principle. For instance, we considered (but rejected) the possibility that attention-capture by salient dots on the figural side of the biased edge might advantage that side, with this advantage then potentially propagating via object-based or grouping-constrained attention (e.g., Matsukura & Vecera, 2006; Egly et al., 1994; Behrmann et al., 2000) to the corresponding side of the unbiased edge. In Experiments 2 and 4 we addressed this issue for the specific proposal that attention might be captured by salient moving dots, by manipulating which side(s) of the biased dividing edge contained such dots. We found that the critical results remained even when moving dots appeared only on the ground side of the biased dividing edge, thus arguing against any simple account for our results in terms of attention capture by moving dots per se.
It might nevertheless be possible still to argue that despite this specific result, figural regions tend intrinsically to attract attention following edge-assignment to that side (e.g., see Nelson & Palmer, 2007 for evidence that this happens for familiar shapes). On such an argument, the figural side of the biased section might thereby become the attended side also, making it potentially ambiguous whether figural assignment per se, or attention (or both) is what propagates to the corresponding side of the unbiased section. We think that our dot-salience manipulation provides some initial evidence against such a proposal, or at least evidence which makes such a proposal less parsimonious. On our account figural assignment can propagate, under the constraint of grouping, from a biased dividing edge to an unbiased dividing edge. It seems to us that any alternative account in terms of an intervening role for attention probably has to postulate a similar causal chain, but with the added premise that redirection of attention might intervene between initial figural assignment of the biased dividing edge, and propagation of the resulting advantage for one side to the corresponding side of the unbiased section, again under the constraint of grouping. Occam's razor would thus suggest that any such intervening role for attention has to be proven rather than merely assumed. Moreover, it is no longer uncontested that figural regions and attended regions are inevitably associated (Huang & Pashler, 2009). For these reasons, we prefer for now to retain our hypothesis that figural assignment for one edge can propagate to related but separate edges, under the constrain of grouping, without necessarily advocating any essential or additional role for attention. Nevertheless, now that we have established the new principle of figural assignment propagating under the constraint of perceptual grouping, it may be interesting to study whether this applies equally for attended and unattended stimuli, in future variants of the paradigms we have introduced.
Our manipulation of the occluder in Experiment 5 provides a further demonstration, now in relation to pictorial cues for 3D depth, that when perceptual evidence suggests that the edges and regions in the biased and unbiased-sections of the display are unlikely to be related, then figural edge-assignment will not propagate between them. One way of understanding this occluder manipulation is as another example of a grouping manipulation. When the occluder is “in front” it suggests that the edges and regions should be completed behind it by virtue of the aligned t-junctions on either side of it, which provide a strong perceptual cue for a partially occluded relation. However, when the occluder is pictorially “behind”, the evidence that these junctions are actually corners suggests that the two sections should not group with one another and so that the regions should not be interpolated.
Our results provide a new rationale for investigating how neural populations involved in visual contour grouping (e.g., Polat et al., 1998) may relate to those involved in figural edge-assignment (e.g., Zhou et al., 2000). Such interactions have previously been suggested by computational modeling work (Craft et al., 2007; Heitger et al., 1998; Peterhans & Heitger, 2001; Thielscher & Neumann, 2008) in which edge-assignment caused by t-junctions is considered to propagate along contours. One possible neural correlate for our new results on human perception might be that the neural populations involved in contour-grouping (e.g., Polat et al., 1998) are also involved in figural edge-assignment (e.g., von der Heydt et al., 2000; Zhou et al., 2000). Alternatively, two distinct populations (one responsible for grouping, another for edge-assignment) might interact. Neural measures in paradigms relating to those that we have introduced here may be required to resolve this.
Our results cast some doubt on Palmer & Rock's (1994b) proposal that figure-ground assignment should always precede grouping. Their claim was largely based on arguments from first principles, within their overarching perspective on perceptual organization. They initially proposed that grouping may only occur after figure-ground organization, arguing that figure-ground organization provides the elements that can then be subject to grouping processes. Here we have clearly established the converse influence for human perception, namely that grouping can affect figure-ground organization. Grouping clearly determined figure-ground assignment for the unbiased section of our displays. But one might still ask whether figural assignment within the biased section preceded, followed, or overlapped with the grouping between biased and unbiased edges and regions. Some of the neural measures as suggested above might shed further light on the exact time course of the underlying processes.
Despite the new issues arising for future research, the present results clearly establish a new contextual influence on figure-ground assignment, and show that this contextual influence is gated by perceptual grouping. By integrating information about edge-assignment only between grouped contextual edges, figure-ground processes may be able to maintain consistent figure-ground organization along edges where inconsistent local cues or partial occlusion in the image might otherwise disrupt this continuity.
Supplementary Material
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Acknowledgments
Joseph L. Brooks and Jon Driver, UCL Institute of Cognitive Neuroscience, 17 Queen Square, London, WC1N 3AR, United Kingdom. JLB was supported by a Royal Society Post-Doctoral Fellowship. JD was supported by the Wellcome Trust, MRC, BBSRC, and a Royal Society Anniversary Research Professorship. We thank Sergei Gepshtein for early discussions leading to this work.
References
- Albert MK, Hoffman DD. Genericity in spatial vision. In: Luce RD, D'Zmura M, Hoffman DD, Iverson GJ, Romney AK, editors. Geometric representations of perceptual phenomena: Papers in honor of Tarow Indow on his 70th birthday. Erlbaum; Mahwah, N.J.: 1995. [Google Scholar]
- Appelbaum LG, Wade AR, Vildavski VY, Pettet MW, Norcia AM. Cue-invariant networks for figure and background processing in human visual cortex. Journal of Neuroscience. 2006;26:11695–11708. doi: 10.1523/JNEUROSCI.2741-06.2006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Barenholtz E, Tarr MJ. Figure-ground assignment to a translating contour: a preference for advancing vs. receding motion. Journal of Vision. 2009;9:27–29. doi: 10.1167/9.5.27. [DOI] [PubMed] [Google Scholar]
- Baylis GC, Cale EM. The figure has a shape, but the ground does not: Evidence from a priming paradigm. Journal of Experimental Psychology: Human Perception and Performance. 2001;27:633–643. doi: 10.1037//0096-1523.27.3.633. [DOI] [PubMed] [Google Scholar]
- Baylis GC, Driver J. One-sided edge assignment in vision: I. Figure-ground segmentation and attention to objects. Current Directions in Psychological Science. 1995;4:140–146. [Google Scholar]
- Baylis GC, Driver J. Shape-coding in IT cells generalizes over contrast and mirror reversal, but not figure-ground reversal. Nature Neuroscience. 2001;4:937–942. doi: 10.1038/nn0901-937. [DOI] [PubMed] [Google Scholar]
- Behrmann M, Zemel RS, Mozer MC. Occlusion, symmetry, and object-based attention: reply to Saiki (2000) Journal of Experimental Psychology: Human Perception and Performance. 2000;26:1497–1505. doi: 10.1037//0096-1523.26.4.1497. [DOI] [PubMed] [Google Scholar]
- Clowes MB. On seeing things. Artificial Intelligence. 1971;2:79–116. [Google Scholar]
- Craft E, Schutze H, Niebur E, von der HR. A neural model of figure-ground organization. Journal of Neurophysiology. 2007;97:4310–4326. doi: 10.1152/jn.00203.2007. [DOI] [PubMed] [Google Scholar]
- Driver J, Baylis GC. Edge-assignment and figure-ground segmentation in short-term visual matching. Cognitive Psychology. 1996;31:248–306. doi: 10.1006/cogp.1996.0018. [DOI] [PubMed] [Google Scholar]
- Egly R, Driver J, Rafal RD. Shifting visual attention between objects and locations: evidence from normal and parietal lesion subjects. Journal of Experimental Psychology: General. 1994;123:161–177. doi: 10.1037//0096-3445.123.2.161. [DOI] [PubMed] [Google Scholar]
- Franconeri SL, Simons DJ. The dynamic events that capture visual attention: A reply to Abrams and Christ (2005) Perception & Psychophysics. 2005;67:962–966. doi: 10.3758/bf03193623. [DOI] [PubMed] [Google Scholar]
- Guzman A. Decomposition of a visual scene into three-dimensional bodies. In: Griselli A, editor. Automatic interpretation and classification of images. Academic Press; New York: 1969. pp. 243–276. [Google Scholar]
- Heitger F, von der Heydt R, Peterhans E, Rosenthaler L, Kubler O. Simulation of neural contour mechanisms: representing anomalous contours. Image and Vision Computing. 1998;16:407–421. [Google Scholar]
- Hillstrom AP, Yantis S. Visual motion and attentional capture. Perception & Psychophysics. 1994;55:399–411. doi: 10.3758/bf03205298. [DOI] [PubMed] [Google Scholar]
- Huang L, Pashler H. Reversing the attention effect in figure-ground perception. Psychological Science. 2009;20:1199–1201. doi: 10.1111/j.1467-9280.2009.02424.x. [DOI] [PubMed] [Google Scholar]
- Huffman DA. In: Impossible objects as nonsense sentences. Meltzerm M, Michie D, editors. Edinburgh University Press; Edinburgh, Scotlland: 1971. [Google Scholar]
- Hulleman J, Humphreys GW. A new cue to figure-ground coding: Top-bottom polarity. Vision Research. 2004;44:2779–2791. doi: 10.1016/j.visres.2004.06.012. [DOI] [PubMed] [Google Scholar]
- Kanizsa G, Gerbino W. Convexity and symmetry in figure-ground organization. In: Henle M, editor. Vision and Artifact. Springer; New York: 1976. [Google Scholar]
- Kourtzi Z, Kanwisher N. Representation of perceived object shape by the human lateral occipital complex. Science. 2001;293:1506–1509. doi: 10.1126/science.1061133. [DOI] [PubMed] [Google Scholar]
- Lowe DG. Perceptual organization and visual recognition. Kluwer Academic Publishers; Boston, MA: 1985. [Google Scholar]
- Matsukura M, Vecera SP. The return of object-based attention: selection of multiple-region objects. Perception & Psychophysics. 2006;68:1163–1175. doi: 10.3758/bf03193718. [DOI] [PubMed] [Google Scholar]
- Metzger . Gesetze des Sehens. Waldemar Kramer; Frankfurt-am-Main: 1953. [Google Scholar]
- Nelson R, Palmer SE. Familiar Shapes Attract Attention in Figure-Ground Displays. Perception and Psychophysics. 2007;69:382–392. doi: 10.3758/bf03193759. [DOI] [PubMed] [Google Scholar]
- Palmer SE. Vision Science: Photons to Phenomenology. MIT Press; Cambridge, MA: 1999. [Google Scholar]
- Palmer SE, Brooks JL. Edge-region grouping in figure-ground organization and depth perception. Journal of Experimental Psychology: Human Perception and Performance. 2008;34:1353–1371. doi: 10.1037/a0012729. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Palmer SE, Rock I. On the nature and order of organizational processing: A reply to Peterson. Psychonomic Bulletin & Review. 1994a;1:515–519. doi: 10.3758/BF03210957. [DOI] [PubMed] [Google Scholar]
- Palmer SE, Rock I. Rethinking perceptual organization: The role of uniform connectedness. Psychonomic Bulletin & Review. 1994b;1:29–55. doi: 10.3758/BF03200760. [DOI] [PubMed] [Google Scholar]
- Peterhans E, Heitger F. Simulation of neuronal responses defining depth order and contrast polarity at illusory contours in monkey area V2. Journal of Computational Neuroscience. 2001;10:195–211. doi: 10.1023/a:1011273115282. [DOI] [PubMed] [Google Scholar]
- Peterson MA, Salvagio E. Inhibitory competition in figure-ground perception: context and convexity. Journal of Vision. 2008;8:4–13. doi: 10.1167/8.16.4. [DOI] [PubMed] [Google Scholar]
- Peterson MA. Object recognition processes can and do operate before figure-ground organization. Current Directions in Psychological Science. 1994;3:105–111. [Google Scholar]
- Peterson MA, Gibson BS. Must figure-ground organization precede object recognition? An assumption in peril. Psychological Science. 1994a;5:253–259. [Google Scholar]
- Peterson MA, Gibson BS. Object recognition contributions to figure ground organization: Operations on outlines and subjective contours. Perception & Psychophysics. 1994b;56:551–564. doi: 10.3758/bf03206951. [DOI] [PubMed] [Google Scholar]
- Polat U, Mizobe K, Pettet MW, Kasamatsu T, Norcia AM. Collinear stimuli regulate visual responses depending on cell's contrast threshold. Nature. 1998;391:580–584. doi: 10.1038/35372. [DOI] [PubMed] [Google Scholar]
- Rock I. The logic of perception. MIT Press; Cambridge, MA: 1983. [Google Scholar]
- Rubin E. Visuell Wahrgenommene Figuren. Glydendalske; Copenhagen: 1921. [Google Scholar]
- Stevens KA, Brookes A. The concave cusp as a determiner of figure-ground. Perception. 1988;17:35–42. doi: 10.1068/p170035. [DOI] [PubMed] [Google Scholar]
- Thielscher A, Neumann H. Globally consistent depth sorting of overlapping 2D surfaces in a model using local recurrent interactions. Biological Cybernetics. 2008;98:305–337. doi: 10.1007/s00422-008-0211-7. [DOI] [PubMed] [Google Scholar]
- Townsend JT, Ashby FG. Methods of modeling capacity in simple processing systems. In: Castellan NJ, Restle F, editors. Cognitive Theory. Earlbaum; Hillsdale, NJ: 1978. pp. 199–239. [Google Scholar]
- Vecera SP, Flevaris AV, Filapek JC. Exogenous Spatial Attention Influences Figure-Ground Assignment. Psychological Science. 2004;15:20–26. doi: 10.1111/j.0963-7214.2004.01501004.x. [DOI] [PubMed] [Google Scholar]
- Vecera SP, Vogel EK, Woodman GF. Lower region: A new cue for figure-ground assignment. Journal of Experimental Psychology: General. 2002;131:194–205. doi: 10.1037//0096-3445.131.2.194. [DOI] [PubMed] [Google Scholar]
- von der Heydt R, Zhou H, Friedman HS. Representation of stereoscopic edges in monkey visual cortex. Vision Research. 2000;40:1955–1967. doi: 10.1016/s0042-6989(00)00044-4. [DOI] [PubMed] [Google Scholar]
- Wagner G, Boynton RM. Comparison of four methods of heterochromatic photometry. Journal of the Optical Society of America. 1972;62:1508–1515. doi: 10.1364/josa.62.001508. [DOI] [PubMed] [Google Scholar]
- Yonas A, Craton LG, Thompson WB. Relative motion: Kinetic information for the order of depth at an edge. Perception & Psychophysics. 1987;41:53–59. doi: 10.3758/bf03208213. [DOI] [PubMed] [Google Scholar]
- Zhou H, Friedman HS, von der Heydt R. Coding of border ownership in monkey visual cortex. Journal of Neuroscience. 2000;20:6594–6611. doi: 10.1523/JNEUROSCI.20-17-06594.2000. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.