

Abstract
The molecule ATP, famous as an essential energy source inside cells, also carries critical messages between cells. That dual role is suggesting fresh ideas for fighting human diseases
One of the first and most enduring facts most students learn in biology class is that all living cells use a small molecule called adenosine triphosphate (ATP) as fuel. That universal energy currency drives the biological reactions that allow cells to function and life to flourish—making ATP a crucial player in the biological world.
Less commonly known, however, is that what is perhaps the most produced and consumed molecule in the human body also has a completely separate but no less essential role outside of cells. A long series of discoveries has now demonstrated beyond doubt that ATP is a critical signaling molecule that allows cells and tissues throughout the body to communicate with one another. The universal fuel, in effect, serves as a common language as well.
When ATP’s dual function was first proposed nearly 50 years ago, the idea met with considerable skepticism. But an avalanche of findings in the past 15 years has detailed how ATP acts on cells from the outside and how it serves in the development and daily operation of organs and tissues. Because ATP is so ubiquitous, its signaling actions have a uniquely broad influence on physiological functioning and offer unusually diverse opportunities to improve human health. Laboratories worldwide are now racing to turn these insights into therapies.
ATP Unmasked, Twice
When ATP was discovered in 1929, investigators around the world were seeking the elusive source of cellular energy. In nearly simultaneous breakthroughs, Karl Lohmann, working with the 1922 Nobel Laureate Otto Meyerhof of the Kaiser Wilhelm Institute for Medical Research in Heidelberg, and Cyrus H. Fiske, working with his graduate student Yellapragada SubbaRow of Harvard Medical School, showed that intracellular activities that allow muscle cells to contract depended on a molecule made of a purine—adenosine, a combination of the base adenine with a sugar—and three phosphates. By 1935 Katashi Makino of Dailen Hospital in Manchuria had proposed a structure for the molecule, which was confirmed 10 years later, by Basil Lythgoe and Alexander R. Todd of the University of Cambridge Chemical Laboratory.
During this period no one envisioned a role for the molecule outside of the cell. That was still the case in 1962, when one of us (Burn-stock) was a young neurophysiologist at the University of Melbourne in Australia, studying the nerves that control smooth muscle tissue. In the course of investigating signaling by the autonomic nervous system (which controls such basic muscle-dependent functions as intestinal and bladder contractions), he saw evidence of neural signaling that did not involve the classical neurotransmitter chemicals acetylcholine or noradrenaline. Intrigued by data published in 1959 by Pamela Holton of the Cambridge Physiological Laboratory suggesting that sensory nerves released ATP molecules, Burn-stock set out to determine whether ATP could be responsible for signaling between motor nerves and muscle. Through a series of experiments in which he applied chemicals to block signaling by the classical neurotransmitters to smooth muscle tissue, he was able to demonstrate that any continued signaling from the nerves to the muscle had to be conveyed by ATP. Pursuing this lead for more than a decade, Burnstock felt confident enough by 1972 to propose the existence of “purinergic nerves” that release ATP as a neurotransmitter.
Nerve cells generate electrical impulses that travel the length of a single neuron, but the charge does not cross the tiny gap between the cells known as the synaptic cleft, or the gap between nerve cells and muscles. The message is forwarded from cell to cell by chemical transmitters, such as acetylcholine, glutamate, dopamine and others, released from the firing neuron into the cleft. These chemicals cross the gap and bind to receptor proteins on the receiving cell, causing that cell to undergo a series of internal changes that alter its activity; recipient neurons might fire impulses of their own, and muscle cells might contract or relax. A message can thus be transmitted down the line from neuron to neuron by an alternating series of impulses and chemical discharges.
Individual neurons were long believed to emit only a single neurotransmitter type, and cells that released acetylcholine came to be described as cholinergic; those that released dopamine, were dopaminergic, and so forth. Burn-stock’s concept of purinergic neurons was based not only on his own observations by that point but also on the early work of a series of outstanding students and collaborators, including Max Bennett, Graeme Campbell, David Satchell, Mollie Holman and Mike Rand of the universities of Melbourne and London.
Despite a huge amount of data showing ATP release from neurons into muscle, gut and bladder tissue, many neurophysiologists nonetheless remained skeptical about the existence of nerves releasing ATP as a messenger, largely because they thought it unlikely that such a ubiquitous substance could perform such a specific role. Moreover, for a signaling molecule to be able to function, it must find a suitable receptor on its target cell. The first receptor for a neurotransmitter had been isolated only in 1970; therefore, the hunt was on for the receptors for ATP.
Well before they were found, however, many researchers continued to use pharmacological methods to examine how ATP released by neurons delivered messages into muscle and other cells of the body. Based on this work, Burnstock suggested in 1978 that separate families of receptors existed for ATP (which he designated P2 receptors) and for its final breakdown product, adenosine (which he called P1 receptors). Further studies showed that ATP activation of P2 receptors could produce different cellular effects. That led Burnstock and his collaborator Charles Kennedy to anticipate the existence of P2 receptor subtypes, which they named P2X and P2Y.
Still, the idea of nerves that released ATP as a neurotransmitter remained controversial and was dismissed by many for years to come. In the 1990s, though, molecular tools became available that allowed many research groups to isolate ATP receptors and to further explore their many fascinating effects on cells of the nervous system and beyond.
Interplay and Dynamics
The early 1990s saw the initiation of the Human Genome Project and the beginning of an era of prolific discovery of the genes that encode important proteins in the human body. Among these genes were several for ATP receptors, which allowed scientists to locate the receptors themselves on many different cell types. Studies of ATP signaling entered a new and exciting era. Attempts to characterize the molecular structure of purine receptors proved the existence of a large family of receptors and identified a number of channels and enzymes on the surface of cells that participate in ATP signaling.
As predicted, two broad classes of receptor were identified, but the work also revealed many more receptor subtypes than expected within those classes. This diversity implied that particular receptor subtypes could be targeted with highly selective drugs to modulate ATP signaling only in specific tissues or cell types—a prospect that is bearing fruit today [see table on page 92].
[DRUGS]. TARGETING ATP RECEPTORS.
Identification of the specific receptor subtypes responsible for ATP’s signaling effects in various tissues has allowed pharmaceutical companies to begin developing therapies for a range of disorders. Two of the drugs listed below are already marketed; the rest are still under study.
| DISORDER | DRUG | MECHANISM | TESTING STAGE |
|---|---|---|---|
| Cystic fibrosis | Denufosol | Activates P2Y2 receptors | Late-stage human efficacy tests under way |
| Dry eye | Diquafosol | Activates P2Y2 receptors | Late-stage human efficacy tests under way |
| Inflammation | EVT 401 | Inhibits P2X7 receptors | Human safety tests completed |
| Pain | GSK1482160 | Inhibits P2X7 receptors | Human safety tests under way |
| Unnamed compounds (from Evotec AG) | Inhibit P2X3 and P2X2/3 receptors | Cell and animal testing | |
| Rheumatoid arthritis | CE-224,535 | Inhibits P2X7 receptors | Late-stage human efficacy tests completed |
| AZD9056 | Inhibits P2X7 receptors | Human safety tests completed | |
| Thrombosis (aberrant blood clotting) | Clopidogrel | Inhibits P2Y12 receptors | Approved |
| Prasugrel | Inhibits P2Y12 receptors | Approved | |
| PRT060128 | Inhibits P2Y12 receptors | Human safety and efficacy tests under way | |
| Ticagrelor | Inhibits P2Y12 receptors | Late-stage human efficacy tests under way |
After the initial isolation of ATP receptors, various investigators showed that the two main classes operate in significantly different ways. P2X receptors belong to a “superfamily” of transmitter-gated ion channels. One of us (Khakh), along with other investigators, showed that when bound by ATP, P2X receptors literally open to form a channel that allows sodium and large amounts of calcium ions to rush into cells. P2Y receptors, in contrast, do not open in the same way, but ATP binding to their extra-cellular surface sets off a cascade of molecular interactions inside cells that results in intracellular calcium stores being released. In both cases, the calcium can then set off further molecular events that alter cell behavior.
Although ATP stays in the synaptic cleft only briefly, the cellular effects of receptor activation can occur quickly in some instances—within milliseconds—but slowly in others—sometimes over the course of years. For example, an inrush of calcium ions through P2X channels may cause the cell to release other transmitters, as Khakh has shown in brain tissue, or calcium released by P2Y activation may alter gene activity involved in cell proliferation that causes changes in the tissue with lifelong consequences. Even though the presence of ATP molecules in the extracellular space is fleeting, therefore, their biological effects can be quite pervasive.
The mechanisms of ATP signaling become even more fascinating when its interactions with other signaling systems outside of cells are taken into account. A large family of enzymes known as ectoATPases sit on the surface of most cells, where they quickly strip ATP of its phosphates one by one—sequentially turning an ATP molecule into adenosine diphosphate (ADP), adenosine monophosphate (AMP) and finally adenosine alone. Each of ATP’s breakdown products may have an effect of its own on a cell—such as when adenosine binds to P1 receptors.
Fusao Kato of the Jikei University School of Medicine in Tokyo has shown, for instance, that ATP and adenosine act in concert in the brain stem network responsible for basic body functions such as breathing, heart rhythm and gastrointestinal action. Other situations exist, however, where ATP and adenosine oppose each other, such as during neuron-to-neuron transmission, where adenosine can inhibit a neuron from releasing ATP into the synaptic cleft. The interconnected effects of ATP, its component parts and the extracellular ectoATPases can thus be seen as forming a self-regulating signaling loop in many circumstances.
It is not only ATP breakdown products that influence the molecule’s effects on cells. In the nervous system, ATP also acts in concert with other neurotransmitters as a co-transmitter. Discovery of this phenomenon in 1976 by Burn-stock helped to revise the long-standing view that any given neuron can synthesize, store and release just one kind of neurotransmitter. Today a substantial body of evidence shows that ATP is typically released along with classical neurotransmitters, such as noradrenaline or acetylcholine. Although co-transmission was first proposed and proved for ATP, the phenomenon of neurons that co-release transmitter molecules has now also been demonstrated for a variety of other transmitters, including GABA with glycine, dopamine with serotonin, and acetylcholine with glutamate. Co-transmission is thus another example of how studies of ATP signaling have revealed more general physiological principles as well as shaped and guided research in other fields.
ATP in Health and Disease
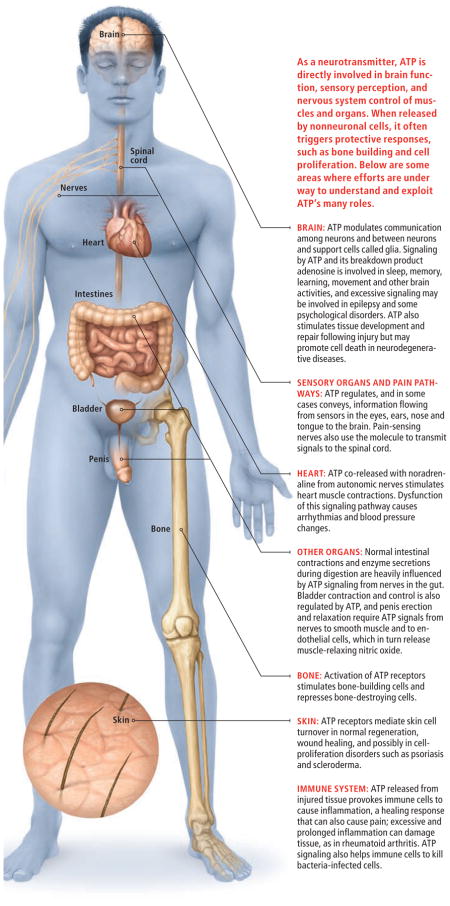
In light of ATP’s established role in signaling among cells of the nervous system, it may come as no surprise that ATP plays an important part in the functioning of the five senses. In the eye, for example, ATP receptors on nerve cells in the retina influence the cells’ responses to information received from rods and cones, the eyes’ light detectors. The retinal nerves, in turn, dispatch ATP and acetylcholine as co-transmitters to convey their information to sensory-processing centers in the brain. In addition to this everyday function for ATP, several research groups have shown that ATP signaling at a key point during an embryo’s eye development can have effects that last a lifetime. Indeed, Nicholas Dale of the University of Warwick in England and his colleagues have shown that release of ATP at a critical time in the early embryo is the signal for the development of eyes.
Release of ATP during development is also essential for the proper formation of the cochlea, the organ responsible for hearing, and ATP signaling continues to be crucial to the workings of the inner ear in adults. Some 50,000 hair cells—the sound-transducing neurons of the inner ear—line the human cochlea, and about half of those display receptors for ATP, which has been shown to ease neural firing in some circumstances. In addition, taste buds, the sensory nerve endings in the tongue, possess P2X receptors that mediate taste. In a particularly well-designed study, Sue C. Kinnamon and her colleagues at Colorado State University have demonstrated that ATP is vital as a transmitter from taste bud cells to gustatory nerves and that mice lacking both P2X2 and P2X3 receptor sub-types are incapable of tasting.
Interestingly, the P2X2 and P2X3 receptors present on taste buds are the same ones involved in certain types of pain signaling. For decades scientists have known that ATP introduced into the skin causes pain. Stephen B. McMahon and his colleagues at Guy’s, King’s and St. Thomas’ School of Biomedical Sciences in London, recently showed that the pain is triggered by the activation of P2X3 ATP receptors on the sensory nerve endings in the skin that mediate responses to both touch and pain. Another form of pain, one associated with damage to nerves, is called neuropathic pain and involves ATP by a different route. Elegant studies from Kazuhide Inoue of Kyushu University in Japan and Michael Salter of the University of Toronto show that a key step in the development of this type of pain involves activation of ATP receptors on spinal cord immune cells called microglia. The microglia, in turn, release molecules that irritate nerve fibers, leading to chronic pain [see “New Culprits in Chronic Pain,” by R. Douglas Fields; Scientific American, November 2009].
Because of such insights into ATP’s signaling role, several pharmaceutical companies are now pursuing P2X receptors as new drug targets for neuropathic pain or pain caused by inflammation. And pain is but one aspect of human health that may soon benefit from therapies aimed at ATP or its receptors.
People with heart and blood vessel disorders are among those who stand to benefit from future drugs that act on ATP receptors. The reason becomes clear when one looks at the events that follow an injury. Cells that are distressed or physically damaged can release or spill ATP into the extracellular space. In those situations, ATP signaling often results in protective and healing responses, including by blood platelets, the cells responsible for forming a clot to stop bleeding from a new wound. Platelets display the P2Y12 receptor subtype, and its activation by extracellular ATP causes them to undergo changes that lead to clot formation. Of course, this same process contributes to the formation of the clots in blood vessels that can cause heart attacks and stroke. An existing “blockbuster” drug, clopidogrel, works by blocking the P2Y12 receptor on platelets and thereby preventing ATP from promoting clots. A handful of drugs that function in related ways are also in advanced clinical trials for coronary disorders.
A similarly promising therapeutic area is the digestive system. James J. Galligan of Michigan State University and others have demonstrated that ATP sent from the intestinal nervous system to the intestinal wall acts on P2X and P2Y receptors to control the rhythmic contractions that move food through the tract. Meanwhile ATP that binds to P2Y receptors on cells lining the inner surface of the gut wall triggers secretion of digestive enzymes. Agents that act on those receptors to modulate these functions are therefore being hotly pursued by pharmaceutical companies as potential treatments for irritable bowel syndrome and its more severe form, Crohn’s disease.
The involvement of ATP in healthy functioning of other organs and tissues makes it a possible drug target in a long list of disorders, including diseases of the kidney, bone, bladder, skin, and even neurological and psychiatric illnesses. What is more, ATP may be one of the body’s natural cancer-fighting tools. Eliezer Rapaport, when at the Boston University School of Medicine, first described a tumor-killing effect of ATP in 1983. He, too, was met with skepticism, but research since then by a number of laboratories working independently has shown that ATP can inhibit the growth of tumors, including prostate, breast, colorectal, ovarian and esophageal cancers, as well as melanoma cells. ATP signaling acts in part to promote suicide of the tumor cells and in part to promote cell differentiation, which slows tumor cell proliferation.
Much work remains to be done to translate new insights into ATP signaling gathered so far into novel medicines ready for use in the clinic. But many laboratories and drug companies are actively looking for drugs that can selectively activate or silence specific ATP receptor subtypes, inhibit or enhance the release of ATP, or inhibit the breakdown of ATP after it has been released from cells.
The Ultimate Messenger
ATP’s ubiquity as a signaling molecule does pose at least one major challenge: developing drugs targeted only to a single organ or tissue without causing side effects in other body systems. This concern is not unique to ATP, however, and the great variety of subunit configurations found on different cell types will make targeting specific tissues more feasible. Khakh has been experimenting with creating “designer” ATP receptors that can be incorporated into cultured cells or even living laboratory mice and used to test the effects of subtly changing the function of a P2X receptor protein. This is just one approach that allows researchers to manipulate ATP signaling in a controlled manner and study the results in living organisms.
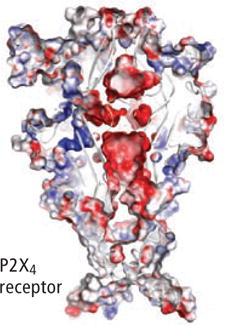
One of the most important breakthroughs in the past 20 years has been the recent determination of the crystal structure of a zebra fish P2X channel by Eric Gouaux and his colleagues at Oregon Health and Science University. This landmark achievement shows atomic-scale details of how an ATP receptor works and paves the way for an understanding of ATP signaling from the level of molecules to that of whole physiological systems. It will also significantly accelerate the process of drug discovery.
Recent evidence of ATP receptors in plants and primitive organisms, such as green algae, amoebas and parasitic schistosomes, offers the possibility that targeting ATP signaling may also be useful in agriculture and in the treatment of infectious diseases. The presence of ATP signaling in such diverse life-forms suggests, too, that ATP’s function as a signaling molecule appeared early in the evolution of life—perhaps more or less simultaneously with its adoption as an energy source. Many reports of potent effects caused by ATP and its derivatives in most invertebrate and lower vertebrate animals also suggest that ATP’s influence could be widespread indeed.
It is gratifying for us to see how the role of ATP as a signaling molecule has gone from an idea that was widely deemed dubious 50 years ago to a large and vibrant field of inquiry today of interest to the entire biology community and of great potential import to medicine. We look forward to seeing how further breakthroughs in understanding the fascinating double life of ATP are exploited to improve the quality of human life.
ATP SIGNALING: A BRIEF HISTORY
1929 ATP discovered to be the energy source in muscle tissue.
1929 Albert Szent-Györgyi finds purines (ATP’s chemical family) have potent effects on the heart.

1945 ATP structure confirmed.
1959 Pamela Holton shows ATP release from sensory nerves.
1962 Geoffrey Burnstock demonstrates message transmission from neurons to muscle by a new neurotransmitter.

1972 Burnstock proposes the existence of nerves that signal using ATP.
1976 Burnstock proposes that ATP acts as a co-transmitter with other neurotransmitters.
1993 and 1994 P2X and P2Y receptors for ATP isolated from cells.
1998 Clopidogrel, a drug that acts on platelet P2Y receptors, introduced to prevent clot formation in blood vessels.
2009 Crystal structure of a P2X receptor revealed, which should aid drug discovery.
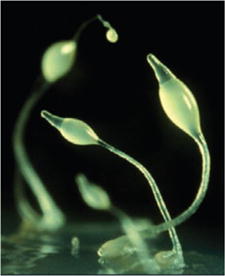
EARLY ORIGIN
The discovery of ATP receptors in plants and primitive life-forms, such as amoebas and worms, suggests that the molecule took on a signaling role very early in the evolution of life. In the slime mold Dictyostelium discoideum (below), ATP-activated receptors that resemble human P2X channels control the flow of water into and out of cells.
KEY CONCEPTS
ATP, best known as a universal fuel inside living cells, also serves as a molecular signal that affects cell behavior.
A leading investigator and the discoverer of ATP’s messenger role describe how ATP signals work and why they are essential to basic bodily functions and development.
Because ATP is so ubiquitous, the molecule’s influences can vary from tissue to tissue, offering new insights into a wide range of disorders and diverse ways to treat them.
–The Editors
[DUAL ROLES].
ATP INSIDE CELLS … AND OUTSIDE
Students routinely learn that the small molecule adenosine triphosphate (ATP) is a critical cellular power source; it fuels the activities of the molecular machinery that allows all cells to function and thrive (below). But not all intracellular ATP is used up by cellular processes. Cells of all kinds also release ATP to send messages to nearby cells (right).
![[DUAL ROLES]](https://cdn.ncbi.nlm.nih.gov/pmc/blobs/4b76/2877495/8c6ab349753e/nihms199392u4.jpg)
[ATP IN ACTION].
ONE SIGNAL, MANY MESSAGES
ATP-signaling activity was first detected between nerve cells and muscle tissue but is now known to operate within a wide variety of cell types in the body. Select examples from the cardiovascular system illustrate how diverse ATP’s effects can be in their nature and duration.
![[ATP IN ACTION]](https://cdn.ncbi.nlm.nih.gov/pmc/blobs/4b76/2877495/16f1514cbc1f/nihms199392u5.jpg)
ANATOMY OF ATP
Biographies

Baljit S. Khakh is an Assistant professor of physiology and neurobiology at the David Geffen School of Medicine at the University of California, Los Angeles. He has developed novel tools, such as designer ATP receptors that can be monitored by light, to probe how cells sense and respond to ATP

Geoffrey Burnstock, the first to show that ATP acts as a signaling molecule, was chair of the department of anatomy and developmental biology at University College London for 22 years and is now president of the Autonomic Neuroscience Center at the Royal Free and University College Medical School in London. He has won numerous awards and honors. He and Khakh met in 1994 at a coffee shop in Vienna, where they discussed ATP over apple strudel