

Abstract
Bacterial secondary transporters of the DctA family mediate ion-coupled uptake of C4-dicarboxylates. Here, we have expressed the DctA homologue from Bacillus subtilis in the Gram-positive bacterium Lactococcus lactis. Transport of dicarboxylates in vitro in isolated membrane vesicles was assayed. We determined the substrate specificity, the type of cotransported ions, the electrogenic nature of transport, and the pH and temperature dependence patterns. DctA was found to catalyze proton-coupled symport of the four C4-dicarboxylates from the Krebs cycle (succinate, fumurate, malate, and oxaloacetate) but not of other mono- and dicarboxylates. Because (i) succinate-proton symport was electrogenic (stimulated by an internal negative membrane potential) and (ii) the divalent anionic form of succinate was recognized by DctA, at least three protons must be cotransported with succinate. The results were interpreted in the light of the crystal structure of the homologous aspartate transporter GltPh from Pyrococcus horikoshii.
The DctA family is one of several diverse families of secondary transporters that catalyze the uptake of C4-dicarboxylates from the Krebs cycle in bacteria (16, 27). In Escherichia coli, DctA mediates the uptake of succinate, fumurate, and malate under aerobic conditions; genomic disruption of dctA in E. coli prevents growth with malate or fumarate as the sole carbon source, and the mutant grows poorly on succinate (5). Similarly, a dctA knockout mutant of Bacillus subtilis cannot grow with succinate or fumarate as the sole carbon source (1). DctA plays a major role in the symbiotic relationship between nitrogen-fixing rhizobia (43) and root nodule-forming plants (30, 37, 38). Transport assays with Sinorhizobium meliloti cells showed previously that in addition to succinate, malate, and fumarate, orotate is transported and that a range of other substrates such as succinamic acid and succinamide may be transported, because they inhibit the transport of orotate (42). In Corynebacterium glutamicum, malate transport by DctA is inhibited by α-ketoglutarate, oxaloacetate, and glyoxylate, indicating that these compounds may be substrates also (41).
DctA transporters belong to a large family of secondary transporters (the DAACS [dicarboxylate/amino acid:cation symporter] family), which also comprises well-characterized glutamate/aspartate transporters and neutral amino acid transporters (32, 33). While DctA-type dicarboxylate transporters are found only in bacteria, glutamate/aspartate transporters of the DAACS family are found both in prokaryotes (e.g., GltT in Bacillus stearothermophilus, GltP in E. coli, and GltPh in Pyrococcus horikoshii [2, 7, 34]) and in higher eukarya, where they play a pivotal role in the reuptake of the excitatory neurotransmitter glutamate from the synaptic cleft (4). Neutral amino acid (alanine, serine, and threonine) transporters are found in mammals (see, e.g., references 36 and 44) as well as bacteria (17).
Secondary transporters of the DAACS family use (electro)chemical gradients of cations across the membrane to drive transport. The type of cotransported ions varies among family members: eukaryotic glutamate transporters couple the transport of glutamate to the symport of one proton and three sodium ions and the antiport of one potassium ion (24, 45). Bacterial and archaeal glutamate transporters utilize either sodium ions or protons for symport (2) and are independent of potassium ions (28, 31). The bacterial and mammalian neutral amino acid transporters are sodium ion coupled. Glutamate/aspartate transporters and bacterial serine/threonine transporters (SstTs) are electrogenic, but mammalian neutral amino acid transporters are obligate electroneutral amino acid antiporters (44).
Insight into the structure-function relationships of the DAACS family members has greatly increased since crystal structures of the P. horikoshii aspartate transporter GltPh have been determined (2, 29, 40). The protein consists of eight membrane-spanning helices and two reentrant regions (helical hairpins HP1 and HP2) (40). The C-terminal part of the protein (helices 7 and 8 and HP1 and HP2) is most strongly conserved with respect to other family members and binds the substrate and cotransported ions, with HP1 and HP2 functioning as lids that allow alternating access to the substrate- and ion-binding sites from either side of the membrane (3, 29). GltPh forms a homotrimeric complex in which each protomer functions independently of the other subunits (11, 12, 18, 19, 23). The fold and oligomeric state are likely to be conserved throughout the family.
Whereas the transport mechanisms of bacterial glutamate and neutral amino acid transporters of the DAACS family have been studied extensively in vitro, the C4-dicarboxylate transporters of the DAACS family (DctA proteins) have been studied using whole cells only. To fully characterize these transporters, in vitro activity assays using either membrane vesicles or proteoliposomes containing purified protein are necessary. In such assays, the internal and external buffer compositions can be controlled, thus allowing manipulation of the chemical ion gradients and the electrical potential across the membrane. Here, we present the first biochemical characterization of a DctA family member in membrane vesicles. We have studied the DctA homologue from B. subtilis, which is annotated as DctP (1) but which we propose to rename DctA to reflect the homology to other DctA proteins. B. subtilis DctA (DctABs) has 30 to 32% sequence identity to the aspartate transporter GltPh and human excitatory amino acid transporter (EAAT) family members, over 40% sequence identity to the characterized bacterial glutamate transporters from E. coli and B. stearothermophilus, and 41 and 56% identity to DctA homologues from C. glutamicum and E. coli, respectively. We determined the substrate specificity of DctABs, the type of cotransported ions, the electrogenic nature of transport, and the pH and temperature dependence patterns.
MATERIALS AND METHODS
Chemicals.
Restriction enzymes and polymerases were purchased from Fermentas Life Sciences (Burlington, CA). [14C]succinate (1.85 MBq/ml [2.04 GBq/mmol]) was purchased from ARC Radiochemicals (St. Louis, MO). Lipids were purchased from Avanti Polar Lipids (Alabaster, AL). All chemicals were of analytical grade.
DNA manipulations.
The dctA gene from B. subtilis was amplified from genomic DNA by PCR using Phusion polymerase according to the manufacturer's instructions. For expression in Lactococcus lactis, the PCR products were cloned into vector pRE-Nlic, essentially as described by Geertsma and Poolman (9). The pRE-Nlic vector was converted into a plasmid for expression in L. lactis by the vector-backbone exchange (VBeX) method (9). Vectors were designed in such a way that an N-terminal decahistidine tag followed by a tobacco etch virus (TEV) protease cleavage site was added to the protein product. DNA sequencing by Service XS (Leiden, Netherlands) confirmed that no unwanted mutations had occurred.
Expression and purification of DctABs.
DctABs was expressed in L. lactis NZ9000 cells (21) grown on M17 medium (Oxoid, Basingstoke, United Kingdom) supplemented with 1% glucose at a constant pH (6.5) and temperature (30°C). Expression was induced by the addition of nisin A to a 1:5,000 (vol/vol) dilution of the supernatant from a stationary-phase L. lactis NZ9700 culture at an optical density at 660 nm of ∼1.5 (20). Two hours after induction, cells were harvested with a Beckman JLA 8.1000 rotor at a relative centrifugal force [RCF] of 10,000 for 10 min at 4°C and resuspended in 20 mM Tris-HCl (pH 8). DNase (10 μg/ml), phenylmethylsulfonyl fluoride (PMSF; 200 μM), and MgSO4 (1 mM) were added, and cells were passed twice through a cell disrupter at 45,000 lb/in2 (Constant Systems Ltd., Daventry, United Kingdom) and cooled at 4°C. Unbroken cells and cell debris were pelleted using a Harrier 18/80 swing-out centrifuge at an RCF of 5,000 for 25 min at 4°C, and the supernatant was subjected to ultracentrifugation using a Beckman 50.2 Ti rotor at an RCF of 150,000 for 90 min at 4°C. Membrane pellets were resuspended in 20 mM Tris-HCl (pH 8) and stored at −80°C. The protein concentrations in the membranes were determined using Bradford reagent with bovine serum albumin as a standard.
For protein purification (7), membrane vesicles were solubilized in buffer A (50 mM Tris-HCl [pH 8.0], 300 mM NaCl), containing 15 mM imidazole (pH 8.0) and 1% n-dodecyl-β-d-maltopyranoside (DDM), at a final protein concentration of 5 mg/ml. After incubation on ice for 45 min with occasional gentle shaking, the solution was centrifuged with a Beckman TLA 100.4 rotor at an RCF of 267,000 for 15 min at 4°C. Supernatants were incubated on a rotating platform for 60 min at 4°C with a nickel-Sepharose slurry (Fast Flow; GE Healthcare) preequilibrated with buffer A containing 15 mM imidazole, pH 8.0. The mixture was loaded onto a Bio-Rad Poly-Prep column, and unbound protein was allowed to flow through. The column was washed with 20 column volumes of buffer A supplemented with 60 mM imidazole and 0.04% DDM. Protein was eluted from the column in three fractions of 300, 500, and 500 μl by using buffer A supplemented with 500 mM imidazole, pH 8.0, and 0.04% DDM. The second elution fraction from the affinity chromatography contained most of the purified protein and was loaded onto a Superdex-200 gel filtration column using 20 mM potassium phosphate, pH 7.0, 150 mM NaCl, and 0.04% DDM as eluents.
Throughout the purification procedure, samples were taken and analyzed by sodium dodecyl sulfate (SDS)-12.5% polyacrylamide gel electrophoresis. Gels were stained using Serva Blue G Coomassie brilliant blue.
Fused membrane vesicles.
L. lactis membrane vesicles were mixed in a 1:10 (wt/wt) ratio of protein (from the membrane vesicle preparation) to lipid (from liposomes) with liposomes made from E. coli polar lipids and egg phosphatidylcholine (3:1, wt/wt) that had been extruded nine times through a 400-nm-pore-size polycarbonate filter by using a miniextruder (Hamilton, Bonaduz, Switzerland). The mixture was frozen in liquid nitrogen, thawed three times, and subjected to centrifugation with a Beckman TLA 100.4 rotor at an RCF of 267,000 for 20 min at 4°C. Fused membranes were then resuspended in the luminal buffer of choice (see the figure legends), again frozen and thawed three times, and stored in liquid nitrogen. Freezing, thawing, and subsequent extrusion (see below) were used to load the vesicles with the desired buffer.
Transport assays using fused membrane vesicles.
Fused membranes were thawed, extruded as described above, and centrifuged with a Beckman TLA 100.4 rotor at an RCF of 267,000 for 20 min at 4°C. Membranes were resuspended in the internal (luminal) buffer to a total protein concentration of 12.5 μg/μl. In transport assays, fused membrane vesicles were diluted 100× into 200 μl of external buffer (described in Results) supplemented with [14C]succinate (a 3.1 μM concentration, unless otherwise indicated) and, if applicable, 0.5 μM valinomycin (3 mM stock solution in ethanol). All assays were performed at 25°C, except when otherwise indicated, and internal and external buffers were isosmotic. Transport reactions were stopped by the addition of 2 ml of ice-cold 100 mM LiCl, followed by rapid filtration with BA-85 nitrocellulose filters and an additional wash step with lithium chloride. Levels of radioactivity were determined by the addition of 2 ml of Emulsifier-Scintillator Plus liquid (PerkinElmer, Waltham, MA) and analysis with a PerkinElmer Tri-Carb 2800 TR isotope counter.
RESULTS
Expression and purification of DctABs.
Initially we aimed at the purification of DctABs and reconstitution in proteoliposomes. A histidine-tagged variant of the protein was expressed in the Gram-positive host L. lactis (22). The DctABs protein was extracted from L. lactis membrane vesicles by solubilization using the detergent DDM and purified by nickel affinity and size exclusion chromatography (Fig. 1A). The protein eluted at a volume of ∼11 ml from the size exclusion column (Fig. 1B). The trimeric prokaryotic glutamate/aspartate transporters of the DAACS family elute at similar volumes (10, 39, 40), indicating that DctABs is probably also a homotrimer. However, the size exclusion chromatogram showed that the protein was prone to aggregation in detergent solution. Static light-scattering (size exclusion chromatography-multiangle laser light-scattering [SEC-MALLS]) analysis of DctABs indeed showed that the elution peak was not monodisperse, but the results were consistent with a trimeric oligomeric state of the protein in the main peak (data not shown). Varying the buffer composition (by the addition of glycerol or salt) or the pH (from 6 to 8.5) did not improve the stability of the protein, nor did the addition of a substrate (succinate) to the buffers throughout the purification. The chromatogram for DctABs shown in Fig. 1B was the best obtained.
FIG. 1.

Purification of DctABs. (A) Protein samples from different steps throughout the purification were run on an SDS-12.5% polyacrylamide gel, which was stained with Coomassie brilliant blue. Lanes: 1, membrane vesicles (10 μg of protein); 2, DDM-soluble fraction; 3, insoluble fraction; 4, flowthrough from the nickel-Sepharose column; 5, wash fraction from the Ni-Sepharose column; 6 to 8, three elution fractions from the nickel-Sepharose column; and 9, peak fraction (at an elution volume of ∼11 ml) from the size exclusion chromatography step. The arrow indicates DctABs. M, markers; Mw, molecular mass. (B) The peak elution fraction from the nickel-Sepharose column (lane 7) was subjected to size exclusion chromatography with a Superdex-200 column. The chromatogram is shown.
The DctABs protein from the peak fraction around the elution volume of 11 ml was used in reconstitution trials. In addition, DctABs that had been partially purified by only metal affinity chromatography was also used in reconstitution trials. All attempts to functionally reconstitute the purified protein into proteoliposomes failed. Different detergents (maltoside series and octyl glucoside) and reconstitution protocols (detergent removal by rapid dilution [14] and BioBead adsorption [8]) were tested, but none resulted in functional protein.
DctABs is a proton-coupled succinate transporter.
It was possible that the failure to reconstitute DctABs in an active form had been caused by instability of the protein in detergent solution. To avoid the need for solubilization of DctABs by detergents, in vitro transport assays using membrane vesicles prepared from L. lactis cells overexpressing DctABs were attempted. We monitored the uptake of radiolabeled succinate into membrane vesicles in the presence of chemical gradients for sodium ions and protons and a negative membrane potential (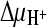 ,
, 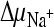 , and ΔΨ, respectively) (Fig. 2). Rapid succinate transport in membrane vesicles containing DctABs was observed. Transport was DctA mediated, because no accumulation of succinate was observed in membrane vesicles in which the unrelated thiamine transporter ThiT from L. lactis was overexpressed (6). To establish which ion gradients were required for transport, we varied the composition of the external buffers. Omission of the sodium ion gradient (
, and ΔΨ, respectively) (Fig. 2). Rapid succinate transport in membrane vesicles containing DctABs was observed. Transport was DctA mediated, because no accumulation of succinate was observed in membrane vesicles in which the unrelated thiamine transporter ThiT from L. lactis was overexpressed (6). To establish which ion gradients were required for transport, we varied the composition of the external buffers. Omission of the sodium ion gradient (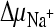 ) did not affect transport rates, but when the pHs on the two sides of the membrane were equal, transport rates and accumulation levels were strongly reduced, regardless of the presence of a gradient of sodium ions. An internal negative membrane potential stimulated succinate transport rates and accumulation levels both with and without a pH gradient, indicating that transport by DctABs was electrogenic, with a net positive charge moving in the same direction as the dicarboxylate substrate.
) did not affect transport rates, but when the pHs on the two sides of the membrane were equal, transport rates and accumulation levels were strongly reduced, regardless of the presence of a gradient of sodium ions. An internal negative membrane potential stimulated succinate transport rates and accumulation levels both with and without a pH gradient, indicating that transport by DctABs was electrogenic, with a net positive charge moving in the same direction as the dicarboxylate substrate.
FIG. 2.

Succinate transport by DctABs in membrane vesicles. (A) Membrane vesicles containing DctABs (circles) or ThiT (negative control; triangles) were loaded with 100 mM K-HEPES (pH 7.5). The vesicles were diluted 100-fold in an isosmotic Na-MES buffer, pH 5.5 (consisting of ∼135 mM MES [morpholineethanesulfonic acid] adjusted to pH 5.5 with NaOH), containing 3.1 μM [14C]succinate in the presence (closed symbols) or absence (open symbols) of 0.5 μM valinomycin. In this way, chemical gradients for protons and sodium ions (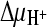 and
and 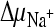 ) were created. In the presence of valinomycin, an additional K+ diffusion potential, ΔΨ (Nernst potential, −118 mV), was created. (B) Initial [14C]succinate transport rates in the presence of various gradients. Columns correspond to the presence of gradients as follows: 1,
) were created. In the presence of valinomycin, an additional K+ diffusion potential, ΔΨ (Nernst potential, −118 mV), was created. (B) Initial [14C]succinate transport rates in the presence of various gradients. Columns correspond to the presence of gradients as follows: 1, 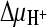 ,
, 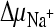 , and ΔΨ; 2,
, and ΔΨ; 2, 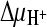 and ΔΨ; 3,
and ΔΨ; 3, 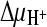 and
and 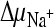 ; 4,
; 4, 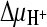 ; 5,
; 5, 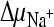 and ΔΨ; and 6, negative control. Initial transport rates were calculated from data for the 15-s time point (see panel A). Data were normalized, and the highest measured rate [47.4 pmol (mg protein·s)−1] was set at 100%. Proton and sodium ion gradients (
and ΔΨ; and 6, negative control. Initial transport rates were calculated from data for the 15-s time point (see panel A). Data were normalized, and the highest measured rate [47.4 pmol (mg protein·s)−1] was set at 100%. Proton and sodium ion gradients (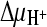 and
and 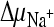 ) in the presence or absence of ΔΨ were created as described in the legend to panel A. To create
) in the presence or absence of ΔΨ were created as described in the legend to panel A. To create 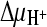 in the absence of
in the absence of 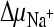 , external methylglucamine-MES buffer (pH 5.5) was used instead of Na-MES buffer. Again, the addition of valinomycin was used to create a K+ diffusion membrane potential (ΔΨ) where indicated. To create
, external methylglucamine-MES buffer (pH 5.5) was used instead of Na-MES buffer. Again, the addition of valinomycin was used to create a K+ diffusion membrane potential (ΔΨ) where indicated. To create 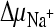 and ΔΨ in the absence of
and ΔΨ in the absence of 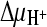 , Na-HEPES (pH 7.5) was used as the external buffer and valinomycin was present. ThiT-harboring vesicles were used as a control in the presence of all three gradients. All data are averages of three independent measurements; error bars indicate standard deviations.
, Na-HEPES (pH 7.5) was used as the external buffer and valinomycin was present. ThiT-harboring vesicles were used as a control in the presence of all three gradients. All data are averages of three independent measurements; error bars indicate standard deviations.
Succinate transport by DctABs is inhibited by other C4-dicarboxylates.
To determine the substrate specificity of DctABs, we tested whether the addition of a 50-fold excess of unlabeled putative substrates inhibited transport of radiolabeled succinate (Fig. 3). Only the addition of the C4-dicarboxylates succinate, fumarate, malate, and oxaloacetate strongly inhibited uptake of [14C]succinate into the membrane vesicles. The addition of maleate and various C3- and C5-dicarboxylates did not inhibit transport. None of the natural amino acids were able to inhibit succinate transport. Also, the addition of cysteate, gamma-aminobutyric acid (GABA), orotate, glyoxylate, and α-ketoglutarate had no effect on succinate uptake. It must be noted that the results do not exclude the possibility that the noninhibitory compounds may be low-affinity substrates, because only 170 μM concentrations were used in the inhibition assay. However, as discussed below, a counterflow assay showed that it is very unlikely that the compounds are (low-affinity) substrates of DctABs.
FIG. 3.

Succinate transport by DctABs in membrane vesicles in the presence of putative transport inhibitors. Transport of [14C]succinate (3.1 μM) in the presence of 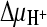 ,
, 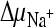 , and ΔΨ (−118 mV) was measured as described in the legend to Fig. 2A. The external buffer was supplemented with a 50-fold excess of the indicated compounds. Initial transport rates were determined as described in the legend to Fig. 2B and normalized with respect to the rate without inhibitors [100%, corresponding to 37.5 pmol (mg protein·s)−1]. Measurements were performed in quadruplicate, and standard deviations are shown.
, and ΔΨ (−118 mV) was measured as described in the legend to Fig. 2A. The external buffer was supplemented with a 50-fold excess of the indicated compounds. Initial transport rates were determined as described in the legend to Fig. 2B and normalized with respect to the rate without inhibitors [100%, corresponding to 37.5 pmol (mg protein·s)−1]. Measurements were performed in quadruplicate, and standard deviations are shown.
Fumarate, oxaloacetate, and malate are substrates for DctABs.
The results of the inhibition assay (Fig. 3) demonstrated that DctABs might transport fumarate, malate, and oxaloacetate. On the other hand, these compounds could also be competitive inhibitors that bind to the transporter without being transported themselves and, thus, block transport of radiolabeled succinate. To establish whether fumarate, oxaloacetate, and malate were indeed transported by DctABs, we used a counterflow assay. Membrane vesicles were loaded with 1 mM unlabeled (putative) substrate and diluted in the same buffer containing radiolabeled succinate. If the unlabeled dicarboxylate was indeed transported, exchange of the internal (unlabeled) substrate and external (labeled) succinate would lead to accumulation of [14C]succinate, due to the concentration gradient across the membrane. In the counterflow experiments, internalization of the radiolabeled substrate was observed when membrane vesicles were loaded with succinate (demonstrating homologous counterflow) (Fig. 4). Also, the presence of malate, fumarate, or oxaloacetate in the lumina of the membrane vesicles resulted in uptake of radiolabeled succinate into the vesicles. The presence of orotate, citrate, and maleate in the lumina of the vesicles did not result in the induction of succinate counterflow by DctABs, nor did the presence of aspartate, glyoxylate, or α-ketoglutarate (data not shown). We also did not observe succinate counterflow when no substrate was added to the internal buffer. Control membranes in which ThiT was expressed instead of DctABs also did not show counterflow of the radiolabeled substrate, regardless of the presence of the internal substrate. DctABs thus appears to be specific for a narrow set of C4-dicarboxylates. The counterflow assay also showed that aspartate, glyoxylate, α-ketoglutarate, orotate, citrate, and maleate are not low-affinity substrates of DctABs because they were not transported even though they were present at high (1 mM) concentrations.
FIG. 4.
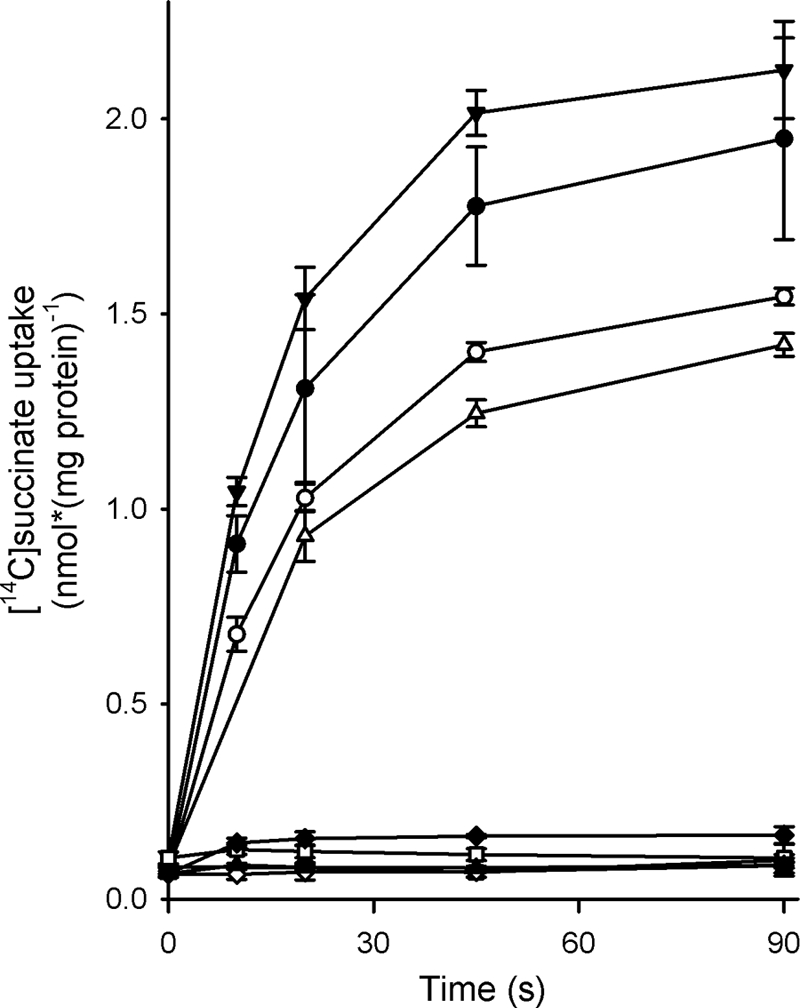
Succinate counterflow by DctABs. Membrane vesicles were loaded with 50 mM potassium phosphate (pH 7.0) supplemented with 1 mM succinate (closed circles), fumarate (open circles), malate (closed triangles), oxaloacetate (open triangles), maleate (open squares), citrate (closed squares), or orotate (closed diamonds) and diluted 200-fold in external buffer (50 mM potassium phosphate, pH 7.0) supplemented with 3.1 μM [14C]succinate. As a negative control, membrane vesicles that were not loaded with any substrate (open diamonds) or vesicles loaded with 1 mM succinate (crosses) in which ThiT was expressed were used. Measurements were done in triplicate; error bars indicate standard deviations.
pH dependence of DctA.
The experimental findings presented in Fig. 2 indicate that DctABs couples succinate transport to protons and not to sodium ions. This observation corresponds with results obtained for DctA of C. glutamicum in a whole-cell experimental setup (41). To gain more insight into both the optimal pH for transport by DctA and the protonation state of the transported dicarboxylate, we measured the pH dependence of succinate transport into membrane vesicles. Interpretation of the pH dependence of succinate transport by DctABs is complicated because (i) protons are substrates (cotransported ions), (ii) the protonation state of the dicarboxylate substrates depends on the pH, and (iii) there may be other (regulatory) sites on the protein which can be protonated and the protonation state of which may affect activity. We performed three experiments to study the pH dependence of DctABs by using both net uptake and counterflow assays.
In the first experiment, we varied the external pH (5.5 or 7) while keeping the internal pH (8) constant. We determined the apparent Km and the Vmax for succinate transport and found that the transporter follows Michaelis-Menten kinetics, which has also been shown previously for other members of the glutamate transporter family (31, 35). The Vmax for succinate transport was approximately 3-fold lower at an external pH of 7 [1.4 nmol (mg protein)−1 s−1] than at pH 5.5 [3.9 nmol (mg protein)−1 s−1]. This result is consistent with the protons being cosubstrates. Although protonation of a pH-sensitive regulatory site at the extracellular face of the protein could also explain the results, we will show below that such regulation is less likely. The apparent Km values for succinate transport were identical within the range of error (means ± standard deviations, 2.8 ± 1.0 μM at pH 5.5 and 2.6 ± 1.4 μM at pH 7; n = 3). The pKa values for the two carboxylate groups in succinate are 4.2 and 5.6, respectively. At pH 5.5, ∼60% of succinate is in a protonated state (single or double), whereas at pH 7 this percentage is only ∼4%. The 15-fold decrease in the concentration of the protonated substrate was not reflected in the apparent Km values, indicating that the doubly deprotonated form of succinate must be recognized by the transporter. Similar interpretations of apparent Km values at different pHs have been made to determine the protonation state of the substrate of a glutamate transporter in L. lactis (26). The concentration of divalent anionic succinate is ∼2-fold higher at pH 7 than at pH 5.5. The increase was also not reflected in the apparent Km values. Although this result could indicate that both protonated and unprotonated forms of succinate were recognized by DctABs, the experimental error in the determination of the Km values was too large to accurately measure a 2-fold difference.
In the second experiment, we varied the external pH from 5 to 7 and simultaneously varied the internal pH from 6 to 8, so that a pH gradient of 1 unit was maintained. Succinate transport rates increased by ∼3-fold when the internal pH was raised from 6 to 8 (Fig. 5). The transport rates increased despite a reduction in the concentration of the cosubstrate (protons) on the outside, which as described above, was shown in the previous experiment to decrease the transport rate when the internal pH was constant. It is therefore likely that the release of protons on the inside was rate limiting, with the stimulating effect of a higher internal pH being dominant over the inhibitory effect of a higher external pH. An alternative explanation would be that there is a pH-sensitive regulatory site at the intracellular face of the protein resulting in higher activity at alkaline pHs. However, this interpretation would imply that the pH regulation of the protein on the inside and that on the outside work in opposite directions, which is unlikely. The more likely explanation for the observations is that the measured pH dependence directly indicates proton binding on the outside and release on the inside.
FIG. 5.

pH dependence of succinate transport. Membrane vesicles were loaded with a mixture of 50 mM K-MES and 50 mM K-HEPES adjusted to a pH of 6, 7, or 8, and the loaded vesicles were diluted in isosmotic Na-MES-Na-HEPES buffer adjusted to a pH of 5, 6, or 7. The external buffer contained 3.1 μM [14C]succinate and 0.5 μM valinomycin.
In the third experiment, the pH dependence of succinate transport by DctABs during counterflow with identical pHs on the two sides of the membrane was measured. Maximal succinate counterflow activity was observed at pHs of 6.5 to 7.0 (Fig. 6). At lower pHs and at a pH of 7.5, the rate of exchange was significantly lower. These results suggest that in addition to the substrate, protons are exchanged during the counterflow assay. The observed pH optimum for counterflow activity may thus be a reflection of the stimulation of transport rates by a compromise between (i) high proton concentrations on the outside and (ii) low concentrations on the inside during the counterflow reaction. These results corroborate the interpretation of the findings from the succinate uptake experiments described above, from which we concluded that the measured pH dependence of succinate transport directly represents proton binding on the outside and release on the inside. Exchange of a coupling ion during counterflow has also been observed previously, e.g., with the neutral amino acid transporter ASCT-1 (44) of the DAACS family. In this case, a sodium ion was exchanged.
FIG. 6.

pH dependence of succinate transport during counterflow. Membrane vesicles were loaded with 50 mM potassium phosphate, supplemented with 1 mM unlabeled succinate, at the indicated pHs. Membrane vesicles were diluted 100-fold in the same external buffer but supplemented with 3.1 μM [14C]succinate instead of the unlabeled succinate. Error bars indicate the standard deviations, taken from three measurements.
A negative membrane potential stimulates succinate transport by DctA.
Succinate transport by DctABs is electrogenic, as depicted in Fig. 2. To further analyze the effect of the membrane potential on succinate transport by DctABs, the magnitude and sign of the membrane potential were varied. Membrane vesicles were loaded with a buffer containing 10 mM potassium ions. By varying the external concentration of potassium ions, potassium diffusion potentials of different magnitudes were established in the presence of the potassium-selective ionophore valinomycin. In the absence of the ionophore valinomycin, transport was driven solely by the proton gradient and transport rates and accumulation levels were the same regardless of the external potassium ion concentration (data not shown), indicating that the potassium ion concentration does not affect transport. Lack of potassium ion dependence has also been observed previously for other prokaryotic transporters of the DAACS family. In contrast, eukaryotic glutamate transporters require potassium ions as countertransported ions (24, 28, 31, 45).
The addition of valinomycin resulted in an increase of the initial transport rates when the external concentration of potassium was lower than the internal concentration and negative K+ diffusion potentials were generated (Fig. 7). When K+ concentrations on the two sides of the membrane were equal, the addition of valinomycin also resulted in an increase in transport rates. This result indicates that the net positive charge was translocated inward across the membrane. In the absence of valinomycin, the translocated charge led to the buildup of a positive membrane potential, thus impeding further transport. By clamping the membrane potential at voltages lower than the (positive) equilibrium potential for succinate-proton symport, such impediment was alleviated.
FIG. 7.

Succinate transport in the presence of membrane potentials of different magnitudes. Membrane vesicles were loaded with 100 mM Na-HEPES buffer (pH 7.5) supplemented with 10 mM KCl. Membrane vesicles were diluted in external buffer consisting of Na-MES, pH 5.5, and 3.1 μM [14C]succinate supplemented with 0 mM KCl (closed circles), 2 mM KCl (open circles), 10 mM KCl (closed triangles), or 50 mM KCl (open triangles) in the presence of 0.5 μM valinomycin. The concentration of Na-MES was adjusted to ensure that the external and internal solutions were isosmotic. As a control, succinate transport in membrane vesicles in which ThiT was expressed was measured with 0 mM KCl in the external buffer (closed squares). The values of the K+ equilibrium potentials are indicated.
Temperature optimum and Q10 value.
We measured initial rates of [14C]succinate transport by DctABs at temperatures between 10 and 45°C (data not shown). The rates increased when the temperature was raised from 10 up to 40°C. The Q10 value was approximately 1.6 for temperatures between 20 and 40°C, suggesting that significant conformational changes take place during transport. This finding is consistent with results obtained previously for GltPh (Q10 of 3.7) (31) and various other bacterial glutamate transporters (Q10s of ∼2.2) (M. Groeneveld, unpublished data), although the Q10 value for DctABs is lower than those for the aforementioned proteins.
DISCUSSION
Prokaryotic transporters of the DAACS family can be subdivided into three separate classes based on substrate specificity (33): glutamate/aspartate transporters, neutral amino acid transporters, and transporters for C4-dicarboxylates of the Krebs cycle. Glutamate/aspartate transporters and the serine/threonine transporter SstT have been studied in vitro using membrane vesicles and purified proteins reconstituted into liposomes, but DctA-type dicarboxylate transporters have been studied in whole-cell assays only.
In this report, we describe the characterization of DctABs in membrane vesicles from L. lactis in which DctABs was overproduced. We observed DctABs-specific succinate transport in these membranes, and a series of inhibition and counterflow assays showed that fumarate, malate, and oxaloacetate were also transported by DctABs. The substrate specificity of DctABs is narrower than that of other DctA homologues, which have been reported to transport other compounds such as orotate, glyoxylate, and α-ketoglutarate (41, 42). None of these substrates were transported by DctABs.
We made a homology model of the substrate-binding site of DctABs by using the crystal structure of GltPh as a template (data not shown). When the amino acids in GltPh involved in substrate binding (2) were compared with the corresponding amino acids in DctABs, only a few differences were found. The most obvious difference is the replacement of Asp394 from GltPh by Ser377 in DctABs. In GltPh, Asp394 interacts with the α-amino group of the substrate aspartate, which is not present in the dicarboxylate substrates of DctABs. The binding pocket in GltPh for the two carboxylates of aspartate is strongly conserved in DctABs. The pH dependence of succinate transport by DctABs suggested that the doubly deprotonated (divalent anionic) form of succinate was recognized by the transporter, which would be consistent with the presence of a conserved binding site for the two carboxylate groups, because the divalent anionic form of aspartate is also transported by GltPh (2, 31).
Substrate transport by DctABs was coupled to proton transport (symport) and was not dependent on sodium ions. Succinate transport was stimulated by a negative membrane potential, indicating that transport was electrogenic. Therefore, at least three protons must be transported with the divalent anion succinate. This pattern is in contrast with succinate transport by the Staphylococcus aureus protein SdcS, a sodium-coupled C4-dicarboxylate transporter of the divalent anion:Na+ symporter (DASS) family, which has been characterized using proteoliposomes containing purified protein. SdcS transports a doubly deprotonated substrate together with two sodium ions, resulting in electroneutral transport (13). On the other hand, transport by eukaryotic homologues of SdcS (members of the SlC13 subfamily) is electrogenic; three sodium ions are cotransported with a divalent substrate (25).
Glutamate/aspartate transporters of the DAACS family use either protons (in the case of GltP from E. coli) or sodium ions (in the case of GltPh) as coupling ions. Sodium ion binding by the aspartate transporter GltPh, for which a crystal structure has been determined, has been studied in detail. By using thallium ions to replace sodium in the crystals of GltPh, Boudker et al. found two distinct sodium-binding sites in the protein, in close proximity to the aspartate-binding site (2). We compared the sodium ion-binding sites in the crystal structure of GltPh with sites in the homology model of DctABs and found surprisingly few differences. In the first site, the sodium ion is coordinated by three backbone carbonyl oxygen atoms and a carboxylate from Asp405 in GltPh. This aspartate residue is conserved among human homologues (40), and another acidic residue (glutamate) is present in the binding site of the sodium-coupled serine/threonine transporter SstT from E. coli. Proton-coupled transporters such as DctA or bacterial glutamate transporters such as GltT from B. stearothermophilus (40) lack the aspartate residue in the first binding site and contain an asparagine residue instead. In the second predicted sodium-binding site, the sodium ion is coordinated by backbone carbonyls only (33, 40). The homology model of DctABs does not allow us to pinpoint the locations of the three proton-binding sites, but based on tertiary-structure conservation, it seems likely that the proton-binding sites in DctA are at positions similar to those of the predicted sodium-binding sites in GltPh (2). The third proton may bind in an additional ion-binding site, as predicted by Holley and Kavanaugh (15).
Acknowledgments
We thank Ruben Vis for his initial experiments with E. coli DctA and Guus Erkens for his kind gift of L. lactis cells harboring a ThiT expression vector.
This work was supported by a vidi grant from the Netherlands Organization for Scientific Research (NWO) to D.J.S.
Footnotes
Published ahead of print on 2 April 2010.
REFERENCES
- 1.Asai, K., S. H. Baik, Y. Kasahara, S. Moriya, and N. Ogasawara. 2000. Regulation of the transport system for C4-dicarboxylic acids in Bacillus subtilis. Microbiology 146(Pt. 2):263-271. [DOI] [PubMed] [Google Scholar]
- 2.Boudker, O., R. M. Ryan, D. Yernool, K. Shimamoto, and E. Gouaux. 2007. Coupling substrate and ion binding to extracellular gate of a sodium-dependent aspartate transporter. Nature 445:387-393. [DOI] [PubMed] [Google Scholar]
- 3.Crisman, T., S. Qu, B. Kanner, and L. Forrest. 2009. Inward-facing conformation of glutamate transporters as revealed by their inverted-topology structural repeats. Proc. Natl. Acad. Sci. U. S. A. 106:20752-20757. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Danbolt, N. C. 2001. Glutamate uptake. Prog. Neurobiol. 65:1-105. [DOI] [PubMed] [Google Scholar]
- 5.Davies, S. J., P. Golby, D. Omrani, S. A. Broad, V. L. Harrington, J. R. Guest, D. J. Kelly, and S. C. Andrews. 1999. Inactivation and regulation of the aerobic C4-dicarboxylate transport (dctA) gene of Escherichia coli. J. Bacteriol. 181:5624-5635. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Eudes, A., G. B. Erkens, D. J. Slotboom, D. A. Rodionov, V. Naponelli, and A. D. Hanson. 2008. Identification of genes encoding the folate- and thiamine-binding membrane proteins in Firmicutes. J. Bacteriol. 190:7591-7594. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Gaillard, I., D. J. Slotboom, J. Knol, J. S. Lolkema, and W. N. Konings. 1996. Purification and reconstitution of the glutamate carrier GltT of the thermophilic bacterium Bacillus stearothermophilus. Biochemistry 35:6150-6156. [DOI] [PubMed] [Google Scholar]
- 8.Geertsma, E. R., N. A. B. Nik Mahmood, G. K. Schuurman-Wolters, and B. Poolman. 2008. Membrane reconstitution of ABC transporters and assays of translocator function. Nat. Protoc. 3:256-266. [DOI] [PubMed] [Google Scholar]
- 9.Geertsma, E. R., and B. Poolman. 2007. High-throughput cloning and expression in recalcitrant bacteria. Nat. Methods 4:705-707. [DOI] [PubMed] [Google Scholar]
- 10.Gendreau, S., S. Voswinkel, D. Torres-Salazar, N. Lang, H. Heidtmann, S. Detro-Dassen, G. Schmalzing, P. Hidalgo, and C. Fahlke. 2004. A trimeric quaternary structure is conserved in bacterial and human glutamate transporters. J. Biol. Chem. 279:39505-39512. [DOI] [PubMed] [Google Scholar]
- 11.Grewer, C., P. Balani, C. Weidenfeller, T. Bartusel, Z. Tao, and T. Rauen. 2005. Individual subunits of the glutamate transporter EAAC1 homotrimer function independently of each other. Biochemistry 44:11913-11923. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Groeneveld, M., and D.-J. Slotboom. 2007. Rigidity of the subunit interfaces of the trimeric glutamate transporter GltT during translocation. J. Mol. Biol. 372:565-570. [DOI] [PubMed] [Google Scholar]
- 13.Hall, J. A., and A. M. Pajor. 2007. Functional reconstitution of SdcS, a Na+-coupled dicarboxylate carrier protein from Staphylococcus aureus. J. Bacteriol. 189:880-885. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Heuberger, E. H., E. Smits, and B. Poolman. 2001. Xyloside transport by XylP, a member of the galactoside-pentoside-hexuronide family. J. Biol. Chem. 276:34465-34472. [DOI] [PubMed] [Google Scholar]
- 15.Holley, D. C., and M. P. Kavanaugh. 2009. Interactions of alkali cations with glutamate transporters. Philos. Trans. R. Soc. Lond. B Biol. Sci. 364:155-161. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Janausch, I. G., E. Zientz, Q. H. Tran, A. Kroger, and G. Unden. 2002. C4-dicarboxylate carriers and sensors in bacteria. Biochim. Biophys. Acta 1553:39-56. [DOI] [PubMed] [Google Scholar]
- 17.Kim, Y.-M., W. Ogawa, E. Tamai, T. Kuroda, T. Mizushima, and T. Tsuchiya. 2002. Purification, reconstitution, and characterization of Na+/serine symporter, SstT, of Escherichia coli. J. Biochem. 132:71-76. [DOI] [PubMed] [Google Scholar]
- 18.Koch, H. P., R. L. Brown, and H. P. Larsson. 2007. The glutamate-activated anion conductance in excitatory amino acid transporters is gated independently by the individual subunits. J. Neurosci. 27:2943-2947. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Koch, H. P., and H. P. Larsson. 2005. Small-scale molecular motions accomplish glutamate uptake in human glutamate transporters. J. Neurosci. 25:1730-1736. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Kuipers, O. P., M. M. Beerthuyzen, R. J. Siezen, and W. M. de Vos. 1993. Characterization of the nisin gene cluster nisABTCIPR of Lactococcus lactis. Requirement of expression of the nisA and nisI genes for development of immunity. Eur. J. Biochem. 216:281-291. [DOI] [PubMed] [Google Scholar]
- 21.Kuipers, O. P., P. G. de Ruyter, M. Kleerebezem, and W. M. de Vos. 1998. Quorum sensing controlled gene expression in lactic acid bacteria. J. Biotechnol. 64:15-21. [Google Scholar]
- 22.Kunji, E. R. S., D.-J. Slotboom, and B. Poolman. 2003. Lactococcus lactis as host for overproduction of functional membrane proteins. Biochim. Biophys. Acta 1610:97-108. [DOI] [PubMed] [Google Scholar]
- 23.Leary, G. P., E. F. Stone, D. C. Holley, and M. P. Kavanaugh. 2007. The glutamate and chloride permeation pathways are colocalized in individual neuronal glutamate transporter subunits. J. Neurosci. 27:2938-2942. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Levy, L. M., O. Warr, and D. Attwell. 1998. Stoichiometry of the glial glutamate transporter GLT-1 expressed inducibly in a Chinese hamster ovary cell line selected for low endogenous Na+-dependent glutamate uptake. J. Neurosci. 18:9620-9628. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Markovich, D., and H. Murer. 2004. The SLC13 gene family of sodium sulphate/carboxylate cotransporters. Pflugers Arch. 447:594-602. [DOI] [PubMed] [Google Scholar]
- 26.Poolman, B., E. J. Smid, and W. N. Konings. 1987. Kinetic properties of a phosphate-bond-driven glutamate-glutamine transport system in Streptococcus lactis and Streptococcus cremoris. J. Bacteriol. 169:2755-2761. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Prakash, S., G. Cooper, S. Singhi, and M. H. J. Saier. 2003. The ion transporter superfamily. Biochim. Biophys. Acta 1618:79-92. [DOI] [PubMed] [Google Scholar]
- 28.Raunser, S., M. Appel, C. Ganea, U. Geldmacher-Kaufer, K. Fendler, and W. Kuhlbrandt. 2006. Structure and function of prokaryotic glutamate transporters from Escherichia coli and Pyrococcus horikoshii. Biochemistry 45:12796-12805. [DOI] [PubMed] [Google Scholar]
- 29.Reyes, N., C. Ginter, and O. Boudker. 2009. Transport mechanism of a bacterial homologue of glutamate transporters. Nature 462:880-885. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Ronson, C. W., P. M. Astwood, and J. A. Downie. 1984. Molecular cloning and genetic organization of C4-dicarboxylate transport genes from Rhizobium leguminosarum. J. Bacteriol. 160:903-909. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Ryan, R. M., E. L. R. Compton, and J. A. Mindell. 2009. Functional characterization of a Na+-dependent aspartate transporter from Pyrococcus horikoshii. J. Biol. Chem. 284:17540-17548. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Saier, M. H. J., C. V. Tran, and R. D. Barabote. 2006. TCDB: the Transporter Classification Database for membrane transport protein analyses and information. Nucleic Acids Res. 34:D181-D186. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Slotboom, D., W. Konings, and J. Lolkema. 1999. Structural features of the glutamate transporter family. Microbiol. Mol. Biol. Rev. 63:293-307. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Tolner, B., T. Ubbink-Kok, B. Poolman, and W. N. Konings. 1995. Cation-selectivity of the L-glutamate transporters of Escherichia coli, Bacillus stearothermophilus and Bacillus caldotenax: dependence on the environment in which the proteins are expressed. Mol. Microbiol. 18:123-133. [DOI] [PubMed] [Google Scholar]
- 35.Tolner, B., T. Ubbink-Kok, B. Poolman, and W. N. Konings. 1995. Characterization of the proton/glutamate symport protein of Bacillus subtilis and its functional expression in Escherichia coli. J. Bacteriol. 177:2863-2869. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Utsunomiya-Tate, N., H. Endou, and Y. Kanai. 1996. Cloning and functional characterization of a system ASC-like Na+-dependent neutral amino acid transporter. J. Biol. Chem. 271:14883-14890. [DOI] [PubMed] [Google Scholar]
- 37.van Slooten, J. C., T. V. Bhuvanasvari, S. Bardin, and J. Stanley. 1992. Two C4-dicarboxylate transport systems in Rhizobium sp. NGR234: rhizobial dicarboxylate transport is essential for nitrogen fixation in tropical legume symbioses. Mol. Plant Microbe Interact. 5:179-186. [DOI] [PubMed] [Google Scholar]
- 38.Yarosh, O. K., T. C. Charles, and T. M. Finan. 1989. Analysis of C4-dicarboxylate transport genes in Rhizobium meliloti. Mol. Microbiol. 3:813-823. [DOI] [PubMed] [Google Scholar]
- 39.Yernool, D., O. Boudker, E. Folta-Stogniew, and E. Gouaux. 2003. Trimeric subunit stoichiometry of the glutamate transporters from Bacillus caldotenax and Bacillus stearothermophilus. Biochemistry 42:12981-12988. [DOI] [PubMed] [Google Scholar]
- 40.Yernool, D., O. Boudker, Y. Jin, and E. Gouaux. 2004. Structure of a glutamate transporter homologue from Pyrococcus horikoshii. Nature 431:811-818. [DOI] [PubMed] [Google Scholar]
- 41.Youn, J.-W., E. Jolkver, R. Kramer, K. Marin, and V. F. Wendisch. 2009. Characterization of the dicarboxylate transporter DctA in Corynebacterium glutamicum. J. Bacteriol. 191:5480-5488. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Yurgel, S., M. W. Mortimer, K. N. Rogers, and M. L. Kahn. 2000. New substrates for the dicarboxylate transport system of Sinorhizobium meliloti. J. Bacteriol. 182:4216-4221. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Yurgel, S. N., and M. L. Kahn. 2004. Dicarboxylate transport by rhizobia. FEMS Microbiol. Rev. 28:489-501. [DOI] [PubMed] [Google Scholar]
- 44.Zerangue, N., and M. P. Kavanaugh. 1996. ASCT-1 is a neutral amino acid exchanger with chloride channel activity. J. Biol. Chem. 271:27991-27994. [DOI] [PubMed] [Google Scholar]
- 45.Zerangue, N., and M. P. Kavanaugh. 1996. Flux coupling in a neuronal glutamate transporter. Nature 383:634-637. [DOI] [PubMed] [Google Scholar]