

Abstract
Four major malaria-causing Plasmodium spp. and lymphatic filariasis-causing Wuchereria bancrofti are co-endemic in many tropical and sub-tropical regions. Among molecular diagnostic assays, multiplex polymerase chain reaction (PCR)–based assays for the simultaneous detection of DNAs from these parasite species are currently available only for P. falciparum and W. bancrofti or P. vivax and W. bancrofti. Using a post-PCR oligonucleotide ligation detection reaction–fluorescent microsphere assay (LDR-FMA), we developed a multiplex assay that has the capability to simultaneously detect all four Plasmodium spp. and W. bancrofti infections in blood samples. Compared with microfilarial positivity in the blood, the LDR-FMA assay is highly concordant (91%), sensitive (86%), and specific (94%), and has high reproducibility for Plasmodium spp. (85–93%) and W. bancrofti (90%) diagnoses. The development of this assay for the simultaneous diagnosis of multiple parasitic infections enables efficient screening of large numbers of human blood and mosquito samples from co-endemic areas.
Globally, malaria and lymphatic filariasis are the most threatening of the mosquito-transmitted parasitic diseases.1 Among the three parasites that cause lymphatic filariasis, Wuchereria bancrofti, Brugia malayi, and B. timori, W. bancrofti is the most widely distributed and is responsible for 90% of lymphatic filariasis infections (bancroftian filariasis) worldwide.2 Malaria and bancroftian filariasis are co-endemic in many tropical and sub-tropical regions, such as Southeast Asia, including the western Pacific, Africa, and Central and South America, and are transmitted by a number of common vector species.3,4 Thus, co-infections with malaria and bancroftian parasites in humans5–7 and mosquitoes7,8 are found in these regions.
Because of their significant impact on public health, global campaigns with a variety of approaches have been launched for the control/elimination of these diseases.9,10 These approaches range from the treatment of clinical patients to the control of disease transmission by preventative chemotherapy and vector control.9,10 However, challenges lie ahead for the success of these control/elimination programs without thoughtful and appropriate use of highly sensitive and specific diagnostic methods.
Parasitologic diagnosis of malaria and bancroftian filariasis is typically made by microscopic examination of stained blood smears or membrane filtrates.5–7 In addition to microscopic detection of microfilariae, detection of circulating filarial antigen(s) by enzyme-linked immunosorbent assay and immunochromatographic test are other commonly used methods to diagnose bancroftian filariasis.11–13 A number of polymerase chain reaction (PCR)–based assays are available to separately detect malaria14,15 and bancroftian filariasis16–19 parasites. However, only two assays are available to detect these parasite species simultaneously: a multiplex PCR assay for detection of Plasmodium falciparum and W. bancrofti in humans,20 and a real-time multiplex quantitative PCR assay for detection of P. falciparum and W. bancrofti, or P. vivax and W. bancrofti in mosquitoes.21
Malaria is endemic at altitudes below 1,300–1,600 meters in Papua New Guinea and is the leading cause of illness and death in this country.22 Four major parasite species, P. falciparum, P. vivax, P. malariae, and P. ovale, are transmitted in Papua New Guinea and mixed-species infections are common.23,24 Recently, we developed a 96-well format, post-PCR ligation detection reaction–fluorescent microsphere assay (LDR-FMA) for multiplex detection of the four major Plasmodium spp,25 and validated its utility in diverse epidemiologic settings.26,27 Bancroftian filariasis is also endemic in several areas in Papua New Guinea and is a major cause of chronic and acute morbidity.28
We have been using the density of microfilariae in blood and an enzyme-linked immunosorbent assay (detection of Og4C3 antigen and anti-Bm14 IgG4) as measures of W. bancrofti infection in our ongoing lymphatic filariasis–related epidemiologic studies.29–31 However, with decreasing prevalence of W. bancrofti infections, lower microfilaremia, and increasing importance of xenodiagnosis of infection in mosquitoes because of the anticipated success of filariasis elimination programs, DNA-based methods may be more efficient for performing the population-level diagnostic surveillance. Expanding our existing post-PCR LDR-FMA assay, we report the development of a multiplex assay that has the capability to simultaneously detect P. falciparum, P. vivax, P. malariae, and P. ovale, and W. bancrofti infections with high sensitivity and specificity in blood samples.
The study was performed according to protocols approved by Institutional Review Boards of University Hospitals Case Medical Center (Protocol 08-05-13) and the Papua New Guinea Institute of Medical Research (Protocol 07-16). Further approval was obtained from the Papua New Guinea Medical Research Advisory Committee (Protocol 6.09). Informed consent was obtained from all study participants at the time of enrollment.
This new assay involves a multiplex PCR to amplify genomic regions from Plasmodium spp. (small subunit ribosomal RNA gene fragment)25 and W. bancrofti (long DNA repeat region),18 followed by a multiplex LDR-FMA to detect P. falciparum, P. vivax, P. malariae, and P. ovale,25 and W. bancrofti in a sequence-specific manner. The PCR reagents and conditions for Plasmodium spp. amplification have been described.24,25
For the multiplex PCR, we evaluated the dNTP concentrations (dATP, dTTP, dGTP, and dCTP) from 200 μM to 800 μM to ensure nucleotide availability for the amplification of both Plasmodium spp. and W. bancrofti genomic regions, and added 0.12 μM of each of W. bancrofti UP (5¢-GATGGTGTATAATAGCAGCA-3¢) and W. bancrofti DN (5¢-GTCATTTATTTCTCCGTCGACTGTC-3¢) amplification primers to the PCR master mixture. The dNTP concentration that performed with consistently high efficiency was 400 μM. The PCR products were subjected to electrophoresis on agarose gels to visualize distinct Plasmodium spp. (491–500 basepairs)25 and W. bancrofti (174 basepairs) amplicons. The PCR products were then subjected to LDR-FMA as described,25 with minor modifications that included use of LDR primers: a W. bancrofti-specific primer (tacactttatcaaatcttacaatcTATATCTGCCCATAGAAATAACTA [sequence in lower case letters represents a 24-basepair oligonucleotide tag]) and a W. bancrofti common primer (Phos 5¢-CGGTGGATCTCTGGTTATCACTCTG-3¢Biotin). In the LDR-fluorescent microsphere hybridization solution containing Plasmodium species-specific fluorescent microspheres,25 we added W. bancrofti -specific fluorescent microsphere #3. Our W. bancrofti PCR and LDR primer sequences are based on the W. bancrofti sequence in GenBank (accession no. AY297458).18
To confirm the specificity of our W. bancrofti PCR primers, we amplified an approximately 170-basepair genomic DNA region from one microfilaria isolated from a person in Papua New Guinea and sequenced it. We also sequenced the PCR products from two microfilaria-positive persons from Papua New Guinea whose blood samples were collected as a part of ongoing studies.32 Furthermore, we sequenced the PCR product from five pooled third-stage larvae dissected from mosquitoes that were collected in the Dreikikir District (East Sepik Province, Papua New Guinea), where malaria and lymphatic filariasis are endemic.24,29–31 All sequences were 100% identical with the W. bancrofti sequence in GenBank (accession no. AY297458).
The specificity of the assay was further demonstrated by using P. falciparum, P. vivax, P. malariae, and P. ovale, and W. bancrofti genomic DNA samples (Figure 1). Using these genomic DNA samples individually in LDR-FMA reactions containing primers and microspheres for all five species, we found that the assay detected only the parasite species DNA present, and background signals for all other species DNAs were below a median fluorescence intensity of 500. We then performed multiplex PCR LDR-FMA diagnosis to detect Plasmodium spp. and W. bancrofti infections in the blood samples from 517 persons living in the Dreikikir District (East Sepik Province, Papua New Guinea).
Figure 1.
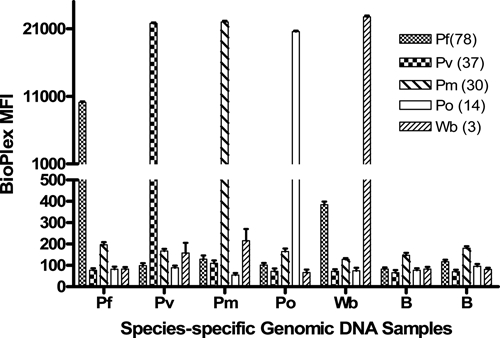
Detection of species-specific DNAs by the five-species multiplex polymerase chain reaction oligonucleotide ligation detection reaction–fluorescent microsphere assay (LDR-FMA). Data represent a summary of five positive-control experiments detecting Plasmodium falciparum, P. vivax, P. malariae, P. ovale, and Wuchereria bancrofti genomic DNAs. Whereas genomic DNAs were added individually, LDR-FMA reactions included oligonucleotide primers and microspheres representing all five species. Numbers in parentheses next to parasite-species designations in the legend ((78), (37), (30), (14), (3)) identify FlexMap™ microspheres (Luminex Corp., Austin, TX). B identifies two blank samples to which no genomic DNAs were added.
Using this assay, we found that 443 persons (86%) were infected with at least one of the parasites. Overall infection counts were P. falciparum 346 (67%), P. vivax 176 (34%), P. malariae 116 (22%), P. ovale 35 (7%), and W. bancrofti 175 (34%). Reproducibility of these results was tested for 174 samples (34%). In this analysis, we observed 85–93% concordance for Plasmodium spp. diagnosis and 90% concordance for W. bancrofti diagnosis. Thus, development of this 96-well format assay for simultaneous diagnosis of multiple parasitic infections enables efficient screening of large numbers of samples.
Finally, we categorized Plasmodium spp. and W. bancrofti infections into parasite assemblages, which are shown in Table 1. Overall infection prevalences of P. falciparum, P. vivax, P. malariae, and P. ovale and the results regarding single-species and mixed-species infections (Table 1) were consistent with our previous observations from the same area.24 Most W. bancrofti infections (150 of 175) were observed in various Plasmodium spp. assemblages containing the most prevalent P. falciparum infections (Table 1), which is similar to other findings.5–7
Table 1.
Prevalence (counts) of not infected and all infections detected by ligation detection reaction–fluorescent microsphere assay*
| Infection status | No. of persons |
|---|---|
| Not infected | 74 |
| Pf | 100 |
| Pv | 30 |
| Pm | 7 |
| Po | 2 |
| Wb | 25 |
| Pf, Pv | 51 |
| Pf, Pm | 27 |
| Pf, Po | 4 |
| Pv, Pm | 8 |
| Pv, Po | 1 |
| Pm, Po | 1 |
| Pf, Pv, Pm | 22 |
| Pf, Pv, Po | 5 |
| Pf, Pm, Po | 5 |
| Pv, Pm, Po | 2 |
| Pf, Pv, Pm, Po | 3 |
| Pf, Wb | 68 |
| Pv, Wb | 11 |
| Pm, Wb | 6 |
| Po, Wb | 2 |
| Pf, Pv, Wb | 22 |
| Pf, Pm, Wb | 16 |
| Pf, Po, Wb | 4 |
| Pv, Pm, Wb | 1 |
| Pv, Po, Wb | 1 |
| Pf, Pv, Pm, Wb | 14 |
| Pf, Pv, Po, Wb | 1 |
| Pf, Pv, Pm, Po, Wb | 4 |
Pf = Plasmodium falciparum; Pv = P. vivax; Pm = P. malariae; Po = P. ovale; Wb = Wuchereria bancrofti.
For diagnosis of Plasmodium spp. infections, we have observed that LDR-FMA is significantly more sensitive than light microscopy.25–27 In the present study, we compared microfilarial positivity in the blood, determined by microscopic examination of membrane filtrates,31 with the W. bancrofti LDR-FMA results (Table 2). We observed high concordance (91%) between the two methods, and high sensitivity (86%) and specificity (94%) of the LDR-FMA. When we further analyzed 26 false-negative results (microfilariae positive and LDR-FMA negative), we found that 69% (18 of 26) of them had low microfilaria counts (1–5 microfilariae/mL). Thus, false-negative results could be attributed to low parasite density, technical issues related to genomic DNA extraction and PCR/LDR-FMA, and/or W. bancrofti sequence polymorphism. Conversely, 19 false-positive results (microfilariae negative and LDR-FMA positive) could be attributed to misreadings during microscopy. Furthermore, false-negative and false-positive results may be partly attributed to non-uniform distribution of microfilariae in the blood.33
Table 2.
Comparison between microfilaria counts and Wuchereria bancrofti LDR-FMA results*
| LDR-FMA | Microfilariae | |
|---|---|---|
| Positive | Negative | |
| Positive | 156 | 19 |
| Negative | 26 | 316 |
Concordance between microfilariae detection by microscopy and ligation detection reaction–fluorescent microsphere assay (LDR-FMA) was calculated as (156 + 316)/517 = 91%. Sensitivity of the LDR-FMA assay was calculated as 156/(156 + 26) = 86%, and specificity of the assay was calculated as 316/(316 + 19) = 94%.
Wuchereria bancrofti is most commonly diagnosed when microfilariae or circulating parasite antigen(s) are detected in the blood.10 Antibody testing10 and parasite DNA detection in either human blood samples20 or mosquitoes21 could also be used for this purpose. All of these diagnostic tests have their own advantages and limitations, depending upon the lymphatic filariasis–related epidemiologic and clinical characteristics of the region.10 In the regions where W. bancrofti and multispecies Plasmodium infections are co-endemic, sensitive and specific molecular diagnosis of the full spectrum of these infections is highly desirable. Furthermore, the availability of accurate tools to monitor presence or absence of parasites in humans and mosquitoes is essential to better assess the effectiveness of intervention programs aimed at controlling or eliminating these diseases. In this direction, the multiplex post-PCR LDR-FMA assay described here represents a significant advance because it detects the four major Plasmodium spp. and W. bancrofti infections in humans with high sensitivity and specificity in various assemblages.
Using the same assay, studies to detect these infections in mosquitoes collected from the same area in Papua New Guinea are underway. It is envisaged that because of its high-throughput format and high sensitivity and specificity, this multiplex assay has the potential to serve as a useful monitoring tool during and after implementation of large-scale control/elimination programs. In this regard, the assay can be successfully implemented whether DNA extraction was performed using whole blood, mosquitoes, or filter paper–dried blood spots. In addition to the four major Plasmodium spp. and W. bancrofti, future investigations can also include detection of the recently emerged human malaria parasite P. knowlesi34 and another important, but less prevalent, filariasis parasite, B. malayi, for worldwide application of this assay to the control/elimination programs targeting malaria and lymphatic filariasis.
Acknowledgments
We thank study participants in Papua New Guinea, and Dave McNamara and Brian Grimberg for critical comments on the manuscript.
Footnotes
Financial support: This study was supported by grants from the National Institutes of Health (AI065717 to James W. Kazura and Peter A. Zimmerman) and the Fogarty International Center (Ecology of Infectious Diseases, TW007872 to Peter A. Zimmerman).
Authors' addresses: Rajeev K. Mehlotra, Laurie R. Gray, Melinda J. Blood-Zikursh, Zachary Kloos, Cara N. Henry-Halldin, Daniel J. Tisch, Will Kastens, James W. Kazura, and Peter A. Zimmerman, Center for Global Health and Diseases, Case Western Reserve University School of Medicine, Wolstein Research Building, 4-125, Cleveland, OH. Edward Thomsen and Lisa Reimer, Center for Global Health and Diseases, Case Western Reserve University School of Medicine, Wolstein Research Building, 4-125, Cleveland, OH; and Papua New Guinea Institute of Medical Research, Madang 511, Papua New Guinea. Manasseh Baea, Kaye Baea, Moses Baisor, and Nandao Tarongka, Papua New Guinea Institute of Medical Research, Madang 511, Papua New Guinea.
References
- 1.Hotez PJ, Remme JH, Buss P, Alleyne G, Morel C, Breman JG. Combating tropical infectious diseases: report of the Disease Control Priorities in Developing Countries Project. Clin Infect Dis. 2004;38:871–878. doi: 10.1086/382077. [DOI] [PubMed] [Google Scholar]
- 2.Michael E, Bundy DA. Global mapping of lymphatic filariasis. Parasitol Today. 1997;13:472–476. doi: 10.1016/s0169-4758(97)01151-4. [DOI] [PubMed] [Google Scholar]
- 3.Muturi EJ, Jacob BG, Kim CH, Mbogo CM, Novak RJ. Are coinfections of malaria and filariasis of any epidemiological significance? Parasitol Res. 2008;102:175–181. doi: 10.1007/s00436-007-0779-1. [DOI] [PubMed] [Google Scholar]
- 4.Reeder JC. Health research in Papua New Guinea. Trends Parasitol. 2003;19:241–245. doi: 10.1016/s1471-4922(03)00089-8. [DOI] [PubMed] [Google Scholar]
- 5.Chadee DD, Rawlins SC, Tiwari TS. Short communication: concomitant malaria and filariasis infections in Georgetown, Guyana. Trop Med Int Health. 2003;8:140–143. doi: 10.1046/j.1365-3156.2003.01001.x. [DOI] [PubMed] [Google Scholar]
- 6.Ghosh SK, Yadav RS. Naturally acquired concomitant infections of bancroftian filariasis and human plasmodia in Orissa. Indian J Malariol. 1995;32:32–36. [PubMed] [Google Scholar]
- 7.Muturi EJ, Mbogo CM, Mwangangi JM, Ng'ang'a ZW, Kabiru EW, Mwandawiro C, Beier JC. Concomitant infections of Plasmodium falciparum and Wuchereria bancrofti on the Kenyan coast. Filaria J. 2006;5:8. doi: 10.1186/1475-2883-5-8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Burkot TR, Molineaux L, Graves PM, Paru R, Battistutta D, Dagoro H, Barnes A, Wirtz RA, Garner P. The prevalence of naturally acquired multiple infections of Wuchereria bancrofti and human malarias in anophelines. Parasitology. 1990;100:369–375. doi: 10.1017/s003118200007863x. [DOI] [PubMed] [Google Scholar]
- 9.Campbell CC. Halting the toll of malaria in Africa. Am J Trop Med Hyg. 2008;78:851–853. [PubMed] [Google Scholar]
- 10.Weil GJ, Ramzy RM. Diagnostic tools for filariasis elimination programs. Trends Parasitol. 2007;23:78–82. doi: 10.1016/j.pt.2006.12.001. [DOI] [PubMed] [Google Scholar]
- 11.Melrose WD, Durrheim DD, Burgess GW. Update on immunological tests for lymphatic filariasis. Trends Parasitol. 2004;20:255–257. doi: 10.1016/j.pt.2004.04.002. [DOI] [PubMed] [Google Scholar]
- 12.Nuchprayoon S, Porksakorn C, Junpee A, Sanprasert V, Poovorawan Y. Comparative assessment of an Og4C3 ELISA and an ICT filariasis test: a study of Myanmar migrants in Thailand. Asian Pac J Allergy Immunol. 2003;21:253–257. [PubMed] [Google Scholar]
- 13.Pani SP, Hoti SL, Vanamail P, Das LK. Comparison of an immunochromatographic card test with night blood smear examination for detection of Wuchereria bancrofti microfilaria carriers. Natl Med J India. 2004;17:304–306. [PubMed] [Google Scholar]
- 14.Berry A, Benoit-Vical F, Fabre R, Cassaing S, Magnaval JF. PCR-based methods to the diagnosis of imported malaria. Parasite. 2008;15:484–488. doi: 10.1051/parasite/2008153484. [DOI] [PubMed] [Google Scholar]
- 15.Erdman LK, Kain KC. Molecular diagnostic and surveillance tools for global malaria control. Travel Med Infect Dis. 2008;6:82–99. doi: 10.1016/j.tmaid.2007.10.001. [DOI] [PubMed] [Google Scholar]
- 16.Bisht R, Hoti SL, Thangadurai R, Das PK. Isolation of Wuchereria bancrofti microfilariae from archived stained blood slides for use in genetic studies and amplification of parasite and endosymbiont genes. Acta Trop. 2006;99:1–5. doi: 10.1016/j.actatropica.2005.12.009. [DOI] [PubMed] [Google Scholar]
- 17.Kanjanavas P, Tan-ariya P, Khawsak P, Pakpitcharoen A, Phantana S, Chansiri K. Detection of lymphatic Wuchereria bancrofti in carriers and long-term storage blood samples using semi-nested PCR. Mol Cell Probes. 2005;19:169–172. doi: 10.1016/j.mcp.2004.11.003. [DOI] [PubMed] [Google Scholar]
- 18.Rao RU, Atkinson LJ, Ramzy RM, Helmy H, Farid HA, Bockarie MJ, Susapu M, Laney SJ, Williams SA, Weil GJ. A real-time PCR-based assay for detection of Wuchereria bancrofti DNA in blood and mosquitoes. Am J Trop Med Hyg. 2006;74:826–832. [PMC free article] [PubMed] [Google Scholar]
- 19.Zhong M, McCarthy J, Bierwert L, Lizotte-Waniewski M, Chanteau S, Nutman TB, Ottesen EA, Williams SA. A polymerase chain reaction assay for detection of the parasite Wuchereria bancrofti in human blood samples. Am J Trop Med Hyg. 1996;54:357–363. doi: 10.4269/ajtmh.1996.54.357. [DOI] [PubMed] [Google Scholar]
- 20.Chansiri K, Kwoasak P, Tananyutthawongese C, Sukhumsirichart W, Sarataphan N, Phantana S. Detection of Plasmodium falciparum and Wuchereria bancrofti infected blood samples using multiplex PCR. Mol Cell Probes. 2001;15:201–207. doi: 10.1006/mcpr.2001.0359. [DOI] [PubMed] [Google Scholar]
- 21.Rao RU, Huang Y, Bockarie MJ, Susapu M, Laney SJ, Weil GJ. A qPCR-based multiplex assay for the detection of Wuchereria bancrofti, Plasmodium falciparum and Plasmodium vivax DNA. Trans R Soc Trop Med Hyg. 2009;103:365–370. doi: 10.1016/j.trstmh.2008.07.012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Muller I, Bockarie M, Alpers M, Smith T. The epidemiology of malaria in Papua New Guinea. Trends Parasitol. 2003;19:253–259. doi: 10.1016/s1471-4922(03)00091-6. [DOI] [PubMed] [Google Scholar]
- 23.Mehlotra RK, Kasehagen LJ, Baisor M, Lorry K, Kazura JW, Bockarie MJ, Zimmerman PA. Malaria infections are randomly distributed in diverse holoendemic areas of Papua New Guinea. Am J Trop Med Hyg. 2002;67:555–562. doi: 10.4269/ajtmh.2002.67.555. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Mehlotra RK, Lorry K, Kastens W, Miller SM, Alpers MP, Bockarie M, Kazura JW, Zimmerman PA. Random distribution of mixed species malaria infections in Papua New Guinea. Am J Trop Med Hyg. 2000;62:225–231. doi: 10.4269/ajtmh.2000.62.225. [DOI] [PubMed] [Google Scholar]
- 25.McNamara DT, Kasehagen LJ, Grimberg BT, Cole-Tobian J, Collins WE, Zimmerman PA. Diagnosing infection levels of four human malaria parasite species by a polymerase chain reaction/ligase detection reaction fluorescent microsphere-based assay. Am J Trop Med Hyg. 2006;74:413–421. [PMC free article] [PubMed] [Google Scholar]
- 26.Kasehagen LJ, Mueller I, McNamara DT, Bockarie MJ, Kiniboro B, Rare L, Lorry K, Kastens W, Reeder JC, Kazura JW, Zimmerman PA. Changing patterns of Plasmodium blood-stage infections in the Wosera region of Papua New Guinea monitored by light microscopy and high throughput PCR diagnosis. Am J Trop Med Hyg. 2006;75:588–596. [PMC free article] [PubMed] [Google Scholar]
- 27.Mueller I, Widmer S, Michel D, Maraga S, McNamara DT, Kiniboro B, Sie A, Smith TA, Zimmerman PA. High sensitivity detection of Plasmodium species reveals positive correlations between infections of different species, shifts in age distribution and reduced local variation in Papua New Guinea. Malar J. 2009;8:41. doi: 10.1186/1475-2875-8-41. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Bockarie MJ, Kazura JW. Lymphatic filariasis in Papua New Guinea: prospects for elimination. Med Microbiol Immunol (Berl) 2003;192:9–14. doi: 10.1007/s00430-002-0153-y. [DOI] [PubMed] [Google Scholar]
- 29.Bockarie MJ, Tisch DJ, Kastens W, Alexander ND, Dimber Z, Bockarie F, Ibam E, Alpers MP, Kazura JW. Mass treatment to eliminate filariasis in Papua New Guinea. N Engl J Med. 2002;347:1841–1848. doi: 10.1056/NEJMoa021309. [DOI] [PubMed] [Google Scholar]
- 30.Tisch DJ, Bockarie MJ, Dimber Z, Kiniboro B, Tarongka N, Hazlett FE, Kastens W, Alpers MP, Kazura JW. Mass drug administration trial to eliminate lymphatic filariasis in Papua New Guinea: changes in microfilaremia, filarial antigen, and Bm14 antibody after cessation. Am J Trop Med Hyg. 2008;78:289–293. [PMC free article] [PubMed] [Google Scholar]
- 31.Tisch DJ, Hazlett FE, Kastens W, Alpers MP, Bockarie MJ, Kazura JW. Ecologic and biologic determinants of filarial antigenemia in bancroftian filariasis in Papua New Guinea. J Infect Dis. 2001;184:898–904. doi: 10.1086/323324. [DOI] [PubMed] [Google Scholar]
- 32.Bockarie MJ, Alexander ND, Hyun P, Dimber Z, Bockarie F, Ibam E, Alpers MP, Kazura JW. Randomised community-based trial of annual single-dose diethylcarbamazine with or without ivermectin against Wuchereria bancrofti infection in human beings and mosquitoes. Lancet. 1998;351:162–168. doi: 10.1016/S0140-6736(97)07081-5. [DOI] [PubMed] [Google Scholar]
- 33.Ngwira BM, Jabu CH, Kanyongoloka H, Mponda M, Crampin AC, Branson K, Alexander ND, Fine PE. Lymphatic filariasis in the Karonga district of northern Malawi: a prevalence survey. Ann Trop Med Parasitol. 2002;96:137–144. doi: 10.1179/0003498302125000411. [DOI] [PubMed] [Google Scholar]
- 34.Cox-Singh J, Davis TM, Lee KS, Shamsul SS, Matusop A, Ratnam S, Rahman HA, Conway DJ, Singh B. Plasmodium knowlesi malaria in humans is widely distributed and potentially life threatening. Clin Infect Dis. 2008;46:165–171. doi: 10.1086/524888. [DOI] [PMC free article] [PubMed] [Google Scholar]