

Abstract
Toll-like receptor (TLR) agonists induce potent innate immune responses and can be used in the development of novel vaccine adjuvants. However, access to TLRs can be challenging as exemplified by TLR 7, which is located intracellularly in endosomal compartments. To increase recognition and subsequent stimulatory effects of TLR 7, imiquimod was encapsulated in acetalated-dextran (Ac-DEX) microparticles. Ac-DEX, a water-insoluble and biocompatible polymer, is relatively stable at pH 7.4, but degrades rapidly under acidic conditions, such as those found in lysosomal vesicles. To determine the immunostimulatory capacity of encapsulated imiquimod, we compared the efficacy of free versus encapsulated imiquimod in activating RAW 264.7 macrophages, MH-S macrophages, and bone marrow derived dendritic cells. Encapsulated imiquimod significantly increased IL-1β, IL-6, and TNF-α cytokine expression in macrophages relative to the free drug. Furthermore, significant increases were observed in classic macrophage activation markers (iNOS, PD1-L1, and NO) after treatment with encapsulated imiquimod over the free drug. Also, bone marrow derived dendritic cells produced significantly higher levels of IL-1β, IL-6, IL-12p70, and MIP-1α as compared to their counterparts receiving free imiquimod. These results suggest that encapsulation of TLR ligands within Ac-DEX microparticles results in increased immunostimulation and potentially better protection from disease when used in conjunction with vaccine formulations.
Keywords: Vaccination, microparticles, acid-degradable materials, dendritic cell, imiquimod, macrophage
Introduction
Vaccines prepared from attenuated or heat-inactivated viruses have been very successful in treating pathogens such as poliomyelitis (1), Variola major (2), and influenza (3). However, due to safety concerns, attenuated viruses are not used for pathogens such as HIV and Hepatitis C (4).
Subunit vaccines are considered to be a safer alternative to live or attenuated virus vaccine formulations because they can establish protective immunity, yet lack the molecular machinery to cause an infection. However, subunit vaccines typically lack the inherent immunostimulatory capacity of attenuated viruses. In general, the use of immunostimulatory molecules, which are typically recognized by the innate immune system, drastically increase the immune response generated against the specific antigen in subunit vaccines (5,6). Activation of an innate immune response occurs by stimulation of pattern recognition receptors (PRR), such as toll-like receptors (TLRs) and the more recently identified NOD-like receptors (NLRs) (7,8). PRRs are highly conserved structures amongst an array of species that activate after binding of pathogen associated molecular patterns (PAMPs) such as lipopolysacharride (LPS) and unmethylated CpG DNA. The binding of PAMPs to TLRs results in significant immune activation characterized by dendritic cell maturation, inflammatory cytokine production, trafficking of dendritic cells to secondary lymphoid tissue, and activation of adaptive immune responses through interactions with naïve T cells (7). This mechanism allows TLR agonists to drastically increase the potency of subunit vaccines.
Imidazoquinolines (imiquimod or resiquimod) are synthetic FDA approved immunostimulants that have recently been successful in improving the efficacy of subunit vaccines (9). Traditionally, imidazoquinolines have been used for the treatment of several ailments including genital warts (10), actinic keratosis (11,12), and several forms of carcinoma and melanoma (13-16). Imidazoquinolines bind TLR 7 and TLR 8, which are expressed within endosomal compartments of macrophages and multiple subsets of dendritic cells (17,18). Stimulation of TLR 7 or TLR 8 induces the activation of the MyD88 signaling cascade (18), ultimately resulting in the production of the inflammatory cytokines IL-1β, IL-6, TNF-α, and IFN-α (7). The activity of these compounds has been attributed to their ability to stimulate antibody and T cell mediated responses as well as inducing significant inflammatory cytokine production when topically administered (19,20). Despite the successful use of this molecule in the treatment of several illnesses, it is not without limitations. In particular, imiquimod is a poorly soluble compound limiting parenteral administration in vivo (21). In addition, when delivered systemically, imiquimod initiates a cascade of inflammatory cytokines and pro-apoptotic factors that prove detrimental to the host (22). For this reason, delivery of imiquimod is restricted to topical administration, which limits the range of ailments to which it may be applied.
Encapsulation of TLR agonists in polymeric carriers is a common technique for passively targeting antigen presenting cells (APCs) due to their inherent ability to phagocytose/endocytose particulate material. CpG, a TLR 9 agonist (23-25), and poly (I:C), a TLR 3 agonist (26), have been co-encapsulated in polymeric carriers with antigenic proteins resulting in an enhancement of both CD4 and CD8 T cell activation against the encapsulated protein. The TLR ligands for both CpG and poly (I:C) are intracellular. Depending on the size of a particle, the particle can be internalized by endocytosis via clathrin-coated pits (20-200 nm), taken up by macropinocytosis (0.5-5 μm), or internalized via phagocytosis (> 0.5 μm) (27). Although not completely understood, recent data suggest that in analogy to MHC class II molecules, intracellular TLRs such as 7 and 9 are trafficked through the Golgi via the secretory pathway to endolysosomal(28) and phagolysosomal compartments (29). Regardless of the mechanism of internalization, intracellular TLRs should have access to particulate carriers and their encapsulated cargo. Ideally, when particles encapsulating CpG or poly (I:C) are exposed to lysosomal conditions, the particles should be capable of releasing the encapsulated adjuvant, thus activating the phagocytic cell. Typically, the incorporation of the hydrophilic molecules CpG and poly (I:C) in microparticles requires the adjuvant to be complexed with a polycation (30,31). The incorporation of a polycation within the polymeric carrier may increase the toxicity of the microparticle. To date, no research has studied the incorporation of the hydrophobic immunostimulant imiquimod in microparticles and their subsequent activity in macrophages and dendritic cells. Since imiquimod is hydrophobic, and the TLR ligand it interacts with is intracellular, the encapsulation of imiquimod is highly desirable. The incorporation of imiquimod in a hydrophobic microparticle should have a high encapsulation efficiency, not require the incorporation of a polycation, and since imiquimod can interact with intracellular TLRs the activation of dendritic cells should be relatively high.
The most common polymer used as a carrier for drug delivery applications is poly (lactic-co-glycolic acid) (PLGA) due to its inherent biodegradability and low toxicity. However, PLGA may not be ideally suited for vaccine applications. For example, it may be beneficial to have a material that is sensitive to the acidic environment present in the phago-lysosomal compartments of macrophages and dendritic cells (∼pH 5) (32). An acid-sensitive material allows for the expedient delivery of protein to the cross-presentation machinery present in the phagosome (33,34). This release mechanism cannot be easily controlled when pH-insensitive carrier materials such as PLGA are used. In addition, as PLGA degrades, an acidic microenvironment is created within the particle matrix as the byproducts of PLGA degradation, lactic and glycolic acid, accumulate. Prolonged exposure to acidic environments may be harmful to the stability of recombinant proteins used in vaccines, e.g., tetanus toxoid and diphtheria toxoid (35). Due to these inherent issues with PLGA, new polymeric carriers are desired for vaccine applications.
Acetalated dextran (Ac-DEX) is a recently described polymer that is made by appending acetals onto the hydroxyl groups of dextran, a homopolysaccharide of glucose (36). This reaction renders the modified dextran soluble in common organic solvents, but insoluble in water. These properties allow for the facile processing of Ac-DEX into microparticles encapsulating antigens for vaccine applications through the use of standard emulsion techniques. Microparticles made from Ac-DEX are acid-sensitive due to the pH-dependency of acetal hydrolysis, and degrade more quickly under lysosomal conditions (∼ pH 5) compared to the extracellular environment (pH 7.4). This pH-sensitivity allows for a significant increase in both MHC I and MHC II presentation relative to other carrier materials (37). In addition, the degradation of Ac-DEX does not lead to acidic by-product, and therefore may be more suitable for pH-sensitive antigens.
In this study, we investigated the potential of using Ac-DEX microparticles as a delivery system for imiquimod induced stimulation. Encapsulated imiquimod was delivered to macrophages and dendritic cells and assayed for inflammatory cytokine production and activation. Herein, we find that Ac-DEX particles are an ideal delivery vehicle for the encapsulation and delivery of imiquimod to macrophages and dendritic cells.
Materials and Methods
Cell lines and bone marrow-derived cells
MH-S and RAW 264.7 macrophages were purchased from ATCC (Manassas, VA). Cells were grown and maintained as per guidelines provided by the manufacturer. Bone marrow-derived dendritic cells were prepared as previously described (37). Briefly, bone marrow was harvested from 6-8 week old A/J mice and treated with 3 ng/mL each of IL-4 and GM-CSF for 5 days. Cells were harvested after 5 days and sorted via magnetic bead isolation for CD11c expression. The experiments reported herein were conducted in compliance with the Animal Welfare Act and in accordance with the principles set forth in the “Guide for the Care and Use of Laboratory Animals,” Institute of Laboratory Animals Resources, National Research Council, National Academy Press, 1996.
General Materials and Methods
All reagents were purchased from commercial sources and used without further purification. Water (dd-H2O) for buffers and particle washing steps was purified to a resistance of 18 MΩ using a NANOpure purification system (Barnstead, USA). When used in the presence of acetal-containing materials, dd-H2O was rendered basic (pH 9) by the addition of triethylamine (TEA) (approximately 0.01%). Fluorescence measurements were obtained on a Spectra Max Gemini XS microplate reader (Molecular Devices, USA), usage courtesy of Prof. Jonathan Ellman.
Synthesis of Acetalated Dextran (Ac-DEX)
Ac-DEX was synthesized from 10 kDa dextran as described previously (36).
Preparation of Single Emulsion Particles Encapsulating Imiquimod
Microparticles containing imiquimod were prepared using an oil-in-water emulsion method similar to the technique described by Bachelder et al. (38). Imiquimod (Sigma-Aldrich) (4 mg) was dissolved in chloroform (2 mL) by heating the imiquimod/chloroform solution for 15 minutes at 50 °C. The solution was allowed to cool to room temperature, and then used to dissolve Ac-DEX (100 mg). The resulting solution was added to an aqueous solution of poly(vinyl alcohol) (PVA, MW = 13,000 – 23,000 g/mol, 87-89% hydrolyzed) (2 mL, 3% w/w in PBS) and sonicated for 30 s on ice using a probe sonicator (Branson Sonifier 450, with a 0.5 in. flat tip) with an output setting of 3 and a duty cycle of 10%. The resulting single emulsion was immediately poured into a second PVA solution (10 ml, 0.3% w/w in PBS) and stirred for 4 h to allow the organic solvent to evaporate. The particles were isolated by centrifugation (14,800 × g, 15 min, 4°C) and washed with dd-H2O (3 × 50 mL, pH 9). The washed particles were re-suspended in dd-H2O (2 mL, pH 9) and lyophilized to yield a white fluffy solid (87 mg).
Preparation of Empty Single Emulsion Particles
Empty Ac-DEX particles were made in the same manner as above omitting imiquimod.
Quantification of Encapsulated Imiquimod
Particles (1 mg) were dissolved in DMSO (1 mL) and analyzed for imiquimod content by measuring fluorescence using a Spectra Max Gemini XS microplate reader (Molecular Devices, Sunnyvale, CA) (ex. 325 nm, em. 365 nm). The results were compared to a standard curve and the mass of imiquimod was calculated. The imiquimod loading was found to be 4.1 ± 0.2 wt% and the loading efficiency was essentially quantitative.
Scanning Electron Microscopy
Microparticles were characterized by scanning electron microscopy using an S-5000 microscope (Hitachi, Japan). Particles were suspended in dd-H2O (pH 9) at a concentration of 1 mg/mL and the resulting dispersions were dripped onto a silicon wafer. After 15 min, the remaining water was wicked away using tissue paper and the samples were allowed to air dry. The particles were then sputter coated with a 2 nm layer of a palladium/gold alloy and imaged.
Release profile of imiquimod
4 mg of particles (in triplicate per pH) were suspended in either 1ml of PBS (pH 7.4) or 0.3 M acetate buffer (pH 5.0). The particle suspensions were then injected into a presoaked slide-a-lyzer cassette per the manufacturer's instructions (Thermo Scientific, Waltham, MA). The slide-a-lyzer cassette was placed in 800 ml of buffer at 37°C and stirred moderately. Samples were taken at indicated times and stored at -20 °C. Imiquimod release was calculated by measuring fluorescence using a Spectra Max Gemini XS microplate reader (ex. 325 nm, em. 365 nm).
Particle Size Analysis by Dynamic Light Scattering
Particle size distributions were determined by dynamic light scattering using a Zetasizer Nano ZS (Malvern Instruments, United Kingdom). Particles were suspended in dd-H2O (pH 9) at a concentration of 1 mg/mL and three measurements were taken of the resulting dispersions. The results in the text are presented as average particle diameters ± width of the distribution at half maximal height.
In vitro Toxicity
To assess of the relative toxicity of Ac-DEX microparticles, MH-S and RAW 264.7 macrophages were plated at 1×104 cells/mL and incubated overnight in RPMI medium 1640/10% FBS/1% penicillin-streptomycin. The medium in each well was replaced by 100 μL of new medium containing either empty Ac-DEX particles, imiquimod encapsulated in Ac-DEX particles or free imiquimod, ranging from 0.25 μg/mL-4 mg/mL. Assays were conducted in replicates of three for each concentration. After incubation for 20 h, a 40 μL solution of MTT solution (2.91 mg/mL) was added to each well. The cells were incubated for 3 h, after which time the medium was carefully removed. DMSO (200 μL) was added to the resulting purple crystals followed by a glycine buffer (25 μL, pH 10, 0.1 M glycine, 0.1 M NaCl). The optical densities at 570 nm were measured using a SpectraMAX 190 microplate reader (Molecular Devices, Sunnyvale, CA).
Nitrite Analysis
Briefly, the concentration of nitrite in supernatants of RAW 264.7 and MH-S alveolar macrophages was determined spectrophotometrically using the Griess reagent. Macrophages were plated at 5 × 105 cells/mL in 24-well plates and left overnight to adhere. Cells were treated with serial dilutions of medium, free imiquimod, imiquimod encapsulated in Ac-DEX particles, or empty particles for 24 hours. Supernatants were collected after 24 hours, mixed 1/1 with Griess reagent, and absorbances were measured at 543 nm using a SpectraMax 360 microplate reader (Molecular Devices, Sunnyvale, CA). The nitrite concentration was determined using sodium nitrite as a standard.
RNA isolation and Purification and Real Time Polymerase Chain Reaction
MH-S alveolar macrophages and RAW 264.7 macrophages were plated in 24-well plates overnight at 5×105 cells/mL and treated the next day with medium, free imiquimod, imiquimod encapsulated in Ac-DEX particles, or empty particles for 6 hours. Cells were lysed with Buffer RLT (Qiagen Sciences, Valencia, CA), containing 2-β-mercaptoethanol, and spun through Qiashredder tubes (Qiagen Sciences, Valencia, CA). Total RNA was extracted using the RNeasy Mini Kit (Qiagen Sciences, Valencia, CA). Individual sample RNA (0.1 μg) was reverse-transcribed using Superscript II (Invitrogen, Carlsbad, CA) and a mixture of oligo (dT) and random primers. Real-time polymerase chain reaction (RT-PCR) was performed on an ABI Prism 7900HT sequence detection system (Applied Biosystems, Foster City, CA). Sequences for hypoxanthine guanine phosphoribosyl transferase (HPRT) were published in (39), while sequences for IL-1, IL-6, TNF-, iNOS, and PD1-L1 were generated using the Universal Probe Library Assay Design Center (Roche Applied Science). Sequences are as follows: IL-1β Forward-TGTAATGAAAGACGGCACACC Reverse-TCTTCTTTGGGTATTGCTTGG, IL-6 Forward-TCCAGTTGCCTTCTTGGGAC Reverse-GTGTAATTAAGCCTCCGACTTG, TNF-α Forward-GCCTCTTCTCATTCCTGCTTGT Reverse-GGCCATTTGGGAACTTCTCAT, iNOS Forward-TGCCCCTTCAATGGTTGGTA Reverse-ACTGGAGGGACCAGCCAAAT, PD1-L1 Forward-CCATCCTGTTGTTCCTCATTG Reverse-TCCACATCTAGCATTCTCACTTG. Relative quantities of mRNA for several genes was determined using SYBR Green PCR Master Mix (Applied Biosystems, Foster City, CA) and by the comparative threshold cycle method as described by Applied Biosystems for the ABI Prism 7900HT sequence detection systems. In this method, mRNA levels for each sample were normalized to HPRT mRNA levels and then expressed as a relative increase or decrease compared with levels in untreated controls.
Multiplex-bead analysis
MH-S alveolar macrophages and RAW 264.7 macrophages were plated in 24-well plates overnight at 5×10 cells/mL, and treated the next day with medium, free imiquimod, imiquimod encapsulated in Ac-DEX particles, or empty particles for 24 hours. Cytokine production was assayed from supernatants of cells treated with various concentrations of imiquimod (0.5 μg/mL-4 μg/mL). Eight cytokines/chemokines were assayed using a mouse multiplex assay system (BioRad, Hercules, CA): IL-1β, IL-2, IL-6, IL-12p70, IFN-γ, TNF-α, and MIP-1α. Samples were run individually, but replicated in multiple experiments. Individual standards provided by the manufacturer were run on each plate. Data was collected using a BioRadLuminex-200 system, and analyzed using the BioRad Bio-Plex Manager Software (version 4.1).
Statistics
Differences in cytokine, mRNA expression, nitrite, and protein levels were evaluated by ANOVA. Differences were considered significant when p<0.05*, p<0.01**, or p<0.001***.
Results and Discussion
Ac-DEX microparticles exhibit low toxicity in vitro
Under acidic conditions, the pendant acetals on Ac-DEX hydrolyze to yield the parent water-soluble polymer, dextran (Figure 1A) (36). Microparticles were fabricated from Ac-DEX using a standard emulsion technique and appeared as shown in the SEM micrograph in Figure 1B. The imiquimod loading of the particles is 4 wt% and the loading efficiency is 100%. This loading efficiency is substantially higher compared to what can be obtained using other adjuvants such as CpG and poly (I:C) (30,31). Since imiquimod is hydrophobic it can easily be incorporated into the microparticles, and does not require complexation with a polycation. The average size of the particles as determined by dynamic light scattering was 233 nm ± 105 nm (Figure 1C), which has previously been shown to be in a suitable range for targeting phagocytic cells (e.g., dendritic cells) in vitro and in vivo (40,41).
Figure 1.
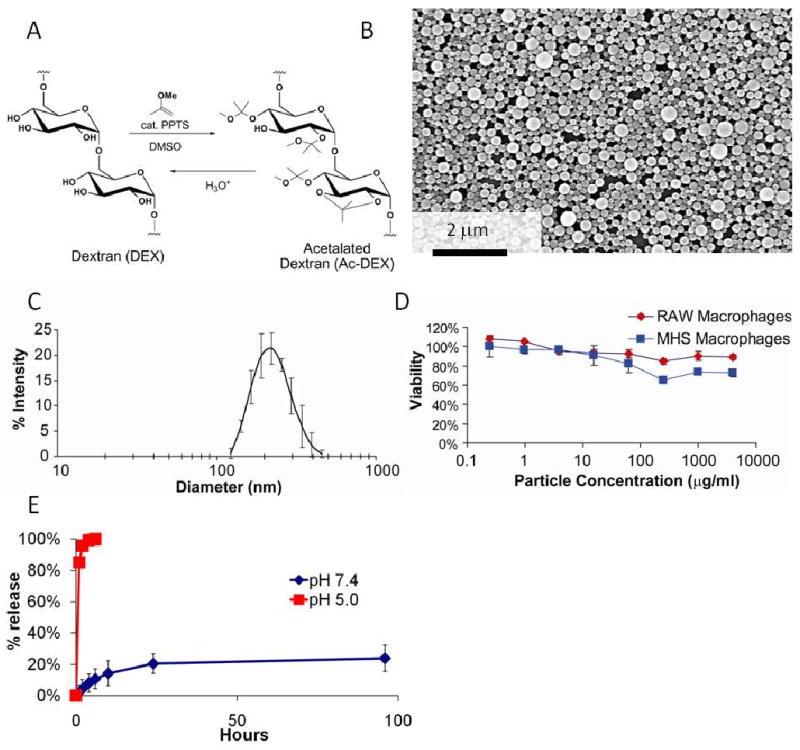
Ac-DEX microparticles are well tolerated by macrophages in vitro. (A) Synthesis of Ac-DEX. (B), SEM of Ac-DEX microparticles. (C), Size distribution of Ac-DEX particles, as measured by dynamic light scattering. (D) The cytotoxicity of empty Ac-DEX particles were determined by using the MTT assay with RAW 264.7 macrophages and MH-S alveolar macrophages. (E) Release of imiquimod from microparicles at pH 5.0 or 7.4.
To asses the biocompatibility of Ac-DEX microparticles, viability assays were performed with APCs. As shown in Figure 1D, empty Ac-DEX microparticles (0.25 μg/mL to 4 mg/mL) had little effect on the viability of both MH-S and RAW 264.7 macrophages, suggesting the particles were well tolerated even at high concentrations. Due to their pH-sensitive degradation properties, appropriate size, and negligible toxicity, Ac-DEX microparticles are a promising delivery platform for adjuvants and immunostimulatory ligands.
The pH sensitivity of the particles was tested by incubating imiquimod encapsulated in Ac-DEX in either a pH 5.0 or pH 7.4 buffer and measuring the release of imiquimod at the designated times (Figure 1E). At pH 5.0 the Ac-DEX particles quickly released the encapsulated imiquimod. This quick release should be beneficial for the activation of dendritic cells; upon internalization, Ac-DEX particles exposed to the low pH environment present in lysosomal compartments, release a bolus of imiquimod in proximity to TLR7/8. At pH 7.4 Ac-DEX microparticles released approximately 20% of the encapsulated drug within 48 hours, after which time the release rate leveled off. This initial burst release is very common with the encapsulation of drugs using single emulsion encapsulation techniques (42,43). With the use of Ac-DEX particles we could potentially decrease the amount of imiquimod used as an adjuvant in a potential vaccine application. Future experiments will be done to decrease the initial burst release of imiquimod in our microparticle carrier system.
Macrophage activation is enhanced through the encapsulation of imiquimod
Macrophages are a key component of the immune response, serving as both effectors during innate responses and antigen presenting cells during adaptive immune responses. Furthermore, alveolar macrophages provide a critical first line of protection during pulmonary exposure to pathogens, and due to the ability of Ac-DEX microparticles to potentially be delivered either as an aerosol or as an injection, it is important to evaluate immune responses induced by various types of macrophages. Therefore, we chose to examine the activity of imiquimod-loaded Ac-DEX particles in both MH-S alveolar macrophages and RAW 264.7 macrophages.
We hypothesized that the encapsulation of imiquimod in Ac-DEX microparticles would result in higher inflammatory cytokine production by macrophages because Ac-DEX particles are stable at physiological pH, and can release a bolus of imiquimod following internalization by APCs. We reasoned that endocytosis of the microparticles by macrophages, and eventual degradation of the particle in the low pH environment within phagolysosomes, would efficiently deliver imiquimod at a higher concentration than free imiquimod, resulting in higher TLR 7/8 activation. To explore our hypothesis, we assayed gene expression levels and protein concentrations of the inflammatory cytokines IL-1β, IL-6, and TNF-α in both macrophage cell lines after treatment with several concentrations of free or encapsulated imiquimod. MH-S macrophages demonstrated significantly increased gene expression for IL-1β, IL-6, and TNF-α at multiple dosages, with greater differences observed at the lowest concentrations (1 μg/mL and 0.5 μg/mL) for all three cytokines (p<0.001 for all cytokines and dosages with the exception of TNF-α at 1 μg/mL with a p<0.05) (Figure 2A). Similarly, protein production of IL-6 and TNF-α in MH-S macrophages exposed to encapsulated imiquimod was much higher compared to free imiquimod at lower concentrations. Interestingly, both free imiquimod and encapsulated imiquimod were capable of boosting IL-1β transcript and protein levels above baseline production in MH-S macrophages, which are known to constitutively secrete this cytokine (44) (Figure 2A). Protein concentrations of IL-1β were similar for both free and encapsulated imiquimod at doses greater than 1μg/mL. A difference was observed at the lowest concentration, where only encapsulated imiquimod was able to maintain elevated production. Similarly, IL-1β was upregulated in RAW 264.7 macrophages at both the transcript and protein level. Significant differences in IL-1β protein production were observed at the lowest concentrations of encapsulated imiquimod (p<0.01 for 1 μg/mL and p<0.001 for 0.5 μg/mL dosages) (Figure 2B). Notably, MH-S and RAW 264.7 macrophages treated with empty particles induced little to no production of cytokines at either the transcript or protein level. Furthermore, similar patterns were observed in measurements of IL-6 and TNF-α in both cell lines consistent with the concept that encapsulation of imiquimod provides significantly more efficient delivery of the TLR ligand. Overall, imiquimod encapsulated in Ac-DEX particles induced production of a relatively high amount of IL-1β, IL-6, and TNF-α at lower concentrations compared to free imiquimod. Based on this data, we predict that the encapsulation of imiquimod will allow dose sparing when performing treatments in vivo.
Figure 2.
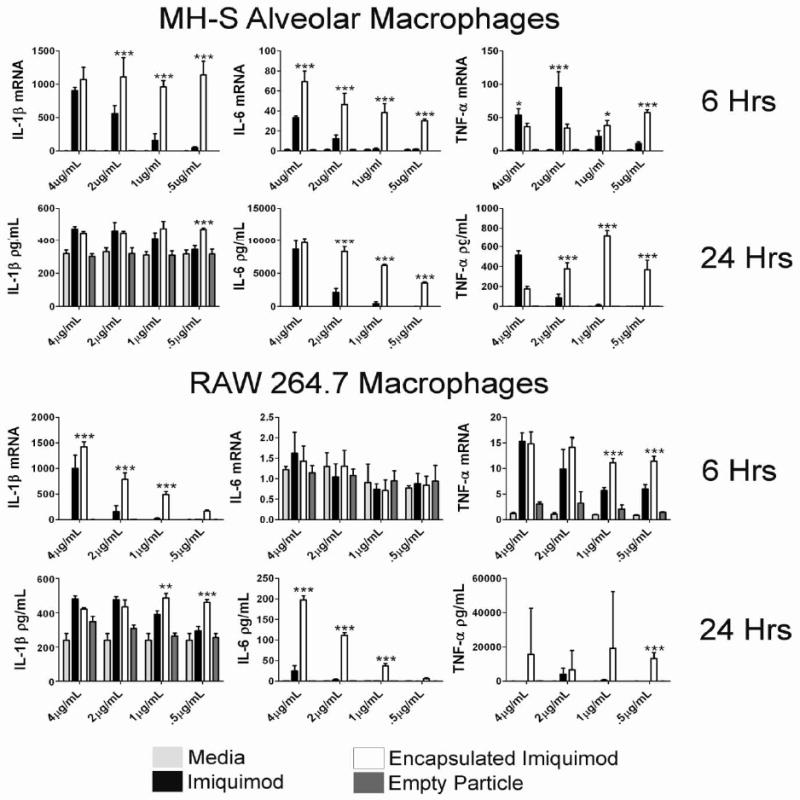
Encapsulation of imiquimod in Ac-DEX microparticles results in significantly higher inflammatory cytokine production in vitro. (A&B) 5×106 cells/mL MH-S and RAW 264.7 macrophages (N=3) were stimulated for 6 hours with medium, free imiquimod, encapsulated imiquimod, or empty particles, and prepared individually for real-time PCR analysis for IL-1β, IL-6, and TNF-α expression. Gene expression (mean ± SEM) is expressed as the fold increase over medium controls after normalization to HPRT. In separate experiments 1×106 cells/mL RAW 264.7 and MH-S (N=3) were stimulated with medium, free imiquimod, encapsulated imiquimod, or empty particles. After 24 hours the supernatants were removed and assayed for protein concentration of IL-1β, IL-6, and TNF-α via Luminex. Graphs are representative of at least two individual experiments. *=p value <0.05, **=p value <0.01, ***=p value <0.001
We also assessed the level of macrophage activation by examining the production of classical activation markers, including PD1-L1, iNOS, and the downstream product of iNOS activity, nitric oxide. Prior research has shown that nitric oxide production in macrophages is crucial for clearance of intracellular bacteria (45). As shown in Figure 3A, encapsulation of imiquimod resulted in significantly higher iNOS expression (p<0.001 for all dosages), compared to free imiquimod, when tested in both MH-S and RAW 264.7 macrophages. In addition, as shown in Figure 3B, nitric oxide (NO) production was significantly upregulated in encapsulated versus free imiquimod (p<0.001) at all concentrations in both MH-S and RAW 264.7 macrophages after 24 hours. Imiquimod encapsulated in Ac-Dex particles led to significantly higher levels of PD1-L1 than free imiquimod in MH-S macrophages at the three lowest concentrations (minimum of p<0.01 for all dosages), while RAW 264.7 macrophages yielded significantly higher PD1-L1 expression at the three highest concentrations (p<0.001).
Figure 3.
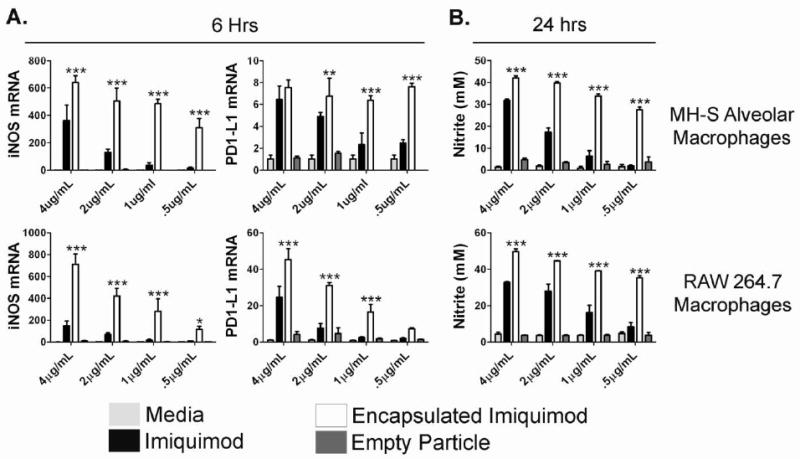
Encapsulation of imiquimod in Ac-DEX microparticles increases macrophage activation in vitro. (A) 5×106/mL RAW 264.7 and MH-S macrophages (N=3) were stimulated for 6 hours with medium, free imiquimod, encapsulated imiquimod, or empty particles, and prepared individually for real-time PCR analysis for iNOS and PD1-L1 expression. Gene expression (mean ± SEM) is expressed as the fold increase over medium controls after normalization to hypoxanthine guanine phosphoribosyl transferase HPRT. (B) 1×106/mL RAW 264.7 and MH-S (N=3) were stimulated with medium, free imiquimod, encapsulated imiquimod, or empty particles. After 24 hours, the supernatants were removed and the presence of nitrite was quantified by the Griess assay. Graphs are representative of at least two individual experiments. *=p value <0.05, **=p value <0.01, ***=p value <0.001
Examination of these two markers suggests that imiquimod is capable of inducing macrophage activation on its own, but that this activation can be significantly augmented by the encapsulation of the TLR7/8 ligand in an acid-sensitive vehicle. Furthermore, when we measured the responses for these markers with cells that had been treated with empty particles we observed no significant increases in any of the cytokines or macrophage activation markers. These data suggest that these particles are not stimulatory on their own, and that the changes in activation status are due to their ability to deliver an immunostimulatory payload. Taken together, this data clearly indicates that the encapsulation of imiquimod in Ac-DEX particles leads to significantly increased cytokine production by macrophages in vitro.
Encapsulation of imiquimod significantly increases cytokine and chemokine production in bone marrow derived dendritic cells
While macrophages are capable of antigen presentation, they are most typically noted for their effector abilities. In contrast, myeloid dendritic cells are widely considered to be professional antigen presenting cells, and as such, we wanted to assess the ability of Ac-DEX particles to effectively deliver immunostimulatory ligands to this cell type. Therefore, bone marrow-derived dendritic cells (BMDCs) were used to assess the activity of imiquimod encapsulated in Ac-DEX particles. Imiquimod concentrations were similar to those analyzed in previous experiments (i.e. serial dilutions from 4 μg/mL to 0.5 μg/mL). Supernatants from cell cultures were harvested and assayed for the production of protein using a multiplexed bead assay. As shown in Figure 4, BMDCs exposed to encapsulated imiquimod yielded significantly higher levels of IL-1β and IL-6 at the two highest concentrations (p<0.001), as compared to similar amounts of free imiquimod. Similarly, we observed significant increases in the amounts of IL-12p70 (p<0.01 or greater for all dosages) and MIP-1α (p<0.001 for 4 μg/mL and 2 μg/mL dosages) in encapsulated imiquimod groups as opposed to non-encapsulated groups.
Figure 4.
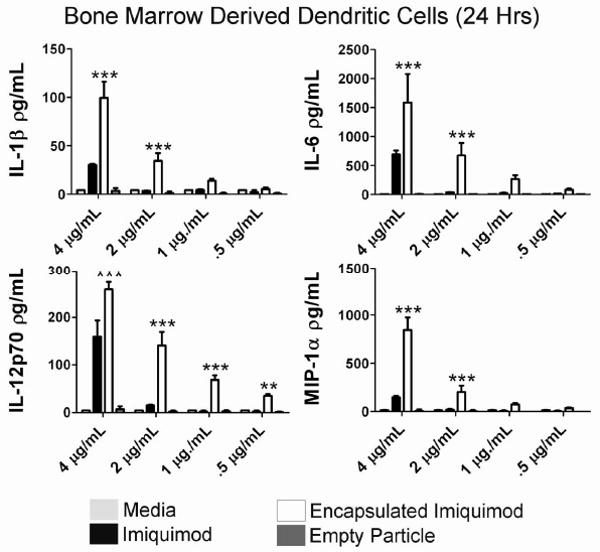
Microencapsulation of imiquimod using Ac-DEX microspheres results in higher BMDC activation. BMDCs (N=3) were stimulated for 24 hours with medium, free imiquimod, encapsulated imiquimod, or empty particles. Supernatants were collected assayed for protein concentration of IL-1β, IL-6, IL-12p70, and MIP-1α using a multiplexed assay. Graphs are representative of at least two individual experiments. **=p value <0.01, ***=p value <0.001
Previous work has shown that dendritic cells treated with free imiquimod will produce the inflammatory cytokines IL-1β and IL-6 and IL-12p70 (46). To the best of our knowledge this is the first published report showing that encapsulated imiquimod drastically increases the production of inflammatory cytokines in dendritic cells and macrophages compared to free imiquimod. As stated previously, the incorporation of imiquimod is relatively easy compared to the methods required to encapsulate other adjuvants such as CpG and poly (I:C). The efficacy of encapsulating imiquimod is illustrated by the upregulation of IL-12p70 in encapsulated imiquimod treated DCs. IL-12p70 upregulation is beneficial in initiating an immune response since it is known to induce IFN-γ production from NK and NKT cells, and induces the differentiation of Th1 cells (47). In addition, the increase in the production of MIP-1α by dendritic cells treated with encapsulated imiquimod suggests an enhanced ability to attract T cells and subsequently establishing a cytotoxic or adaptive immune response. Taken together, these results indicate that the encapsulation of imiquimod in acid-sensitive microparticles leads to enhanced TLR 7/8 signaling, results in a greater effect on dendritic cell activation and has the potential to drastically enhance the efficacy of in vivo vaccine applications. Prior work has shown that the imidazoquinoline 3M-019, when encapsulated in a liposome, will increase the production of antibodies compared to the free imidazoquinoline (48). In addition, Heit et al. (49) has shown that the encapsulation of imiquimod drastically enhances CD8 T cell presentation in vitro. With this prior research, and our data showing an increase in DC activation, the encapsulation of small molecule TLR agonists has the potential of increasing the efficacy of subunit vaccines.
Conclusions
The implications of our results are broad in their applicability. First, we have shown that encapsulation of the FDA approved imiquimod in acid-degradable particles can significantly enhance the in vitro activation of macrophages and dendritic cells at levels which exceed treatment with free imiquimod alone. These data set the stage for targeted co-delivery of vaccine constructs along with a potent adjuvant to intracellular compartments, thus reducing the amount of vaccine and adjuvant necessary to induce protection. Second, we have illustrated that dose sparing occurs with the encapsulation of imiquimod. Through encapsulation, we can reduce the amount of imiquimod required and potentially alleviate side effects associated with its use. Third, the methods required to encapsulate imiquimod are relatively easy compared to the methods required to encapsulate other adjuvants. Based on the ability of Ac-DEX microparticles to effectively deliver imiquimod to macrophages and dendritic cells in vitro, we feel this is a successful platform that can be partnered with DNA and protein based vaccine candidates for future in vivo testing.
Acknowledgments
This work was supported by the Defense Threat Reduction Agency program ZZ0020_06_NM_B. The development of Ac-Dex bead carriers was supported by the National Institutes of Health (NBIB grant R01-EB005824).
I am a military service member (or employee of the U.S. Government). This work was prepared as part of my official duties. Title 17 U.S.C. §105 provides that ‘Copyright protection under this title is not available for any work of the United States Government.’ Title 17 U.S.C. §101 defines a U.S. Government work as a work prepared by a military service member or employee of the U.S. Government as part of that person's official duties.
Contributor Information
Eric M. Bachelder, Department of Chemistry, University of California, Berkeley, CA 94720-1460.
Tristan T. Beaudette, Department of Chemistry, University of California, Berkeley, CA 94720-1460
Kyle E. Broaders, Department of Chemistry, University of California, Berkeley, CA 94720-1460
Jean MJ. Fréchet, Department of Chemistry, University of California, Berkeley, CA 94720-1460
Mark T. Albrecht, Vaccines and Medical Countermeasures Group, Biological Defense Research Directorate, Navy Medical Research Center, 12300 Washington Avenue, Rockville, MD 20852
Alfred J. Mateczun, Vaccines and Medical Countermeasures Group, Biological Defense Research Directorate, Navy Medical Research Center, 12300 Washington Avenue, Rockville, MD 20852
Kristy M. Ainslie, Vaccines and Medical Countermeasures Group, Biological Defense Research Directorate, Navy Medical Research Center, 12300 Washington Avenue, Rockville, MD 20852.
John T. Pesce, Vaccines and Medical Countermeasures Group, Biological Defense Research Directorate, Navy Medical Research Center, 12300 Washington Avenue, Rockville, MD 20852.
Andrea M. Keane-Myers, Vaccines and Medical Countermeasures Group, Biological Defense Research Directorate, Navy Medical Research Center, 12300 Washington Avenue, Rockville, MD 20852
References
- 1.Minor P. Vaccine-derived poliovirus (VDPV): Impact on poliomyelitis eradication. Vaccine. 2009;27(20):2649–52. doi: 10.1016/j.vaccine.2009.02.071. [DOI] [PubMed] [Google Scholar]
- 2.Artenstein AW, Grabenstein JD. Smallpox vaccines for biodefense: need and feasibility. Expert Rev Vaccines. 2008;7(8):1225–37. doi: 10.1586/14760584.7.8.1225. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Garulli B, Castrucci MR. Protective immunity to influenza: lessons from the virus for successful vaccine design. Expert Rev Vaccines. 2009;8(6):689–93. doi: 10.1586/erv.09.35. [DOI] [PubMed] [Google Scholar]
- 4.Foster JL, Garcia JV. HIV-1 Nef: at the crossroads. Retrovirology. 2008;5:84. doi: 10.1186/1742-4690-5-84. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Kovacs-Nolan J, Latimer L, Landi A, Jenssen H, Hancock RE, Babiuk LA, van Drunen Littel-van den Hurk S. The novel adjuvant combination of CpG ODN, indolicidin and polyphosphazene induces potent antibody- and cell-mediated immune responses in mice. Vaccine. 2009;27(14):2055–64. doi: 10.1016/j.vaccine.2009.01.118. [DOI] [PubMed] [Google Scholar]
- 6.Burke B, Gomez-Roman VR, Lian Y, Sun Y, Kan E, Ulmer J, Srivastava IK, Barnett SW. Neutralizing antibody responses to subtype B and C adjuvanted HIV envelope protein vaccination in rabbits. Virology. 2009;387(1):147–56. doi: 10.1016/j.virol.2009.02.005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Medzhitov R, Janeway CA., Jr Innate immunity: the virtues of a nonclonal system of recognition. Cell. 1997;91(3):295–8. doi: 10.1016/s0092-8674(00)80412-2. [DOI] [PubMed] [Google Scholar]
- 8.Mitchell JA, Fitzgerald KA, Coyle A, Silverman N, Cartwright N. TOLLing away in Brazil. Nat Immunol. 2006;7(7):675–9. doi: 10.1038/ni0706-675. [DOI] [PubMed] [Google Scholar]
- 9.Othoro C, Johnston D, Lee R, Soverow J, Bystryn JC, Nardin E. Enhanced immunogenicity of Plasmodium falciparum peptide vaccines using a topical adjuvant containing a potent synthetic Toll-like receptor 7 agonist, imiquimod. Infect Immun. 2009;77(2):739–48. doi: 10.1128/IAI.00974-08. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Gollnick H, Barasso R, Jappe U, Ward K, Eul A, Carey-Yard M, Milde K. Safety and efficacy of imiquimod 5% cream in the treatment of penile genital warts in uncircumcised men when applied three times weekly or once per day. Int J STD AIDS. 2001;12(1):22–8. [PubMed] [Google Scholar]
- 11.Stockfleth E, Meyer T, Benninghoff B, Christophers E. Successful treatment of actinic keratosis with imiquimod cream 5%: a report of six cases. Br J Dermatol. 2001;144(5):1050–3. doi: 10.1046/j.1365-2133.2001.04197.x. [DOI] [PubMed] [Google Scholar]
- 12.Lebwohl M, Dinehart S, Whiting D, Lee PK, Tawfik N, Jorizzo J, Lee JH, Fox TL. Imiquimod 5% cream for the treatment of actinic keratosis: results from two phase III, randomized, double-blind, parallel group, vehicle-controlled trials. J Am Acad Dermatol. 2004;50(5):714–21. doi: 10.1016/j.jaad.2003.12.010. [DOI] [PubMed] [Google Scholar]
- 13.Sterry W, Ruzicka T, Herrera E, Takwale A, Bichel J, Andres K, Ding L, Thissen MR. Imiquimod 5% cream for the treatment of superficial and nodular basal cell carcinoma: randomized studies comparing low-frequency dosing with and without occlusion. Br J Dermatol. 2002;147(6):1227–36. doi: 10.1046/j.1365-2133.2002.05069.x. [DOI] [PubMed] [Google Scholar]
- 14.Bath-Hextall F, Bong J, Perkins W, Williams H. Interventions for basal cell carcinoma of the skin: systematic review. Bmj. 2004;329(7468):705. doi: 10.1136/bmj.38219.515266.AE. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Bath-Hextall FJ, Perkins W, Bong J, Williams HC. Interventions for basal cell carcinoma of the skin. Cochrane Database Syst Rev. 2007;(1):CD003412. doi: 10.1002/14651858.CD003412.pub2. [DOI] [PubMed] [Google Scholar]
- 16.Fleming CJ, Bryden AM, Evans A, Dawe RS, Ibbotson SH. A pilot study of treatment of lentigo maligna with 5% imiquimod cream. Br J Dermatol. 2004;151(2):485–8. doi: 10.1111/j.1365-2133.2004.05983.x. [DOI] [PubMed] [Google Scholar]
- 17.Hassan F, Islam S, Tumurkhuu G, Dagvadorj J, Naiki Y, Komatsu T, Koide N, Yoshida T, Yokochi T. Involvement of interleukin-1 receptor-associated kinase (IRAK)-M in toll-like receptor (TLR) 7-mediated tolerance in RAW 264.7 macrophage-like cells. Cell Immunol. 2009;256(1-2):99–103. doi: 10.1016/j.cellimm.2009.01.013. [DOI] [PubMed] [Google Scholar]
- 18.Doxsee CL, Riter TR, Reiter MJ, Gibson SJ, Vasilakos JP, Kedl RM. The immune response modifier and Toll-like receptor 7 agonist S-27609 selectively induces IL-12 and TNF-alpha production in CD11c+CD11b+CD8- dendritic cells. J Immunol. 2003;171(3):1156–63. doi: 10.4049/jimmunol.171.3.1156. [DOI] [PubMed] [Google Scholar]
- 19.Gerster JF, Lindstrom KJ, Miller RL, Tomai MA, Birmachu W, Bomersine SN, Gibson SJ, Imbertson LM, Jacobson JR, Knafla RT, Maye PV, Nikolaides N, Oneyemi FY, Parkhurst GJ, Pecore SE, Reiter MJ, Scribner LS, Testerman TL, Thompson NJ, Wagner TL, Weeks CE, Andre JD, Lagain D, Bastard Y, Lupu M. Synthesis and structure-activity-relationships of 1H-imidazo [4,5-c]quinolines that induce interferon production. J Med Chem. 2005;48(10):3481–91. doi: 10.1021/jm049211v. [DOI] [PubMed] [Google Scholar]
- 20.Stanley MA. Imiquimod and the imidazoquinolones: mechanism of action and therapeutic potential. Clin Exp Dermatol. 2002;27(7):571–7. doi: 10.1046/j.1365-2230.2002.01151.x. [DOI] [PubMed] [Google Scholar]
- 21.Chollet JL, Jozwiakowski MJ, Phares KR, Reiter MJ, Roddy PJ, Schultz HJ, Ta QV, Tomai MA. Development of a topically active imiquimod formulation. Pharm Dev Technol. 1999;4(1):35–43. doi: 10.1080/10837459908984222. [DOI] [PubMed] [Google Scholar]
- 22.Schon MP, Schon M. TLR7 and TLR8 as targets in cancer therapy. Oncogene. 2008;27(2):190–9. doi: 10.1038/sj.onc.1210913. [DOI] [PubMed] [Google Scholar]
- 23.Standley SM, Mende I, Goh SL, Kwon YJ, Beaudette TT, Engleman EG, Frechet JM. Incorporation of CpG oligonucleotide ligand into protein-loaded particle vaccines promotes antigen-specific CD8 T-cell immunity. Bioconjug Chem. 2007;18(1):77–83. doi: 10.1021/bc060165i. [DOI] [PubMed] [Google Scholar]
- 24.Kaiser-Schulz G, Heit A, Quintanilla-Martinez L, Hammerschmidt F, Hess S, Jennen L, Rezaei H, Wagner H, Schatzl HM. Polylactide-coglycolide microspheres co-encapsulating recombinant tandem prion protein with CpG-oligonucleotide break self-tolerance to prion protein in wild-type mice and induce CD4 and CD8 T cell responses. J Immunol. 2007;179(5):2797–807. doi: 10.4049/jimmunol.179.5.2797. [DOI] [PubMed] [Google Scholar]
- 25.Wack A, Baudner BC, Hilbert AK, Manini I, Nuti S, Tavarini S, Scheffczik H, Ugozzoli M, Singh M, Kazzaz J, Montomoli E, Del Giudice G, Rappuoli R, O'Hagan DT. Combination adjuvants for the induction of potent, long-lasting antibody and T-cell responses to influenza vaccine in mice. Vaccine. 2008;26(4):552–61. doi: 10.1016/j.vaccine.2007.11.054. [DOI] [PubMed] [Google Scholar]
- 26.Heffernan MJ, Kasturi SP, Yang SC, Pulendran B, Murthy N. The stimulation of CD8+ T cells by dendritic cells pulsed with polyketal microparticles containing ion-paired protein antigen and poly(inosinic acid)-poly(cytidylic acid) Biomaterials. 2009;30(5):910–8. doi: 10.1016/j.biomaterials.2008.10.034. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Xiang SD, Scholzen A, Minigo G, David C, Apostolopoulos V, Mottram PL, Plebanski M. Pathogen recognition and development of particulate vaccines: does size matter? Methods. 2006;40(1):1–9. doi: 10.1016/j.ymeth.2006.05.016. [DOI] [PubMed] [Google Scholar]
- 28.Barton GM, Kagan JC. A cell biological view of Toll-like receptor function: regulation through compartmentalization. Nat Rev Immunol. 2009;9(8):535–42. doi: 10.1038/nri2587. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Mancuso G, Gambuzza M, Midiri A, Biondo C, Papasergi S, Akira S, Teti G, Beninati C. Bacterial recognition by TLR7 in the lysosomes of conventional dendritic cells. Nat Immunol. 2009;10(6):587–94. doi: 10.1038/ni.1733. [DOI] [PubMed] [Google Scholar]
- 30.Fischer S, Schlosser E, Mueller M, Csaba N, Merkle HP, Groettrup M, Gander B. Concomitant delivery of a CTL-restricted peptide antigen and CpG ODN by PLGA microparticles induces cellular immune response. J Drug Target. 2009;17(8):652–61. doi: 10.1080/10611860903119656. [DOI] [PubMed] [Google Scholar]
- 31.San Roman B, Irache JM, Gomez S, Tsapis N, Gamazo C, Espuelas MS. Co-encapsulation of an antigen and CpG oligonucleotides into PLGA microparticles by TROMS technology. Eur J Pharm Biopharm. 2008;70(1):98–108. doi: 10.1016/j.ejpb.2008.03.015. [DOI] [PubMed] [Google Scholar]
- 32.Ji J, Rosenzweig N, Griffin C, Rosenzweig Z. Synthesis and application of submicrometer fluorescence sensing particles for lysosomal pH measurements in murine macrophages. Anal Chem. 2000;72(15):3497–503. doi: 10.1021/ac000080p. [DOI] [PubMed] [Google Scholar]
- 33.Haining WN, Anderson DG, Little SR, von Bergwelt-Baildon MS, Cardoso AA, Alves P, Kosmatopoulos K, Nadler LM, Langer R, Kohane DS. pH-triggered microparticles for peptide vaccination. J Immunol. 2004;173(4):2578–85. doi: 10.4049/jimmunol.173.4.2578. [DOI] [PubMed] [Google Scholar]
- 34.Houde M, Bertholet S, Gagnon E, Brunet S, Goyette G, Laplante A, Princiotta MF, Thibault P, Sacks D, Desjardins M. Phagosomes are competent organelles for antigen cross-presentation. Nature. 2003;425(6956):402–6. doi: 10.1038/nature01912. [DOI] [PubMed] [Google Scholar]
- 35.Mallapragada SK, Narasimhan B. Immunomodulatory biomaterials. Int J Pharm. 2008;364(2):265–71. doi: 10.1016/j.ijpharm.2008.06.030. [DOI] [PubMed] [Google Scholar]
- 36.Bachelder EM, Beaudette TT, Broaders KE, Dashe J, Frechet JM. Acetal-derivatized dextran: an acid-responsive biodegradable material for therapeutic applications. J Am Chem Soc. 2008;130(32):10494–5. doi: 10.1021/ja803947s. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Broaders KE, Cohen JA, Beaudette TT, Bachelder EM, Frechet JM. Acetalated dextran is a chemically and biologically tunable material for particulate immunotherapy. Proc Natl Acad Sci U S A. 2009;106(14):5497–502. doi: 10.1073/pnas.0901592106. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Bachelder EM, Beaudette TT, Broaders KE, Paramonov SE, Dashe J, Frechet JM. Acid-degradable polyurethane particles for protein-based vaccines: biological evaluation and in vitro analysis of particle degradation products. Mol Pharm. 2008;5(5):876–84. doi: 10.1021/mp800068x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Pesce J, Kaviratne M, Ramalingam TR, Thompson RW, Urban JF, Jr, Cheever AW, Young DA, Collins M, Grusby MJ, Wynn TA. The IL-21 receptor augments Th2 effector function and alternative macrophage activation. J Clin Invest. 2006;116(7):2044–55. doi: 10.1172/JCI27727. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Foged C, Brodin B, Frokjaer S, Sundblad A. Particle size and surface charge affect particle uptake by human dendritic cells in an in vitro model. Int J Pharm. 2005;298(2):315–22. doi: 10.1016/j.ijpharm.2005.03.035. [DOI] [PubMed] [Google Scholar]
- 41.Hirota K, Hasegawa T, Hinata H, Ito F, Inagawa H, Kochi C, Soma G, Makino K, Terada H. Optimum conditions for efficient phagocytosis of rifampicin-loaded PLGA microspheres by alveolar macrophages. J Control Release. 2007;119(1):69–76. doi: 10.1016/j.jconrel.2007.01.013. [DOI] [PubMed] [Google Scholar]
- 42.Allison SD. Analysis of initial burst in PLGA microparticles. Expert Opin Drug Deliv. 2008;5(6):615–28. doi: 10.1517/17425247.5.6.615. [DOI] [PubMed] [Google Scholar]
- 43.Yeo Y, Park K. Control of encapsulation efficiency and initial burst in polymeric microparticle systems. Arch Pharm Res. 2004;27(1):1–12. doi: 10.1007/BF02980037. [DOI] [PubMed] [Google Scholar]
- 44.Mbawuike IN, Herscowitz HB. MH-S, a murine alveolar macrophage cell line: morphological, cytochemical, and functional characteristics. J Leukoc Biol. 1989;46(2):119–27. doi: 10.1002/jlb.46.2.119. [DOI] [PubMed] [Google Scholar]
- 45.Shimada K, Chen S, Dempsey PW, Sorrentino R, Alsabeh R, Slepenkin AV, Peterson E, Doherty TM, Underhill D, Crother TR, Arditi M. The NOD/RIP2 pathway is essential for host defenses against Chlamydophila pneumoniae lung infection. PLoS Pathog. 2009;5(4):e1000379. doi: 10.1371/journal.ppat.1000379. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Testerman TL, Gerster JF, Imbertson LM, Reiter MJ, Miller RL, Gibson SJ, Wagner TL, Tomai MA. Cytokine induction by the immunomodulators imiquimod and S-27609. J Leukoc Biol. 1995;58(3):365–72. doi: 10.1002/jlb.58.3.365. [DOI] [PubMed] [Google Scholar]
- 47.Trinchieri G. Interleukin-12 and the regulation of innate resistance and adaptive immunity. Nat Rev Immunol. 2003;3(2):133–46. doi: 10.1038/nri1001. [DOI] [PubMed] [Google Scholar]
- 48.Johnston D, Zaidi B, Bystryn JC. TLR7 imidazoquinoline ligand 3M-019 is a potent adjuvant for pure protein prototype vaccines. Cancer Immunol Immunother. 2007;56(8):1133–41. doi: 10.1007/s00262-006-0262-3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Heit A, Schmitz F, Haas T, Busch DH, Wagner H. Antigen co-encapsulated with adjuvants efficiently drive protective T cell immunity. Eur J Immunol. 2007;37(8):2063–74. doi: 10.1002/eji.200737169. [DOI] [PubMed] [Google Scholar]