

Summary
A central problem in posttraumatic stress disorder (PTSD) is the inability to suppress fear under safe conditions. We have previously shown that PTSD patients cannot inhibit conditioned fear. Another relevant finding in PTSD is the hypersensitivity of the hypothalamic-pituitary adrenal (HPA) axis feedback. Given their common neurobiological pathways, alterations in HPA function in PTSD may be associated with impaired fear inhibition. The present study examined the relationship between HPA axis function and fear-potentiated startle and inhibition of conditioned fear in trauma-exposed individuals. We used a conditional discrimination procedure (AX+/BX−), in which one set of shapes (AX+) was paired with aversive airblasts to the throat (danger signal), and the same X shape with a different shape (BX−) were presented without airblasts (safety signal). The paradigm also included a transfer of fear inhibition test (AB). In addition to fear-potentiated startle, blood was drawn for neuroendocrine analysis and the dexamethasone suppression test (DEX) was performed; cortisol and ACTH were assessed at baseline and post-DEX. Ninety highly traumatized individuals recruited from Grady Hospital in Atlanta, GA participated in the study. The sample was divided into those who met DSM-IV criteria for PTSD (n=29) and Non-PTSD controls (n=61) using the PTSD symptom scale (PSS). Both groups showed significant reduction in cortisol and ACTH levels after DEX. Subjects with PTSD had higher fear-potentiated startle to the safety signal, BX− (F(1,88)=4.44, p<0.05) and fear inhibition trials, AB (F(1,88)=5.20, p<0.05), both indicative of less fear inhibition in the presence of B, compared to control subjects. In addition, fear-potentiated startle to AX+, BX−, and AB was positively correlated with baseline and post-DEX ACTH in PTSD subjects. These results suggest that impaired fear inhibition and associated alterations in HPA feedback may reflect amygdala hyperactivity in subjects with PTSD.
Keywords: PTSD, Trauma, Physiology, Startle Response, HPA, Cortisol
Introduction
Posttraumatic stress disorder (PTSD) can develop in individuals who experience severely traumatic events. While both genes and environment interact to increase an individual’s risk of developing PTSD, it is unclear how the underlying neurobiology is shaped by these factors to result in the observed dysregulations. Animal models of early life stress have found long-term effects on the hypothalamic-pituitary-adrenal (HPA) axis (Plotsky and Meaney, 1993). Similar persistent changes in HPA function as a result of early life stress have been found in human studies. Adult women who were abused in childhood show increased adrenocorticotropin hormone (ACTH) and cortisol reactivity to a psychosocial stressor (Heim et al., 2000). Exaggerated suppression of HPA activity following dexamethasone challenge has been a consistent finding in PTSD (Yehuda et al., 2002; Yehuda et al., 2004a; Yehuda et al., 2004b) and in traumatized individuals regardless of PTSD status (de Kloet et al., 2007). On the other hand, concentrations of corticotropin releasing hormone (CRH) in the cerebrospinal fluid are higher in PTSD patients (Bremner et al., 1997; Baker et al., 1999). Furthermore, elevated CRH levels are associated with increased fear response (Kalin and Takahashi, 1990) and anxiety (Sutton et al., 1982), including the startle response (Liang et al., 1992; Lee and Davis, 1997; Keen-Rhinehart et al., 2008) and enhanced fear conditioning (Swerdlow et al., 1989; Roozendaal et al., 2002).
Fear conditioning is based on a simple Pavlovian conditioning model in which a neutral conditioned stimulus (CS, for example, a light) is paired with an aversive unconditioned stimulus (US, for example, electric shock). After a number of pairings, the association is formed so that the CS alone elicits the conditioned response (CR, for example, a fear response). Two physiological responses have been used as behavioral outcome measures for fear conditioning in humans: acoustic startle response and skin conductance responses. The acoustic startle response is characterized by an integrative, reflex contraction of the skeletal musculature in response to a sudden intense environmental stimulus (Landis and Hunt, 1939; Yeomans and Frankland, 1995; Yeomans et al., 2002). It is mediated by a simple subcortical three-neuron circuit (Davis et al., 1982; Lee et al., 1996; Koch, 1999), but is modulated by limbic brain structures such as the amygdala. Fear-potentiated startle is the relative increase in the startle response elicited in the presence of a conditioned stimulus that was previously paired with an aversive stimulus (Davis, 1992).
Fear acquisition refers to learning that something is dangerous, whereas fear inhibition is a mechanism by which an individual learns that something which previously elicited fear is no longer dangerous, i.e., that it is safe. The two major laboratory models used for behavioral testing of fear inhibition, extinction and conditioned inhibition, focus on active suppression of fear responses through learned safety signals. A recent meta-analysis of 15 studies using fear conditioning found that patients with anxiety disorders showed greater levels of fear responses compared to healthy controls (Lissek et al., 2005). These data suggest that the fear response is dysregulated in PTSD, which has led researchers to use fear conditioning models to examine some of the core PTSD symptoms. One study (Grillon and Morgan, 1999) used a fear-potentiated startle paradigm with veterans diagnosed with PTSD and found equivalent levels of fear potentiation to the danger signal in the PTSD and control groups. However, the PTSD subjects also potentiated to the safety cue, whereas the controls did not.
Our recent data also show that increased fear responses to safety cues are related to the severity of current PTSD symptoms (Jovanovic et al., 2009). A recent study of patients with panic disorder found that these patients also had increased fear-potentiated startle responses to the safety cue; this finding may have been related to the patients’ increased expectancy of the US during the safety cue (Lissek et al., 2009). In this study the impaired discrimination between danger and safety appeared to involve both cognitive as well as physiological deficits (Lissek et al., 2009); in our study of veterans with PTSD the impairment was only seen on the startle measure (i.e., they reported that they did not expect to receive an airblast US during the CS− trial). A study by Orr and colleagues that looked at fear conditioning in PTSD subjects using skin conductance found that PTSD subjects discriminated between the danger and safety cues better than controls (Orr et al., 2000). Similarly enhanced conditionability in PTSD patients was found when trauma-related cues were used as the US in fear conditioning; in this study the enhanced fear conditioning was also related to slower extinction (Wessa and Flor, 2007).
Deficient fear extinction in PTSD has been found in several studies that used skin conductance as the physiological measure (Peri et al., 2000; Bremner et al., 2005; Wessa and Flor, 2007). Blechert and colleagues examined fear acquisition and extinction using skin conductance and US expectancy and reported that delayed extinction in PTSD was paralleled by a continued expectation of the US during extinction (Blechert et al., 2007). Another recent study of Vietnam veterans and their twins found that PTSD subjects did not have impaired extinction learning, but rather had less extinction retention on the day after acquisition and extinction compared to veterans without PTSD (Milad et al., 2008). Furthermore, impaired retention of extinction appeared to be an acquired trait related to the disorder since the twins of the PTSD subjects did not show the same impairment.
Given the large number of studies that have examined HPA function in PTSD (de Kloet et al., 2007), and the examination of startle in PTSD (Grillon and Baas, 2003) there is a surprising paucity in the literature on the relationship between HPA function and the startle response in humans. To our knowledge, only two studies have examined cortisol and startle response in healthy human volunteers. The first study found that diurnal variation in cortisol was inversely associated with startle magnitude, in that high morning cortisol was associated with low startle magnitude (Miller and Gronfier, 2006). The second study used fear-potentiated startle and found that high cortisol to dehydroepiandrosterone-sulfate (DHEA-S) ratios after fear conditioning were associated with increased startle potentiation (Grillon et al., 2006).
We have recently developed an experimental paradigm that allows for the independent evaluation of fear-potentiated startle and the inhibition of fear-potentiated startle (Jovanovic et al., 2005). Our earlier study found that PTSD symptom severity is associated with impaired inhibition of fear-potentiated startle (Jovanovic et al., 2008). The current study used this protocol to investigate the relationship between fear-potentiated startle and HPA function in individuals with PTSD. We hypothesized that ACTH hypersuppression in response to dexamethasone would be associated with fear-potentiated startle in PTSD patients.
Methods
Study Subjects
Ninety participants were recruited as part of a larger study investigating the genetic and environmental factors that contribute to PTSD in a primarily African-American, low socioeconomic, inner city population. Exclusion criteria for participation in the study included active psychosis and major medical illnesses as assessed by health and physical examinations conducted by medical professionals. A urine pregnancy screen was used to exclude pregnant women from the dexamethasone test. Prior to their participation, all participants provided written informed consents that were approved by the Emory University Institutional Review Board.
Psychological Assessment
The Structured Clinical Interview for DSM IV, SCID, (First et al., 1995) was administered to all subjects. In addition to the diagnostic interview, all participants completed the Symptom Scale for PTSD and the Beck Depression Inventory for depression assessment. The Childhood Trauma Questionnaire was used to examine early adverse events and the Traumatic Events Inventory was used to assess adult levels of trauma.
Modified PTSD Symptom Scale
The modified PTSD Symptom Scale (PSS) is a psychometrically valid 17-item self-report scale assessing PTSD symptomatology over the two weeks prior to rating (Falsetti et al., 1993; Foa and Tolin, 2000; Schwartz et al., 2005). The PSS interview has been validated with the widely used measure of PTSD – the Clinician Administered PTSD Scale, CAPS; (Foa et al., 1993; Foa and Tolin, 2000). The categorical definition of PTSD was determined based on DSM-IV A–E criterion responses to the PSS questionnaire (A, presence of trauma; B, presence of at least 1 intrusive symptom; C, presence of at least 3 avoidance/numbing symptoms; and D, presence of at least 2 hyperarousal symptoms; and E, present for at least 1 month). A current diagnosis of PTSD was also confirmed by the SCID.
The Beck Depression Inventory (BDI) was administered to measure depressive symptoms. This interview was conducted at least 1-week prior to the conditioning session. The BDI consists of a 21-item questionnaire (Beck et al., 1961). Each of the items measures the presence and severity of depressive symptoms which are rated on a scale from 0 to 3. A categorical definition of MDD was determined by a cutoff score of 15 on the BDI. A current diagnosis of MDD was also confirmed by the SCID.
Childhood Trauma Questionnaire (CTQ)
The CTQ is a self-report inventory assessing childhood physical, sexual and emotional abuse. Studies have established the internal consistency, stability over time, and criterion validity of both the original 70-item CTQ and the current brief version (Bernstein and Fink, 1998; Bernstein et al., 2003). The CTQ yields a total score and subscale scores for each of the types of child abuse. The CTQ total score was used as a covariate in the analyses.
Traumatic Events Inventory (TEI)
The traumatic events inventory (Schwartz et al., 2005) assesses lifetime history of trauma exposure and is a measure of both child abuse and non-child abuse trauma. The TEI assesses past experience and frequency of 13 separate types of traumatic events as well as feelings of terror, horror, and helplessness with such events.
HPA Hormone Assays
Blood was drawn on two separate visits: the baseline visit, and the post-dexamethasone (post-DEX) visit. At the end of the baseline visit, medically cleared subjects and women with negative pregnancy tests were given a 0.5 mg dexamethasone tablet and instructed to take it at 11 pm the night before the next study visit. Subjects were instructed not to take anything by mouth other than water after midnight the night before each study visit. Fasting blood specimens were obtained by venipuncture between 8 a.m. and 9 a.m. on the mornings of the baseline and post-DEX visits. After collection, samples underwent routine processing and were transferred to a −80° C freezer for storage until analysis.
Baseline hormone measures included plasma cortisol and ACTH levels. Post-DEX measures included plasma cortisol and ACTH levels as well as a dexamethasone detection assay to confirm that subjects had taken the dexamethasone tablets as instructed. Cortisol, ACTH, and dexamethasone determinations were completed by the Yerkes Biomarkers Core Laboratory at Emory University. Cortisol and ACTH levels were measured by commercially available radioimmunoassay kits (Cortisol RIA from Diagnostic Systems Laboratories, Webster, TX and ACTH RIA from DiaSorin, Inc., Stillwater, MN). The presence or absence of Dexamethasone was ascertained by Elisa using commercially prepared kits from Neogen (Lexington, KY). All intra-assay and inter-assay coefficients of variability were under 10%.
Startle Response Measurements
Startle testing was conducted on the first visit, after the baseline blood draw. Given that the subjects had been fasting for the blood draw, they were given breakfast prior to the startle phase of the study. All subjects were screened for auditory impairment using an audiometer (Grason-Stadler, Model GS1710). The subjects were required to detect tones at 30 dB(A)SPL at frequencies ranging from 250 to 4000 Hz. The startle response data were acquired using Biopac MP150 for Windows (Biopac Systems, Inc., Aero Camino, CA) and stored on the hard drive of a Windows XP laptop. All data were sampled at 1000 Hz and amplified with a gain of 5000 using the EMG module of the Biopac system. The acquired data were filtered, rectified, and smoothed using MindWare software (MindWare Technologies, Ltd., Gahanna, OH) and exported for statistical analyses. The EMG signal was filtered with low- and high- frequency cutoffs at 28 and 500 Hz, respectively. The maximum amplitude of the eyeblink muscle contraction 20 – 200 ms after presentation of the startle probe was used as a measure of the acoustic startle response.
As previously described (Jovanovic et al., 2005; Jovanovic et al., 2006) the eyeblink component of the acoustic startle response was measured by EMG recordings of the right orbicularis oculi muscle with two 5-mm Ag/AgCl electrodes filled with electrolyte gel. One electrode was positioned 1cm below the pupil of the right eye and the other was 1cm below the lateral canthus. We used disposable electrodes from Biopac (EL504) pre-coated with electrolyte gel. Impedance levels were less than 6 kilo-ohms for each participant as measured by a Checktrode impedance meter (1089 MKIII, UFI, Morro Bay, CA). A background white noise of 70-dB (A) SPL was presented continuously throughout the session; startle probe delivery was superimposed on the background noise. The startle probe was a 108-dB (A) SPL, 40ms burst of broadband noise with near instantaneous rise time, delivered binaurally through headphones (Model TDH-39-P, Maico, Minneapolis, MN).
Response Keypad
A response keypad unit (SuperLab, Cedrus, Corp., San Pedro, CA) was incorporated into the startle session in order to assess trial-by-trial US expectancy. Subjects were instructed to respond on each CS trial by pressing one of three buttons: one when they expected the US, a second button when they did not expect the US, and a third button when they were uncertain of the contingency. The exact instructions given to the subjects were: “During this experiment you will hear some sudden tones and noises in addition to seeing several colored lights turn on. The tones are there to elicit startle and occur every time something happens. However, some of the lights will be followed by the blast of air while other lights will not. Throughout the experiment please press the button on the keypad to tell us whether you think a light will be followed by air (the plus sign), or will not be followed by air (the minus sign). If you do not know, press the 0 sign. You should press a button for each light.”
Experimental Design
Fear-potentiated startle and inhibition of fear-potentiated startle was assessed using a conditional discrimination paradigm termed AX+/BX− (Jovanovic et al., 2005). Each session consisted of a startle habituation phase followed by three blocks of conditioning that occurred without any breaks. The conditioning phase was seamlessly followed by a testing block for fear inhibition. Each conditioned stimulus (CS) was a compound of two shapes presented on a computer monitor. The AX+ compound served as the reinforced stimulus (CS+), and the BX− compound served as the non-reinforced stimulus (CS−). The AX+ and BX− cues consisted of a set of 2 blue, black or purple shapes (star, triangle or square) presented centrally on a monitor (with counterbalanced shape assignment across the CSs). Each compound CS had one novel cue (A or B) and one common cue ‘X’ (see Figure 1A and 1B). The fear inhibition test stimulus was a compound of the previously conditioned A and B cues (Figure 1C) that was used to determine transfer of inhibition (by B) to the fear response to A (Myers and Davis, 2004; Jovanovic et al., 2005). For each compound stimulus, the shapes were presented simultaneously and in one of two pseudorandom sequences. The aversive stimulus (US) was a 250 ms air blast with an intensity of 140 psi directed to the larynx. The air blast was emitted by a compressed air tank attached to the polyethylene tubing and controlled by a solenoid switch. This US has been used in our studies previously (Jovanovic et al., 2005; Norrholm et al., 2006) and produces robust fear-potentiated startle.
Figure 1. Examples of the CSs presented on a computer monitor during conditioning.
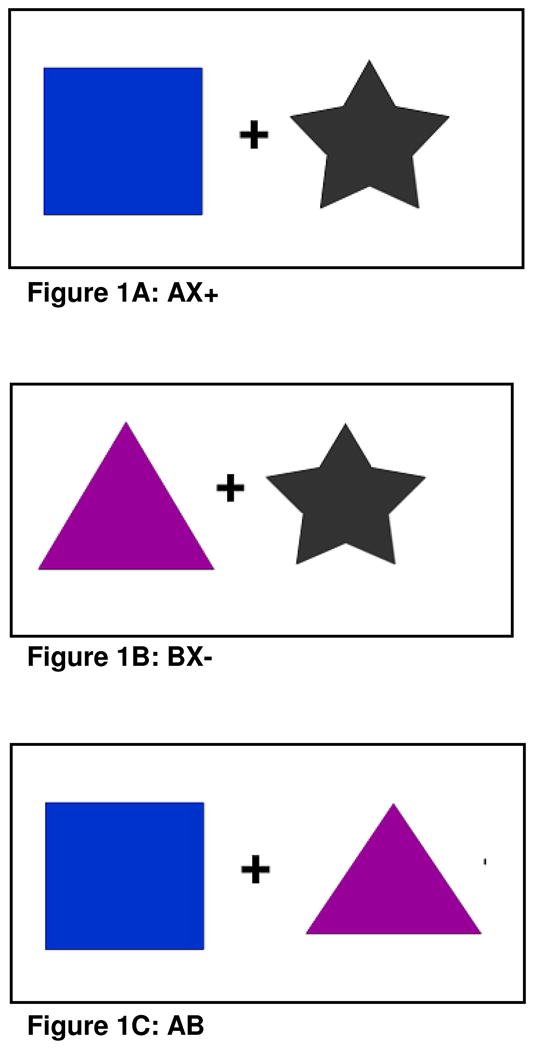
A. AX+, the reinforced stimulus, i.e. “the danger signal”; B. BX−, the non-reinforced stimulus, “the safety signal”; and C. AB, conditioned inhibition test trial, i.e., “transfer of safety”. Shape and color assignment was counterbalanced across subjects.
The habituation phase consisted of six startle probes presented alone (noise-alone trials, NA). Immediately following habituation, participants underwent the conditioning phase, which consisted of three blocks, each of which included four trials of each CS type and four NA trials for a total of 12 trials per block. A block of three AB trials was presented after the conditioning phase. All AX+ trials were reinforced with the US, while the BX− trials were not reinforced. Both conditioned stimuli were 6 sec in duration. During AX+ trials, the 250 ms air blast co-terminated with the stimulus, and the 40ms startle probe preceded the US (air blast) by 500 ms. The BX− trials terminated immediately after the presentation of the startle probe. The AB trials were designed the same as the BX− trials. In all phases of the experiment, inter-trial intervals were of randomized duration ranging from 9 to 22 seconds.
Data Analysis
Demographic and clinical data such as age, PTSD and depression symptoms, as well as childhood and adult trauma levels were compared between the PTSD and Non-PTSD groups using multivariate analyses of variance (MANOVA); categorical data, such as sex and race were analyzed using Chi-square analyses.
HPA data were analyzed separately for baseline cortisol and ACTH and post-DEX cortisol and ACTH. In addition, a change score was calculated for cortisol and ACTH by subtracting post-DEX levels from baseline levels. This resulted in 6 dependent variables that were analyzed using a MANOVA with the between groups factor of diagnosis group (2 levels). In order to account for the effects of current symptoms of depression, we included the BDI categorical score as a covariate in the multivariate analyses. Dexamethasone suppression was tested using a repeated measures analysis of variance (RM ANOVA) comparing baseline and post-DEX cortisol and ACTH as within-subjects factors separately for each diagnostic group. The BDI score was used as a covariate in these analyses as well.
Fear-potentiated startle was assessed by comparing startle magnitude to the noise alone (NA) trials to startle magnitude on CS+ trials (AX+) and CS− trials (BX−) in each conditioning block. All startle magnitude variables met the homogeneity of variance assumption. These variables were analyzed in a 3-way mixed ANOVA with the within-subject factor of block (3 levels), trial type (3 levels), and the between-groups factor of diagnosis (2 levels), with BDI score as a covariate in order to control for comorbidity with depression. However, significant differences in Non-PTSD and PTSD groups were also followed up with analyses without BDI as a covariate. Fear conditioning was assessed by the contrast of NA to AX+, and discrimination between the CS+ and CS− was assessed by the contrast between AX+ and BX−.
The degree of differential conditioning to AX+ and BX−, and fear inhibition to AB was measured by calculating percent potentiation for each CS type, in order to account for individual differences in startle magnitude as well as startle habituation. This value was derived as follows: Percent Startle Potentiation = 100 × (startle magnitude during CS trials – NA startle)/(NA startle), with the NA derived from the same conditioning block as the CS. The percent potentiation variables for AX, BX, and AB met the homogeneity of variance assumption. These variables were compared between groups using a MANOVA. In these analyses we also included the BDI score as a covariate. In order to assess whether there were group differences in CS discrimination, we used a RM ANOVA of CS type with a contrast comparing AX+ to BX− as a within-subjects factor separately for each group. Similarly, we tested fear inhibition by comparing AX+ to AB in a RM ANOVA within each group. Contingency awareness was analyzed by comparing US expectancy using keypad responses to AX+ and BX− with a RM MANOVA within each group. Additionally, the distribution of aware vs. unaware participants was compared between groups using a Chi-square analysis.
In all MANOVAs we used Roy’s Largest Root statistic with an alpha level of 0.05. All RM ANOVAs used the Sphericity Assumed statistic given that all analyses involved only 2 levels. All analyses were performed in SPSS 15.0 for Windows (SPSS, Inc., Chicago, IL).
In order to investigate the association between hormones and startle data, we ran partial two-tailed correlations between the HPA hormone variables (baseline and post-DEX) and the startle potentiation variables. Three fear-potentiated startle variables were generated: an average of the percent potentiation across blocks for AX and BX, and percent potentiation to the for the test block of AB trials. These variables were square root transformed in order to control for any effect of outliers. We examined these correlations separately in the PTSD and Non-PTSD group. Again, BDI was used as a covariate in the analyses. In these analyses we used an alpha level of 0.01 in order to control for Type I error without over-inflating Type II error rate.
Results
Subject Characteristics
HPA and startle data were collected on 90 subjects. Of these, 55 returned for the second, post-dexamethasone visit. One subject did not test positive for dexamethasone on the post-DEX day and their post-DEX data were not included in the analyses. The resulting sample had 90 subjects (29 PTSD and 61 Non-PTSD) with startle and baseline hormone data, and 54 subjects (18 PTSD and 36 Non-PTSD) with post-DEX data. We compared the subjects for the baseline sample and the subjects for the post-DEX sample on clinical data (PSS and BDI score), fear-potentiated startle data, and baseline cortisol and ACTH data—the two samples did not differ on any of these variables. All analyses of hormone data, including the correlations with startle were performed only on the 54 subjects who had both baseline and post-DEX data. All startle analyses were conducted on the 90 subjects who had baseline data. In the entire sample, the subjects’ age ranged from 18–63 years old and 61.3% were female. All subjects were African American. Table 1 list the rates of traumas in the total sample. Table 2 shows the demographic and clinical information of the subjects across the different groups, as well as trauma history.
Table 1.
Rates of traumatic experiences in the entire sample.
| TRAUMATIC EVENTS INVENTORY | Percent of Sample |
|---|---|
| Natural Disaster | 27.7 |
| Serious Accident or Injury | 54.7 |
| Sudden Life-Threatening Illness | 39.8 |
| Military Combat | 6.2 |
| Close Friend or Family Member Murdered | 6.1 |
| Close Friend or Family Member Committed Suicide | 7.5 |
| Attacked with Weapon | 32.5 |
| Attacked Without Weapon | 30.0 |
| Beaten as a Child | 31.7 |
| Sexual Contact Before Age 13 | 24.4 |
| Forced Sexual Contact Between 14 and 17 | 10.0 |
| Forced Sexual Contact After Age 17 | 10.3 |
Table 2.
The table displays the demographic and clinical information across the two diagnostic groups.
| PTSD N=29 | NON-PTSD N=61 | P | |
|---|---|---|---|
| SEX: | |||
| MALE | 34.5% | 40.4% | Ns |
| FEMALE | 65.5% | 59.6% | Ns |
| AGE (MEAN, SD) | 44.4 (12.0) | 45.2 (12.2) | Ns |
| CTQ SCORE (MEAN, SD) | 42.1 (22.6) | 44.5 (17.6) | Ns |
| TEI SCORE (MEAN, SD) | 4.1 (2.4) | 3.2 (2.0) | Ns |
| BDI SCORE (MEAN, SD) | 17.6 (12.7) | 10.7 (8.6) | <0.001 |
| PSS SCORE (MEAN, SD) | 22.9 (9.2) | 5.6 (5.3) | <0.001 |
Acronyms: CTQ: Childhood Trauma Questionnaire; TEI: Traumatic Events Inventory (mean indicates number of traumatic events experienced) BDI: Beck Depression Inventory; PSS; PTSD Symptoms Scale.
Clinical Assessments
As shown in Table 2, all subjects had approximately equivalent levels of childhood (F(1,73)=0.24, p=0.63) and adult (F(1,73)=2.36, p=0.13) trauma. However, the subjects that met current criteria for PTSD had significantly higher PSS scores, F(1,73)=105.18, p<0.0001, than traumatized controls without PTSD. Furthermore, the PTSD subjects had significantly greater current symptoms of depression compared to Non-PTSD controls (F(1,73)=7.25, p<0.01). Subjects were excluded from the study if they had schizophrenia. The prevalence of other current comorbid Axis I disorders, as assessed by the SCID, were as follows: major depression, 12.5% of sample (no difference in distribution between groups, χ2=0.20,p=0.67)—depression symptoms were also controlled statistically by covarying for BDI score; bipolar disorder, 1.3% of sample (only in Non-PTSD group, χ2=0.49,p=0.48); alcohol dependence, 6% of total sample (no difference in distribution between groups, χ2=2.57,p=0.11); drug dependence, 5% of sample (equally distributed between groups, χ2=0.08,p=0.78); panic disorder, 2.5% of sample (equally distributed between groups, χ2=0.34,p=0.56); social phobia, 3.9% of total sample (all of those were in the PTSD group, χ2=6.89,p<0.01); specific phobia, 8% of sample (equally distributed between groups, χ2=1.51,p=0.22); obsessive-compulsive disorder, 1.3% of sample (equally distributed between groups, χ2=2.10,p=0.15); and generalized anxiety disorder, 12.3% of sample (trend for higher likelihood in PTSD group, χ2=3.65,p=0.06).
HPA Function
Figure 2A shows baseline and post-DEX cortisol data, and figure 2B shows baseline and post-DEX ACTH data in the diagnostic groups. Multivariate analyses of variance (MANOVA) were conducted on baseline and post-DEX levels of cortisol and ACTH, as well as change scores for the two variables, comparing PTSD and Non-PTSD subjects. Current depression was included as a covariate in these analyses. There were no group differences in baseline hormone levels, or change scores from baseline to post-DEX levels. However, there was a significant main effect of group on post-DEX cortisol levels, with PTSD subjects having lower cortisol levels after dexamethasone compared to Non-PTSD subjects (F(1,42)=3.86,p=0.05).
Figure 2. Baseline and post-dexamethasone (post-DEX) levels and change scores of HPA hormones across the two diagnostic groups.

A. Mean+SE cortisol levels. * denotes p<0.05.
B. Mean+SE adenocorticotropin hormone (ACTH) levels. * denotes p<0.05; ** denotes p<0.01; *** denotes p<0.001
A RM ANOVA of dexamethasone effects showed a significant suppression of cortisol levels in both groups: PTSD, F(1,14)=17.67, p<0.001; Non-PTSD subjects, F(1,31)=53.94, p<0.01, see Figure 2A. ACTH levels were also significantly suppressed in the PTSD group, F(1,13)=5.36, p<0.05 and the Non-PTSD group (F(1,28)=10.13, p<0.01), see Figure 2B.
Fear-Potentiated Startle
Startle magnitude was assessed during noise alone (NA) trials and in the presence of conditioned stimuli across 3 blocks of conditioning using a 3-way mixed ANOVA with block (3 levels) X trial type (NA, AX+, BX−) X group (PTSD, Non-PTSD) and BDI score as a covariate. Fear acquisition was assessed using a contrast between NA and the reinforced stimulus (AX+), while differential conditioning to the CS+ and CS− was assessed by contrasting AX+ to BX. There was a trend toward a three-way interaction for fear potentiation (F(1,79)=3.80, p=0.06) and differential conditioning (F(1,79)=3.26,p=0.08). There was no main effect of group on startle magnitude to the NA and CS trials.
The interactions were followed up by separate two-way RM ANOVAs of block X trial type within each group. Figure 3 shows startle magnitude to the CS+ and CS− across the conditioning blocks for each group. In Non-PTSD subjects there was a significant linear effect for block (F(1,53)=21.29, p<0.001), while the PTSD subjects did not show a significant effect of block (F(1,25)=2.55, p=0.13) indicating deficient habituation to the startle stimulus. Fear conditioning was significant in both groups, as startle magnitude to AX+ was greater than NA in PTSD subjects (F(1,25)=4.18, p=0.05) and Non-PTSD subjects (F(1,53)=15.41, p<0.001), see Figure 3. Finally, startle magnitude was larger in the presence of AX+ than BX− in Non-PTSD (F(1,53)=5.03, p<0.05), but not in PTSD (F(1,25)=0.04, p=0.85) subjects. In order to determine that PTSD subjects were not simply slower at acquiring the discrimination, we compared startle magnitude on AX+ and BX− trials on the last block of conditioning in each group. Again, the Non-PTSD subjects had significant discrimination (F(1,53)=4.31, p<0.05), while the PTSD subjects did not (F(1,25)=0.22, p=0.65). The two-way RM ANOVAs within each group were also conducted without using BDI as a covariate, and the results were replicated, i.e., Non-PTSD subjects showed startle magnitude discrimination between AX+ and BX− (F(1,60)=4.17, p<0.05) and the PTSD subjects did not discriminate (F(1, 28)=0.39, p=0.54).
Figure 3.

Startle magnitude to Noise alone (NA), AX+ trials (danger signal), and BX− trials (safety signal) during the conditioning phase for the two groups. * denotes p<0.05 for trial type (NA vs. AX+, p<0.05; AX+ vs. BX−, ns); *** denotes p<0.001 for trial type (NA va AX+, p<0.001; AX+ vs. BX−, p<0.05); † denotes p<0.001 for Block
The degree of differential conditioning to AX+ and BX−, and fear inhibition on AB trials was assessed with a MANOVA comparing the groups on average percent startle potentiation to AX+, BX−, and AB. Computing percent potentiation from noise alone in each block accounted for individual variability in startle magnitude as well as habituation to the startle stimulus. This analysis showed a significant main effect of group on BX− trials (F(1,88)=4.44, p<0.05) and AB trials (F(1,88)=5.20, p<0.05), with higher fear potentiation in PTSD compared to Non-PTSD subjects, indicative of less fear inhibition, see Figure 4. In order to examine the reduction in the percent fear potentiation to BX− and AB relative to AX+ in PTSD and Non-PTSD subjects, a RM ANOVA of CS trial type was performed within each group. We found significant discrimination between AX+ and BX− in the controls, F(1,60)=14.52, p<0.001 while PTSD subjects did not show differential conditioning (F(1,28)=1.86,p=0.18), replicating the finding in startle magnitude. The contrast between AB and AX+ indicated that controls had significant fear inhibition on the AB trials, F(1,60)=7.14, p=0.01, while PTSD subjects did not (F(1,28)=0.05, p=0.82), Figure 4.
Figure 4.

Percent fear-potentiated startle to AX+ (danger signal), BX− (safety signal), and AB (safety transfer) across the two diagnostic groups. * denotes p<0.05; ** denotes p<0.01.
We also examined the correlation between percent fear potentiation on AX+, BX−, and AB with PTSD symptom severity in the three major symptom clusters: re-experiencing, avoidance and hyper-arousal symptoms. Percent potentiation to the nonreinforced cue, BX−, was positively correlated with avoidance symptoms (r(77)=0.23,p<0.05) and hyper-arousal symptoms (r(77)=0.23,p<0.05) on the PSS. Percent potentiation to the fear inhibition test trials, AB, was positively correlated with avoidance symptoms (r(77)=0.23,p<0.05). However, there were no significant correlations with startle to the danger signal, AX+.
Contingency Awareness
The results of the response keypad data showed that, across both groups, subjects understood the experimental contingencies. Four subjects (2 PTSD and 2 Non-PTSD) had missing response pad data. A mixed ANOVA in the two groups showed a main effect of CS type, with higher US expectancy for AX+ than BX−: PTSD (F(1, 26)=13.93, p<0.001), Non-PTSD (F(1, 58)=52.44, p<0.001). However, there were no between-group differences in US expectancy to either trial type and there were no CS type by group interactions. We used the response keypad data to categorize individuals as to their awareness of the reinforcement contingencies in the experiment. Individuals were considered aware if they had two consecutive correct responses on the keypad. In this sample, 20.9% of the subjects were classified as unaware; however, the distribution of aware and unaware subjects did not vary in the two diagnostic groups (χ2 (86)=0.59, p=0.44).
In order to eliminate the effect of unawareness on discrimination, we re-analyzed the differential conditioning data by restricting the dataset to only the aware subjects. We compared startle on the last conditioning block of AX+ to BX−, with BDI again as a covariate. These analyses replicated the earlier findings: Non-PTSD subjects had significantly higher startle magnitude in the presence of AX+ than BX− (F(1,40)=11.51,p<0.01), while PTSD subjects did not (F(1,16)=1.05, p=0.32). Similarly, percent startle potentiation on AX+ trials was significantly higher than percent potentiation on BX− trials in the controls (F(1,40)=6.81,p=0.01) but not the PTSD subjects (F(1,16)=0.004, p=0.95).
HPA and Fear-Potentiated Startle Correlations
We performed partial correlations on the HPA and startle data separately for each group, with depression as a covariate. Table 3 shows the correlations for all variables. We found that the PTSD group had a positive correlation between ACTH levels and fear-potentiated startle. This finding was true of baseline ACTH levels and post-DEX ACTH levels, and the relationship held for each trial type, see Table 3. Figure 5A shows the scatter diagram of startle to AX+ and BX− with baseline ACTH, and Figure 5B shows the scatter diagram of startle to AX+ and BX− with post-DEX ACTH. Cortisol levels were negatively associated with fear-potentiated startle in PTSD; however, the level of significance did not meet the criterion for multiple comparisons.
Table 3.
Correlations between HPA variables and fear-potentiated startle variables (square root) in PTSD and Non-PTSD subjects.
| Baseline Cortisol (g/L) | Baseline ACTH (pg/L) | Post-DEX Cortisol (g/L) | Post-DEX ACTH (pg/L) | Change Score Cortisol (g/L) | Change Score ACTH (pg/L) | |
|---|---|---|---|---|---|---|
| Startle to AX | ||||||
| PTSD | −0.49 | 0.71** | −0.28 | 0.67** | −0.41 | 0.17 |
| NON-PTSD | −0.04 | 0.10 | −0.12 | 0.11 | 0.05 | −0.03 |
| Startle to BX | ||||||
| PTSD | −0.17 | 0.53* | −0.17 | 0.79** | 0.13 | −0.22 |
| NON-PTSD | −0.04 | −0.13 | 0.05 | 0.00 | −0.07 | −0.20 |
| Startle to AB | ||||||
| PTSD | −0.52 | 0.66** | −0.35 | 0.52* | −0.44 | 0.27 |
| NON-PTSD | 0.09 | 0.07 | 0.03 | −0.20 | 0.05 | 0.39 |
p=0.01;
p<0.005
Figure 5. Scatter diagram of the percent fear-potentiated startle (square root) to AX+ (danger signal) and BX− (safety signal) and adenocorticotropin hormones (ACTH) in PTSD subjects.

A. Baseline ACTH and fear-potentiatiated startle, AX+ (p=0.01); BX− (p=0.05).
B. Post-DEX ACTH and fear-potentiatiated startle, AX+ (p<0.01); BX− (p<0.01).
Discussion
The objective of this study was to examine the relationship between fear-potentiated startle and HPA function in individuals with PTSD. We found that individuals who met criteria for PTSD had higher potentiation of the startle response to safety signals than traumatized controls and furthermore did not demonstrate startle magnitude discrimination between danger and safety cues. Importantly, impaired discrimination was limited to the startle data; the contingency awareness data demonstrates that both groups learned to expect the US only on the reinforced trials. The discrepancy between the awareness data and the startle data suggests that PTSD patients have difficulty appropriately responding to safety cues physiologically, i.e. they cannot inhibit the fear response even when they know they are safe. We also found that PTSD patients did not inhibit fear on the safety transfer trials; however, this finding may be secondary to the lack of physiological discrimination. Fear-potentiated startle to the safety cue was positively correlated with avoidance and hyper-arousal symptoms of PTSD.
These civilian PTSD data are consistent with our previous study from combat populations that found that PTSD symptom severity was associated with impaired inhibition of fear-potentiated startle (Jovanovic et al., 2009). One major difference between these studies is that the combat PTSD patients did learn to discriminate between danger sand safety cues, but could not use that knowledge to transfer safety on the conditioned inhibition trials. One possible explanation for this difference is that our civilian subjects were still living in their traumatizing environments, while the war veterans were removed from the combat zone both spatially and temporally—safety cues may not be salient events in a high trauma, inner-city environment.
The results of the present study are consistent with an increasing number of investigations that have found deficits in safety signal processing associated with PTSD (Peri et al., 2000; Bremner et al., 2005; Blechert et al., 2007; Wessa and Flor, 2007; Milad et al., 2008; Jovanovic et al., 2009; Lissek et al., 2009). A small number of prospective studies suggest that impaired suppression of fear during extinction may be a risk factor for PTSD (Guthrie and Bryant, 2006; Pole et al., 2009). While cognitive measures of safety signal learning parallel the physiological findings in some studies (Blechert et al., 2007; Lissek et al., 2009), our results show a dissociation between cognitive awareness of safety and startle potentiation, in that PTSD subjects who were aware that they would not receive the aversive airblast still showed heightened responding to the CS−. Exaggerated fear responses and deficient fear inhibition in PTSD are supported by neuroimaging studies showing amygdala hyperactivity in these patients in response to fearful stimuli (Shin et al., 2006; Liberzon and Sripada, 2007).
A novel finding in the present study was the positive correlation between fear-potentiated startle and plasma ACTH. Both PTSD and Non-PTSD subjects had lower levels of cortisol and ACTH in response to dexamethasone, as has been found in previous studies (Yehuda et al., 2004b; de Kloet et al., 2007); however the groups did not differ in the degree of cortisol or ACTH suppression. Interestingly, we found that baseline and post-DEX ACTH levels were positively correlated with startle responses to danger and safety cues in the PTSD subjects, but not in the controls. On the other hand, neither baseline nor post-DEX cortisol were associated with startle.
The differing results between the resilient individuals and PTSD patients may point to an underlying mechanism for the observed psychopathology. A positron emission tomography study of PTSD patients during fear conditioning indicated that they had higher amygdala activation during acquisition and decreased anterior cingulated cortex activation during extinction compared to controls (Bremner et al., 2005). Resilient individuals may have good inhibitory control of the amygdala by the prefrontal cortex (Milad et al., 2007) which would be reflected in lower potentiation on the safety and fear inhibition trials; these individuals also do not appear to have hypersensitivity of the HPA feedback function which has been reported in PTSD (Yehuda et al, 2004). On the other hand, PTSD patients show higher fear potentiation to the BX− trials (i.e., safety signals) and AB trials (i.e. less transfer of inhibition), associated with increased ACTH levels. These results appear to contradict earlier findings in which PTSD symptoms are associated with hypersuppression of ACTH following DEX (Yehuda et al, 2002,Yehuda et al, 2004 Yehuda et al, 2006). It is possible that increased fear-potentiated startle on these trials is a marker of CRH hypersecretion in the amygdala, which could result in higher baseline ACTH levels as well as interfere with DEX suppression at the level of the pituitary. This possibility is supported by recent findings that overexpression of CRH in the amygdala interferes with DEX feedback on the HPA axis and increases acoustic startle in female rats (Keen-Rhinehart et al., 2008). The two studies that have examined the startle response in relation to cortisol in humans would support this idea since low diurnal cortisol during rest is associated with high CRH levels (Miller and Gronfier, 2006). On the other hand, high CRH levels in the amygdala could increase both the cortisol/DHEA-S ratio and fear-potentiated startle (Grillon et al., 2006). Given these two studies, it is unclear why our data do not show an association between startle and cortisol. Possibly, the dexamethasone suppression effect reduced cortisol levels to such a low level that any significant relationship between these two factors was obscured.
A limitation of the present study is that it relies on correlational data; thus we cannot determine the cause of either impaired differential conditioning or HPA function. Although we controlled for demographic data (sex, age and race) and levels of childhood and adult trauma by having groups that were matched on these variables, another limitation is the lack of control of many other variables that could influence HPA function, such as body mass index, gonadal hormone levels, medication status, or substance abuse and dependence. A replication of the study with a larger sample size with enough power to covary for these variables would provide strong support for these findings and may replicate the reports in the literature with respect to ACTH hypersuppression. It is also important to keep in mind that PTSD is a heterogeneous disorder and that individual patients may vary in the degree to which they present with different symptom clusters. Therefore, it is unlikely that a single neurobiological system, such as amygdala hyperactivation, will explain all aspects of this complex disease. However, each biomarker can contribute valuable information on vulnerability to the development of the disorder as well as treatment resistance.
In conclusion, this study found that PTSD was associated with greater fear-potentiated startle to safety cues and a lack of differential fear-potentiated startle to danger and safety cues, despite showing intact cognitive discrimination learning between these cues. Higher fear-potentiation to danger as well as safety cues was associated with higher baseline and post-DEX ACTH levels in PTSD. Taken together, the psychophysiological and neuroendocrine findings suggest that deficient responding to safety cues, or over-generalized responding to danger cues, may serve as an intermediate biological phenotype for PTSD; such indices are of crucial importance to progress in this field.
Acknowledgments
This work was primarily supported by National Institutes of Mental Health (MH071537). Support was also Emory and Grady Memorial Hospital General Clinical Research Center, NIH National Centers for Research Resources (M01 RR00039), and the Burroughs Wellcome Fund. We thank Allen Graham, BA, Joshua Castleberry, BS, Daniel Crain, BS, Abby Powers, BS, Rachel Herschenberg, BS as well as the nurses and staff of the Grady GCRC for their assistance with data collection and support.
Footnotes
Publisher's Disclaimer: This is a PDF file of an unedited manuscript that has been accepted for publication. As a service to our customers we are providing this early version of the manuscript. The manuscript will undergo copyediting, typesetting, and review of the resulting proof before it is published in its final citable form. Please note that during the production process errors may be discovered which could affect the content, and all legal disclaimers that apply to the journal pertain.
References
- Baker DB, West SA, Nicholson WE, Ekhator NN, Kasckow JW, Hill KK, Bruce AB, Orth DN, Geracioti TD. Serial CSF corticotropin-releasing hormone levels and adrenocortical activity in combat veterans with posttraumatic stress disorder. American Journal of Psychiatry. 1999;156:585–588. doi: 10.1176/ajp.156.4.585. [DOI] [PubMed] [Google Scholar]
- Beck AT, Ward CH, Mendelsohn M, Mock J, Erbaugh J. An inventory for measuring depression. Archives of General Psychiatry. 1961;4:561–571. doi: 10.1001/archpsyc.1961.01710120031004. [DOI] [PubMed] [Google Scholar]
- Bernstein DP, Fink L. Childhood Trauma Questionnaire A retrospective self-report manual. The Psychological Corporation; San Antonio, TX: 1998. [Google Scholar]
- Bernstein DP, Stein JA, Newcomb MD, Walker E, Pogge D, Ahluvalia T, Stokes J, Handelsman L, Medrano M, Desmond D, Zule W. Development and validation of a brief screening version of the Childhood Trauma Questionnaire. Child Abuse & Neglect. 2003;27:169–190. doi: 10.1016/s0145-2134(02)00541-0. [DOI] [PubMed] [Google Scholar]
- Blechert J, Michael T, Vriends N, Margraf J, Wilhelm FH. Fear conditioning in posttraumatic stress disorder: Evidence for delayed extinction of autonomic, experiential, and behavioural responses. Behaviour Research and Therapy. 2007;45:2019–2033. doi: 10.1016/j.brat.2007.02.012. [DOI] [PubMed] [Google Scholar]
- Bremner JD, Licinio J, Darnell A, Krystal JH, Owens MJ, Southwick SM, Nemeroff CB, Charney DS. Elevated CSF corticotropin-releasing factor concentrations in posttraumatic stress disorder. American Journal Psychiatry. 1997;154:624–629. doi: 10.1176/ajp.154.5.624. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bremner JD, Vermetten E, Schmahl C, Vaccarino V, Vythilingam M, Afzal N, Grillon C, Charney DS. Positron emission tomographic imaging of neural correlates of a fear acquisition and extinction paradigm in women with childhood sexual-abuse-related post-traumatic stress disorder. Psychological Medicine. 2005;35:791–806. doi: 10.1017/s0033291704003290. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Davis M. The role of the amygdala in fear-potentiated startle: implications for animal models of anxiety. Trends Pharmacol Sci. 1992;13:35–41. doi: 10.1016/0165-6147(92)90014-w. [DOI] [PubMed] [Google Scholar]
- Davis M, Gendelman DS, Tischler MD, Gendelman PM. A primary acoustic startle circuit: Lesion and stimulation studies. Journal of Neuroscience. 1982;6:791–805. doi: 10.1523/JNEUROSCI.02-06-00791.1982. [DOI] [PMC free article] [PubMed] [Google Scholar]
- de Kloet CS, Vermetten E, Heijnen CJ, Geuze E, Lentjes EG, Westenberg HG. Enhanced cortisol suppression in response to dexamethasone administration in traumatized veterans with and without posttraumatic stress disorder. Psychoneuroendocrinology. 2007;32:215–226. doi: 10.1016/j.psyneuen.2006.12.009. [DOI] [PubMed] [Google Scholar]
- Falsetti S, Resnick H, Resick P, Kilpatrick D. The Modified PTSD Symptom Scale: A brief self-report measure of posttraumatic stress disorder. The Behavior Therapist. 1993;16:161–162. [Google Scholar]
- First MB, Spitzer RL, Williams JBW, Gibbon M. Structured Clinical Interview for DSMIV-Patient Edition (SCID-P) American Psychiatric Press; Washington, D.C: 1995. [Google Scholar]
- Foa EB, Riggs DS, Dancu CV, Rothbaum BO. Reliability and validity of a brief instrument for assessing post-traumatic stress disorder. Journal of Traumatic Stress. 1993;6:459–473. [Google Scholar]
- Foa EB, Tolin DF. Comparison of the PTSD symptom scale-interview version and the clinician-administered PTSD scale. Journal of Traumatic Stress. 2000;13:181–191. doi: 10.1023/A:1007781909213. [DOI] [PubMed] [Google Scholar]
- Grillon C, Baas J. A review of the modulation of the startle reflex by affective states and its application in psychiatry. Clin Neurophysiol. 2003;114:1557–1579. doi: 10.1016/s1388-2457(03)00202-5. [DOI] [PubMed] [Google Scholar]
- Grillon C, Morgan CA., 3rd Fear-potentiated startle conditioning to explicit and contextual cues in Gulf War veterans with posttraumatic stress disorder. Journal of Abnormal Psychology. 1999;108:134–142. doi: 10.1037//0021-843x.108.1.134. [DOI] [PubMed] [Google Scholar]
- Grillon C, Pine DS, Baas JM, Lawley M, Ellis V, Charney DS. Cortisol and DHEA-S are associated with startle potentiation during aversive conditioning in humans. Psychopharmacology (Berl) 2006;186:434–441. doi: 10.1007/s00213-005-0124-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Guthrie RM, Bryant RA. Extinction learning before trauma and subsequent posttraumatic stress. Psychosomatic Medicine. 2006;68:307–311. doi: 10.1097/01.psy.0000208629.67653.cc. [DOI] [PubMed] [Google Scholar]
- Heim C, Newport DJ, Heit S, Graham YP, Wilcox M, Bonsall R, Miller AH, Nemeroff CB. Pituitary-adrenal and autonomic responses to stress in women after sexual and physical abuse in childhood. Journal of the American Medical Association. 2000;284:592–597. doi: 10.1001/jama.284.5.592. [DOI] [PubMed] [Google Scholar]
- Jovanovic T, Keyes M, Fiallos A, Myers KM, Davis M, Duncan EJ. Fear Potentiation and Fear Inhibition in a Human Fear-Potentiated Startle Paradigm. Biological Psychiatry. 2005;57:1559–1564. doi: 10.1016/j.biopsych.2005.02.025. [DOI] [PubMed] [Google Scholar]
- Jovanovic T, Norrholm SD, Fennell JE, Keyes M, Fiallos A, Myers KM, Davis M, Duncan EJ. Posttraumatic stress disorder may be associated with impaired fear inhibition: relation to symptom severity. Psychiatry Res. 2009;167:151–160. doi: 10.1016/j.psychres.2007.12.014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jovanovic T, Norrholm SD, Keyes M, Fiallos A, Jovanovic S, Myers KM, Davis M, Duncan EJ. Contingency awareness and fear inhibition in a human fear-potentiated startle paradigm. Behav Neurosci. 2006;120:995–1004. doi: 10.1037/0735-7044.120.5.995. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kalin NH, Takahashi LK. Fear-motivated behavior induced by prior shock experience is mediated by corticotropin-releasing hormone systems. Brain Research. 1990;509:80–84. doi: 10.1016/0006-8993(90)90311-x. [DOI] [PubMed] [Google Scholar]
- Keen-Rhinehart E, Michopoulos V, Toufexis DJ, Martin EI, Nair H, Ressler KJ, Davis M, Owens MJ, Nemeroff CB, Wilson ME. Continuous expression of corticotropin-releasing factor in the central nucleus of the amygdala emulates the dysregulation of the stress and reproductive axes. Mol Psychiatry. 2008;14:37–50. doi: 10.1038/mp.2008.91. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Koch M. The neurobiology of startle. Progress in Neurobiology. 1999;59:107–128. doi: 10.1016/s0301-0082(98)00098-7. [DOI] [PubMed] [Google Scholar]
- Landis C, Hunt W. The Startle Paradigm. Farrar and Rinehart; New York: 1939. [Google Scholar]
- Lee Y, Davis M. Role of the hippocampus, the bed nucleus of the stria terminalis, and the amygdala in the excitatory effect of corticotropin-releasing hormone on the acoustic startle reflex. Journal of Neuroscience. 1997;17:6434–6446. doi: 10.1523/JNEUROSCI.17-16-06434.1997. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lee Y, Lopez DE, Meloni EG, Davis M. A primary acoustic startle pathway: obligatory role of cochlear root neurons and the nucleus reticularis pontis caudalis. Journal of Neuroscience. 1996;16:3775–3789. doi: 10.1523/JNEUROSCI.16-11-03775.1996. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Liang KC, Melia KR, Miserendino MJD, Falls WA, Campeau S, Davis M. Corticotropin-releasing factor: long-lasting facilitation of the acoustic startle reflex. Journal of Neuroscience. 1992;12:2303–2312. doi: 10.1523/JNEUROSCI.12-06-02303.1992. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Liberzon I, Sripada CS. Progress in Brain Research. Elsevier; 2007. The functional neuroanatomy of PTSD: a critical review; pp. 151–169. [DOI] [PubMed] [Google Scholar]
- Lissek S, Powers AS, McClure EB, Phelps EA, Woldehawariat G, Grillon C, Pine DS. Classical fear conditioning in the anxiety disorders: a meta-analysis. Behaviour Research & Therapy. 2005;43:1391–1424. doi: 10.1016/j.brat.2004.10.007. [DOI] [PubMed] [Google Scholar]
- Lissek S, Rabin SJ, McDowell DJ, Dvir S, Bradford DE, Geraci M, Pine DS, Grillon C. Impaired discriminative fear-conditioning resulting from elevated fear responding to learned safety cues among individuals with panic disorder. Behaviour Research & Therapy. 2009;47:111–118. doi: 10.1016/j.brat.2008.10.017. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Milad MR, Orr SP, Lasko NB, Chang Y, Rauch SL, Pitman RK. Presence and acquired origin of reduced recall for fear extinction in PTSD: Results of a twin study. Journal of Psychiatric Research. 2008;42:515–520. doi: 10.1016/j.jpsychires.2008.01.017. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Milad MR, Wright CI, Orr SP, Pitman RK, Quirk GJ, Rauch SL. Recall of Fear Extinction in Humans Activates the Ventromedial Prefrontal Cortex and Hippocampus in Concert. Biological Psychiatry. 2007;62:446–454. doi: 10.1016/j.biopsych.2006.10.011. [DOI] [PubMed] [Google Scholar]
- Miller MW, Gronfier C. Diurnal variation of the startle reflex in relation to HPA-axis activity in humans. Psychophysiology. 2006;43:297–301. doi: 10.1111/j.1469-8986.2006.00400.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Myers KM, Davis M. AX+, BX− discrimination learning in the fear-potentiated startle paradigm: possible relevance to inhibitory fear learning in extinction. Learning & Memory. 2004;11:464–475. doi: 10.1101/lm.74704. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Norrholm SD, Jovanovic T, Vervliet B, Myers KM, Davis M, Rothbaum BO, Duncan EJ. Conditioned fear extinction and reinstatement in a human fear-potentiated startle paradigm. Learning & Memory. 2006;13:681–685. doi: 10.1101/lm.393906. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Orr SP, Metzger LJ, Lasko NB, Macklin ML, Peri T, Pitman RK. De novo conditioning in trauma-exposed individuals with and without posttraumatic stress disorder. Journal of Abnormal Psychology. 2000;109:290–298. [PubMed] [Google Scholar]
- Peri T, Ben-Shakhar G, Orr SP, Shalev AY. Psychophysiologic assessment of aversive conditioning in posttraumatic stress disorder. Biological Psychiatry. 2000;47:512–519. doi: 10.1016/s0006-3223(99)00144-4. [DOI] [PubMed] [Google Scholar]
- Plotsky PM, Meaney MJ. Early, postnatal experience alters hypothalamic corticotropin-releasing factor (CRF) mRNA, median eminence CRF content and stress-induced release in adult rats. Molecular Brain Research. 1993;18:195–200. doi: 10.1016/0169-328x(93)90189-v. [DOI] [PubMed] [Google Scholar]
- Pole N, Neylan TC, Otte C, Henn-Hasse C, Metzler TJ, Marmar CR. Prospective Prediction of Posttraumatic Stress Disorder Symptoms Using Fear Potentiated Auditory Startle Responses. Biological Psychiatry. 2009;65:235–240. doi: 10.1016/j.biopsych.2008.07.015. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Roozendaal B, Brunson KL, Holloway BL, McGaugh JL, Baram TZ. Involvement of stress-released corticotropin-releasing hormone in the basolateral amygdala in regulating memory consolidation. Proceedings of the National Academy of Sciences of the United States of America. 2002;99:13908–13913. doi: 10.1073/pnas.212504599. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Schwartz AC, Bradley RL, Sexton M, Sherry A, Ressler KJ. Posttraumatic Stress Disorder Among African Americans in an Inner City Mental Health Clinic. Psychiatr Serv. 2005;56:212–215. doi: 10.1176/appi.ps.56.2.212. [DOI] [PubMed] [Google Scholar]
- Shin LM, Rauch SL, Pitman RK. Amygdala, Medial Prefrontal Cortex, and Hippocampal Function in PTSD. Ann NY Acad Sci. 2006;1071:67–79. doi: 10.1196/annals.1364.007. [DOI] [PubMed] [Google Scholar]
- Sutton RE, Koob GF, Le Moal M, Rivier J, Vale W. Corticotropin releasing factor produces behavioural activation in rats. Nature. 1982;297:331–333. doi: 10.1038/297331a0. [DOI] [PubMed] [Google Scholar]
- Swerdlow NR, Britton KT, Koob GF. Potentiation of acoustic startle by corticotropin-releasing factor (CRF) and by fear are both reversed by alpha-helical CRF (9–41) Neuropsychopharmacology. 1989;2:285–292. doi: 10.1016/0893-133x(89)90033-x. [DOI] [PubMed] [Google Scholar]
- Weike AI, Schupp HT, Hamm AO. In dubio pro defensio: Initial activation of conditioned fear is not cue specific. Behavioral Neuroscience. 2008;122:685–696. doi: 10.1037/0735-7044.122.3.685. [DOI] [PubMed] [Google Scholar]
- Wessa M, Flor H. Failure of Extinction of Fear Responses in Posttraumatic Stress Disorder: Evidence From Second-Order Conditioning. American Journal Psychiatry. 2007;164:1684–1692. doi: 10.1176/appi.ajp.2007.07030525. [DOI] [PubMed] [Google Scholar]
- Yehuda R, Golier JA, Halligan SL, Meaney M, Bierer LM. The ACTH response to dexamethasone in PTSD. American Journal Psychiatry. 2004a;161:1397–1403. doi: 10.1176/appi.ajp.161.8.1397. [DOI] [PubMed] [Google Scholar]
- Yehuda R, Halligan SL, Golier JA, Grossman R, Bierer LM. Effects of trauma exposure on the cortisol response to dexamethasone administration in PTSD and major depressive disorder. Psychoneuroendocrinology. 2004b;29:389–404. doi: 10.1016/s0306-4530(03)00052-0. [DOI] [PubMed] [Google Scholar]
- Yehuda R, Halligan SL, Grossman R, Golier JA, Wong C. The cortisol and glucocorticoid receptor response to low dose dexamethasone administration in aging combat veterans and holocaust survivors with and without posttraumatic stress disorder. Biol Psychiatry. 2002;52:393–403. doi: 10.1016/s0006-3223(02)01357-4. [DOI] [PubMed] [Google Scholar]
- Yeomans JS, Frankland PW. The acoustic startle reflex: neurons and connections. Brain Research - Brain Research Reviews. 1995;21:301–314. doi: 10.1016/0165-0173(96)00004-5. [DOI] [PubMed] [Google Scholar]
- Yeomans JS, Li L, Scott BW, Frankland PW. Tactile, acoustic and vestibular systems sum to elicit the startle reflex. Neuroscience & Biobehavioral Reviews. 2002;26:1–11. doi: 10.1016/s0149-7634(01)00057-4. [DOI] [PubMed] [Google Scholar]