

Abstract
Emotional resilience can be defined as the ability to maintain healthy and stable levels of psychological functioning in the wake of stress and trauma. Although genes that contribute to psychopathology (often in interaction with environmental stressors) are being detected with increasing consistency, genes that influence resilience to stress have been less studied. In this study, 423 undergraduate college students completed a psychometrically sound 10-item self-report measure of resilience (CDRISC-10) and provided blood for DNA. Linear and logistic regression analyses were used to model relationships between the serotonin transporter promoter polymorphism (5HTTLPR) and CDRISC-10 scores and categories, respectively. CDRISC–10 scores were normally distributed (mean 26.17 SD 5.88 [range 5 – 40]). In models adjusting for ancestry proportion scores (to mitigate confounding by population stratification) and other covariates, each copy of the “s” allele of 5HTTLPR was associated with ~1-point lower CDRISC-10 score. Each copy of the “s” allele was associated with increased (adjusted OR = 1.53, 95% CI 1.06-2.21, p = 0.024) odds of being in the low resilient category (> 1 SD below the mean), compared to being homozygous for the “l” allele. These findings suggest that variation in 5HTTLPR is associated with individual differences in emotional resilience, defined as an individual's ability to withstand and bounce back from stress. This relationship may explain the frequently observed interaction between 5HTTLPR and life stressors in predicting adverse mental health outcomes (e.g., depressive symptoms). Replication is needed, in concert with identification of other genes that influence emotional resilience and related phenotypes.
Keywords: emotional resilience, stress, depression, 5HTTLPR, serotonin, gene
INTRODUCTION
Resilience can be defined as an individual's ability to thrive despite the experience of stress or trauma (Luthar et al. 2000;Connor and Davidson 2003;Charney 2004). As such, emotional resilience can be considered a trait that confers protection against the development of mental disorders in the face of adversity (Bonanno 2004). This psychological hardiness may emanate from constitutional variables like adaptive personality traits as well as specific learned behaviors such as problem-focused coping (Tsuang 2000;Campbell-Sills et al. 2006). Resilience as a personal characteristic is distinct from, but likely influenced by, external variables (e.g., adequate social support, good schools) that reduce risk for stress-related mental disorders by serving as “buffers” to stress (Luthar et al. 2000).
Recovery and adaptation in the face of stress is a common part of human experience, and despite exposure to many traumatic events in their lifetimes (Kessler et al. 1995)— even extreme events such as terrorism (Galea et al. 2003) or natural catastrophes such as hurricanes (Galea et al. 2007) – most individuals do not go on to develop psychopathology. Empirical evidence suggests that the determinants of resilience are complex, with social (King et al. 1999), psychological (Tugade and Fredrickson 2004;Campbell-Sills et al. 2006), and biological (Charney 2004) factors all believed to play a role. In terms of the latter, it is likely that genetic factors influence resilience to stress (Caspi et al. 2003;Kendler 2005). Preclinical studies indicate that functional variation in certain genes can influence coping in the face of stress (Hariri and Holmes 2006). For example, abnormalities in stress-coping – including depressive-like behaviors – have been observed in serotonin transporter (5HTT) knock-out mice, suggesting that reduced resilience to stress is a behavioral consequence of loss of 5HTT gene function (Wellman et al. 2007).
Although no specific genetic factors for emotional resilience have yet been identified in humans, a large body of research points to a functional length polymorphism in the promoter of the serotonin transporter gene (gene, SLC6A4; variant, 5HTTLPR) as a prime candidate. This polymorphism has functional importance in that the “short” (S) variant is less transcriptionally efficient than the “long” (L) variant (Lesch et al. 1996). The 5HTTLPR polymorphism has been shown in many (Caspi et al. 2003;Eley et al. 2004;Grabe et al. 2005;Kaufman et al. 2006;Kaufman et al. 2004;Kendler et al. 2005;Zalsman et al. 2006;Wilhelm et al. 2006;Jacobs et al. 2006), but not all (Surtees et al. 2006;Gillespie et al. 2005) studies to interact with stressful life events (though in one study, the interaction was with the purported low-risk allele (Chorbov et al. 2007)) to increase susceptibility to major depression or depressive symptoms. We have also shown that this polymorphism – when interacting with childhood stressors – contributes to variance in anxiety sensitivity, a risk factor for panic and other anxiety disorders (Stein et al. 2008), and risk for early alcohol use (Kaufman et al. 2007).
Several meta-analyses have also concluded that there is a small but significant association between 5HTTLPR and neuroticism (Schinka et al. 2004;Sen et al. 2004), a known personality risk factor for affective illness. It is therefore possible that there are direct effects of 5HTTLPR on risk for mood and anxiety disorders that do not necessarily operate through its effect on stress resilience. But the confluence of findings is consistent with the notion that 5HTTLPR operates pleiotropically to moderate or buffer emotional responsivity to stress. Following this line of reasoning, we hypothesized that variation in 5HTTLPR is associated directly (i.e., rather than through a requisite interaction with life stress) and specifically (i.e., more than with related phenotypes such as neuroticism or state depression) with the construct of emotional resilience – an individual's capacity to handle or rebound from stressful experience.
METHODS
Sample and Procedures
Participants (N = 423) were an unselected sample of 318 (75%) female and 105 male (mean age 18.83 sd 1.67 years; median age 19 years) undergraduate psychology students at San Diego State University (SDSU) who completed self-report questionnaires about personality characteristics in return for course credit. Self-reported ethnicity was predominantly European American (N = 260; 61.5%), Hispanic (N = 68; 16.1%), Asian American (N = 53; 12.5%), or miscellaneous others. Subjects each came for a scheduled appointment at which a blood sample (60 ml) was drawn for genetic studies and additional questionnaires were completed. Subjects gave informed, written consent to participate in the study, which was approved by the Human Research Protection Programs at both SDSU and University of California San Diego. Subjects received $25 for providing the blood sample.
Questionnaires
Although some subjects participated in subsequent studies for which a psychiatric diagnostic interview was administered, diagnostic information was not available for the vast majority of subjects and was therefore not considered here. All subjects completed a panel of self-report measures relevant to the study of personality and anxiety-related traits. Measures that were examined for this report are:
Connor-Davidson Resilience Scale, 10-item version (CDRISC-10), a measure of emotional resilience that has been well-studied in general population and patient samples (Connor and Davidson 2003). In the present study we used the 10-item version of the CDRISC, which is unidimensional and displays good internal consistency, convergent validity, and discriminant validity (Campbell-Sills and Stein 2007). In a sample of 439 undergraduates which partially overlapped with the present sample, the correlation of the 10-item score with the 25-item score was r = 0.70. Women score lower on average than men on the CDRISC-10, as do individuals with lower education (Campbell-Sills L, Stein MB, Forde DR, submitted). Sample items on the CDRISC-10 are “I am able to adapt when changes occurs” and “Under pressure, I stay focused and think clearly”. Range of the CDRISC-10 is 0-40, with each of the 10 items scored 0 “Not True at All (of Me)” through 4 “True Nearly All of the Time (of Me)”.
NEO-Personality Inventory-Revised (NEO-FFI), a widely used, 60-item self-report measure of personality (Costa, Jr. and McCrae 1992). We limited analyses to the relationship of 5HTTLPR to neuroticism – a well established, non-specific risk factor for anxiety and mood disorders (Middeldorp et al. 2005). T scores were calculated directly from college-age, sex-specific norms. CDRISC-10 scores correlate moderately with NEO neuroticism (NEO-N) scores (r = −0.65) (Campbell-Sills and Stein 2007)
Beck Depression Inventory (BDI), a well established, 21-item self-report measure of depressive symptoms (Beck et al. 1961).
Childhood Trauma Questionnaire (CTQ), a 28-item retrospective self-report questionnaire designed to assess five types of negative childhood experiences (Bernstein et al. 1994;Scher et al. 2001). We limited analyses to the Emotional Abuse (CTQ-EA) component of this instrument which, in our hands, has consistently shown evidence of interaction with 5HTTLPR to explain variance in psychopathology (Stein et al. 2008).
Genotype Analysis
Genomic DNA was extracted from whole blood and the 5HTTLPR variant was analyzed by polymerase chain reaction (PCR) amplification followed by size fractionation, as described elsewhere for the diallelic L-S classification (Gelernter et al. 1997). We also genotyped an a/g single nucleotide polymorphism (SNP; rs25531) in the variable region that defines the variable number of tandem repeats (VNTR) S/L polymorphism divides L alleles of 5-HTTLPR into two functionally different variants, La and Lg (Hu et al. 2005;Wendland et al. 2006), the latter of which has been reported to behave equivalently to the S allele in terms of its low transcriptional efficiency. For quality control, 8% of genotypes were repeated, with no discrepancies.
Ancestral Proportion Scores
Subjects' ancestries were estimated using a set of unlinked genetic markers by Bayesian cluster analysis, using the procedures and software developed by Pritchard and colleagues (http://pritch.bsd.uchicago.edu/software.html) (Falush et al. 2003;Pritchard and Rosenberg 1999;Pritchard et al. 2000). Pritchard's software program STRUCTURE implements Bayesian cluster modeling that can recognize cryptic population genetic patterns without prior information of population origins. Data were submitted to STRUCTURE using 35 markers with models specified as “admixture” and “allele frequencies correlated” and 500,000 burn-in and 500,000 MCMC (Markov Chain Monte Carlo) iterations. The markers were the STRs described previously (Yang et al. 2005).
A two-cluster solution fit the data best and classified 87.3% of self-identified European American (EA) subjects (N = 260) in this sample in concordance with their self-reported ethnicity. Within the EA sample, distribution of 5HTTLPR alleles was L 55.0% and S 45.0%, and genotype observations were consistent with no significant deviation from HWE expectations (chi-square = 2.53, df = 1, p = 0.11).
Statistical Analysis
Analyses were conducted using the biostatistical analysis program STATA (Version 10.0) (StataCorp 2007). Linear regression techniques in STATA (“rreg”) (StataCorp 2007) that are considered robust to outliers were used to model the relationship between 5HTTLPR genotype (coded as 0 for LL, 1 for LS, and 2 for SS) and CDRISC-10 scores. In a series of sensitivity analyses (see Results), linear regressions were rerun using covariates such as sex and ancestral population scores (modeled using two ancestral coefficients). Parallel sets of analyses were conducted using 5HTTLPR genotype to predict BDI and NEO-N scores, respectively, to test the hypothesis that the relationship between 5HTTLPR and CDRISC-10 was specific.
We repeated analyses using the 5HTTLPR triallelic genotypes (we found no instances of Sg alleles). They were reclassified by their level of expression as follows: Lg/S, Lg/Lg, and S/S were reclassified as S′S′, La/S and La/Lg were reclassified as L′S′, and La/La was reclassified as L′L′ (Parsey et al. 2006).
Analyses were not corrected for multiple testing for the following reasons: We were testing only one hypothesis, i.e., that 5HTTLPR would explain variance in self-reported resilience. The analyses (shown in Table 1) are meant to show that this relationship is robust to the presence of covariates that could otherwise be confounders of this relationship. In other words, these multiple models can be considered “sensitivity analyses”. The analyses with BDI and NEO-N are meant to show that the relationship of 5HTTLPR is specific, not that 5HTTLPR explains variance in these other constructs.
Table 1.
Parameter Estimates for (Adjusted) Main Effects of 5HTTLPR from Robust Multivariate Linear Regression Analyses Predicting CDRISC-10 Scores
| Beta | 95% CI Beta | p value | |
|---|---|---|---|
| Model | |||
| Model 1a (N = 423) | −1.10 | −1.88, −0.318 | 0.006 |
| Model 2b (N = 423) | −1.07 | −1.85, −0.301 | 0.007 |
| Model 3c (N = 423) | −0.751 | −1.39, −0.114 | 0.021 |
| Model 4d (N = 318) | −0.978 | −1.83, −0.126 | 0.025 |
| Model 5e (N = 318) | −1.06 | −1.91, −0.203 | 0.015 |
| Model 6f (N = 423) | −0.955 | −1.74, −0.171 | 0.017 |
Model 1: Covariates: None
Model 2: Covariates: Sex
Model 3: Covariates: Sex and Neuroticism Score
Model 4: Covariates: Sex and Ethnicity in European-American + Hispanic Subsample
Model 5: Covariates: Sex and Ancestral Proportion Scores (2) in European-American + Hispanic Subsample
Model 6: Covariates: Sex and Ancestral Proportion Scores (2)
RESULTS
CDRISC-10 Scores in the Sample
Mean CDRISC-10 score in the sample (N = 423) was 26.17 SD 5.88 [range 5-40]. The range of CDRISC-10 scores is shown in Figure 1, which shows them to be approximately normally distributed. Women (N = 318) had significantly lower CDRISC-10 scores (25.62 SD 5.87) than men (N = 105; 27.84 SD 5.62) (t = −3.39, df = 421, p = 0.0008). Comparison of the two largest self-identified ethnic groups in the sample also revealed that European-Americans (N = 260; 26.72 SD 5.51) had significantly higher CDRISC-10 scores than Hispanics (N = 68; 24.76 SD 6.24) (t = 2.54, df = 326, p = 0.012).
FIGURE 1. Distribution of CDRISC-10 Scores in the Sample (N=423).
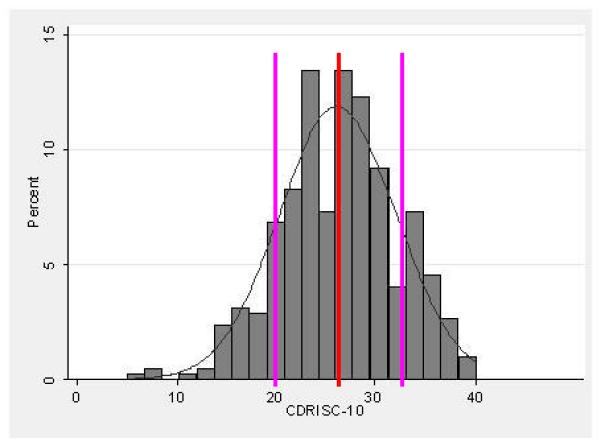
Distribution of CDRISC-10 scores in the sample of N = 423 subject. The red bar shows the mean value for the sample with the pink bars showing 1 standard deviation (SD) below and above the mean, respectively.
Relationship of 5HTTLPR to CDRISC-10 Scores
Subjects with 1 or 2 copies of the S allele (N = 307; 25.81 SD 5.90) had significantly lower CDRISC-10 scores than those with zero copies of the S allele (N = 116; 27.14 SD 5.75) (t = 2.08, df = 421, p = 0. 038).
There were no significant interactions found between 5HTTLPR genotype and childhood emotional abuse (CTQ-EA) scores in predicting CDRISC-10 scores (data not shown). Consequently, subsequent analyses focused on main effects of 5HTTLPR.
A series of multivariate linear regression models was run using 5HTTLPR genotype to predict CDRISC-10 scores, incorporating additional covariates as follows: Model 1: no covariates; Model 2: sex; Model 3: sex and neuroticism scores; Model 4: sex and self-reported ethnicity (in the two largest ethnic groups, Caucasians and Hispanics); Model 5: sex and ancestral proportion scores in Caucasians and Hispanics; and Model 6: sex and ancestral proportion scores in the entire sample. The regression coefficients (and their standard errors) for 5HTTLPR – adjusted for all other covariates in the model – are shown in Table 1. In all models, which explained approximately 5% of the variance in resilience scores, increased number of S copies was linearly associated with significantly reduced resilience scores.
The triallelic 5HTTLPR classification system failed to show any association with CDRISC-10 scores (data not shown, but available upon request from the corresponding author) in any of the aforementioned models.
Relationship of 5HTTLPR Genotype to Resilience Risk Categorization
To express the impact of 5HTTLPR genotype in a different way, we examined the relationship between 5HTTLPR genotype (coded 0, 1, 2 per copy of the S allele) and odds of being in the lowest category of resilience (i.e., 1 SD or more below the sex-specific group mean; N = 70 [16.55% of subjects]).
Each copy of the S allele was associated with a 63% increase in the likelihood of being in the low-resilient category: Odds Ratio (OR) = 1.63, 95% CI 1.14-2.34, p = 0.008. When adjusted for ancestral proportion scores, the finding remained essentially unchanged: OR = 1.53, 95% CI 1.06-2.21, p = 0.024.
Specificity of Findings: Relationship of 5HTTLPR to Depression and Neuroticism
To determine the specificity of the findings for emotional resilience, a series of multivariate linear regression models was run using 5HTTLPR genotype to predict depression (BDI) and neuroticism (NEO-N) scores, respectively. These models paralleled those described above for the prediction of resilience. None of the models found statistically significant effects of 5HTTLPR to predict either BDI or NEO-N scores, respectively (data not shown, but available upon request from the corresponding author).
DISCUSSION
We found that variation in 5HTTLPR genotype was significantly (although modestly, in terms of the proportion of variance explained) associated in a dose-response fashion with individual differences in self-reported emotional resilience. Specifically, a greater number of “S” copies was associated with lower resilience. This is the first study, to the best of our knowledge, to find a relationship between a gene and emotional resilience. Importantly, whereas many prior studies have detected an interaction between 5HTTLPR and stressful life experiences (e.g., childhood maltreatment) (Caspi et al. 2003) that predicted psychopathology (e.g., depressive symptoms or disorder), the relationship observed here between 5HTTLPR and resilience was independent of childhood maltreatment (the only life stressor assessed in this study). Moreover, in the present study, the relationship between 5HTTLPR and resilience was specific to the extent that no such relationship was observed with another dispositional trait (i.e., neuroticism) thought to predispose to psychopathology, or with the severity of a particular set of state-dependent behavioral (i.e., depressive) symptoms.
Since the construct of emotional resilience is defined as an individual's habitual capacity to withstand stress, it stands to reason that a main effect of 5HTTLPR would be observed here, whereas an interaction with stress would be required to see a relationship with neurobehavioral outcomes (e.g., depressive illness). Many investigators have argued that 5HTTLPR operates pleiotropically to influence emotional responsivity to stress (e.g., (Caspi and Moffitt 2006;Kendler et al. 2005;Stein et al. 2008)). The present findings support that argument to the extent that they specifically implicate the construct of emotional resilience as a likely mediator of that relationship. It must be noted, however, that our failure to find a significant association between 5HTTLPR and neuroticism may reflect insufficient power to detect such an association and, as such, cannot be considered definitive evidence of this type of specificity.
This study has a number of limitations. First and foremost, the observed effect size is small, and replication will be mandatory. Second, although the sample is drawn from a college student sample, it is not population based and, moreover, has a predominance of women, raising questions about generalizability; once again, replication in larger, more diverse samples is needed. Third, the two ancestral components used as covariates in the analyses are unlikely to represent all of the ethnic diversity inherent in the sample. Fourth, the assessment of emotional resilience is based exclusively on self-report. As such, it will be important to extend and replicate the findings using additional behavioral and biological (e.g., functional neuroimaging) measures of emotional resilience.
Although this is the first study to specifically link 5HTTLPR to emotional resilience, there are several studies in the recent literature that could have presaged such a finding. In a study of 350 college students asked to record their daily stressors, researchers found a significant association between 5HTTLPR genotype (using the triallelic classification) and anxious mood on stressful days, wherein La/La subjects were less emotionally reactive to stressors than Lg or S carriers (Gunthert et al. 2007). In a study of 557 outpatients with chronic coronary disease, S carriers were not only more likely to be depressed, but also had higher mean scores for perceived stress than L/L homozygotes (Otte et al. 2007). These findings are compatible with the notion that individual differences in emotional resilience – conceptualized as the ability to buffer or rebound from stressful life experiences – are, in part, influenced by variation in 5HTTLPR. As is the case in much of the recent literature, findings are inconsistent as to whether genetic vulnerability is more closely associated with the biallelic or triallelic classification of 5HTTLPR, making functional attributions somewhat speculative at this juncture. In this regard, we do not have a good explanation for why the triallelic (a/g SNP) classification failed to show an association with resilience in out data.
Reference to the functional neuroimaging literature may be informative for proposing neural mechanisms by which 5HTTLPR may contribute to increased resilience. For example, emotional regulation is one psychological mechanism thought to contribute to emotional resilience (Gross and John 2003), and it is thought to involve the capacity to regulate amygdala activity (probably through top-down effects from lateral and medial prefrontal cortex) during emotional processing (Hare et al. 2005;Phan et al. 2003;Phan et al. 2005). An important role has been posited for 5HTTLPR in emotion regulation and other types of social cognition (Canli and Lesch 2007). A remarkably consistent finding across imaging studies has been the observation that 5HTTLPR “S” carriers have increased amygdala activation during emotion processing tasks (e.g., viewing emotional faces) (Hariri et al. 2005;Bertolino et al. 2005), including ambiguous presentations that may be particularly stressful (Heinz et al. 2007). Given our findings, we hypothesize that individual differences in resilience mediate the relationship between 5HTTLPR genotype and amygdala response to emotion processing, and propose that this hypothesis be tested in future functional neuroimaging studies.
5HTTLPR is not the only plausible candidate variant for emotional resilience. Indeed, our observation that 5HTTLPR explains only a small proportion of the variance in resilience suggests that other influences must exist. Further research should determine whether other polymorphisms conferring relative alterations in brain serotonin availability similarly bias emotional resiliency, or govern different aspects of resiliency, either independently or in concert with 5HTTLPR. Good candidates to consider would be the MAO-A VNTR low activity variant (Schulze et al. 2000), the norepinephrine transporter (NET) which in gene knockout studies in mice (Dziedzicka-Wasylewska et al. 2006) and human clinical studies (Davidson et al. 2005) has shown a relationship to stress resilience, and the HTR1A variant (−1019C) which itself has been shown to be associated with a latent genetic factor underlying susceptibility to anxiety and depression (Hettema et al. 2008b).
To the extent that amygdala reactivity during emotion processing can be considered a correlate of resilience, the functional neuroimaging literature also points to other candidates such as catechol-O-methyltransferase (COMT) (Smolka et al. 2005) (which would also be considered a candidate because of its association with increased susceptibility to anxiety-spectrum phenotypes [(Hettema et al. 2008a)]), and neuropeptide Y (NPY) (Zhou et al. 2008). But other stress-related genes such as FKBP5 – which has recently been shown to interact with childhood abuse to increase risk for posttraumatic stress disorder (Binder et al. 2008) should also be considered in terms of their contribution to individual differences in emotional resilience.
In conclusion, we have demonstrated a “dose-dependent” relationship between a well-studied variant at the serotonin transporter protein gene (SLC6A4) and emotional resilience, an observation that mirrors previous work demonstrating that the same variant predisposes to negative psychiatric outcomes in the presence of stressors. This is a useful step in allowing a deeper conceptualization of the relationship between this variant (and the serotonin system) and mental health and illness (Holmes 2008), a conceptualization that lends itself to several directions of further experimental evaluation, as described above.
Acknowledgments
Supported in part by NIMH grant MH64122 to MBS and NIAAA grants AA11330, and NIDA grants DA12690, DA12849, and DA15105 to JG; and the U.S. Department of Veterans Affairs (the VA Medical Research Program [Merit Review] to JG). The authors are grateful to Carla Hitchcock and Alexis Rochlin for coordinating the study and to Ann Marie Lacobelle and Greg Kay for excellent technical assistance. Thanks also to Kathryn Connor MD and Jonathan Davidson MD for permitting the use of their measure of resilience in this study.
Footnotes
Disclosure/Conflict of Interest
There are no conflicts for any of the authors relating to this submission.
Dr. Stein reports that he has received compensation for professional services in the previous 3 years from the following entities: University of California San Diego; Veterans Affairs Healthcare System (VA); National Institutes of Health (NIMH); Forest Pharmaceuticals; Avera Pharmaceuticals; AstraZeneca; BrainCells, Inc.; Bristol-Myers Squibb; Integral Health Decisions, Inc.; JDS Pharmaceuticals; Johnson & Johnson; Lilly; Jazz Pharmaceuticals; and Comprehensive NeuroScience, Inc.
Dr. Campbell-Sills reports that she has received compensation for professional services in the previous 3 years from the following entities: University of California San Diego; the National Institutes of Health (NIMH); and the American Psychiatric Association.
Dr. Gelernter reports that he has received compensation for professional services in the previous 3 years from the following entities: Yale University School of Medicine; Veterans Affairs Healthcare System (VA) and the National Institutes of Health (NIAAA, NIDA, and NIMH).
References
- Beck AT, Ward CH, Mendelson M, Mock J, Erbaugh J. An inventory for measuring depression. Arch Gen Psychiatry. 1961;4:561–571. doi: 10.1001/archpsyc.1961.01710120031004. [DOI] [PubMed] [Google Scholar]
- Bernstein DP, Fink L, Handelsman L, Lovejoy M, Wenzel K, Sapareto E, Ruggiero J. Initial reliability and validity of a new retrospective measure of child abuse and neglect. Am J Psychiatry. 1994;151:1132–1136. doi: 10.1176/ajp.151.8.1132. [DOI] [PubMed] [Google Scholar]
- Bertolino A, Arciero G, Rubino V, Latorre V, De Candia M, Mazzola V, Blasi G, Caforio G, Hariri A, Kolachana B, Nardini M, Weinberger DR, Scarabino T. Variation of human amygdala response during threatening stimuli as a function of 5′HTTLPR genotype and personality style. Biol Psychiatry. 2005;57:1517–1525. doi: 10.1016/j.biopsych.2005.02.031. [DOI] [PubMed] [Google Scholar]
- Binder EB, Bradley RG, Liu W, Epstein MP, Deveau TC, Mercer KB, Tang Y, Gillespie CF, Heim CM, Nemeroff CB, Schwartz AC, Cubells JF, Ressler KJ. Association of FKBP5 polymorphisms and childhood abuse with risk of posttraumatic stress disorder symptoms in adults. JAMA. 2008;299:1291–1305. doi: 10.1001/jama.299.11.1291. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bonanno GA. Loss, trauma, and human resilience - Have we underestimated the human capacity to thrive after extremely aversive events? American Psychologist. 2004;59:20–28. doi: 10.1037/0003-066X.59.1.20. [DOI] [PubMed] [Google Scholar]
- Campbell-Sills L, Cohan SL, Stein MB. Relationship of resilience to personality, coping, and psychiatric symptoms in young adults. Behav Res Ther. 2006;44:585–599. doi: 10.1016/j.brat.2005.05.001. [DOI] [PubMed] [Google Scholar]
- Campbell-Sills L, Stein MB. Psychometric analysis and refinement of the connor-davidson resilience scale (CD-RISC): Validation of a 10-item measure of resilience. J Trauma Stress. 2007;20:1019–1028. doi: 10.1002/jts.20271. [DOI] [PubMed] [Google Scholar]
- Canli T, Lesch KP. Long story short: the serotonin transporter in emotion regulation and social cognition. Nat Neurosci. 2007;10:1103–1109. doi: 10.1038/nn1964. [DOI] [PubMed] [Google Scholar]
- Caspi A, Moffitt TE. Gene-environment interactions in psychiatry: joining forces with neuroscience. Nat Rev Neurosci. 2006;7:583–590. doi: 10.1038/nrn1925. [DOI] [PubMed] [Google Scholar]
- Caspi A, Sugden K, Moffitt TE, Taylor A, Craig IW, Harrington H, McClay J, Mill J, Martin J, Braithwaite A, Poulton R. Influence of life stress on depression: moderation by a polymorphism in the 5-HTT gene. Science. 2003;301:386–389. doi: 10.1126/science.1083968. [DOI] [PubMed] [Google Scholar]
- Charney DS. Psychobiological mechanisms of resilience and vulnerability: implications for successful adaptation to extreme stress. Am J Psychiatry. 2004;161:195–216. doi: 10.1176/appi.ajp.161.2.195. [DOI] [PubMed] [Google Scholar]
- Chorbov VM, Lobos EA, Todorov AA, Heath AC, Botteron KN, Todd RD. Relationship of 5-HTTLPR genotypes and depression risk in the presence of trauma in a female twin sample. Am J Med Genet B Neuropsychiatr Genet. 2007;144:830–833. doi: 10.1002/ajmg.b.30534. [DOI] [PubMed] [Google Scholar]
- Connor KM, Davidson JR. Development of a new resilience scale: The Connor-Davidson Resilience Scale (CD-RISC) Depress Anxiety. 2003;18:76–82. doi: 10.1002/da.10113. [DOI] [PubMed] [Google Scholar]
- Costa PT, Jr., McCrae RR. NEO PI-R Professional Manual. Psychological Assessment Resources, Inc.; Odessa, FL: 1992. [Google Scholar]
- Davidson J, Watkins L, Owens M, Krulewicz S, Connor K, Carpenter D, Krishnan R, Nemeroff C. Effects of paroxetine and venlafaxine XR on heart rate variability in depression. J Clin Psychopharmacol. 2005;25:480–484. doi: 10.1097/01.jcp.0000177547.28961.03. [DOI] [PubMed] [Google Scholar]
- Dziedzicka-Wasylewska M, Faron-Gorecka A, Kusmider M, Drozdowska E, Rogoz Z, Siwanowicz J, Caron MG, Bonisch H. Effect of antidepressant drugs in mice lacking the norepinephrine transporter. Neuropsychopharmacology. 2006;31:2424–2432. doi: 10.1038/sj.npp.1301064. [DOI] [PubMed] [Google Scholar]
- Eley TC, Sugden K, Corsico A, Gregory AM, Sham P, McGuffin P, Plomin R, Craig IW. Gene-environment interaction analysis of serotonin system markers with adolescent depression. Mol Psychiatry. 2004;9:908–915. doi: 10.1038/sj.mp.4001546. [DOI] [PubMed] [Google Scholar]
- Falush D, Stephens M, Pritchard JK. Inference of population structure using multilocus genotype data: linked loci and correlated allele frequencies. Genetics. 2003;164:1567–1587. doi: 10.1093/genetics/164.4.1567. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Galea S, Brewin CR, Gruber M, Jones RT, King DW, King LA, McNally RJ, Ursano RJ, Petukhova M, Kessler RC. Exposure to hurricane-related stressors and mental illness after Hurricane Katrina. Arch Gen Psychiatry. 2007;64:1427–1434. doi: 10.1001/archpsyc.64.12.1427. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Galea S, Vlahov D, Resnick H, Ahern J, Susser E, Gold J, Bucuvalas M, Kilpatrick D. Trends of probable post-traumatic stress disorder in New York City after the September 11 terrorist attacks. Am J Epidemiol. 2003;158:514–524. doi: 10.1093/aje/kwg187. [DOI] [PubMed] [Google Scholar]
- Gelernter J, Kranzler H, Cubells JF. Serotonin transporter protein (SLC6A4) allele and haplotype frequencies and linkage disequilibria in African- and European-American and Japanese populations and in alcohol-dependent subjects. Hum Genet. 1997;101:243–246. doi: 10.1007/s004390050624. [DOI] [PubMed] [Google Scholar]
- Gillespie NA, Whitfield JB, Williams B, Heath AC, Martin NG. The relationship between stressful life events, the serotonin transporter (5-HTTLPR) genotype and major depression. Psychol Med. 2005;35:101–111. doi: 10.1017/s0033291704002727. [DOI] [PubMed] [Google Scholar]
- Grabe HJ, Lange M, Wolff B, Volzke H, Lucht M, Freyberger HJ, John U, Cascorbi I. Mental and physical distress is modulated by a polymorphism in the 5-HT transporter gene interacting with social stressors and chronic disease burden. Mol Psychiatry. 2005;10:220–224. doi: 10.1038/sj.mp.4001555. [DOI] [PubMed] [Google Scholar]
- Gross JJ, John OP. Individual differences in two emotion regulation processes: implications for affect, relationships, and well-being. J Pers Soc Psychol. 2003;85:348–362. doi: 10.1037/0022-3514.85.2.348. [DOI] [PubMed] [Google Scholar]
- Gunthert KC, Conner TS, Armeli S, Tennen H, Covault J, Kranzler HR. Serotonin transporter gene polymorphism (5-HTTLPR) and anxiety reactivity in daily life: a daily process approach to gene-environment interaction. Psychosom Med. 2007;69:762–768. doi: 10.1097/PSY.0b013e318157ad42. [DOI] [PubMed] [Google Scholar]
- Hare TA, Tottenham N, Davidson MC, Glover GH, Casey BJ. Contributions of amygdala and striatal activity in emotion regulation. Biol Psychiatry. 2005;57:624–632. doi: 10.1016/j.biopsych.2004.12.038. [DOI] [PubMed] [Google Scholar]
- Hariri AR, Drabant EM, Munoz KE, Kolachana BS, Mattay VS, Egan MF, Weinberger DR. A susceptibility gene for affective disorders and the response of the human amygdala. Arch Gen Psychiatry. 2005;62:146–152. doi: 10.1001/archpsyc.62.2.146. [DOI] [PubMed] [Google Scholar]
- Hariri AR, Holmes A. Genetics of emotional regulation: the role of the serotonin transporter in neural function. Trends Cogn Sci. 2006;10:182–191. doi: 10.1016/j.tics.2006.02.011. [DOI] [PubMed] [Google Scholar]
- Heinz A, Smolka MN, Braus DF, Wrase J, Beck A, Flor H, Mann K, Schumann G, Buchel C, Hariri AR, Weinberger DR. Serotonin transporter genotype (5-HTTLPR): effects of neutral and undefined conditions on amygdala activation. Biol Psychiatry. 2007;61:1011–1014. doi: 10.1016/j.biopsych.2006.08.019. [DOI] [PubMed] [Google Scholar]
- Hettema JM, An SS, Bukszar J, van den Oord EJ, Neale MC, Kendler KS, Chen X. Catechol-O-Methyltransferase Contributes to Genetic Susceptibility Shared Among Anxiety Spectrum Phenotypes. Biol Psychiatry. 2008a doi: 10.1016/j.biopsych.2008.03.014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hettema JM, An SS, van den Oord EJ, Neale MC, Kendler KS, Chen X. Association study between the serotonin 1A receptor (HTR1A) gene and neuroticism, major depression, and anxiety disorders. Am J Med Genet B Neuropsychiatr Genet. 2008b;147B:661–666. doi: 10.1002/ajmg.b.30656. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Holmes A. Genetic variation in cortico-amygdala serotonin function and risk for stress-related disease. Neurosci Biobehav Rev. 2008 doi: 10.1016/j.neubiorev.2008.03.006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hu X, Oroszi G, Chun J, Smith TL, Goldman D, Schuckit MA. An expanded evaluation of the relationship of four alleles to the level of response to alcohol and the alcoholism risk. Alcohol Clin Exp Res. 2005;29:8–16. doi: 10.1097/01.alc.0000150008.68473.62. [DOI] [PubMed] [Google Scholar]
- Jacobs N, Kenis G, Peeters F, Derom C, Vlietinck R, van Os J. Stress-related negative affectivity and genetically altered serotonin transporter function: evidence of synergism in shaping risk of depression. Arch Gen Psychiatry. 2006;63:989–996. doi: 10.1001/archpsyc.63.9.989. [DOI] [PubMed] [Google Scholar]
- Kaufman J, Yang BZ, Douglas-Palumberi H, Crouse-Artus M, Lipschitz D, Krystal JH, Gelernter J. Genetic and environmental predictors of early alcohol use. Biol Psychiatry. 2007;61:1228–1234. doi: 10.1016/j.biopsych.2006.06.039. [DOI] [PubMed] [Google Scholar]
- Kaufman J, Yang BZ, Douglas-Palumberi H, Grasso D, Lipschitz D, Houshyar S, Krystal JH, Gelernter J. Brain-derived neurotrophic factor-5-HTTLPR gene interactions and environmental modifiers of depression in children. Biol Psychiatry. 2006;59:673–680. doi: 10.1016/j.biopsych.2005.10.026. [DOI] [PubMed] [Google Scholar]
- Kaufman J, Yang BZ, Douglas-Palumberi H, Houshyar S, Lipschitz D, Krystal JH, Gelernter J. Social supports and serotonin transporter gene moderate depression in maltreated children. Proc Natl Acad Sci U S A. 2004;101:17316–17321. doi: 10.1073/pnas.0404376101. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kendler KS. “A gene for”: The nature of gene action in psychiatric disorders. Am J Psychiatry. 2005;162:1243–1252. doi: 10.1176/appi.ajp.162.7.1243. [DOI] [PubMed] [Google Scholar]
- Kendler KS, Kuhn JW, Vittum J, Prescott CA, Riley B. The interaction of stressful life events and a serotonin transporter polymorphism in the prediction of episodes of major depression: a replication. Arch Gen Psychiatry. 2005;62:529–535. doi: 10.1001/archpsyc.62.5.529. [DOI] [PubMed] [Google Scholar]
- Kessler RC, Sonnega A, Bromet E, Hughes M, Nelson CB. Posttraumatic stress disorder in the National Comorbidity Survey. Arch Gen Psychiatry. 1995;52:1048–1060. doi: 10.1001/archpsyc.1995.03950240066012. [DOI] [PubMed] [Google Scholar]
- King DW, King LA, Foy DW, Keane TM, Fairbank JA. Posttraumatic stress disorder in a national sample of female and male Vietnam veterans: Risk factors, war-zone stressors, and resilience-recovery variables. J Abnorm Psychol. 1999;108:164–170. doi: 10.1037//0021-843x.108.1.164. [DOI] [PubMed] [Google Scholar]
- Lesch K-P, Bengel D, Heils A, Sabol SA, Greenberg BD, Petri S, Benjamin J, Müller CR, Hamer DH, Murphy DL. Association of anxiety-related traits with a polymorphism in the serotonin transporter gene regulatory region. Science. 1996;274:1527–1531. doi: 10.1126/science.274.5292.1527. [DOI] [PubMed] [Google Scholar]
- Luthar SS, Cicchetti D, Becker B. The construct of resilience: A critical evaluation and guidelines for future work. Child Development. 2000;71:543–562. doi: 10.1111/1467-8624.00164. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Middeldorp CM, Cath DC, Van Dyck R, Boomsma DI. The co-morbidity of anxiety and depression in the perspective of genetic epidemiology. A review of twin and family studies. Psychol Med. 2005;35:611–624. doi: 10.1017/s003329170400412x. [DOI] [PubMed] [Google Scholar]
- Otte C, McCaffery J, Ali S, Whooley MA. Association of a Serotonin Transporter Polymorphism (5-HTTLPR) With Depression, Perceived Stress, and Norepinephrine in Patients With Coronary Disease: The Heart and Soul Study. Am J Psychiatry. 2007;164:1379–1384. doi: 10.1176/appi.ajp.2007.06101617. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Parsey RV, Hastings RS, Oquendo MA, Hu X, Goldman D, Huang YY, Simpson N, Arcement J, Huang Y, Ogden RT, Van Heertum RL, Arango V, Mann JJ. Effect of a triallelic functional polymorphism of the serotonin-transporter-linked promoter region on expression of serotonin transporter in the human brain. Am J Psychiatry. 2006;163:48–51. doi: 10.1176/appi.ajp.163.1.48. [DOI] [PubMed] [Google Scholar]
- Phan KL, Fitzgerald DA, Nathan PJ, Moore GJ, Uhde TW, Tancer ME. Neural substrates for voluntary suppression of negative affect: a functional magnetic resonance imaging study. Biol Psychiatry. 2005;57:210–219. doi: 10.1016/j.biopsych.2004.10.030. [DOI] [PubMed] [Google Scholar]
- Phan KL, Taylor SF, Welsh RC, Decker LR, Noll DC, Nichols TE, Britton JC, Liberzon I. Activation of the medial prefrontal cortex and extended amygdala by individual ratings of emotional arousal: a fMRI study. Biol Psychiatry. 2003;53:211–215. doi: 10.1016/s0006-3223(02)01485-3. [DOI] [PubMed] [Google Scholar]
- Pritchard JK, Rosenberg NA. Use of unlinked genetic markers to detect population stratification in association studies. Am J Hum Genet. 1999;65:220–228. doi: 10.1086/302449. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pritchard JK, Stephens M, Rosenberg NA, Donnely P. Association mapping in structured populations. Am J Hum Genet. 2000;67:170–181. doi: 10.1086/302959. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Scher CD, Stein MB, Asmundson GJ, McCreary DR, Forde DR. The childhood trauma questionnaire in a community sample: psychometric properties and normative data. J Traum Stress. 2001;14:843–857. doi: 10.1023/A:1013058625719. [DOI] [PubMed] [Google Scholar]
- Schinka JA, Busch RM, Robichaux-Keene N. A meta-analysis of the association between the serotonin transporter gene polymorphism (5-HTTLPR) and trait anxiety. Mol Psychiatry. 2004;9:197–202. doi: 10.1038/sj.mp.4001405. [DOI] [PubMed] [Google Scholar]
- Schulze TG, Muller DJ, Krauss H, Scherk H, Ohlraun S, Syagailo YV, Windemuth C, Neidt H, Grassle M, Papassotiropoulos A, Heun R, Nothen MM, Maier W, Lesch KP, Rietschel M. Association between a functional polymorphism in the monoamine oxidase A gene promoter and major depressive disorder. Am J Med Genet. 2000;96:801–803. [PubMed] [Google Scholar]
- Sen S, Burmeister M, Ghosh D. Meta-analysis of the association between a serotonin transporter promoter polymorphism (5-HTTLPR) and anxiety-related personality traits. Am J Med Genet. 2004;127B:85–89. doi: 10.1002/ajmg.b.20158. [DOI] [PubMed] [Google Scholar]
- Smolka MN, Schumann G, Wrase J, Grusser SM, Flor H, Mann K, Braus DF, Goldman D, Buchel C, Heinz A. Catechol-O-methyltransferase val158met genotype affects processing of emotional stimuli in the amygdala and prefrontal cortex. J Neurosci. 2005;25:836–842. doi: 10.1523/JNEUROSCI.1792-04.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- StataCorp. Stata Statistical Software. [Release 10.0] Stata Corporation; College Station, TX: 2007. [Google Scholar]
- Stein MB, Schork NJ, Gelernter J. Gene-by-Environment (Serotonin Transporter and Childhood Maltreatment) Interaction for Anxiety Sensitivity, an Intermediate Phenotype for Anxiety Disorders. Neuropsychopharmacology. 2008;33:312–319. doi: 10.1038/sj.npp.1301422. [DOI] [PubMed] [Google Scholar]
- Surtees PG, Wainwright NW, Willis-Owen SA, Luben R, Day NE, Flint J. Social adversity, the serotonin transporter (5-HTTLPR) polymorphism and major depressive disorder. Biol Psychiatry. 2006;59:224–229. doi: 10.1016/j.biopsych.2005.07.014. [DOI] [PubMed] [Google Scholar]
- Tsuang M. Genes, environment, and mental health wellness. Am J Psychiatry. 2000;157:489–491. doi: 10.1176/appi.ajp.157.4.489. [DOI] [PubMed] [Google Scholar]
- Tugade MM, Fredrickson BL. Resilient individuals use positive emotions to bounce back from negative emotional experiences. J Pers Soc Psychol. 2004;86:320–333. doi: 10.1037/0022-3514.86.2.320. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wellman CL, Izquierdo A, Garrett JE, Martin KP, Carroll J, Millstein R, Lesch KP, Murphy DL, Holmes A. Impaired stress-coping and fear extinction and abnormal corticolimbic morphology in serotonin transporter knock-out mice. J Neurosci. 2007;27:684–691. doi: 10.1523/JNEUROSCI.4595-06.2007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wendland JR, Martin BJ, Kruse MR, Lesch KP, Murphy DL. Simultaneous genotyping of four functional loci of human SLC6A4, with a reappraisal of 5-HTTLPR and rs25531. Mol Psychiatry. 2006;11:224–226. doi: 10.1038/sj.mp.4001789. [DOI] [PubMed] [Google Scholar]
- Wilhelm K, Mitchell PB, Niven H, Finch A, Wedgwood L, Scimone A, Blair IP, Parker G, Schofield PR. Life events, first depression onset and the serotonin transporter gene. Br J Psychiatry. 2006;188:210–215. doi: 10.1192/bjp.bp.105.009522. [DOI] [PubMed] [Google Scholar]
- Yang BZ, Zhao H, Kranzler HR, Gelernter J. Practical population group assignment with selected informative markers: Characteristics and properties of Bayesian clustering via STRUCTURE. Genet Epidemiol. 2005;28:302–312. doi: 10.1002/gepi.20070. [DOI] [PubMed] [Google Scholar]
- Zalsman G, Huang YY, Oquendo MA, Burke AK, Hu XZ, Brent DA, Ellis SP, Goldman D, Mann JJ. Association of a triallelic serotonin transporter gene promoter region (5-HTTLPR) polymorphism with stressful life events and severity of depression. Am J Psychiatry. 2006;163:1588–1593. doi: 10.1176/ajp.2006.163.9.1588. [DOI] [PubMed] [Google Scholar]
- Zhou Z, Zhu G, Hariri AR, Enoch MA, Scott D, Sinha R, Virkkunen M, Mash DC, Lipsky RH, Hu XZ, Hodgkinson CA, Xu K, Buzas B, Yuan Q, Shen PH, Ferrell RE, Manuck SB, Brown SM, Hauger RL, Stohler CS, Zubieta JK, Goldman D. Genetic variation in human NPY expression affects stress response and emotion. Nature. 2008;452:997–1001. doi: 10.1038/nature06858. [DOI] [PMC free article] [PubMed] [Google Scholar]